
Abstract
Abuse of cocaine and androgenic–anabolic steroids (AASs) has become a serious public health problem. Despite reports of an increase in the incidence of simultaneous abuse of these substances, potential toxic interactions between cocaine and AASs are poorly known. In the present study, we investigated the effects of either single or combined administration of testosterone and cocaine for one or 10 consecutive days on autonomic (arterial pressure, heart rate and tail cutaneous temperature) and neuroendocrine (plasma corticosterone) responses induced by acute restraint stress in rats. Combined administration of testosterone and cocaine for 10 days reduced the increase in heart rate and plasma corticosterone level, as well as the fall in tail skin temperature induced by restraint stress. Furthermore, repeated administration of cocaine inhibited the increase in arterial pressure observed during restraint, and this effect was not affected by coadministration of testosterone. Ten-day combined administration of testosterone and cocaine increased basal values of arterial pressure. Moreover, chronic administration of testosterone induced rest bradycardia and elevated basal level of plasma corticosterone. One-day single or combined administration of the drugs did not affect any parameter investigated.
In conclusion, the present study demonstrated that combined administration of testosterone and cocaine changed the autonomic and neuroendocrine responses to acute restraint stress. These findings suggest that interaction between AASs and cocaine may affect the ability to cope with stressful events.
Keywords
Introduction
Abuse of cocaine and androgenic–anabolic steroids (AASs) is a serious public health problem (Goldstein et al., 2009; Kanayama et al., 2009a; UNODC, 2010). Emerging results indicate that AAS use is associated with abuse of other psychotropic drugs (Arvary and Pope, 2000). Previous clinical reports have pointed up that cocaine is the drug most frequently coabused by AAS users (DuRant et al., 1993, 1995). Indeed, it has been reported that AAS users are likely to display much higher cocaine intake than non-users (McCabe et al., 2007; Kanayama et al., 2009b). Although many reports have documented the toxic effects of single AAS or cocaine abuse, a limited number of studies have assessed the potential dysfunctions related to the concomitant use of these substances.
Maintenance of homeostasis during stress requires an appropriate and coordinated set of physiological responses (Dampney et al., 2008; Ulrich-Lai and Herman, 2009). Changes in autonomic activity is the initial response during aversive situations (Ulrich-Lai and Herman, 2009). Autonomic responses include arterial pressure and heart rate increase (Vianna and Carrive, 2005; Dampney et al., 2008; Busnardo et al., 2010). Moreover, during the exposure to aversive stimuli hemodynamic changes that involve vasoconstriction in the splanchnic, renal and cutaneous circulations, and vasodilatation in the skeletal muscles are observed (Zhang et al., 1996). Cutaneous vasoconstriction during stress evokes a rapid skin temperature drop (Vianna and Carrive, 2005; Busnardo et al., 2010). Besides autonomic responses, stress evokes activation of the hypothalamic–pituitary–adrenocortical (HPA) axis, which results in elevations of circulating glucocorticoids (Ulrich-Lai and Herman, 2009; Busnardo et al., 2010).
A role for stress in drug addiction has been established (Sinha, 2008; Briand and Blendy, 2010). However, the effect of drug abuse on physiological adjustments to stress is poorly understood. Both cocaine and AAS abuse are related to cardiovascular pathologies and HPA axis dysfunctions (Kloner et al., 1992; Buydens-Branchey et al., 2002; Williamson et al., 2005; van Amsterdam et al., 2010). Furthermore, high-frequency cocaine/alcohol abusers display enhanced cardiovascular and HPA responses to psychological stress (Fox et al., 2005). Nevertheless, preclinical studies have reported either no change or a decrease in stress-evoked glucocorticoid response following cocaine exposure (Levy et al., 1994; Sarnyai et al., 1998). Less is known about the effect of chronic cocaine exposure in cardiovascular adjustments to stress. It has been shown that exogenous testosterone decreases autonomic responses and HPA axis activation during stress (Williamson et al., 2005; Hermans et al., 2007). Although interaction between cocaine and testosterone on behavior and cardiovascular pathologies has been demonstrated (Long et al., 1994; Tseng et al., 1994; Engi et al., 2011; Menendez-Delmestre and Segarra, 2011), a possible influence of the combined administration of these substances in physiological responses to stress has never been investigated.
Therefore, our purpose in the present study was to investigate the effect of either single or combined daily administration of testosterone and cocaine for 10 consecutive days on cardiovascular, neuroendocrine and regional blood flow changes induced by acute restraint stress. Moreover, to test whether repeated administration of the drugs is necessary to affect physiological responses to restraint, the effect of one-day administration of testosterone and/or cocaine was studied in a different set of animals.
Material and methods
Animals
Forty-six 60-day-old (240–260g) male Wistar rats were used. Animals were obtained from the animal breeding facility of the São Paulo State University-UNESP (Botucatu-SP, Brazil) and were housed in plastic cages in a temperature-controlled room at 24°C in the Animal Facility of the Laboratory of Pharmacology, School of Pharmaceutical Sciences, São Paulo State University-UNESP. They were kept under a 12:12 h light–dark cycle (lights on between 06:00 and 18:00) with free access to water and standard laboratory food. Housing conditions and experimental procedures were carried out according to protocols approved by the Ethical Committee for Use of Animal and Subjects of the School of Pharmaceutical Science/UNESP.
Treatment
Different groups of animals were subjected to one-day administration or daily treatment for 10 consecutive days. In each treatment regimen, animals were randomly divided into four groups: (i) vehicle (almond oil, 1 mL/kg, subcutaneous (s.c.)) + saline (0.9% NaCl, 1 mL/kg, intraperitoneal (i.p.)) (veh+sal); (ii) testosterone (10 mg/kg, s.c.) + saline (T+sal); (iii) vehicle + cocaine (20 mg/kg, i.p.) (veh+coc); and (iv) testosterone + cocaine (T+coc) (Tseng et al., 1994; Engi et al., 2011). One- and 10-day treatments were performed once a day. Animals subjected to one-day treatment were left undisturbed, except for cleaning the cages, in the Animal Facility for the same period as the animals submitted to repeated treatment, and administrations were performed on the day corresponding to the 10th day of the repeated treatment.
Surgical preparation
Forty-eight hours after the end of treatments, and 48 hours before the trial, rats were anesthetized with tribromoethanol (250 mg/kg, i.p.) and a catheter (a 4 cm segment of PE-10 heat-bound to a 13 cm segment of PE-50) (Clay Adams, Parsippany, NJ, USA) was inserted into the abdominal aorta through the femoral artery, for cardiovascular recording. The catheter was tunneled under the skin and exteriorized on the animal’s dorsum. After the surgery, rats were treated with a streptomycin and penicillin polyantibiotic formulation (0.27 mg/kg, intramuscular (i.m.); Pentabiotico®; Fort Dodge, Campinas, SP, Brazil) to prevent infection, and received the non-steroidal anti-inflammatory drug flunixine meglumine (0.025 mg/kg, i.m.; Banamine®, Schering Plough, Cotia, SP, Brazil) for postoperative analgesia.
Measurement of cardiovascular parameters
On the day of the experiment, the arterial cannula was connected to a pressure transducer. Pulsatile arterial pressure was recorded using an amplifier (Quad Bridge Amp, ML224, ADInstruments, NSW, Australia) and an acquisition board (PowerLab 4/30, ML866/P, ADInstruments, NSW, Australia) connected to a personal computer. Mean arterial pressure (MAP) and heart rate (HR) values were derived from pulsatile arterial pressure recordings.
Tail skin temperature measurements
Variations in tail cutaneous temperature were recorded with a thermal camera Multi-Purpose Thermal Imager IRI4010 (InfraRed Integrated Systems Ltd, Northampton, UK) at a distance of 50 cm (Vianna and Carrive, 2005; Reis et al., 2011). For analysis of images, temperature measurement was realized in five points of the animal’s tail and the mean was calculated for each recording (Reis et al., 2011).
Plasma corticosterone measurements
Plasma corticosterone concentration was measured by radioimmunoassay. The radioimmunoassay for corticosterone was conducted using antibody obtained from Sigma (St Louis, Missouri, USA) and (3H)-corticosterone from New England Nuclear (Boston, Massachusetts, USA). The method was adapted from that described previously (Sarnyai et al., 1992). Briefly, 20 μL of plasma was diluted 50 times with 0.01M PBS and placed in a water bath at 75°C for 1h for heat inactivation of corticosteroid binding globulin. One hundred microliters of a solution of antibody and (3H)-corticosterone (10,000–20,000 cpm/mL) was added to each sample, mixed and incubated overnight at 4°C. Dextran-coated charcoal was used to adsorb free steroid after incubation. Tubes were centrifuged at 2000×g for 15 min at 4°C, the supernatant from each tube was transferred to scintillation vials and the radioactivity was quantified by liquid scintillation spectrometry. Standard curves were constructed using 25, 50, 100, 250, 500, 750, 1000 and 2000 pg/100 μL of corticosterone (Sigma). After dilution, all concentrations of corticosterone samples were within the linear range of the standard curve. Inter- and intra-assay variation was, respectively, 4.0 and 7.2%.
Acute restraint stress
Animals were submitted to restraint by placing each rat in a plastic cylindrical restraint tube (diameter 6.5 cm, length 15 cm), ventilated by holes (1 cm diameter) that made up approximately 20% of the tube surface. Restraint lasted 30 min, and immediately after the end of the stress exposure rats were returned to their home cages. Each rat was submitted to only one session of restraint in order to avoid habituation.
Drugs
Cocaine hydrochloride (Sigma, St Louis, Missouri, USA) and tribromoethanol (Sigma) were dissolved in saline (0.9% NaCl). Testosterone propionate (PharmaNostra, Rio de Janeiro, RJ, Brazil) was dissolved in almond oil. Flunixine meglumine (Banamine®, Schering Plough, Cotia, SP, Brazil) and polyantibiotic preparation (Pentabiotico®, Fort Dodge, Campinas, SP, Brazil) were used as provided.
Experimental procedures
Animals were transferred to the experimental room in their home cages. Animals were allowed at least 60 min to adapt to the experimental room conditions, such as sound and illumination, before the experiments were started. Experiments were always performed in the period from 9:00 am to 12:00 pm during lights on. The experimental room was temperature controlled (24oC) and acoustically isolated from the other rooms. Animals in all experimental groups were submitted to restraint stress.
Cardiovascular recordings began at least 30 min before restraint onset and were performed throughout the period of exposure to restraint stress. Other measurements began only when arterial pressure and heart rate were stable. Tail skin temperature was measured 10, 5 and 0 min before restraint for baseline values, and at every 5 min during restraint. Blood samples (200 μL) were collected from the femoral artery catheter immediately (basal level) and 15 min after onset of the restraint (stress). Blood samples were collected in heparinized tubes, centrifuged at 2000×g for 10 min at 4°C and plasma was stored at -20°C until radioimmunoassay.
Data analysis
Data were expressed as mean ± SEM. The basal values of arterial pressure, HR and tail temperature, as well as the changes in plasma corticosterone levels induced by acute restraint stress were compared using two-way ANOVA with treatment regimen (one- or 10-day treatment) and drug (testosterone and/or cocaine) as independent factors. The time-course during stress of change in MAP, HR, tail temperature and plasma corticosterone levels induced by restraint were analyzed using three-way ANOVA with treatment regimen (one- or 10-day treatment) and drug (testosterone and/or cocaine) as the main independent factors, and time as a repeated measurement. When interactions between the factors were observed, differences within treatment regimens were compared using one-way ANOVA followed by Bonferroni’s post hoc test.
Results
Effect of one- and 10-day single or combined administration of testosterone and cocaine on basal levels of arterial pressure, heart rate and tail skin temperature
Combined administration of testosterone and cocaine (T+coc group) for 10 days increased basal values of MAP (p<0.001), when compared with the veh+sal group within the same treatment regimen (Table 1). Analysis of baseline MAP indicated a significant effect of drug (F(3,38)=4.8, p<0.002), but not of treatment regimen (F(1,38)=2.1, p>0.05). In addition, an interaction between treatment regimen and drug was detected (F(3,38)=5.2, p<0.002). One-day single or combined administration of testosterone and cocaine did not affect baseline arterial pressure parameters (p>0.05) (Table 1).
Basal values of mean arterial pressure (MAP), heart rate (HR) and tail cutaneous temperature in animals subjected to single or combined administration of testosterone and cocaine for one (One day) or 10 consecutive days (10 days).
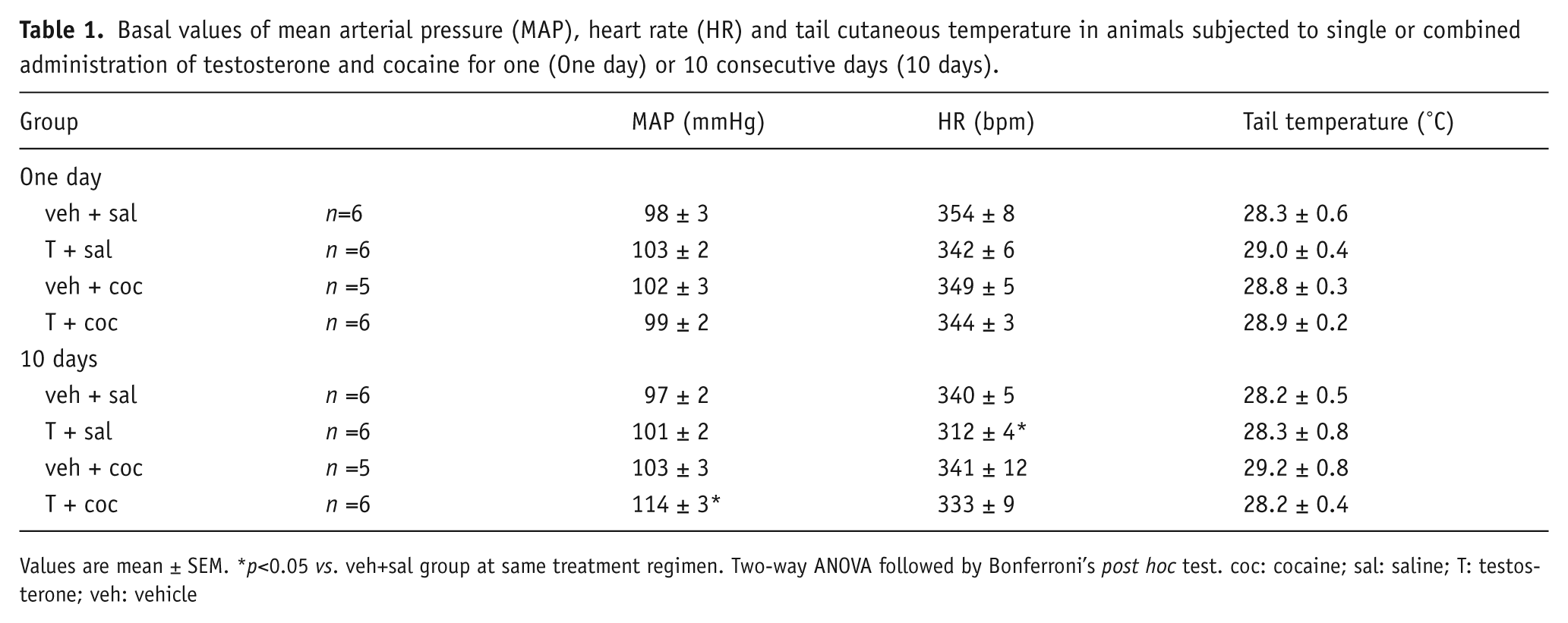
Values are mean ± SEM. *p<0.05 vs. veh+sal group at same treatment regimen. Two-way ANOVA followed by Bonferroni’s post hoc test. coc: cocaine; sal: saline; T: testosterone; veh: vehicle
Administration of T+sal for 10 days decreased basal values of HR (p<0.05), when compared with the veh+sal group within the same treatment regimen (Table 1). There were significant effects of treatment regimen (F(1,38)=8.2, p<0.006) and drug (F(3,38)=2.9, p<0.03), but no interaction between factors (F(3,38)=0.9, p>0.05). One-day single or combined administration of testosterone and cocaine did not affect rest HR (p>0.05) (Table 1).
One- or 10-day administration of testosterone or cocaine did not affect basal values of tail skin temperature (p>0.05) (Table 1). ANOVA did not indicate significant effect of either treatment regimen (F(1,38)=0.8, p>0.05) or drug factors (F(3,38)=0.6, p>0.05).
Effect of one- and 10-day single or combined administration of testosterone and cocaine in autonomic responses to acute restraint stress
Acute restraint stress induced a marked and sustained increase in HR (F(16,608)=123.3, p<0.0001) and MAP (F(16,608)=143.2, p<0.0001), as well as a maintained decrease in tail cutaneous temperature (F(8,304)=160.2, p<0.0001) (Figure 1). Treatment with veh+coc and T+coc for 10 days decreased MAP response to restraint stress (F(3,19)=8.8, p<0.0005) when compared with veh+sal group within the same treatment regimen (Figure 2). There were no statistical differences in restraint-evoked pressor responses between veh+coc and T+coc groups (p>0.05) (Figure 1). Analysis of MAP response indicated a significant effect of treatment regimen (F(1,38)=14.1, p<0.008), drug (F(3,38)=2.9, p<0.004) and time (F(16,608)=143.0, p<0.0001). There were also significant treatment regimen versus drug (F(3,38)=3.2, p<0.03), treatment regimen versus time (F(16,608)=5.9, p<0.06) and drug versus time (F(48,608)=2.3, p<0.006) interactions. One-day single or combined administration of testosterone and cocaine did not affect MAP increase induced by restraint stress (F(3,19)=0.3, p>0.05) (Figure 1).
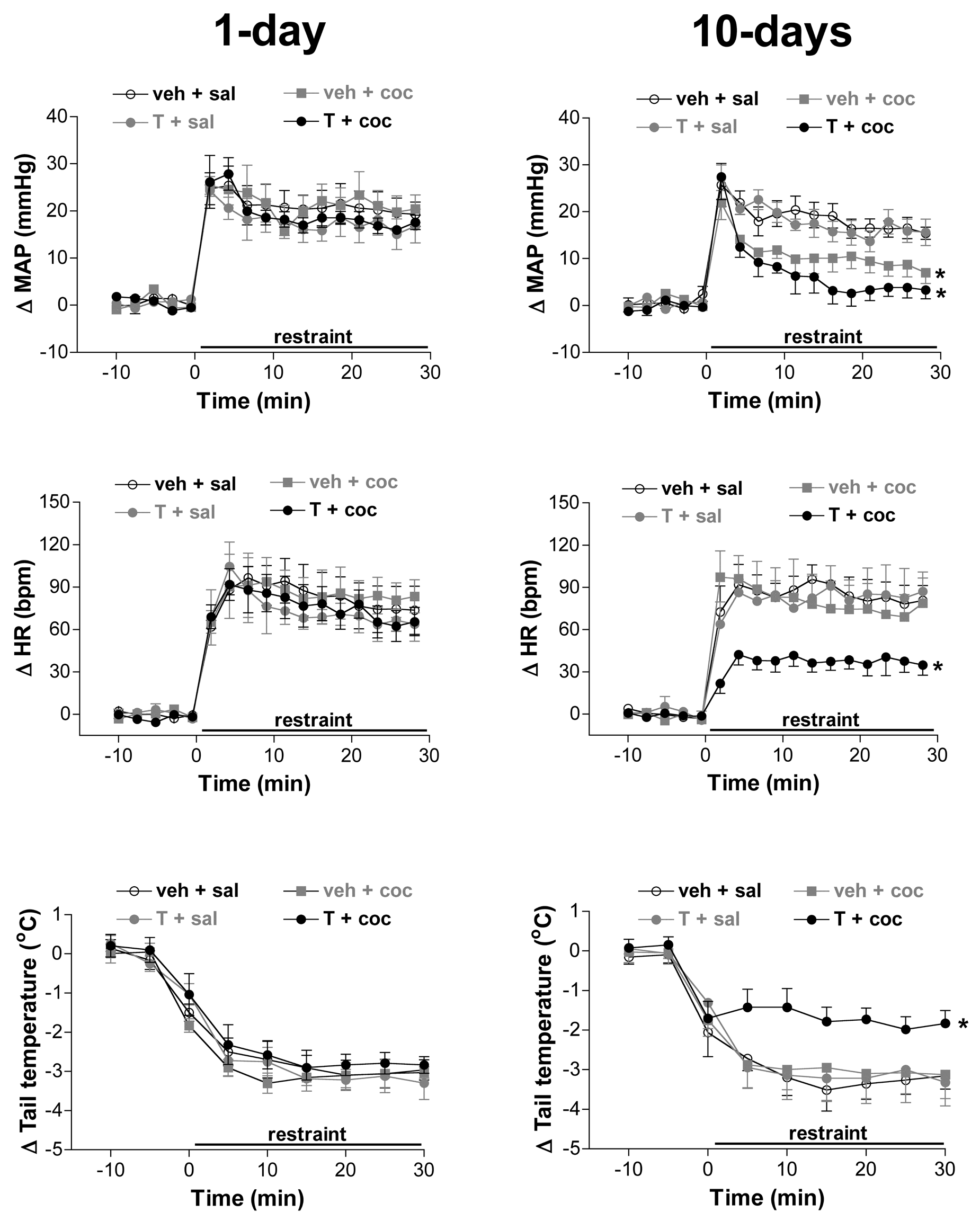
Time course of changes in mean arterial pressure (Δ MAP), heart rate (Δ HR), and tail skin temperature (Δ tail temperature) induced by restraint stress in animals subjected to single or combined administration of testosterone and cocaine for one (One day) or 10 consecutive days (10 days). The onset of restraint was at t=0. Circles represent the mean and bars the SEM. *p<0.05 over the whole restraint period compared with the 10-day veh+sal group, ANOVA followed by Bonferroni’s post hoc test.
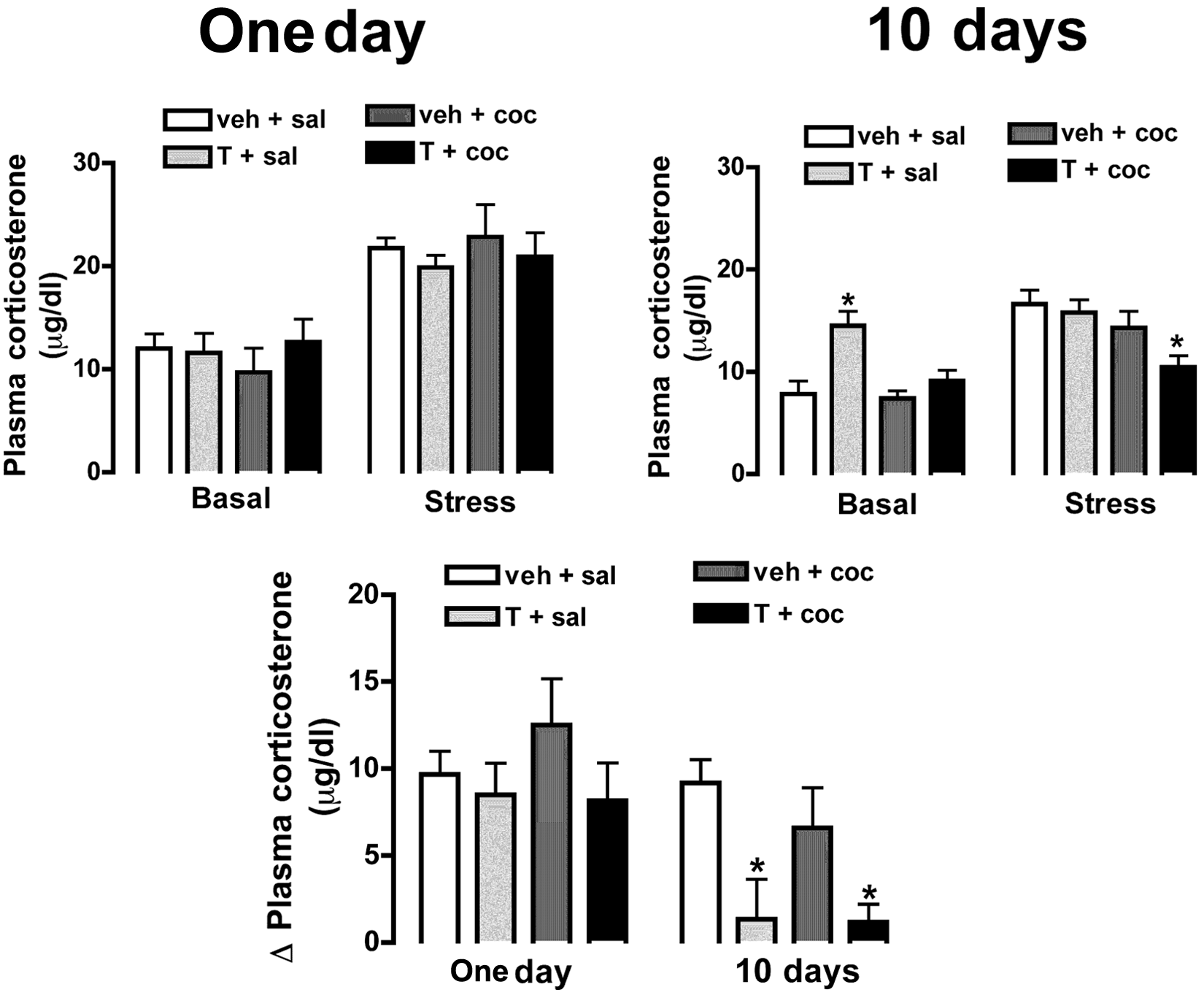
Top: plasma corticosterone levels before (basal) or during restraint stress (stress) in animals subjected to single or combined administration of testosterone and cocaine for one (One day) or 10 consecutive days (10 days). Bottom: change in corticosterone level induced by restraint stress (difference between values during stress and basal) (Δ plasma corticosterone) in animals subjected to single or combined administration of testosterone and cocaine for one (One day) or 10 consecutive days (10 days). Columns represent the mean and bars the SEM. *p<0.05 vs. 10-day veh+sal group at same period (basal or stress), ANOVA followed by Bonferroni’s post hoc test. coc: cocaine; sal: saline; T: testosterone; veh: vehicle
Combined administration of testosterone and cocaine for 10 days reduced tachycardiac response to restraint (F(3,19)=4.8, p<0.008) when compared with the veh+sal group within the same treatment regimen (Figure 1). Analysis of HR response indicated a significant effect of drug (F(3,38)=3.2, p<0.05) and time (F(16,608)=123.2, p<0.0001), but without effect of treatment regimen (F(1,38)=0.8, p>0.05). There were no treatment regimen versus drug (F(3,38)=0.9, p>0.05), treatment regimen versus time (F(16,608)=0.8, p>0.05) or drug versus time (F(48,608)=1.1, p>0.05) interactions. One-day single or combined administration of testosterone and cocaine did not affect restraint-evoked HR increase (F(3,19)=0.2, p>0.05) (Figure 1).
Combined administration of testosterone and cocaine for 10 days reduced the restraint-evoked decrease of tail cutaneous temperature (F(3,19)=3.3, p<0.05) when compared with veh+sal group within the same treatment regimen (Figure 1). Analysis of tail temperature response indicated a significant effect of drug (F(3,38)=3.5, p<0.03) and time (F(8,304)=160.2, p<0.0001), but not of treatment regimen (F(1,38)=0.1, p>0.05). There were no treatment regimen versus drug (F(3,38)=0.6, p>0.05), treatment regimen versus time (F(8,304)=0.8, p>0.05) or drug versus time (F(24,304)=1.7, p>0.05) interactions. One-day single or combined administration of testosterone and cocaine did not affect restraint-evoked tail temperature decrease (F(3,19)=0.5, p>0.05) (Figure 1).
Effect of one- and 10-day single or combined administration of testosterone and cocaine in neuroendocrine response to acute restraint stress
Acute restraint stress induced an increase in plasma concentration of corticosterone (F(1,38)=109.1, p<0.0001) (Figure 2). ANOVA also indicated a significant effect of treatment regimen (F(1,38)=21.3, p<0.003), but not of drug (F(3,38)= 1.4, p>0.05), on stress-induced increase in plasma corticosterone concentration. In addition, there were significant treatment regimen versus time (F(1,38)=14.7, p<0.0005) and drug versus time (F(3,38)=4.8, p<0.01) interactions, while no interaction between treatment regimen and drug was detected (F(3,38)=1.8, p>0.05).
Administration of T+sal for 10 days increased basal levels of plasma corticosterone (F(3,19)=8.3, p<0.001) when compared with veh+sal group within the same treatment regimen (Figure 2). Also, combined administration of testosterone and cocaine for 10 days reduced plasma corticosterone levels during restraint stress (F(3,19)=5.5, p<0.01) when compared with veh+sal group within the same treatment regimen (Figure 2). One-day single or combined administration of testosterone and cocaine did not affect either basal values (F(3,19)=0.2, p>0.05) or stress-induced increase in plasma corticosterone levels (F(3,19)=0.3, p>0.05) (Figure 2).
Analysis of the change in plasma corticosterone levels induced by restraint indicated a significant effect of treatment regimen (F(1,39)=13.7, p<0.001) and drug (F(3,39)=3.3, p<0.01), but there was no interaction between factors (F(3,39)=0.6, p>0.05) (Figure 2). Administration of T+sal (p<0.05) and T+coc (p<0.05) for 10 days reduced restraint-evoked change in plasma corticosterone concentration (F(3,19)=6.3, p<0.009) (Figure 2). One-day single or combined administration of testosterone and cocaine did not affect the increase in plasma corticosterone levels induced by restraint stress (F(3,19)=1.5, p>0.05) (Figure 2).
Discussion
The main findings of the present study are: 1) combined administration of testosterone and cocaine for 10 days significantly increased basal values of arterial pressure; 2) single treatment with testosterone for 10 days caused resting bradycardia and increased plasma baseline levels of corticosterone; 3) combined administration of testosterone and cocaine for 10 days reduced both cardiovascular, tail cutaneous temperature and plasma glucocorticoid responses to acute restraint stress; 4) 10-day single treatment with cocaine reduced restraint-evoked increase in arterial pressure; and 5) one-day single or combined administration of testosterone and cocaine did not produce long-lasting changes that altered either autonomic or neuroendocrine responses induced by restraint stress, suggesting that chronic exposure to these substances is necessary to evoke alterations in physiological responses to stress.
Cocaine abuse is associated with both acute and chronic cardiovascular toxicity (Kloner et al., 1992; Maraj et al., 2010). Acute actions of cocaine include a range of cardiovascular effects such as hypertension, coronary vasoconstriction and cardiac arrhythmias (Iwasaki-Sekino et al., 2009; Maraj et al., 2010). These effects are mediated by the sympathomimetic activity and local anesthetic action of cocaine (Goldstein et al., 2009). However, cocaine-induced cardiovascular complications are mainly due to its chronic abuse, which has been poorly investigated (Isner et al., 1986; Kloner et al., 1992). Unlike cocaine, AAS evokes minor acute cardiovascular side effects (van Amsterdam et al., 2010). However, chronic AAS abuse has been associated with hypertension and cardiac pathologies (Sullivan et al., 1998; van Amsterdam et al., 2010). Preclinical studies suggest that AAS and cocaine are capable of mutually potentiating the cardiovascular effects of each other (Phillis et al., 2000; Togna et al., 2003). Indeed, our findings of increased arterial pressure baseline in the T+coc group corroborate previous preclinical results demonstrating that combined administration of AAS and cocaine increases arterial pressure in normotensive animals and accelerates the rate of development of hypertension in a strain of rats genetically predisposed to hypertension (spontaneously hypertensive rats) (Tseng et al., 1994; Engi et al., 2011). Mechanisms involved in arterial pressure elevation following simultaneous exposure to AAS and cocaine are not entirely understood. Previous results from our laboratory demonstrated that combined treatment with testosterone and cocaine evokes alterations in baroreflex activity and vascular responsiveness to vasoactive agents in rats (Engi et al., 2011), which may contribute to the elevation in arterial pressure. As previously reported (Beutel et al., 2005; Engi et al., 2011), chronic single administration of testosterone caused rest bradycardia. However, the mechanism remains unclear.
There is evidence demonstrating that cocaine and testosterone interact to induce behavioral changes in rodents (Long et al., 1994; Martinez-Sanchis et al., 2002; Menendez-Delmestre and Segarra, 2011). However, to our knowledge, this is the first study to show the interaction between these substances in autonomic and neuroendocrine responses to stress. We observed that combined administration of testosterone and cocaine for 10 days reduced both tachycardiac, skin temperature and corticosterone response to restraint stress. Changes in autonomic activity and HPA axis activation are part of the physiological responses necessary to the metabolic adjustments required to maintain homeostasis during exposure to aversive stimuli (Ulrich-Lai and Herman, 2009). Therefore, changes in these physiological responses may affect the ability to cope with stressful events (Ulrich-Lai and Herman, 2009). Since both testosterone and cocaine exposure affect neural activity of limbic structures involved in physiological responses to stress (Johansson-Steensland et al., 2002; Liu et al., 2005; Chocyk et al., 2006; Orozco-Cabal et al., 2008; Ambar and Chiavegatto, 2009), alterations in neurocircuitry of stress following chronic exposure to testosterone and cocaine could mediate the reduction in autonomic and neuroendocrine response to restraint. However, further studies are required to identify the mechanisms that mediate the combined effects of testosterone and cocaine on stress-evoked physiological adjustments.
Cardiac sympathetic blockers abolish tachycardia response evoked by stress, whereas cardiac parasympathetic blocker increases it (Baudrie et al., 1997; Carrive, 2006; Crestani et al., 2009, 2010). These results suggest a coactivation of cardiac sympathetic and parasympathetic activity during stress. Thus, reduction in restraint-evoked HR rise following combined administration of testosterone and cocaine may result from a decrease in cardiac sympathetic activation or facilitation of cardiac parasympathetic response. However, changes in cardiac response to stress may also be related to a direct action of either testosterone or cocaine on the heart. Prolonged reduction of noradrenaline reuptake in the cardiac sympathetic nerve terminals has been reported in chronic cocaine users (Melon et al., 1997). Moreover, long-term exposure of cultured rat cardiomyocytes to testosterone increases cardiac T-type calcium currents (Michels et al., 2006), thus indicating that this drug could play a role in enhancing HR and cardiac contractility.
The arterial pressure rise during stress is mediated by an increase in vascular sympathetic tone and activation of α1-adrenoreceptors in vascular smooth muscle (Baudrie et al., 1997). Ten-day single administration of cocaine decreased restraint-evoked pressor response, and this effect was not affected by coadministration of testosterone. We have previously reported that chronic cocaine administration reduced vascular responsiveness to the selective α1-adrenoceptor agonist phenylephrine (Engi et al., 2011), suggesting that cocaine may desensitize α1-adrenoceptor in vascular smooth muscles. This finding may explain, at least in part, the reduced pressor response to restraint in cocaine-treated animals. Moreover, cocaine exposure evokes changes in neural activity of limbic structures such as the medial prefrontal cortex, amygdala, hypothalamus regions and lateral septal area (Liu et al., 2005; Chocyk et al., 2006; Orozco-Cabal et al., 2008). Many studies have demonstrated the involvement of these structures in cardiovascular responses to different types of stress in animals (Resstel et al., 2006; Fortaleza et al., 2009; Busnardo et al., 2010; Reis et al., 2011). Therefore, cocaine-induced changes in the neurocircuitry of stress could also mediate the reduction in the pressor response to stress. This hypothesis is further supported by previous results demonstrating that cocaine may reduce sympathetic neural activity through an action in the central nervous system (Raczkowski et al., 1991).
Reduction in cutaneous blood flow caused by stress is mediated by increase in sympathetic vasoconstrictor tone and activation of α-adrenoceptors (Blessing, 2003). Cutaneous vasoconstriction may either prevent blood loss due to injuries, by keeping low amounts of blood in the skin, or redistribute blood to more important organs during stress, such as muscles and the brain. Cutaneous vasoconstriction during stress evokes a rapid skin temperature drop, and fall in tail cutaneous temperature can be used as an indirect measurement of blood flow redistribution by the sympathetic nervous system in the rat (Blessing, 2003; Vianna and Carrive, 2005). As stated above, single administration of cocaine desensitizes α1-adrenoceptor in vascular smooth muscles (Engi et al., 2011). Interestingly, in the present study, single administration of cocaine did not affect restraint-evoked fall of tail skin temperature. Reduction in tail temperature response was observed only after combined administration of testosterone and cocaine. These findings suggest that the effect of cocaine on vascular responsiveness to sympathomimetic substances is vascular bed–specific. However, in vitro studies have demonstrated that AAS is capable of potentiating the vascular effects of cocaine (Togna et al., 2003). Therefore, we cannot exclude the possibility that reduced tail temperature response following combined administration of testosterone and cocaine can also result from a decreased vascular responsiveness to sympathetic activation.
Activation of the HPA axis is a primary response during stress (Ulrich-Lai and Herman, 2009). However, glucocorticoid secretion needs to be tightly regulated since HPA axis hyperactivity is related to metabolic, psychological and cardiovascular dysfunctions (Rosmond and Bjorntorp, 2000; Jokinen and Nordstrom, 2009). Therefore, elevated basal levels of glucocorticoids following testosterone treatment may predispose to the development of several pathologies. Clinical and preclinical evidences of the influence of androgens in basal plasma glucocorticoids level are controversial (Barbosa et al., 1971; Rozenek et al., 1990; Handa et al., 1994; Toufexis and Wilson, 2012). Thus, further studies are necessary to study the mechanisms involved in increased basal level of corticosterone in testosterone-treated animals. However, Williamson and Viau (2008) demonstrated, in gonadectomized rats, that high-dose testosterone replacement increased arginine vasopressin (AVP) staining localized to corticotropin-releasing hormone (CRH) positive terminals in the median eminence without affecting CRH (Williamson and Viau, 2008). Although AVP is a weak adrenocorticotropin (ACTH) secretagog, it enhances the stimulatory effect of CRH on ACTH release (Antoni, 1993). Therefore, increase in AVP synthesis in paraventricular nucleus of the hypothalamus (PVN) parvocellular neurons may explain, at least in part, the elevation in plasma glucocorticoid level following testosterone treatment. Interestingly, coadministration of cocaine inhibited testosterone effect on basal plasma corticosterone. It has been demonstrated that chronic cocaine exposure reduces basal plasma corticosterone (Mantsch and Goeders, 2000), possibly due to a reduced pituitary function (Mantsch et al., 2003). Therefore, cocaine may counteract facilitatory influence of testosterone in the HPA axis.
Our results showed that stress-induced increase in plasma corticosterone levels was attenuated in testosterone-treated animals. This finding can be related to the increased basal levels of plasma corticosterone. Also, sex difference in the ACTH and glucocorticoid response to acute stressors suggests a modulatory effect of androgens in HPA axis response induced by stress (Viau, 2002; Williamson et al., 2005; Handa et al., 2009). Indeed, numerous studies have consistently demonstrated an inhibitory influence of testosterone, as well as other androgens, in HPA axis response to acute stress (Handa et al., 1994; Patchev and Almeida, 1996; Viau and Meaney, 1996; Williamson et al., 2005; Iwasaki-Sekino et al., 2009; Toufexis and Wilson, 2012). Although the above evidences, further studies are necessary to study the mechanisms involved in the influence of exogenous testosterone in the HPA axis.
In summary, the present study adds evidences to previous findings showing interaction between AAS and cocaine. Our findings demonstrate that simultaneous exposure to cocaine and testosterone evokes changes in cardiovascular and neuroendocrine activity. The present study suggests that the interaction between testosterone and cocaine may affect the ability to cope with stressful events. In addition, our data showed that chronic exposure to high levels of testosterone increases the activity of the HPA axis. Together, these results may contribute to the better understanding of the toxicity of testosterone and cocaine reported in clinical research.
Footnotes
Acknowledgements
The authors wish to thank Elisabete ZP Lepera and Rosana FP Silva for technical assistance.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article.
This work was supported by the São Paulo Research Foundation (FAPESP) (grant numbers 2010/16192-8); the National Council for Scientific and Technological Development (CNPq) (grant numbers 474177/2010-6); and PADC-School of Pharmaceutical Sciences-São Paulo State University.
Conflict of interest
The authors declare that they do not have any conflict of interest.