
Abstract
Evidence indicates that the serotonergic system is important in mediating dependence on and craving for alcohol. Among serotonin receptors, 5-hydroxytryptamine 1B (5-HT1B) receptors have been associated with drug abuse including alcohol. In this review, the neurocircuitry involving 5-HT1B receptors in central reward brain regions related to alcohol intake are discussed in detail. Emphasis has been placed on the pharmacological manipulations of 5-HT1B receptor-mediated alcohol intake. Furthermore, 5-HT1B auto- and hetero-receptors regulate alcohol intake through the regulatory mechanism involving release of 5-HT, gamma-aminobutyric acid (GABA), dopamine, and glutamate is evaluated. Thus, interactions between 5-HT1B receptors and these neurotransmitter systems are suggested to modulate alcohol-drinking behavior. This review on the role of 5-HT1B receptors in neurotransmitter release and consequent alcohol intake provides important information about the potential therapeutic role of 5-HT1B receptors for the treatment of alcohol dependence.
Introduction
Alcohol abuse and dependence continue to be significant public health problems (Harper, 2009; Heilig and Egli, 2006; Li, 2008; Room et al., 2005; Sullivan and Zahr, 2008; Volkow and Li, 2005). Alcohol dependence, known also as alcoholism, is considered one of the high severity disorders (Schuckit, 2009). Alcohol dependence is a cluster of cognitive behavior and physiological functions that can be developed after repeated use of alcohol. Chronic consumption of alcohol can induce alteration in neuronal function, which may lead to sensitization, tolerance, withdrawal, and dependence to alcohol (Diana et al., 2003). Emerging evidence indicates that many aspects of alcohol dependence involve the serotonergic system (for review, see Sari et al., 2011).
The serotonin (5-hydroxytryptamine (5-HT)) system is involved in alcohol intake, abuse and dependence (Ait-Daoud et al., 2009; LeMarquand et al., 1994a; LeMarquand et al., 1994b; Sari et al., 2011; Sellers et al., 1992). Decreased 5-HT function in central brain reward regions is associated with increased alcohol consumption (Balldin et al., 1994; LeMarquand et al., 1994a). 5-HT inversely mediates alcohol intake, such that an increase in 5-HT function may lead to a decrease in alcohol intake and a decrease in 5-HT function may lead to an increase in alcohol intake (LeMarquand et al., 1994b). Dysfunction of the 5-HT system often occurs in individuals who either abuse alcohol or are alcohol-dependent (LeMarquand et al., 1994a). Brain imaging studies have shown reduced 5-HT transporter levels in alcoholics (Heinz et al., 1998; Heinz et al., 2001). It is noteworthy that the 5-HT transporter genotype has a significant influence on alcohol craving in alcoholic individuals. Alternatively, the concentration of 5-HT and its metabolite 5-hydroxy indole acetic acid (5-HIAA) in a central reward region such as the nucleus accumbens (NAc) were lower in selectively bred, high alcohol-drinking rats when compared with their low alcohol-drinking counterparts (McBride et al., 1995). This parallels a previous study, which also demonstrated that selectively bred, alcohol-preferring rats have reduced levels of 5-HT and 5-HIAA compared to alcohol-non-preferring rats (Murphy et al., 1982). Note that alcohol-preferring rats exhibit behavioral, physiological and neurochemical traits similar to those observed in alcoholic humans (Bell et al., 2005; Bell et al., 2006a; Bell et al., 2006b; McBride and Li, 1998; Murphy et al., 2002; Rodd et al., 2004).
Clinical studies indicate that selective serotonin reuptake inhibitors (SSRIs; e.g. fluoxetine and paroxetine) can be used to treat alcohol addiction in subgroups of alcoholics (Heinz et al., 1998; Pettinati, 2001). SSRI-induced decreases in 5-HT turnover are associated with a reduced reward response to excessive alcohol consumption in the dorsal raphe nuclei of non-human primates (Wrase et al., 2006). In addition, similar studies have demonstrated that the administrations of SSRIs decrease neuronal firing in the ventral tegmental area (VTA), a central brain reward region (Esposito, 1996; Prisco and Esposito, 1995). Together, these suggest that dysfunction in 5-HT neurotransmission might be a key player in the initiation and maintenance of alcoholism. Changes in the release of 5-HT are considered key factors in mediating the activation of several 5-HT receptors in central brain reward regions. Among these are the 5-HT1A, 5-HT1B and 5-HT3 receptors. The identification of these subtypes of 5-HT receptors which regulate alcohol intake in rodents might be of particular importance in developing pharmacotherapies for alcohol dependence. The present focus has been on the role of the 5-HT1B receptor in alcohol-drinking behavior because of its involvement in the release of several neurotransmitters mediating the regulation of alcohol intake (for review see Sari, 2004). Furthermore, quantitative trait analysis of Bithoraxoid (BXD) recombinant mice demonstrated that the 5-HT1B receptor gene is considered a potential candidate for influencing alcohol preference (Crabbe et al., 1999). Interestingly, polymorphisms of the 5-HT1B receptor gene have been linked to alcoholism (Lappalainen et al., 1998).
In recent decades, findings have demonstrated that 5-HT1B receptors are involved in multiple physiological functions, behaviors and psychiatric diseases (Fernandez-Guasti et al., 1992; Kaiyala et al., 2003; Lin and Parsons, 2002; Ramboz et al., 1996; Sari, 2004). The 5-HT1B receptor (human analog 5-HT1Dβ) contains seven transmembrane domains, of which one is composed of eight amino acids and acts as the binding site for a specific ligand (Findlay and Eliopoulos, 1990; Hibert et al., 1991; Trumpp-Kallmeyer et al., 1992). The 5-HT1B receptor is coupled to an intermediate inhibitory G protein, which acts as a second messenger that can lead to a decrease in the activity of adenylyl cyclase (Bouhelal et al., 1988; Hamblin and Metcalf, 1991; Maroteaux et al., 1992; Schoeffter and Hoyer, 1989; Seuwen et al., 1988).
It is important to note that 5-HT1B receptors have been demonstrated to play a key role in drug abuse reinforcement, including cocaine and alcohol (for review see Sari, 2004). Activation of 5-HT1B receptors potentiates the reinforcing effects of cocaine (Parsons et al., 1999). Studies using pharmacological, molecular, and genetic manipulations have demonstrated that 5-HT1B receptors play a key role in alcohol dependence (Crabbe et al., 1996; Hoplight et al., 2006; Maurel et al., 1999a; Maurel et al., 1999b; Miczek and De Almeida, 2001; Risinger et al., 1999; Tomkins and O’Neill, 2000). Voluntary alcohol consumption appears to be mediated by the 5-HT1B receptors innervating the mesocorticolimbic brain regions involved in the reward circuit. These central reward brain regions originate from VTA neurons and project to the NAc, olfactory tubercle, frontal cortex, and in particular, the amygdala complex (Asan, 1998). Dopaminergic VTA projections are most intensely concentrated in the central nucleus of the amygdala (CeA), the bed nucleus of the stria terminalis (BNST), and the NAc shell; these regions constitute the ‘extended amygdala’ (For review see Alheid, 2003).
Within this neurocircuitry, pharmacological and ligand binding studies show lower densities of 5-HT1B receptors in the central nervous system of alcohol-preferring rats compared to alcohol-non-preferring rats (McBride et al., 1997). In addition, pharmacological studies suggest that both 5-HT1B auto- and hetero-receptors are involved in the modulation of alcohol-drinking behavior. The 5-HT1B receptor mRNA and its protein, 5-HT1B hetero-receptors, are expressed in the NAc (Bruinvels et al., 1993; Bruinvels et al., 1994; Mengod et al., 1990), suggesting that 5-HT1B auto-receptors located at the terminals of dorsal raphe serotonergic neurons in the NAc and VTA may play a significant role in drug reinforcement, including that of alcohol (for review see Sari, 2004). It is noteworthy that there are no structural differences between 5-HT1B auto- and hetero-receptors. However, studies have shown that there exists differential sensitivity between 5-HT1B auto- and hetero-receptors (Rossetti and Carboni, 1995). This differential sensitivity between 5-HT1B auto- and hetero-receptors may be due to differences in the level of receptors at different synapses which may influence sensitivity under conditions such as partial occupancy.
In this review, the pharmacological manipulations of 5-HT1B receptors and the use of 5-HT1B receptor knockout mice have been discussed in detail. The discussion focuses also on the role of 5-HT1B auto- and hetero-receptors in alcohol intake. In addition, the interactions between 5-HT1B receptors and other neurotransmitters, including dopamine, gamma-aminobutyric acid (GABA) and glutamate, in the modulation of alcohol intake have been assessed in this review. Emphasis has been on the role of 5-HT1B receptors in neurotransmitter release for the regulation of alcohol intake.
Animal models and pharmacological manipulations of 5-HT1B receptor-mediated alcohol consumption
Knockout models, alcohol-preferring animal models, and pharmacological manipulations of 5-HT1B receptors in alcohol-drinking behavior
The role of 5-HT1B receptors in alcohol consumption has been investigated in 5-HT1B knockout mice (genotype -/-). The results generated by several studies suggest that 5-HT1B receptors mediate alcohol intake: however, some controversy still exists. Although 5-HT1B receptor knockout mice phenotypically consumed alcohol at an increased rate in initial studies (Crabbe et al., 1996; Risinger et al., 1996), later studies using different substrains of knockout mice have failed to reproduce this phenotypic characteristic (Bouwknecht et al., 2000; Crabbe et al., 1999; Risinger et al., 1999). For example, in one study, 5-HT1B receptor knockout mice showed greater levels of alcohol consumption for unsweetened 10% ethanol but showed moderate alcohol self-administration overall (Risinger et al., 1999). Contrastingly, other studies demonstrated that alcohol intake (g/kg body weight) was similar for wild type and 5-HT1B receptor knockout mice (Bouwknecht et al., 2000). However, in the latter case, body weights and water intake were elevated in these knockout mice which suggests that the initial finding of elevated ethanol consumption might actually be related to differences in fluid intake. Although alcohol intake is not consistent in knockout mice, these differences might be due to substantial genetic drift over generations, including population adaptations.
Variant results related to 5-HT1B receptor knockout mice in alcohol consumption studies suggest that phenotypic differences in mutants may exist (Gerlai, 1996; Phillips et al., 1999). Previous studies suggest the possibility of genetic drift and genetic crossover from multiple substrains of knockout mice during the course of breeding (Phillips et al., 1999). Additionally, dissimilarities in each experimental design may explain the discrepancies in alcohol consumption in 5-HT1B receptor knockout mice. For example, studies performed in multiple laboratories showed low alcohol intake (6%, v/v) in 5-HT1B receptor knockout mice tested for four days (Crabbe et al., 1999), while other studies showed high alcohol intake (for multiples concentrations of alcohol 3–20%, v/v) in mice tested for 12 days (Crabbe et al., 1996).
The involvement of 5-HT1B receptors in alcohol-drinking behavior has also been supported by pharmacological studies conducted with divergent animal strains (Tables 1 and 2). Administration of CGS12066B, a partial 5-HT1B receptor agonist, in the NAc core decreased ethanol intake in Long-Evans rats (Czachowski, 2005). There are 5-HT1B autoreceptors located at the terminals of the dorsal raphe-NAc pathway and 5-HT1B heteroreceptors located at the terminals of the glutamatergic cortico-accumbal pathway. Therefore, the infusion of 5-HT1B receptor agonist in the NAc may lead to inhibition of glutamate release and in turn lead to disinhibition of dopamine neurons in the VTA–NAc pathway. This regulatory mechanism would lead to a reduction in alcohol consumption.
Summary of findings investigated the effects of intraperitoneal (i.p.) injections of drugs targeting 5-hydroxytryptamine 1B (5-HT1B) receptors in the regulation of alcohol consumption.
Summary of findings investigated the effects of intracerebro-injections of viral vector and agonist of 5-hydroxytryptamine 1B (5-HT1B) receptors in the regulation of alcohol-drinking behavior.

In an operant oral self-administration of alcohol (10%, v/v) study, intraperitoneal (i.p.) injections of CP-94,253, a 5-HT1B receptor agonist, reduced ethanol-reinforced lever pressing (Maurel et al., 1999b). In addition, the 5-HT1B receptor agonist m-chlorophenylpiperazine (mCPP) induced a general decrease in alcohol intake in the Cologne Alcohol-Addicted (cAA) rat model of alcoholism (Maurel et al., 1999a). However, this effect may be relatively nonselective for alcohol consumption because mCPP binds non-selectively to 5-HT1B and 5-HT2C receptors. Thus, it is unclear whether these effects of mCPP are due to stimulation of 5-HT2C receptors, 5-HT1B receptors, or both. Furthermore, studies using Wistar rats have demonstrated that i.p. administration of a partial 5-HT1B receptor agonist, CGS12066B, also reduced oral ethanol self-administration (12%, v/v ethanol) (Tomkins and O’Neill, 2000). Moreover, 5-HT1B receptor agonist, RU24969, is the most commonly used agonist in investigating different behavioral paradigms and neurochemical consequences of 5-HT1B receptor activation (Fletcher et al., 1992; Halford and Blundell, 1996; O’Neill et al., 1996; Oberlander et al., 1987). In Wistar rats, i.p. administration of RU24969 reduced oral ethanol self-administration (12% v/v ethanol) (Tomkins and O’Neill, 2000). Regarding the selectivity of RU24969, this compound has similar affinity for both 5-HT1B and 5-HT1A receptors (Middlemiss and Tricklebank, 1992), but several studies have suggested that the action of RU24969 is more specific to 5-HT1B receptors (Fernandez-Guasti et al., 1992; Fletcher et al., 1992; O’Neill et al., 1996; Oberlander et al., 1987). The action of RU24969 is reversed by the 5-HT1B receptor antagonist GR127935 but not by the 5-HT1A receptor antagonist WAY100135 (O’Neill et al., 1996). GR127935 is a potent and selective 5-HT1B/1D antagonist that shows about 1000-fold selectivity for the 5-HT1B receptor compared to the 5-HT1A, 5-HT2C, and 5-HT2A receptors (Skingle et al., 1996).
Lastly, administration of anpirtoline, another 5-HT1B receptor agonist, significantly decreased ethanol-reinforced behavior in Swiss Webster mice (Miczek and De Almeida, 2001). Anpirtoline binds to both 5-HT1B and 5-HT1A receptors, with higher affinity for 5-HT1B receptors. Again, it is unclear whether its effect on alcohol reinforcement is dependent on 5-HT1B receptors, 5-HT1A receptors, or both. In male ejaculatory behavior study, anpirtoline’s effects were prevented by the application of 5-HT1B receptor antagonists (Hillegaart and Ahlenius, 1998). In addition, a study that investigated the metabolism and synthesis of 5-HT has shown similar effects (Stenfors et al., 2000). This suggests that anpirtoline selectively acts on 5-HT1B receptors, at least in some behavioral and chemical assays. These findings show that ethanol-reinforced behavior is inhibited by selective activation of 5-HT1B receptors (Tomkins and O’Neill, 2000) and reduced alcohol intake. The following mechanism of action is suggested: when 5-HT1B receptor agonist is administered i.p., it deactivates the release of glutamate at either the cortico-NAc or cortico-VTA pathway since the 5-HT1B receptor has an inhibitory effect
Viral vector overexpression of 5-HT1B receptors in the regulation of alcohol-drinking behavior
Studies have utilized herpes simplex virus (HSV) viral vector to overexpress 5-HT1B receptors in the investigation of alcohol consumption. Both hemagglutinin-tagged and green fluorescent protein-tagged (HA1B/GFP) 5-HT1B receptors have been overexpressed using HSV vectors (Clark et al., 2002; Clark et al., 2004; Hoplight et al., 2006). Viral vector gene expression is advantageous due to its high selectivity and low toxicity for target neurons (Carlezon et al., 2000). Bilateral microinjections of HSV vectors carrying 5-HT1B receptors in the medial NAc shell increased voluntary alcohol consumption in Long-Evans rats in a two-week free-choice alcohol (6% or 12%) drinking protocol (Hoplight et al., 2006). Groups that had received the viral vector HA1B/GFP showed an increase in alcohol intake compared to the control groups. Viral vector overexpression of 5-HT1B receptors specifically targets the medium spiny neurons in the NAc shell which express 5-HT1B receptors. Interestingly, the expression of 5-HT1B receptors on 5-HT projections from the dorsal raphe nucleus innervating the NAc were not affected (Clark et al., 2002). This suggests that overexpression is specific for 5-HT1B heteroreceptors located at the terminals of the VTA-NAc pathway. Importantly, a recent study showed that overexpression of 5-HT1B receptors in the NAc shell alters the preference for low versus high ethanol concentration and also alters the drinking behavior pattern during both initiation and maintenance phases of ethanol consumption (Furay et al., 2011).
Pharmacological actions of 5-HT1B receptors in neurotransmitter release and subsequent modulation of alcohol-drinking behavior
Although the mechanism by which 5-HT1B ligands regulate alcohol self-administration is not fully known, it is possible that the regulatory effect is achieved through indirect manipulation of dopamine neurotransmission via GABAergic activity (Figure 1). Previous studies showed that intra-tegmental administration of a 5-HT1B receptor agonist, CP-93,129, increased dopamine release in both the VTA and the NAc (Yan and Yan, 2001; Yan et al., 2004). In contrast, infusion of a 5-HT1B receptor antagonist, SB-216641, in the VTA attenuated ethanol-induced (i.p.) increases in extracellular dopamine in both VTA and NAc (Yan et al., 2005). This suggests that 5-HT1B receptors interact indirectly with mesolimbic dopamine systems during alcohol intake. It is postulated here that activation of 5-HT1B receptor in the VTA may induce inhibition of GABA release, which consequently leads to increases in dopamine release in both the VTA and NAc; this regulatory mechanism would be a key factor mediating increased alcohol intake. However, administration of the 5-HT1B receptor agonist, CGS12066B, in the NAc core decreases ethanol-intake (Czachowski, 2005). It is possible that administering a 5-HT1B receptor agonist directly into the NAc may induce decreased neuronal activity by binding to the 5-HT1B receptor subtype and inhibiting presynaptic release of glutamate in the NAc ((Hoyer et al., 1994; Johnson et al., 1992a; Morikawa et al., 2000; Muramatsu et al., 1998), Figure 1). Therefore, 5-HT1B receptor agonist infusion into NAc may bind to 5-HT1B receptors located at the terminals of cortico-accumbal glutamatergic projections and thus inhibit glutamate release, which consequently decreases dopamine release in the NAc, reducing ethanol-associated reward and leading to reduction in alcohol intake.
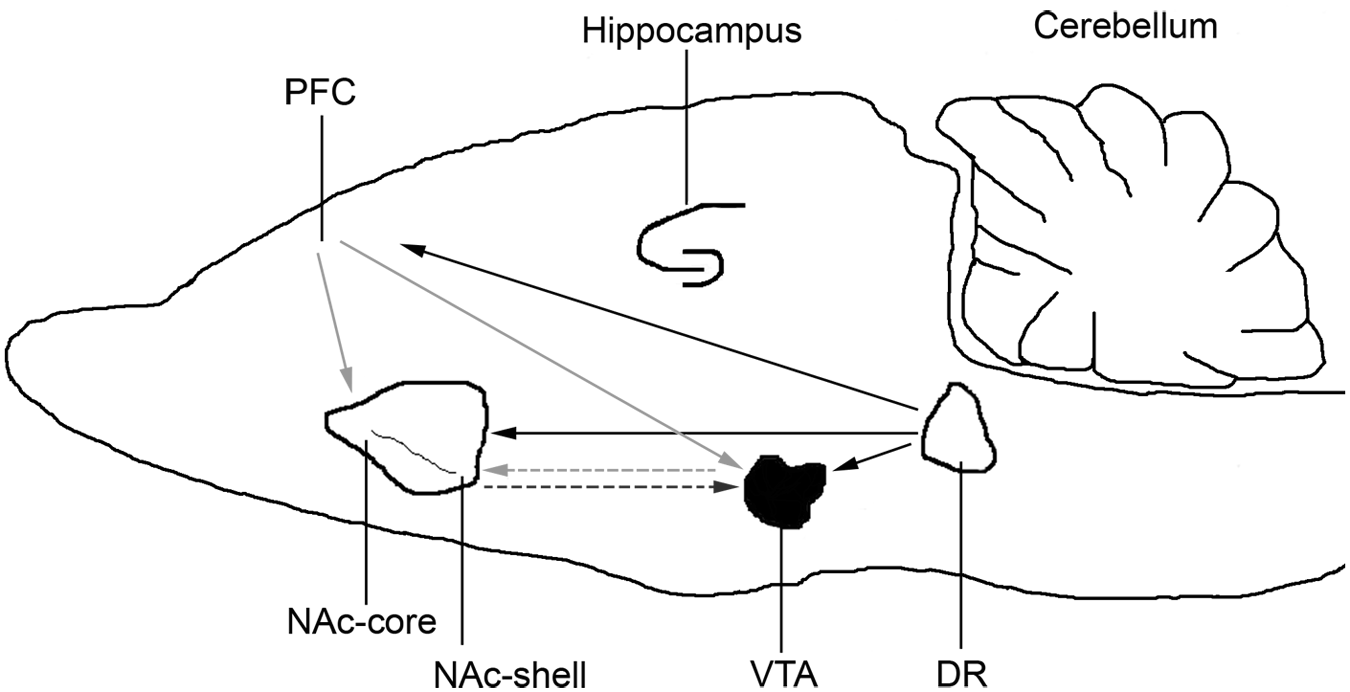
Schematic representation of the neurocircuitry of 5-hydroxytryptamine 1B (5-HT1B) receptors involving releases of neurotransmitters in the central reward brain regions and dorsal raphe (DR) nuclei.
Serotonergic projections from the dorsal raphe innervate the VTA and the NAc (Conrad et al., 1974; Li et al., 1993; Moore et al., 1978; Parent et al., 1981). Importantly, serotonergic projections from the dorsal raphe nucleus to the NAc are suggested to be involved with alcohol reinforcement in alcohol-preferring rats (McBride et al., 1989). These serotonergic projections make synaptic contact with pre- and postsynaptic sites in the NAc (Van Bockstaele et al., 1996; Van Bockstaele and Pickel, 1993). At the ultrastructural level, serotonergic terminals are often found adjacent to GABA-containing neurons in the NAc (Van Bockstaele et al., 1996) and these serotonergic terminals may regulate, or be regulated by, presynaptic inputs with excitatory axons forming asymmetric junctions and also by catecholaminergic afferents (Van Bockstaele and Pickel, 1993). Thus, dendrites of medium spiny neurons which are GABAergic form synaptic contacts with 5-HT, dopamine, and glutamate terminals (Gonzales and Weiss, 1998; Muramatsu et al., 1998; Van Bockstaele et al., 1996; Van Bockstaele and Pickel, 1993; Weiss et al., 1993; Yan, 1999; Yan and Yan, 2001; Yan et al., 2004). Both 5-HT1B receptor mRNA and its protein are expressed in the NAc (Bruinvels et al., 1993; Bruinvels et al., 1994; Mengod et al., 1990) suggesting that 5-HT1B auto-receptors located at the terminals of dorsal raphe serotonergic neurons in the NAc and VTA, and hetero-receptors located at the terminal of medium spiny neurons in the VTA, may play a significant role in drug reinforcement, including alcohol.
It is important to note that there are some differences in sensitivity between presynaptic 5-HT1B auto- and hetero-receptors for a particular agonist (Rossetti and Carboni, 1995). This study investigated the effects of selective 5-HT1B receptor agonists (5-HT and CP93,129) on the K+-evoked overflow of [3H]5-HT, [3H]dopamine and [3H]acetylcholine in the striatum or hippocampus. The 5-HT1B receptor agonists show higher potencies in the inhibition of the K+-evoked overflow of [3H]5-HT from striatal synaptosomes as compared to the inhibition of the K+-evoked overflow of [3H]dopamine in the striatum or [3H]acetylcholine in the hippocampus (Sarhan and Fillion, 1999). These findings demonstrated that 5-HT1B auto- and hetero-receptors located at the terminals of dopaminergic and cholinergic neurons show differences in sensitivity for a 5-HT1B receptor agonist or even antagonist. There appear to be no structural differences between 5-HT1B auto- and hetero-receptors but differences in sensitivity to a selected agonist or antagonist might occur because these receptors might be coupled differently to their effectors. This assumption is based on findings from investigations of D2 receptor function. Thus, studies have shown that D2 receptor agonists display different efficacies in the activation of D2 auto- versus hetero-receptors (Martin et al., 1984; Rosenzweig-Lipson and Barrett, 1995; Rosenzweig-Lipson and Bergman, 1993; Skirboll et al., 1979). In general, D2 receptor agonists displayed dose-dependent effects on the activation of auto- versus hetero-receptors. It was suggested that low doses of D2 receptor agonists may activate only D2 auto-receptors, leading to the inhibition of dopamine release (Rosenzweig-Lipson and Barrett, 1995). This phenomenon may apply to 5-HT1B auto-receptors as well. Studies are needed to investigate this possibility.
Interactions between 5-HT1B receptors and the dopaminergic system involving GABAergic activity in the modulation of alcohol-drinking behavior
The excitatory action of ethanol on dopaminergic neurons in the VTA is potentiated by 5-HT (Rossetti and Carboni, 1995). The 5-HT1B receptors play a key role in the activation of dopaminergic neurons in the VTA. This excitatory effect is further influenced by GABAergic projections (NAc-VTA feedback loop). For example, activation of 5-HT1B receptors in the VTA indirectly disinhibited dopaminergic neurons which reduced alcohol-associated reward and subsequently reduced alcohol intake (Yan et al., 2004; Yan et al., 2005). Thus, the interactions between these reciprocal pathways in the NAc and the VTA regulate alcohol consumption.
Numerous pharmacological studies have demonstrated the role of dopamine in alcohol-drinking behavior using rat models. It has been shown that i.p. injections of D1 and D2 receptor agonists decreased alcohol intake (McBride et al., 1990; Weiss et al., 1990). In addition, systemic administration of D1 and D2, as well as D3, receptor agonists dose-dependently decreased oral self-administration of ethanol (Cohen et al., 1998, 1999). Local injections of D2 receptor antagonists (sulpiride or quinpirole) in the VTA increased alcohol intake (Melendez et al., 2005; Nowak et al., 2000). Furthermore, local injections of sulpiride in the NAc increased ethanol intake in a dose-dependent manner (Levy et al., 1991). Note that any variances in alcohol intake between studies may be due to the specific type of targeted dopamine receptor, the doses of dopamine receptor agonists and antagonists, or differences
The 5-HT1B receptors located at the terminals of NAc GABAergic projections to the VTA are found to be involved in the modulation of mesolimbic dopamine activity (Yan and Yan, 2001; Yan et al., 2004). Pharmacological studies revealed that activation of 5-HT1B receptors in the VTA lead to increased dopamine release in both the VTA and NAc (Yan et al., 2005; Yan et al., 2004). Thus, it is suggested here that activating 5-HT1B receptors in the VTA may potentiate the rewarding effects of ethanol, whereas deactivating 5-HT1B receptors may attenuate the effects of ethanol. This regulation of dopamine release by 5-HT1B receptors in the VTA may result from GABAergic activity. Studies suggest that 5-HT1B receptors indirectly affect the release of dopamine in the VTA and the NAc through GABA release (O’Dell and Parsons, 2004; Parsons and Justice, 1993a; Parsons and Justice, 1993b; Yan and Yan, 2001; Yan et al., 2004). In addition, serotonergic neurons in the dorsal raphe nucleus regulate VTA dopaminergic projections to the NAc (Yoshimoto and McBride, 1992). This suggests that 5-HT affects ethanol-drinking behavior via 5-HT1B auto-receptors located on these dopaminergic projections in the brain regions of drug reward (Czachowski, 2005). In alcohol-preferring rats, imbalances in 5-HT availability may affect the activity of the dopaminergic system in the NAc and VTA. From a therapeutic standpoint, if 5-HT1B auto-receptors are key players in the regulation of dopamine activity to modulate alcohol intake, therefore, targeting the sensitivity of these receptors would be an important step toward developing a treatment for alcohol dependence. As discussed in this review, the doses of 5-HT1B receptor agonists or antagonists administered are important in the apparent differential effects of 5-HT1B auto- or hetero-receptors.
Interactions between the serotonergic and the glutamatergic systems in the modulation of alcohol-drinking behavior
Glutamatergic projections innervating the NAc and the VTA bear 5-HT1B hetero-receptors at their terminals and are critical modulators of alcohol intake. These glutamatergic projections modulate alcohol-drinking behavior in response to inputs from 5-HT1B auto-receptors on dorsal raphe 5-HT neurons. Changes in glutamatergic neurotransmission affect many aspects of neuroplasticity associated with alcohol/drug dependence. Neuroadaptations in both presynaptic and postsynaptic aspects of mesocorticolimbic glutamatergic neurotransmission indicate that the regulation of protein expression is involved in alcohol dependence. Chronic alcohol exposure increased the density of glutamate and N-Methyl-D-aspartic acid (NMDA) antagonist (MK801) binding sites in the rat and mouse brain regions, including the hippocampus and cerebellum (Chen et al., 1997; Grant et al., 1990; Sanna et al., 1993; Snell et al., 1996). Also, cortical glutamatergic projections bearing 5-HT1B heteroreceptors that target the NAc region (Muramatsu et al., 1998) may play a role in the regulation of alcohol-drinking behavior. The exact relationship between the glutamatergic system and 5-HT1B receptors in the central reward regions is not fully defined. Although the glutamatergic projections bearing 5-HT1B hetero-receptors have not been fully delineated, afferent regions in the NAc, VTA, neocortex, and thalamus display high levels of 5-HT1B receptor mRNA (Bruinvels et al., 1994). This may provide a source for 5-HT1B receptors on glutamatergic terminals in the mesocorticolimbic system. If these 5-HT1B hetero-receptors play a role in the regulation of glutamate release (Muramatsu et al., 1998), the aforementioned glutamatergic projections may regulate the activity of associated GABAergic and dopaminergic neurons (Bonci and Malenka, 1999; Johnson et al., 1992b; Sesack et al., 1989). It is noteworthy that 5-HT inhibits synaptic glutamate release in NAc neurons through presynaptic 5-HT1B receptors (Muramatsu et al., 1998). The glutamatergic inputs to the NAc and VTA that bear 5-HT1B hetero-receptors (Figure 1) may be regulated as a consequence of changes in 5-HT release availability. The projections from the dorsal raphe nuclei are considered to be a potential source for 5-HT release in the synaptic clefts (for review see Sari, 2004) (Figure 1) which would subsequently control dopaminergic, glutamatergic and GABAergic release in the control of alcohol-drinking behavior. Therefore, both 5-HT1B auto- and hetero-receptors might be involved in the regulation of glutamate neurotransmission, which leads to the modulation of alcohol intake.
Conclusion
We conclude here that 5-HT1B receptors are involved in the release of at least four neurotransmitters (GABA, dopamine, glutamate and 5-HT) in the central reward brain regions and dorsal raphe nuclei. Pharmacological studies suggested that activation of 5-HT1B receptors in these regions inhibits the release of these neurotransmitters and, by extension, regulates alcohol-drinking behavior. For example, activation of 5-HT1B receptors in VTA can induce inhibition of GABA release, which can consequently lead to increases in dopamine release in both VTA and NAc. This regulatory mechanism would mediate increased alcohol intake. Alternatively, activation of 5-HT1B receptors in the NAc induced decrease in alcohol intake which could be mediated through inhibition of glutamate release in the cortico-accumbal glutamatergic projections. Consequently, this can lead to a reduction in dopamine release in the NAc.
It is suggested here that 5-HT1B auto- or hetero-receptors expressed on serotonergic, glutamatergic, or GABAergic neurons in the central reward regions regulate alcohol-drinking behavior in response to the inhibition of the release of neurotransmitters. The natural 5-HT may have different potencies for 5-HT1B receptors that can lead primarily to the activation of 5-HT release: however, the activation of other neurotransmitters such as glutamate and GABA may occur when there is excessive release of 5-HT. Importantly, differences in sensitivity caused by 5-HT1B auto- or hetero-receptors for the natural agonist 5-HT or a selective agonist or antagonist may lead to the reduction of alcohol intake. Pharmacological manipulations for the determination of lower or higher doses of selective 5-HT1B receptor agonists or antagonists are important for determining doses in treatments for alcohol dependence.
Footnotes
Acknowledgements
The author would like to thank the National Institutes of Health-National Institute on Alcohol Abuse and Alcoholism for their support (R01 AA019458, YS). The author would like also to thank Charisse Montgomery for editing this review article.
Funding
This research was supported by the National Institutes of Health.
Conflicts of interest
The author declares that there is no conflict of interest.