
Abstract
Although the role of the median raphe nucleus (MRN) in the regulation of anxiety has received less attention than that of the dorsal raphe nucleus (DRN) there is substantial evidence supporting this function. Reported results with different animal models of anxiety in rats show that whereas inactivation of serotonergic neurons in the MRN causes anxiolysis, the stimulation of the same neurons is anxiogenic. In particular, studies using the elevated T-maze comparing serotonergic interventions in the MRN and in the DRN indicate that the former affect only the inhibitory avoidance task, which has been related to generalized anxiety. In contrast, similar operations in the DRN change both the inhibitory avoidance and the one-way escape task, the latter being representative of panic disorder. Simultaneous injections of 5-HT-acting drugs in the MRN and in the dorsal hippocampus (DH) suggest that the MRN-DH pathway mediates the regulatory function of the MRN in anxiety. Overall, the results discussed in this review point to a relevant role of the MRN in the regulation of anxiety, but not panic, through the 5-HT pathway that innervates the DH.
Introduction
The median raphe nucleus (MRN) has a dense population of neurons containing serotonin (5-HT) that project to forebrain structures, among which is the dorsal hippocampus (DH); in contrast, the dorsal raphe nucleus (DRN) sends serotonergic nerve fibers mainly to the amygdala and ventral hippocampus, in addition to other important emotion-associated areas such as the midbrain periaqueductal gray matter, striatum and prefrontal cortex (Azmitia and Segal, 1978; Hale and Lowry, 2011; Lechin et al., 2006; Lowry, 2002; McKenna and Vertes, 2001; Ögren et al., 2008; Vertes et al., 1999).
The role of the DRN in fear and anxiety is well established, and has been discussed in several review articles (Graeff and Zangrossi, 2010; Hale and Lowry, 2011; Hale et al., 2012; Lowry et al., 2008; Waselus et al., 2011). However the participation of the MRN and its serotonergic projection to the DH in anxiety regulation has received less attention. Yet, the MRN-DH pathway is central to the hypothesis that the “behavioral inhibition system” represented by the septum-hippocampus is a key neural substrate of anxiety (Gray, 1982, 1987; Gray and McNaughton, 2000).
The original behavior inhibition system theoretical construct assumes that anxiety is due to the activation of an operator by signs of punishment or frustration or by innate fear stimuli, resulting in inhibition of ongoing behavior, increased vigilance and attention directed to potential threats. Its anatomical substrate would be the septum, the hippocampus and the serotonergic and noradrenergic innervation coming from brain stem nuclei (Gray, 1982). Later, it was supposed that the hippocampus would work as a comparator between expected and actual environmental stimuli, and that a mismatch between the two representations would activate the behavior inhibition system (Gray, 1987). In a further development of the idea, the septum-hippocampus became a detector of conflict between approach and avoidance tendencies that biases the conflict resolution towards avoidance. The simultaneous activation of the opposed approach and avoidance trends would characterize anxiety, whereas the fear emotion would engage only the escape-avoidance defence system, without any motivational conflict (Gray and McNaughton, 2000).
The aim of the present article is to review the experimental evidence implicating the MRN in anxiety. For comparison, similar studies on the DRN will also be covered. It should be noted, however, that the far most of the existing behavioral studies with the DRN, as presented below, has been performed with drug injections or lesions aimed at the middle of this nucleus, without taking into account changes in its various subfields. This may be an important drawback since evidence gathered in the last years strongly supports the view that the DRN is not a homogenous structure, but an aggregate of different subpopulations of 5-HT and non-5-HT neurons that are morphologically and functionally distinct (Calizo et al., 2011; Clark et al., 2006; Hale and Lowry, 2011). In contrast, the existence of distinctive sub-areas within the MRN has not been widely acknowledged in the literature. As pointed out by Clark and colleagues (2006), besides the lack of clear anatomical demarcation of subregions within the MRN, this raphe nucleus displays, for instance, less heterogeneity of gene expression (e.g. for tryptophan hydroxylase-2, the serotonin transporter, 5-HT1A and 5-HT1B receptors) across its rostral-caudal extend than for the DRN.
Although critical to the comprehension of MRN and DRN functionality, a more detailed description of their afferents and efferent connections and also, the regulatory interaction between these nuclei are beyond the scope of this review and have been extensively discussed in recent reviews (Calizo et al., 2011; Hale and Lowry, 2011; Hensler, 2006; Paul and Lowry, 2013; Vasudeva et al., 2011).
Median raphe nucleus inactivation
The effects of MRN inactivation in behavioral tests assessing defensive reactions are summarized in Table 1.
Effects of median raphe nucleus (MRN) inactivation or activation on anxiety-related defensive responses in rats.
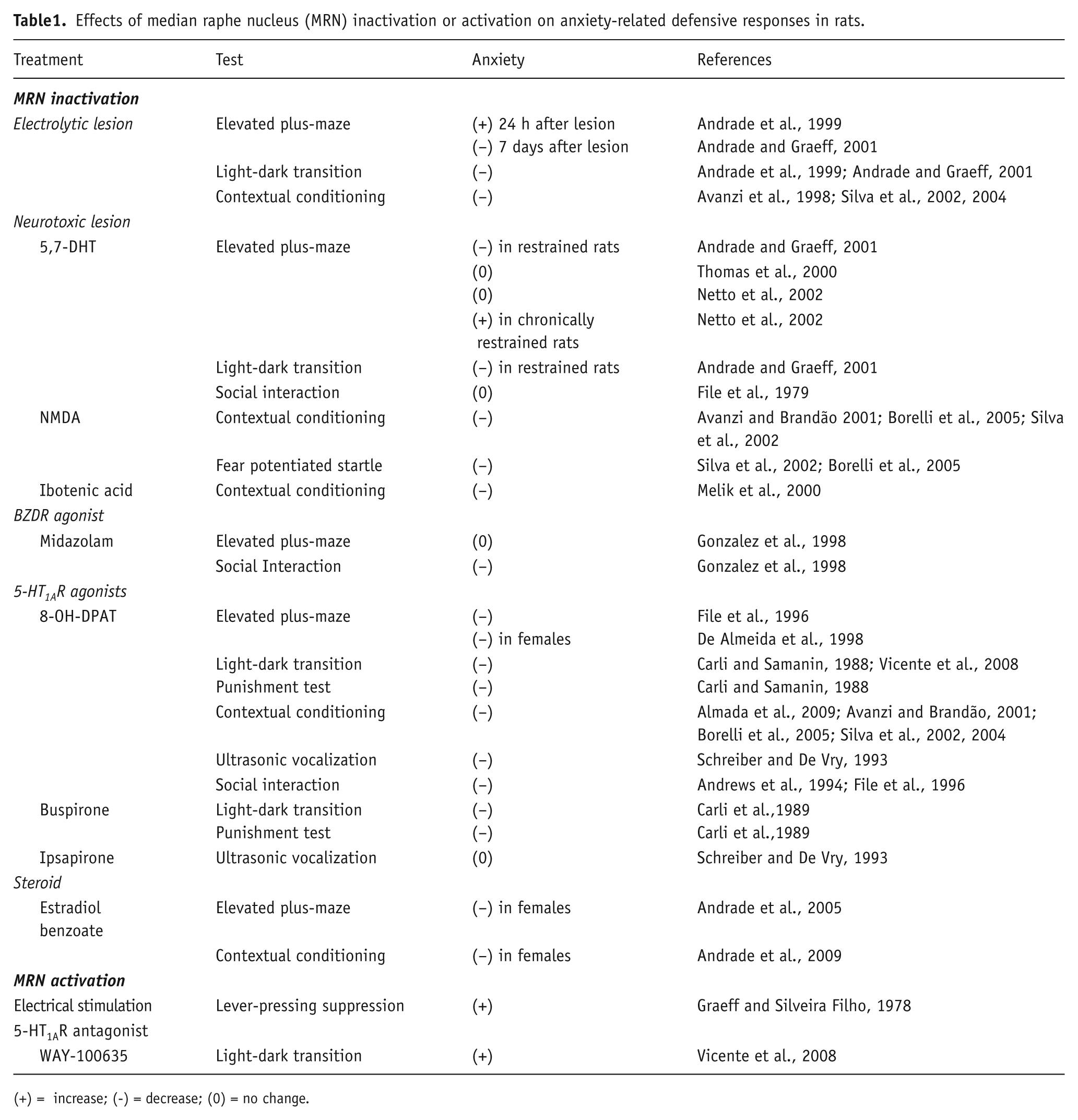
(+) = increase; (-) = decrease; (0) = no change.
Electrolytic lesion of the MRN in rats has been reported to consistently decrease anxiety indices in different animal models of anxiety, such as the elevated plus-maze (Andrade and Graeff, 2001), the light-dark transition test (Andrade et al., 1999; Andrade and Graeff, 2001) and contextual conditioning (Avanzi et al., 1998; Silva et al., 2002, 2004). Anxiolytic-like effects were also reported following neurotoxic damage of this nucleus with excitatory amino acids. For instance, either N-methyl-D-aspartate (NMDA) (Avanzi and Brandão, 2001; Borelli et al., 2005; Silva et al., 2002) or ibotenic acid-induced lesions (Melik et al., 2000) in this area disrupt contextual conditioning. The former type of lesion also decreases fear-potentiated startle, specifically when performed before the training session (Silva et al., 2002).
As observed with these lesion studies, local facilitation of GABAA-mediated inhibitory neurotransmission in the MRN, through microinjection of benzodiazepine (BZD) receptor agonists, also causes anxiolysis, but this seems to be dependent on the experimental model used. Thus, at the same doses, intra-MRN administration of midazolam was anxiolytic in the social interaction test, but ineffective in the elevated plus maze (Gonzalez et al., 1998).
It is worth noting that different neuronal populations within the MRN are affected by these lesions or by the facilitation of GABA-mediated neurotransmission. In the case of electrolytic lesions, besides damage to different cell types (serotonergic and non-serotonergic neurons and glial cells), blood vessels and passing neuronal fibers are also affected (Asin and Fibiger, 1983), and all these changes may ultimately influence the behavioral outcome. In order to circumvent these problems, some studies have employed the neurotoxin 5,7-dihydroxytryptamine (5,7-DHT) to cause selective damage to 5-HT neurons in the MRN. In line with the results found with unspecific manipulations, 5,7-DHT-lesioned rats show decreased anxiety in the light-dark transition model (Andrade and Graeff, 2001; Andrade et al., 1999) and tend to be less anxious in the social interaction test (File et al., 1979). However, the results in the elevated plus maze are inconsistent: a decrease (Andrade and Graeff, 2001), no effect (Thomas et al., 2000) and an increase (Netto et al., 2002) in anxiety were reported. It should be pointed out that the anxiogenic effect in the elevated plus-maze was only observed in rats that were chronically restrained before the test.
Selective inactivation of MRN 5-HT neurons can also be achieved by stimulating autoinhibitory 5-HT1A receptors located at the soma and dendrites of these cells, which results in inhibition of neuronal firing and reduction of 5-HT release in innervated territories (Bosker et al., 1996; Dudley et al., 1999; Hajós et al., 1995; Kreiss and Lucki, 1994). The results so far reported consistently show that intra-MRN administration of different doses of the 5-HT1A receptor agonist 8-OH-DPAT decreases anxiety in the elevated plus-maze (De Almeida et al., 1998; File et al., 1996), light-dark transition (Carli and Samanin, 1988; Vicente et al., 2008), contextual conditioning (Almada et al., 2009; Avanzi and Brandão, 2001; Borelli et al., 2005; Silva et al., 2002, 2004), ultrasonic vocalization (Schreiber and De Vry, 1993), social interaction (Andrews et al., 1994; File et al., 1996) and punishment (Carli and Samanin, 1988) tests.
Anxiolysis has also been reported after intra-MRN injection of buspirone, a partial 5-HT1A receptor agonist, in the light-dark transition and in punishment tests (Carli et al., 1989), but not with ipsapirone, another partial 5-HT1A agonist in the ultrasonic vocalization model (Schreiber and De Vry, 1993).
Although more indirectly, the reported results showing that intra-MRN injection of estradiol benzoate cause an anxiolytic effect in the elevated plus maze and contextual conditioning tests, which is prevented by local 5-HT1A receptor blockade (Andrade et al., 2005, 2009) are consistent with the hypothesis that inhibition of MRN 5-HT neurons decreases anxiety.
MRN activation
The effects of experimental manipulations that result in activation of the MRN are summarized in Table 1.
The results of an early study by Graeff and Silveira Filho (1978) showed that electrical stimulation of the MRN in rats lever-pressing under a variable-interval schedule of water presentation suppressed ongoing responding in the same way as the stimulus paired with foot-shock in the Estes-Skinner conditioned suppression paradigm. In addition, the rats showed behavioral and neurovegetative changes characteristic of the conditioned emotional response, such as freezing, exophthalmos, piloerection, defecation and micturition. Furthermore, the anxiogenic-like effects of MRN electrical stimulation were reduced by administration of the 5-HT synthesis inhibitor p-chlorophenylalanine, implicating 5-HT in their mediation. A subsequent study (Graeff et al., 1980), not listed in Table 1, showed that MRN electrical stimulation induced theta electrical activity in the DH, a phenomenon that has been related to anxiety (McNaughton et al., 2007). However, this electrophysiological effect does not seem to be mediated by 5-HT because stimulation of MRN serotonergic neurons has been shown to desynchronize hippocampal theta (Jackson et al., 2008; Nitz and McNaughton, 1999; Vertes, 1981).
Activation of the MRN-hippocampal pathway may be achieved by acute (diazepam withdrawal) or chronic (14-day social separation) stress, since it was shown that both procedures selectively increased 5-hydroxytryptamine turnover in the hippocampus and median raphe nucleus, and had an anxiogenic effect in the elevated plus-maze. Chronic treatment with imipramine during the 14 days of separation prevented the behavioral and neurochemical changes caused by social separation. Therefore, the increase in anxiety determined by both acute and chronic stress seems to involve the activation of the MRN-hippocampal 5-HT pathway (Dos Santos et al., 2010). As this study did not investigate the consequences of these stressors on other raphe nuclei, it cannot be excluded that activation of 5-HT neurons within the DRN, for instance, may have also contributed to the observed enhancement of anxiety.
An anxiogenic effect was also observed after intra-MRN injection of the 5-HT1A receptor antagonist WAY-100635 in rats tested in the light-dark transition test (Vicente et al., 2008). It was shown that WAY-100635, by counteracting the tonic inhibitory effect of 5-HT on the firing rate of MRN neurons, increases 5-HT release in terminal areas (Dudley et al., 1998). This stimulatory effect on cell membrane activity is selective to 5-HT neurons, since in the MRN, though not in all subdivisions of the DRN, 5-HT1A receptors are located only in serotonergic neurons (Beck et al., 2004; Calizo et al., 2011; Kirby et al., 2003).
In contrast with the aforementioned study in rats, investigations in mice showed that intra-MRN administration of WAY-100635 is anxiolytic in the elevated plus-maze (Canto-de-Souza et al., 2002; Nunes-de-Souza, 2011). The reasons for this discrepant result can be manifold. But, one intriguing possibility is that the 5-HT system of these two species may be composed of subpopulations of serotonergic neurons with qualitatively and/or quantitatively different capacity for adaptation. For instance, a recent study reported that a subpopulation of 5-HT neurons of the DRN and MRN of mice does not express the somatodendritic 5-HT1A autoreceptor (Kiyasova et al., 2013), which is shown to regulate neuronal firing of raphe neurons by a negative feedback mechanism (Bosker et al., 1996; Dudley et al., 1999; Hajós et al., 1995; Kreiss and Lucki, 1994). It is still unknown whether rats have the same population of cells.
It is noteworthy that only two studies were performed to investigate the role played by MRN 5-HT neurons in the regulation of defensive behaviors in mice (Canto-de-Souza et al., 2002; Nunes-de-Souza et al., 2011) and both employed one single experimental model, the elevated plus-maze. Therefore, further analyses are required in order to verify the generality of these findings.
MRN and defensive behaviors measured by the elevated T-maze (ETM)
Overall, most of the studies performed in rats show that whereas either selective or nonselective inactivation of 5-HT neurons in the MRN causes anxiolysis, the stimulation of this nucleus is anxiogenic. The following studies with the elevated T-maze, which are summarized in Table 2, fully support this conclusion and go beyond by showing that 5-HT neurons in this nucleus may be primarily involved in the regulation of defensive behaviors associated with specific anxiety pathology, namely generalized anxiety disorder.
Effects caused by the inactivation or activation of the median raphe nucleus (MRN) or dorsal raphe nucleus (DRN) in rats tested in the elevated T-maze (ETM).
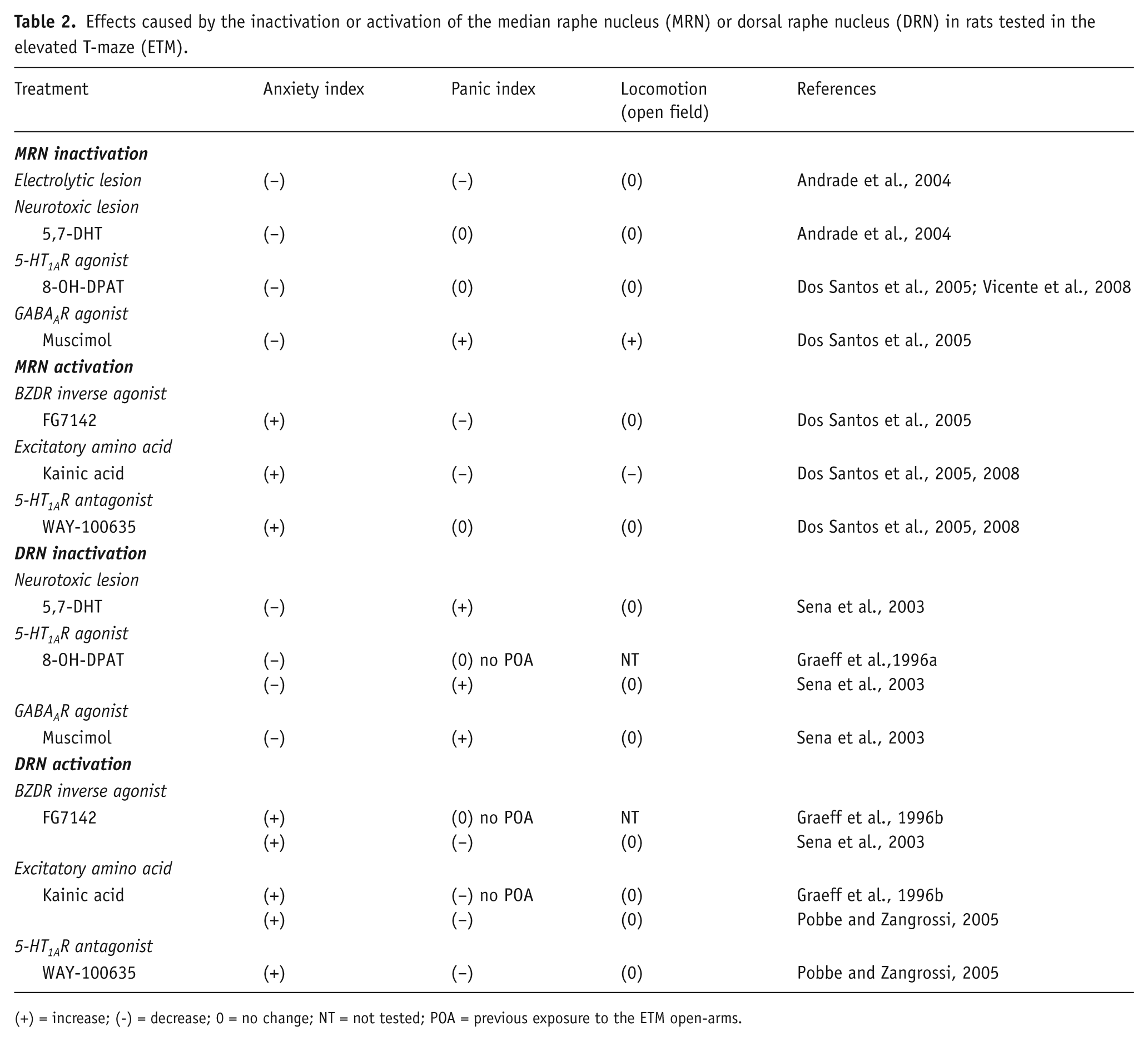
(+) = increase; (-) = decrease; 0 = no change; NT = not tested; POA = previous exposure to the ETM open-arms.
The ETM is derived from the elevated plus-maze (Handley and Mithani, 1984; Pellow et al., 1985) and is made of one arm enclosed by walls, perpendicular to two open arms of equal dimensions, all suspended 50 cm from the floor (Graeff et al., 1993; Viana et al., 1994). The same rat performs an inhibitory avoidance task followed by a one-way escape task. For the first task, the animal is placed at the end of the enclosed arm and the time to withdraw from this arm with the four paws is measured three successive times. The increase in withdrawal latency along the three trials is taken as a measure of inhibitory avoidance acquisition, because the animal is suppressing exploratory activity to avoid exposure to the aversive open arms. For the escape task, the rat is placed at the extremity of one open arm and the time taken to withdraw from this arm with the four paws while going toward the protected arm is measured for three consecutive times. Contrary to what happens in the enclosed arm, the latency to leave the open arm usually does not change along successive trials (Zangrossi and Graeff, 1997).
As discussed in several review articles (Graeff et al., 1998; Graeff and Zangrossi, 2002; McNaughton and Zangrossi, 2008; Pinheiro et al., 2007; Poltronieri et al., 2003), the results of different pharmacological treatments indicate that the avoidance task models generalized anxiety while the escape task represents panic disorder. For instance, clinically effective drugs in patients with generalized anxiety but not panic disorder, such as diazepam and buspirone were shown to impair inhibitory avoidance acquisition without interfering with the escape response, indicating a selective anxiolytic effect (Graeff et al., 1993). Tricyclic antidepressants and selective serotonin re-uptake inhibitors (SSRIs), on the other hand, impair escape response after chronic, but not acute treatment, in line with their effect on panic disorder patients demonstrated in clinical settings (Pinheiro et al., 2007; Poltronieri et al., 2003; Teixeira et al., 2000).
The resemblance of the motor performance in both ETM tasks serves as a control for non-specific drug/lesion effects on motor activity when the latencies to leave the enclosed arm and from the open arm are changed to opposite directions by the treatment used. However, whenever the latencies are similarly increased or decreased, there is need for independent assessment of motor drug effects. This has been achieved by measuring motor activity inside a square arena immediately after testing in the ETM.
Regarding the MRN, it was shown that either its electrolytic- or 5,7-DHT-induced lesions impairs avoidance acquisition indicating an anxiolytic effect. However, only the former procedure interferes with escape expression, impairing it, a panicolytic-like effect (Andrade et al., 2004). Change in both defensive responses was also found after the intra-MRN injection of the GABAA receptor agonist muscimol (Dos Santos et al., 2005), a drug that nonselectively inhibits MRN neurons (Forchetti and Meek, 1981; Judge et al., 2006; Tao and Auerbach, 2000). But in this case, the anxiolytic and panicogenic-like effect observed was accompanied by an increase in locomotion in the open-field, suggesting that a non-specific motor stimulatory effect may have contributed for these results. As reported after 5,7-DHT administration in the MRN, local microinjection of 8-OH-DPAT was anxiolytic upon the inhibitory avoidance task, without affecting escape performance (Dos Santos et al., 2005; Vicente et al., 2008). Importantly, neither of these procedures interfered with locomotion in the square arena.
Three studies explored the behavioral consequences of nonselective chemical stimulation of the MRN in rats performing in the ETM. In two of them (Dos Santos et al., 2005, 2008), neuronal stimulation by locally microinjecting the excitatory amino acid kainic acid facilitated avoidance acquisition and inhibited escape, an effect in direct opposition to that observed after nonselective inhibition of this nucleus after either electrolytic lesions or local microinjection of muscimol. Nevertheless, in agreement with previously reported results (Wirtshafter and McWilliams, 1987) kainic acid also decreased locomotion in the open-field, what makes the interpretation of the behavioral effects in the ETM less clear.
In the same way, local microinjection of inverse benzodiazepine receptor (BZDR) agonist FG-7142 into the MRN, which suppresses GABAergic inhibition and, consequently, causes widespread neuronal overstimulation (Atack et al., 2005; Kurumaji and Nishikawa, 2012; Thiébot et al., 1988), resulted in enhancement of avoidance and impairment of escape (Dos Santos et al., 2005). However, compared with kainic acid, the effects of FG-7142 in the ETM seem to be more genuine, as this drug treatment did not significantly affect locomotion in the open-field.
As mentioned before, intra-MRN injection of the 5-HT1A receptor antagonist WAY-100635 causes a disinhibitory effect on the activity of 5-HT neurons. Two studies were carried out in the ETM using local microinjection of this drug (Dos Santos et al., 2005, 2008), and both showed a marked facilitation of avoidance acquisition without any interference with escape performance or locomotion.
To verify whether the anxiogenic effect of intra-MRN WAY-100635 was enacted through the MRN-DH pathway, in the study of Dos Santos and co-workers (2008) an additional experiment was performed in which one group of rats was given WAY-100635 into the hippocampus 10 min before the intra-MRN administration of the same drug. Another group received intra-DH saline injection as pretreatment, while a control group was injected with saline in both the DH and the MRN. The results showed that after intra-DH saline pretreatment WAY-100635 facilitated avoidance acquisition without interfering with escape expression, as previously reported (Dos Santos et al., 2005). Pre-administration of WAY-100635 into the DH fully counteracted the anxiogenic effect of WAY-100635. These results clearly indicate that activation of the MRN-DH serotonergic pathway enhances anxiety by stimulating post-synaptic 5-HT1A receptors localized in the DH, and that this system does not control panic. Consistent with this interpretation, Dos Santos and colleagues (2008) also showed that intra-HD injection of 8-OH-DPAT facilitated inhibitory avoidance acquisition, without changing escape expression, exactly as observed after injection of WAY-100635 in the MRN.
Serotonergic interventions in the DRN
In contrast to serotonergic manipulations in the MRN, that affected only inhibitory avoidance acquisition in the ETM, interventions in the 5-HT neurotransmission in the DRN change escape performance as well (see a summary of these results in Table 2). Thus, selective lesion of DRN 5-HT neurons with 5,7-DHT impaired avoidance and simultaneously facilitated escape, that is, had both anxiolytic and panicogenic effects (Sena et al., 2003). Inhibiting 5-HT neuronal activity with local administration of 8-OH-DPAT inside the DRN also had both anxiolytic and panicogenic effects in the ETM (Sena et al., 2003). Nevertheless, previously reported results showed only avoidance impairment (Graeff et al., 1996a). This difference is likely to be due to the absence of pre-exposure to one of the open arms the day before the test, a procedure that renders the escape task more sensitive to drugs (Teixeira et al., 2000).
Consistent with the effect observed in the ETM, intra-DRN injection of 8-OH-DPAT had anxiolytic effect in other animal models of anxiety, including the elevated plus-maze (File and Gonzalez, 1996), light-dark transition test (Sena et al., 2003), ultrasonic vocalization (Remy et al., 1996; Schreiber and De Vry, 1993), social interaction (Higgins et al., 1988; Hogg et al., 1994; Merali et al., 2006; Picazo et al., 1995) and the Geller-Seifter conflict test (Cervo et al., 2000; Schreiber and De Vry, 1993).
Results with partial 5-HT1A receptor agonists have been equally consistent. Thus, an anxiolytic effect was obtained in the social interaction test following intra-DRN injection of gepirone (Higgins et al., 1992), buspirone (Higgins et al., 1988) and ipsapirone (Higgins et al., 1988; Picazo et al., 1995). The two latter drugs also decreased anxiety in the ultrasonic vocalization model (Schreiber and De Vry, 1993). However, local microinjection of buspirone was without effect in rats tested in the light-dark transition test (Carli et al., 1989).
In the opposite direction, enhancing the functioning of DRN serotonergic activity by locally injecting the 5-HT1A receptor blocker WAY-100635 facilitated inhibitory avoidance acquisition in the ETM and, at the same time, impaired escape performance, characterizing both an anxiogenic and a panicolytic effect (Pobbe and Zangrossi, 2005). The same effect was also reported after intra-DRN injection of kainic acid (Graeff et al., 1996b; Pobbe and Zangrossi, 2005) and FG-7142 (Sena et al., 2003). An increase in 5-HT release in the dorsal column of the periaqueductal gray matter (DPAG) seems to underlie the effects of WAY-100635 and kainic acid administration in the DRN, because blockage of either 5-HT1A or 5-HT2A/2C receptors in DPAG was shown to counteract the panicolytic effect caused by these DRN manipulations, while antagonism of the latter receptors abolished the facilitation of inhibitory avoidance acquisition (Pobbe and Zangrossi, 2005).
Interestingly, as observed in the MRN, the effects of WAY-100635 administration in the DRN seem to be different between rats and mice. In the latter species, local microinjection of this antagonist was ineffective both in the elevated plus-maze (Canto-de-Souza et al., 2002) and in the mouse defense test battery (Pobbe et al., 2011).
Morphological evidence
The difference between the effects in the ETM of serotonergic manipulations in the MRN and in the DRN is consistent with the differential distribution of the 5-HT-containing fibers originating from these nuclei. Thus, the fibers coming from the MRN reach the DH, and only moderately the DPAG, whereas those stemming from the DRN reach the amygdala and the ventral hippocampus through the medial forebrain bundle, and the DPAG through the DRN-periventricular pathway (Azmitia and Segal, 1978, Vertes et al., 1999). According to the hypothesis proposed by Deakin and Graeff (1991) 5-HT would enhance anxiety through the forebrain projection and inhibit panic through the midbrain projection. The results reported above are consistent with this view. As to the MRN, the 5-HT input to the DH would enhance information processing in this structure resulting in more anxiety, according to the Gray’s ‘“behavioral inhibition system” theoretical model (Gray, 1982, 1987; Gray and McNaughton, 2000). Because the MRN nucleus does not appreciably innervate the DPAG, no effect on panic is expected. Again, the results obtained in the ETM support this prediction.
The role of the DH in anxiety has been overshadowed by its widely discussed participation in other behavioral processes such as declarative memory and spatial navigation (Bannerman et al., 2004; Bertoglio et al., 2006; Fanselow and Dong, 2010; McHugh et al., 2008). Nevertheless, the results reviewed above, obtained in several animal models of anxiety and in different laboratories, consistently support a function of the MRN-DH pathway in anxiety regulation.
A step forward in the understanding of participation of the DRN and the MRN in defensive emotions has been taken by a recent study in which the number of doubly immunostained cells for Fos protein and tryptophan hydroxylase, a marker of serotonergic neurons, was assessed within the rat DRN, MRN and PAG following inhibitory avoidance and escape performance in the ETM (Spiacci et al., 2012). Inhibitory avoidance, but not escape, significantly increased the number of Fos-expressing serotonergic neurons within the mid-caudal part of the dorsal subnucleus, caudal and interfascicular subnuclei of the DRN and in the MRN. Escape, on the other hand, caused a marked increase in the activity of non-5HT cells within the lateral wings of the DRN and the DPAG. These results strongly corroborate the view that different subsets of neurons in the DRN (Lowry, 2002; Lowry et al., 2008) are activated by anxiety- and panic- relevant stimuli/situations. Moreover, it strengthens the view that 5-HT neurons of the MRN are primarily involved in anxiety, instead of panic-regulation.
Conclusion
Overall, the results discussed in this review point to a relevant role of the MRN in the regulation of anxiety. This can be achieved through the 5-HT pathway that innervates the DH. In contrast, there is no evidence supporting a role of this 5-HT system in panic regulation, although nonserotonergic neurons localized in the MRN may be involved in this function. Alternatively, the DRN seems to participate in both anxiety and panic. Interestingly, the regulation of anxiety by the DRN and by the MRN 5-HT pathway is enacted through different target structures – the amygdala, DPAG and ventral hippocampus in the case of the DRN, and the dorsal hippocampus in that of the MRN. Because these structures have different functional profiles, it is plausible that different aspects of anxiety are underpinned by each of them. In this regard, Gray and McNaughton (2000) have argued that the amygdala may underpin the neurovegetative arousal and affective components of anxiety, whereas the DH would regulate cognitive manifestations, such as worry. This suggestion is supported by the reported interaction between the DH and the prefrontal cortex underpinning fear learning and memory (Zelikowsky et al., 2013).
Footnotes
Acknowledgements
FGG is the recipient of a research fellowship from The National Council for Scientific and Technological Development (CNPq, Brazil) and the Fundação de Amparo ao Ensino, Pesquisa e Assistência do Hospital das Clínicas de Ribeirão Preto (FAEPA, No. 05/11) and HZJ is the recipient of a research fellowship from CNPq, Brazil.
Conflict of interest
The authors declare no conflict of interest.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.