
Abstract
The modulation of genes implicated in synaptic plasticity following administration of antipsychotic drugs has been instrumental in understanding their possible mode of action. Arc (Arg 3.1) is one such gene closely associated with changes in synaptic plasticity. In this study we have investigated the changes in expression of Arc protein following acute and chronic administration of a typical antipsychotic (haloperidol) and an atypical antipsychotic (clozapine) by means of immunohistochemistry compared to the prototypic gene marker c-Fos. In dorsal striatum haloperidol (1 mg/kg) significantly increased Arc expression following both acute and chronic (21 day) administration with evidence of modulation in induction after repeated dosing. No significant changes were observed following either acute or chronic administration of clozapine (20 mg/kg). In the nucleus accumbens shell both clozapine and haloperidol induced Arc expression following acute administration, again with evidence of modulation after chronic dosing. The pattern of induction of Arc expression following haloperidol and clozapine in both dorsal and ventral striatum was similar to that for c-Fos. In medial prefrontal and cingulate cortex, Arc expression was significantly decreased by clozapine but not haloperidol without any indication of modulation following chronic dosing, whereas no significant changes in c-Fos expression were observed with either drug. Since synaptic modulation mediated by Arc is associated with down-regulation of the AMPA glutamate receptor, this study suggests a mechanism whereby enhanced glutamate receptor efficacy in medial cortical areas may be a component of antipsychotic drug action.
Keywords
Introduction
The aetiology of schizophrenia remains unclear, but there is increasing evidence implicating neurodevelopmental dysfunction in the pathophysiology of schizophrenia and synaptic modulation as an essential mode of action of pharmacological treatment (Harrison and Weinberger, 2005; Snyder and Gao, 2013). Whilst blockade of dopamine D2 receptors is generally agreed to be the principal mode of action of both typical and atypical antipsychotic drugs, the need for long-term treatment to achieve lasting clinical benefits suggests that this mechanism is only the primary action leading to persistent neuronal modification (Molteni et al, 2009; de Bartolomeis and Tomasetti, 2012). In this respect several neuronal immediate-early genes have been investigated following antipsychotic drug administration, most extensively c-fos (Dragunow et al, 1990; Robertson et al, 1994) and more recently markers related to synaptic plasticity such as homer, norbin, NARP and arc (Robbins et al, 2008; de Bartolomeis et al, 2013). Arc (also known as Arg 3.1) is particularly involved in the development of activity-dependent synaptic plasticity as well as the maintenance of homeostatic scaling (Béïque et al, 2011; Shepherd and Bear, 2011; Nonaka et al, 2014) and is modulated by changes in neurotransmitter function mediated by various psychotropic drugs, including amphetamines (Beveridge et al, 2004: Horner et al, 2010), antidepressants (Pei et al, 2003; Serres et al, 2012) and antipsychotics (Nakahara et al, 2000; Iasevoli et al, 2010). To date, the studies of changes following antipsychotic drug administration have focused primarily on mRNA levels measured either by polymerase chain reaction (PCR) analysis (Robbins et al, 2008; Bruins Slot et al, 2009: Fumagalli et al, 2009) or in situ hybridisation (Iasevoli et al, 2010; de Bartolomeis et al, 2013). The current study has investigated changes in Arc protein measured by immunohistochemistry following both acute and chronic treatment of rats with the typical antipsychotic haloperidol and the atypical drug clozapine, in part to compare protein with mRNA expression in those situations previously investigated and in part to reveal novel changes in those brain regions not previously studied. The effects of these drugs on c-Fos expression has already been extensively studied and therefore served as a reference marker in this study. Given the relevance of Arc in mediating synaptic plasticity, the study has specifically focused on the regional changes in Arc expression following acute administration of typical compared with atypical antipsychotics and the modulation and persistence of changes of Arc expression following chronic compared with acute administration.
Methods and materials
Animals
Rats were housed in groups of three to five per cage under controlled conditions of temperature (21 ± 1°C) and lighting on a 12 hour light/dark cycle (lights on 7.00 am). Rats were given ad libitum access to food (B & K Universal Ltd., Aldbrough, Hull, or Harlan, Bicester, UK) and water. All experiments complied with the Home Office Guidance on the operation of the UK Animals (Scientific Procedures) Act 1986 under appropriate project and personal licences. Experiments were also performed according to guidelines set by the Ethical Review and Animal Welfare committee, De Montfort University and approved by the GlaxoSmithKline Procedures Review Panel.
Chemicals and drugs
All chemicals were supplied by Sigma, UK, or Fisher Scientific, UK, unless otherwise stated. Pentobarbitol (Euthatal) was purchased from Merial Animal Health, UK, haloperidol was purchased from Sigma-Aldrich, UK and clozapine was purchased from Tocris Cookson, UK. In the study, haloperidol was initially dissolved in 3% acetic acid then pH adjusted to 6.0 with sodium hydroxide. The same vehicle was used in the control samples and in the clozapine group.
Drug treatments
Adult male Sprague–Dawley rats (n=5–6 per group) weighing 260–300 g were randomly assigned to a treatment group and pretreated i.p. with either vehicle (V) (1 mL/kg), haloperidol (H) (1 mg/kg) or clozapine (C) (20 mg/kg) once daily for 21 days. The doses chosen correspond to those used in related previous studies (Nguyen et al, 1992; Robertson et al, 1994; Wan et al, 1995; Sebens et al, 1995; Robbins et al, 2008; Fumagali et al, 2009). The site of injection was varied to avoid local irritation. On day 22 a challenge dose of either vehicle, haloperidol or clozapine was administered i.p. such that seven groups were created corresponding to control (V-V), acute haloperidol (V-H), chronic haloperidol (H-H), chronic haloperidol with 24 h withdrawal (H-V), acute clozapine (V-C), chronic clozapine (C-C), and chronic clozapine with 24 h withdrawal (C-V).
Immunohistochemistry
Rats were anaesthetised i.p. with 100 mg/kg pentobarbital given 2 hours after the final drug administration then transcardially perfused with heparinised saline followed by 4% paraformaldehyde. Brains were removed, post-fixed in 4% paraformaldehyde for 48 hours at 4°C and cryoprotected in 30% sucrose for 48 hours then snap frozen in isopentane. Coronal frontal and striatal sections from each brain were cut at 40 μm at -20°C and collected and stored free floating in immunobuffer (0.1 M NaCl, 8.5 mM Na2HPO4, 6.3 mM NaH2PO4, 0.01 M Tris–HCl, 0.001 M Thimerosal, 0.3% Triton-X-100 (BDH, UK), pH 7.4) at 4°C.
Immunohistochemistry was performed using antibodies against Arc (mouse monoclonal IgG, Santa Cruz, USA) and c-Fos (rabbit polyclonal IgG, Oncogene, UK) based on previously published methods (Jennings et al, 2006; Southam et al, 2009). Briefly, free-floating sections were incubated in 1% hydrogen peroxide for 30 min, washed twice in immunobuffer and incubated in 0.5% bovine serum albumin for 30 min. Then they were washed twice in immunobuffer and blocked in 2% normal serum for 1 h. Sections were incubated with primary antibody diluted in 2% normal serum (c-Fos at 1:20,000 for 48 h; Arc at 1:500 for 24 h) at 4°C and washed with immunobuffer three times for 10 min. Sections were then incubated with biotinylated secondary antibody diluted in 2% normal serum for 2 h (either goat anti-rabbit at 1:200 or horse anti-mouse, rat adsorbed at 1:500 (both from Vector Laboratories, UK)). They were then washed three times in immunobuffer and antigens were visualised using the Vectastain Elite ABC kit (Vector Laboratories, UK) according to the manufacturer’s recommendations, followed by incubation with 3,3′-diaminobenzidine substrate (DAB) (Vector Laboratories, UK). Sections were rinsed for 3 × 10 min in phosphate-buffered saline then mounted onto SuperFrost Plus slides (BDH, UK), air-dried and coverslipped using DPX mountant.
Immunohistochemistry was performed on control sections in which the protocol was identical except for omission of the primary antibody. In both cases there was no immunostaining of control sections.
All the slides were randomly coded to ensure analysis was blind to treatment group. The slides were visualised under a light microscope (Leitz Diaplan, Germany) at ×20 objective and images were captured using a Photometrics CoolSNAP 10-bit monochrome digital camera mounted on a ×0.63 reducing lens and displayed on a PC using MCID Basic™ densitometry software (Interfocus Ltd, UK). A flat field correction was first established to eliminate artefact and enhance the image. Regions of interest (ROIs) were analysed by placing a calibrated digital sampling rectangle (500 × 500 μm or 500 × 300 μm depending on the specific region) over the image and counting the number of immunostained cells within the sampling rectangle. Any cells with more than 50% of their labelling outside the perimeter of the sampling rectangle were not counted. Cell counts were taken from relevant discrete brain regions at two coronal levels of the brain. Medial prefrontal cortex (MPFC) cell counts were taken from frontal cortex sections corresponding to Bregma 3.70 mm to 3.20 mm. Dorsolateral caudate (DLC), dorsomedial caudate (DMC), lateral septum (LS), nucleus accumbens core (NAcC) and shell (NAcS) and cingulate cortex (Cing) cell counts were taken from striatal sections corresponding to Bregma 1.20 mm to 0.70 mm (Paxinos and Watson, 2005) as indicated in Figure 1.
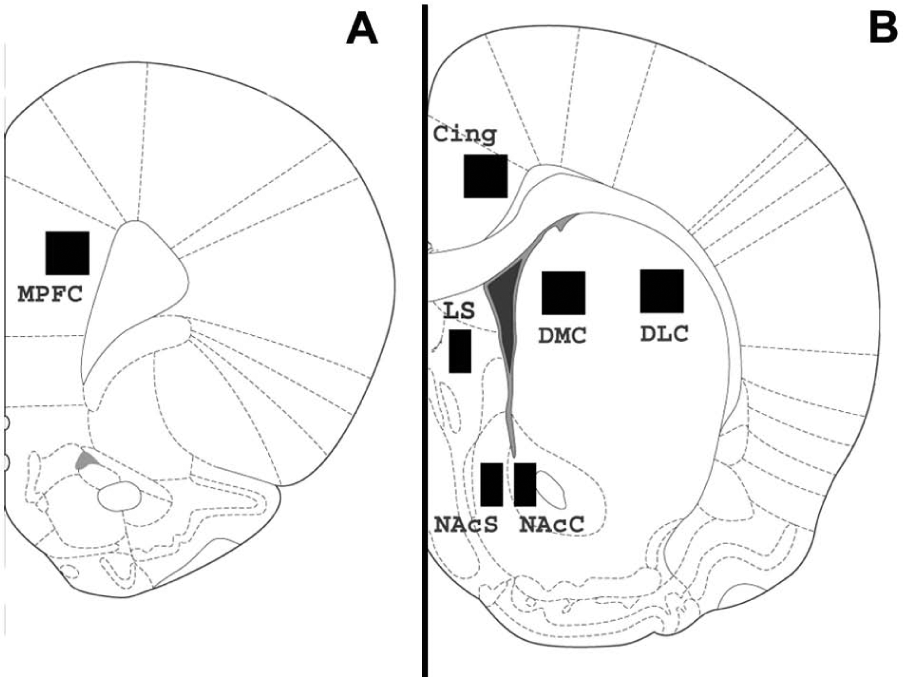
Coronal section of rat brain illustrating the regions selected for immunohistochemical analysis. Section A +3.24 mm relative to Bregma; Section B +1.20 mm relative to Bregma.
Cell counts were expressed as density per mm2. For each rat, an average count of three sections taken from each hemisphere of the brain was used for analysis. The results were decoded, then transferred into GraphPad Prism (GraphPad Software, USA) and the appropriate statistical test was applied.
Statistical analysis
All numerical group data are expressed as mean ± standard error of the mean (SEM). Statistical comparisons were made using one-way ANOVA followed by Bonferroni’s multiple comparisons post-hoc test and the minimum significance level was set at p<0.05.
Results
Caudate nucleus
Expression of Arc-like immunoreactivity (Arc-IR) in dorsolateral caudate was increased following both acute and chronic administration of haloperidol but not after 24 h withdrawal following chronic treatment (Figure 2). A similar pattern of effects was observed in both dorsolateral and dorsomedial caudate with significant increases following acute and chronic haloperidol compared with control but significantly lower Arc-IR expression after chronic than acute administration (Table 1; Figure 3). The relative increases were greater in dorsolateral than dorsomedial caudate. Expression of c-Fos-like immunoreactivity (c-Fos-IR) showed similar changes except for the lack of modulation after chronic versus acute administration in dorsomedial caudate (Table 2). Clozapine did not induce any changes in either Arc-IR or c-Fos-IR expression following either acute or chronic administration in either dorsolateral or dorsomedial caudate.
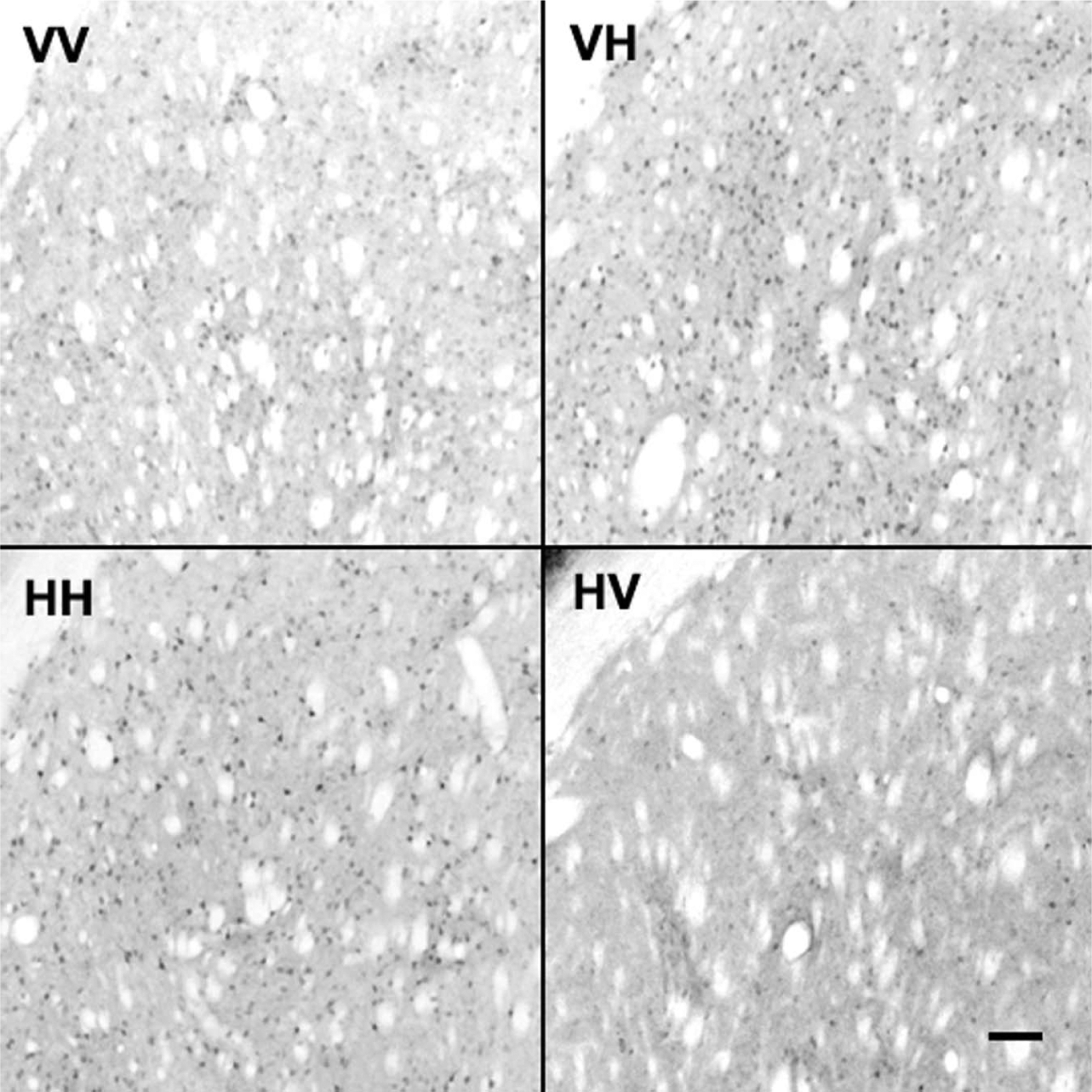
Representative photomicrographs of Arc-like immunoreactive staining in dorsolateral caudate after vehicle or haloperidol pretreatment or challenge. Pretreatment indicates daily administration for 21 days; challenge indicates single administration on day 22; rats were anaesthetised 2 h after the challenge dose and the brain removed as described in Methods. In each case the first letter indicates the pretreatment and the second letter the challenge; ×250 magnification; scale bar 200 μm.
Effect of acute and chronic administration of haloperidol and clozapine on density of cells expressing Arc-like immunoreactivity.
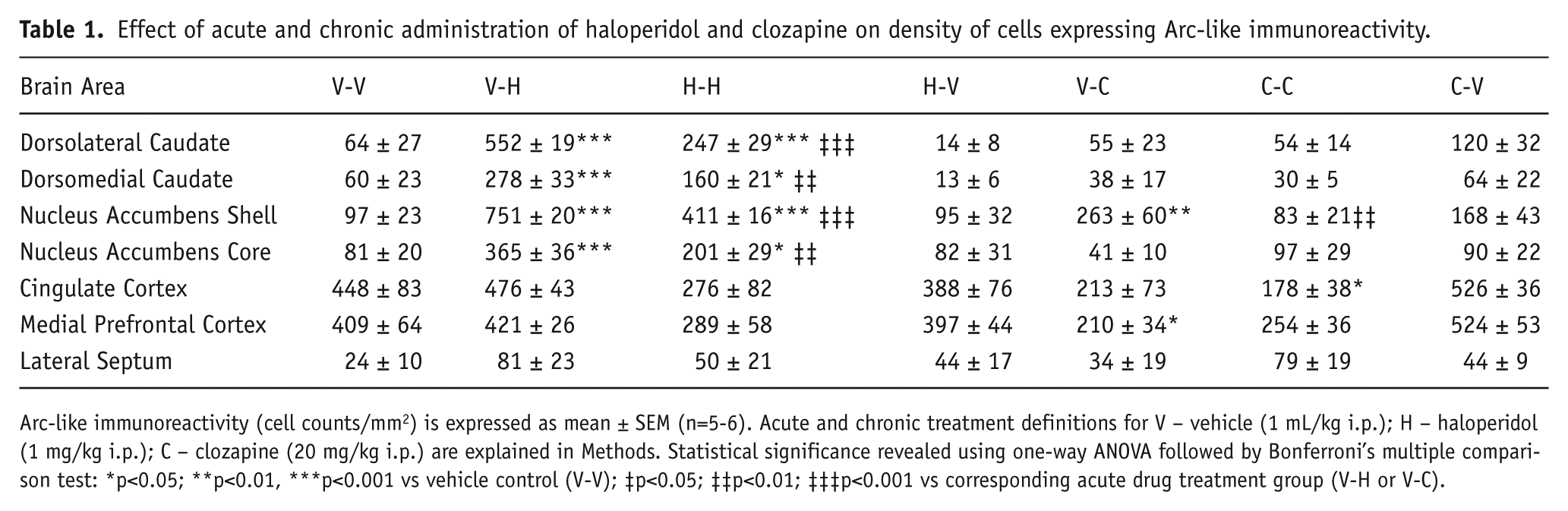
Arc-like immunoreactivity (cell counts/mm2) is expressed as mean ± SEM (n=5-6). Acute and chronic treatment definitions for V – vehicle (1 mL/kg i.p.); H – haloperidol (1 mg/kg i.p.); C – clozapine (20 mg/kg i.p.) are explained in Methods. Statistical significance revealed using one-way ANOVA followed by Bonferroni’s multiple comparison test: *p<0.05; **p<0.01, ***p<0.001 vs vehicle control (V-V); ‡p<0.05; ‡‡p<0.01; ‡‡‡p<0.001 vs corresponding acute drug treatment group (V-H or V-C).

Effects of acute or chronic haloperidol (1 mg/kg i.p.) or clozapine (20 mg/kg i.p.) on expression of Arc-like immunoreactivity in rat brain. Arc-like immunoreactivity (cell counts/mm2) is expressed as mean ± SEM (n=5–6). Treatment groups as defined in Methods. Statistically significant differences revealed by Bonferroni’s multiple comparison test following one-way ANOVA; * p<0.05 vs vehicle control; # p<0.05 vs acute treatment with corresponding drug.
Effect of acute and chronic administration of haloperidol and clozapine on density of cells expressing c-Fos-like immunoreactivity.
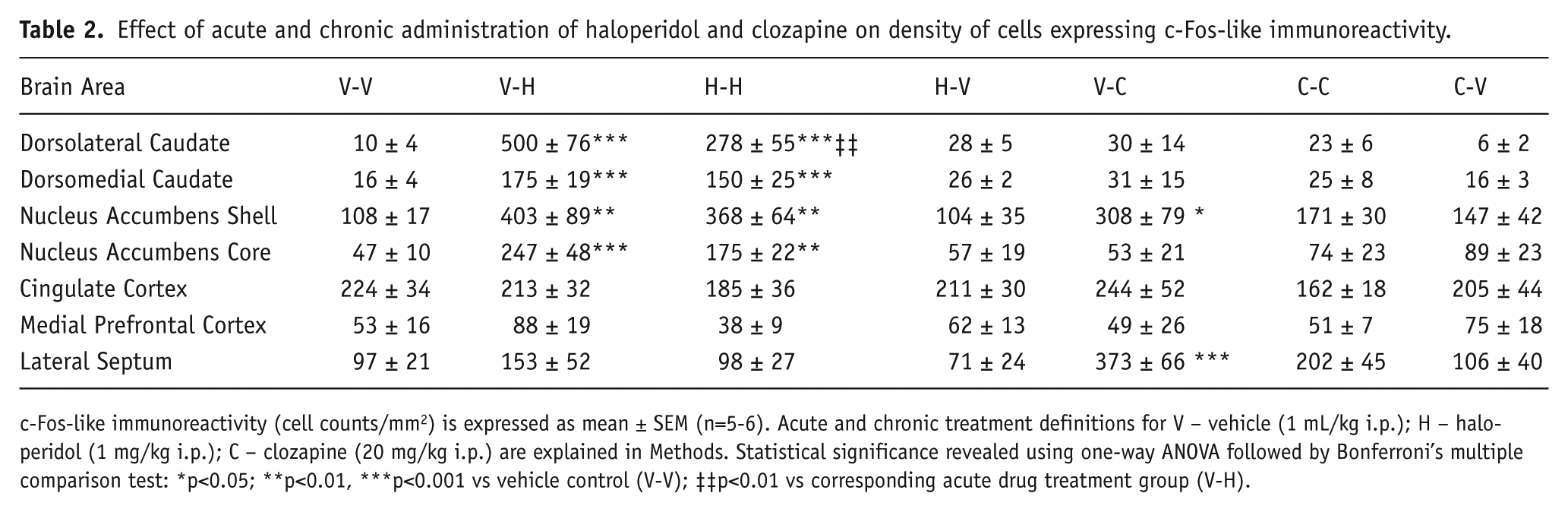
c-Fos-like immunoreactivity (cell counts/mm2) is expressed as mean ± SEM (n=5-6). Acute and chronic treatment definitions for V – vehicle (1 mL/kg i.p.); H – haloperidol (1 mg/kg i.p.); C – clozapine (20 mg/kg i.p.) are explained in Methods. Statistical significance revealed using one-way ANOVA followed by Bonferroni’s multiple comparison test: *p<0.05; **p<0.01, ***p<0.001 vs vehicle control (V-V); ‡‡p<0.01 vs corresponding acute drug treatment group (V-H).
Nucleus accumbens
Arc-IR expression in the nucleus accumbens was increased in the shell but not core region following acute administration of clozapine relative to vehicle control, whereas no increase was evident in either region at 2 h or 24 h following the final dose of chronic treatment (Figure 4). Statistical analysis confirmed a significant increase in the shell following acute administration relative to control and a significant decrease following chronic compared with acute treatment (Table 1; Figure 3). No changes in Arc-IR were evident in the nucleus accumbens core following clozapine. c-Fos-IR expression showed similar changes except the decrease after chronic versus acute administration in the shell was not statistically significant (Table 2). Haloperidol induced significant increases in Arc-IR in both the shell and core following acute and chronic administration compared with control and significantly lower Arc-IR expression after chronic compared with acute treatment (Table 1; Figure 3). Similar changes were observed in c-Fos-IR expression except for the lack of modulation after chronic versus acute administration in both areas (Table 2).
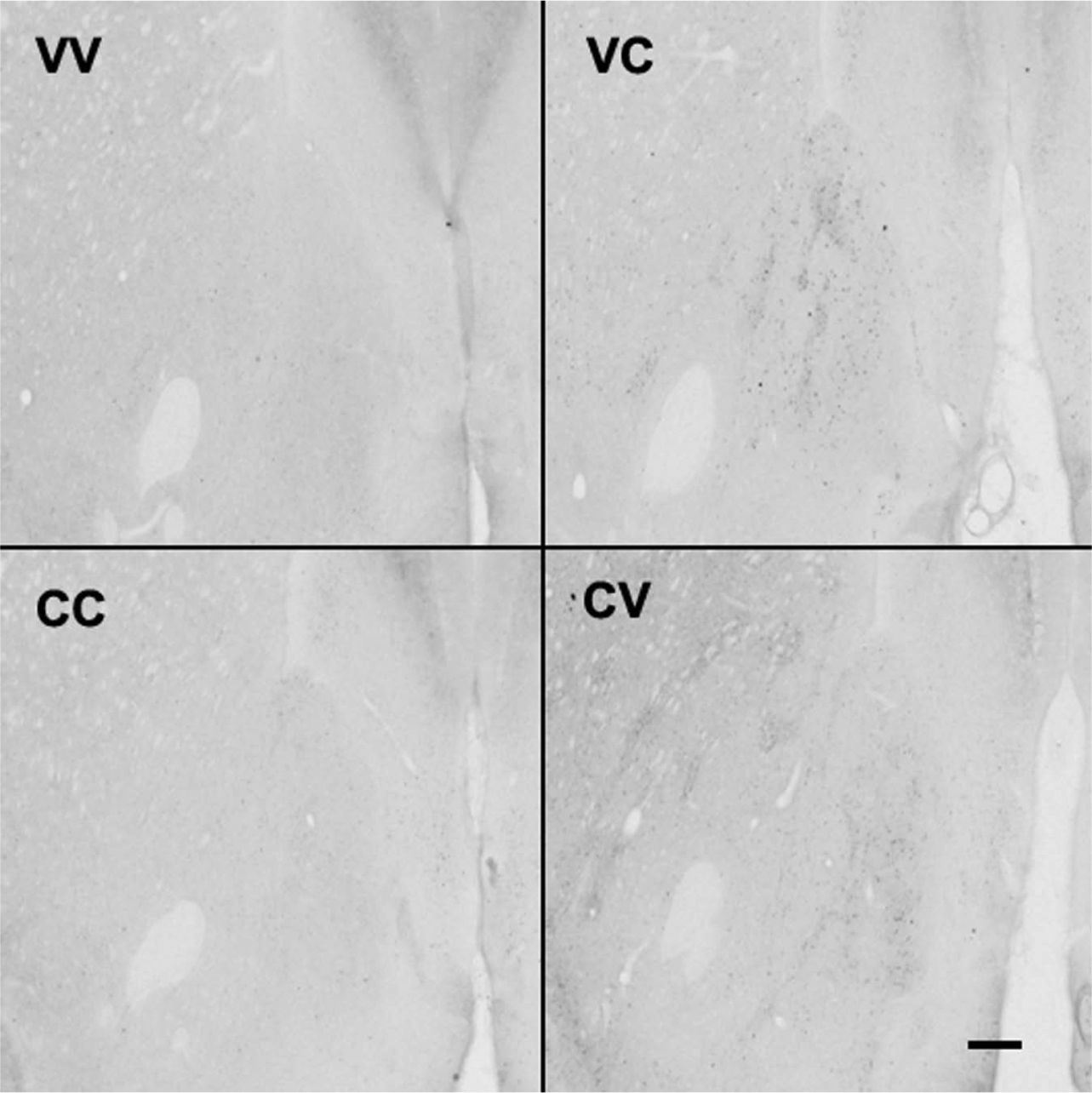
Representative photomicrographs of Arc-like immunoreactive staining in nucleus accumbens after vehicle or clozapine pretreatment or challenge. Pretreatment indicates daily administration for 21 days; challenge indicates single administration on day 22; rats were anaesthetised 2 h after the challenge dose and the brain removed as described in Methods. In each case the first letter indicates the pretreatment and the second letter the challenge; ×125 magnification; scale bar 400 μm.
Medial prefrontal cortex
Arc-IR expression was decreased in medial prefrontal cortex to a similar extent after both acute and chronic administration of clozapine although the latter effect reached statistical significance only on a one-tailed rather than two-tailed test (Table 1; Figure 3). No lasting change was evident at 24 h after the last dose of chronic treatment. No significant changes in c-Fos-IR expression were evident following clozapine administration (Table 2). Haloperidol did not induce any significant changes in Arc-IR or c-Fos-IR expression following either acute or chronic administration.
Cingulate cortex
Similar to the effect observed in MPFC, Arc-IR expression was decreased in the cingulate cortex after both acute and chronic administration of clozapine, although the former effect was statistically significant only by one-tailed test (Table 1; Figure 3). No change was evident at 24 h after the last dose of chronic treatment. No significant changes in c-Fos-IR expression were evident following clozapine administration (Table 2). Haloperidol did not induce any significant changes in either Arc-IR or c-Fos-IR expression following either acute or chronic administration.
Lateral septum
No significant changes in Arc-IR expression were observed in the lateral septum following acute or chronic administration of either haloperidol or clozapine (Table 1). c-Fos-IR expression was significantly increased following acute administration of clozapine but not following either acute or chronic administration of haloperidol.
Discussion
The pattern of expression of Arc protein within the dorsal and ventral striatum demonstrated in the present study following acute administration of the typical antipsychotic drug haloperidol and the atypical drug clozapine broadly matches that previously reported for arc mRNA. Haloperidol strongly induced Arc expression in the dorsolateral caudate whereas clozapine had no such effect, confirming arc mRNA changes both from PCR (Robbins et al, 2008; Bruins Slot et al, 2009: Fumagalli et al, 2009) and in situ hybridisation studies (Iasevoli et al, 2010; de Bartolomeis et al, 2013). These effects are probably mediated by dopamine D2 receptor blockade since a similar increase in arc mRNA was induced by the selective D2 antagonist raclopride whereas arc mRNA was decreased by the D2 agonist quinpirole (Fumagalli et al, 2009). The relatively greater increase in Arc expression induced by haloperidol in dorsolateral than in dorsomedial caudate suggests a greater effect on motor and visuospatial behaviours based on anatomical and functional studies (Featherstone and McDonald, 2004) and corresponds to a similar effect observed for haloperidol compared with amisulpiride on arc mRNA expression (de Bartolomeis et al, 2013). The absence of any Arc induction by clozapine in dorsolateral caudate corresponds to its relative lack of motor effects. In the nucleus accumbens shell both clozapine and haloperidol increased Arc expression whereas haloperidol alone increased Arc in the core region. Since the core region is anatomically linked to motor function via the substantia nigra rather than the association of the shell with limbic structures (Zahm, 2000), this further reinforces the distinction between the typical and atypical antipsychotics characterised by haloperidol and clozapine respectively. The pattern of c-Fos induction observed in this study matches that reported previously in both dorsal and ventral regions of the striatum (Nguyen et al, 1992; Wan et al, 1995). Induction of c-Fos in the nucleus accumbens shell has become a hallmark of the antipsychotic drug profile and the relative scale of induction in this region compared with the dorsolateral caudate has been proposed as a mechanism to differentiate typical from atypical antipsychotics (Robertson et al, 1994). The balance of Arc induction following acute administration of haloperidol and clozapine exhibited in this study suggests a similar pattern applies for Arc as for c-Fos.
Following chronic administration of haloperidol, the extent of Arc and c-Fos induction in dorsolateral and dorsomedial caudate 2 h after the last dose was reduced compared with that observed following acute administration, although remaining significantly greater than control. This mirrors previous studies of arc mRNA and c-Fos (Fumagali et al, 2009; Sebens et al, 1995; Sun et al, 1998) and is probably associated with dopamine D2 receptor up-regulation classically observed following long-term receptor blockade (Huang et al, 1997; Tarazi et al, 1997) and a corresponding increase in dopamine sensitivity (Samaha et al, 2007). In nucleus accumbens shell and core Arc induction was likewise reduced following chronic compared with acute haloperidol administration and a similar effect observed in the shell following chronic clozapine administration, again suggesting a rebound increase in dopamine D2 receptor activity. Although clozapine has lower affinity for the dopamine D2 receptor than haloperidol and in patients D2 receptor occupancy following clozapine is correspondingly lower (Nord and Farde, 2011), previous studies using clozapine at a similar dose over a corresponding treatment period have observed up-regulation of the D2 receptor (Huang et al, 1997). However when examined 24 h after the last chronic dose of either haloperidol or clozapine, neither Arc nor c-Fos levels remained significantly increased relative to control in any of these four striatal regions. This would imply that Arc does not need to be persistently increased to achieve the necessary modulation resulting from chronic neuroleptic drug action. A similar hypothesis has been proposed in relation to occupation of the D2 receptor where persistent rather than intermittent blockade has been associated with the development of vacuous chewing movements in the rat (Turrone et al, 2003) identified as a model of tardive dyskinesia. The pattern of increased striatal Arc expression at 2 h but not at 24 h after the last chronic drug administration mirrors that of other glutamate related genes such as Homer 1a and 1b (Iasevoli et al, 2011) but differs from that of delta-Fos-B (Hiroi and Graybiel, 1996), indicating variation in the degree of tolerance exhibited by different immediate-early genes in response to antipsychotic drug action.
In medial prefrontal and cingulate cortex both acute and chronic clozapine reduced Arc expression albeit with borderline statistical significance. This corresponds with the decrease in arc mRNA following acute and chronic olanzapine reported by Fumagalli et al (2009). Since Arc is associated with internalisation and down-regulation of the AMPA glutamate receptor (Shepherd et al, 2006), these changes may correspond with an increased expression of AMPA receptors and hence glutamate stimulation in these cortical regions. Such an effect would correspond to the glutamatergic hypofunction model of schizophrenia in terms of the putative therapeutic mode of action of antipsychotic drugs (Kantrowitz and Javitt, 2012). Reinforcing this potential impact of Arc reduction, the putative antipsychotic drug LY379268 is reported to increase AMPA receptor sub-unit expression in rat prefrontal cortex and AMPA receptor-mediated current (Wang et al, 2013). Furthermore, behavioural studies indicate that enhancement of AMPA receptor function by administration of ampakine drugs potentiates the effects of olanzapine and risperidone on the conditional avoidance response (Jardemark et al, 2012) and using the sub-chronic phencyclidine schizophrenia model, ampakines attenuate behavioural deficits in a similar way to sertindole (Broberg et al, 2009).
No significant changes in Arc expression were observed following haloperidol administration in contrast to the reduction in arc mRNA reported by Fumagalli et al (2009) although a non-significant decrease did occur following chronic administration in both cortical regions. Autoradiographic studies indicated no change in D2 dopamine receptor density in medial prefrontal cortex following chronic administration of either clozapine or haloperidol (Tarazi et al, 1997) although [3H]spiperone binding was decreased, possibly suggesting mediation via blockade and subsequent down-regulation of 5-HT2A receptors. The absence of any change in c-Fos expression following haloperidol corresponds with previous studies (Wan et al, 1995; Sun et al, 1998).
In conclusion, this is the first study to comprehensively investigate the effects of acute and chronic administration of typical (haloperidol) and atypical (clozapine) antipsychotic drugs on Arc protein expression in rat brain. In dorsal and ventral striatum the changes in Arc protein correspond to previously reported changes in arc mRNA expression and with c-Fos protein, reinforcing the association between induction in the nucleus accumbens shell and antipsychotic drug action. Modulation of Arc induction is evident following chronic administration, which is likely related to D2 dopamine receptor up-regulation but there is no evidence for persistence of induction 24 h following cessation of chronic administration. In medial prefrontal and cingulate cortex Arc protein expression is decreased by clozapine without any evidence of modulation following chronic administration, whereas c-Fos levels are unchanged. This highlights a particular focus of action of antipsychotic drugs on synaptic modulation within these cortical areas that may relate to increased AMPA receptor efficacy, which merits further investigation.
Footnotes
Conflict of interest
The authors declare that there is no conflict of interest.
Funding
We acknowledge financial support from GlaxoSmithKline in partial funding of this project and a doctoral student scholarship from BBSRC for Cheryl Collins.