
Abstract
Administration of neuropeptide S (NPS) elicits anxiolysis, arousal and higher activity in rodents. In humans, the NPS receptor (NPSR1) gene rs324981 A/T (Asn107Ile) polymorphism is associated with fear responses and anxiety. We have recently revealed an association of NPSR1 with impulsivity-related traits and psychopathology. In the present study the association of the NPSR1 genotype with impulsivity and attention-deficit/hyperactivity disorder (ADHD)-related symptoms was re-examined in two independent non-clinical cohorts. We used self-reports of two population-derived samples of the Estonian Psychobiological Study of Traffic Behaviour (EPSTB): a community car driving sample (n=491, MAge=37) and a driving school student sample (n=773, MAge=24). Impulsivity was measured with the Adaptive and Maladaptive Impulsivity Scale (AMIS) in both samples, and with the Barratt Impulsivity Scale (BIS) in driving schools only. For the latter sample, also measurement of ADHD symptoms was carried out with the Adult ADHD Self-Report Scale (ASRS). NPSR1 T-allele carriers had higher scores of impulsivity, motor restlessness and total ADHD scores. The effect on impulsivity originated from male participants but for ADHD symptoms the association was independent of sex. Thus we have confirmed in two additional population-derived samples that the T-allele of the NPSR1 rs324981 polymorphism is associated with increased impulsivity and ADHD-related traits.
Keywords
Introduction
Neuropeptide S is a neurotransmitter mediating central nervous system mechanisms related to arousal, activity and anxiety both in animals and in humans (e.g. Domschke et al., 2011; Reinscheid, 2008). NPS acts via its G-protein coupled receptor NPSR1 that until recently was known as an orphan receptor, GPR154. The NPSR1 gene carries an A/T functional single-nucleotide polymorphism (SNP, rs324981) coding for an Asn-Ile exchange at position 107. NPS has up to 10 times higher potency at the receptor encoded by the T-allele (107Ile) compared to the A-allele encoded receptor, leading to more effective signal transduction with mobilization of intracellular Ca2+, stimulation of cyclic adenosine monophosphate synthesis and induction of mitogen-activated protein kinases phosphorylation (Reinscheid et al., 2005). In animal studies, while administration of NPS elicits an anxiolytic-like effect, it also increases arousal (Lukas and Neumann, 2012; Pulga et al., 2012; Reinscheid, 2008; Rizzi et al., 2008; Smith et al., 2006; Xu et al., 2004). Hence it was predicted that a more active NPS system and the NPSR1 T-allele should be associated with higher dispositional arousal in humans. Indeed, the more ‘efficient’ T-allele has been associated with autonomic arousal reflected by higher heart rate (Domschke et al., 2011), stronger responses to aversive stimuli (Dannlowski et al., 2011; Glotzbach-Schoon et al., 2013; Klauke et al., 2012; Raczka et al., 2010; Tupak et al., 2013), and panic disorder (Domschke et al., 2011; Donner et al., 2010; Okamura et al., 2007). A higher level of internal arousal may lead to impulsive behaviour (Dickman, 2000) and accordingly, in our recent study an association of NPSR1 Asn107Ile with hyperactivity and different facets of impulsivity was revealed (Laas et al., 2014c). In a sample representative of the Estonian population we found that male TT homozygotes displayed the highest level of attention-deficit/hyperactivity disorder (ADHD)-related symptoms overall, and more frequent exposure to stressful life events increased hyperactivity and impulsivity in TT homozygotes in both sexes. Hyperactivity and impulsivity are heritable traits (Crosbie et al., 2013; Faraone et al., 2005) but given the remarkable genetic heterogeneity of ADHD the search for candidate genes has not pointed towards the NPSR1 gene or its locus 7p14.3 (e.g. Banaschewski et al., 2010; Faraone et al., 2005; Lesch et al., 2008). Therefore, we have suggested that NPSR1 may contribute to ADHD-related traits dimensionally through impulsivity and arousal in general population (Laas et al., 2014c).
The purpose of this study was thus to re-examine our previous finding, that the NPSR1 Asn107Ile polymorphism is associated with ADHD-related traits, in additional independent samples hypothesising that this SNP is associated with aspects of impulsivity and hyperactivity in adult males and females. More specifically, we expected to replicate the finding that male T-allele carriers have higher scores of hyperactivity independently of environmental factors.
Method
Sample and data collection
The database of the Estonian Psychobiological Study of Traffic Behaviour (EPSTB; Eensoo et al., 2010; Paaver et al., 2006, 2013) includes two population derived samples. Sample 1 (car drivers) was comprised of car driving male subjects selected randomly from the driving-licence database of the Estonian Motor Vehicle Registration Centre as a control group to traffic law violators (original n=509; mean age (MAge)=36.7±11.8; Paaver et al., 2006). These participants filled in impulsivity self-reports during a visit to laboratory. Sample 2 (driving school; original n=1866, MAge=24.0±8.0) was formed of students applying for a passenger car driving license during an intervention study in driving schools (Eensoo et al., 2010; Paaver et al. 2013). In brief, 24 driving schools out of 54 in the two biggest cities in Estonia, Tallinn and Tartu, were considered eligible and agreed to participate. Subjects from the total of 113 study groups in these driving schools filled in questionnaires during a driving school lesson. Some of them responded to a questionnaire sent by e-mail either two or three years later (see below). Altogether 773 subjects (41%) donated blood samples (males, n=318, MAge=22.6±7.4; females n=455, MAge=25.1±8.2 during the original sampling). Subjects who gave blood samples were older than subjects who refused (24.0±0.3 vs 22.5±0.2, p<0.001); in addition, they had higher adaptive impulsivity (F(1, 1853)=10.5, p=0.001, d=0.015). This study was approved by the Ethics Review Committee at the University of Tartu, Estonia. All participants signed the informed consent form.
Impulsivity and ADHD-related symptoms
All measures were self-reported. For measuring impulsivity, the Adaptive and Maladaptive Impulsivity Scale (AMIS; Paaver et al. 2006) was administered to subjects of both samples. AMIS consists of four subscales in which excitement seeking and fast decision-making reflect adaptive impulsivity, and thoughtlessness and disinhibition reflect maladaptive aspects of impulsivity (Laas et al., 2010). The Driving School sample also reported impulsivity with the Barratt Impulsivity Scale, 11th version (BIS-11, Patton et al., 1995; Paaver et al., 2007). Three years after the initial assessment in traffic schools, the Adult ADHD Self-Report Scale (ASRS, Kessler et al., 2005) for measuring ADHD-related symptoms was delivered and returned by e-mail.
NPSR1 polymorphism genotyping
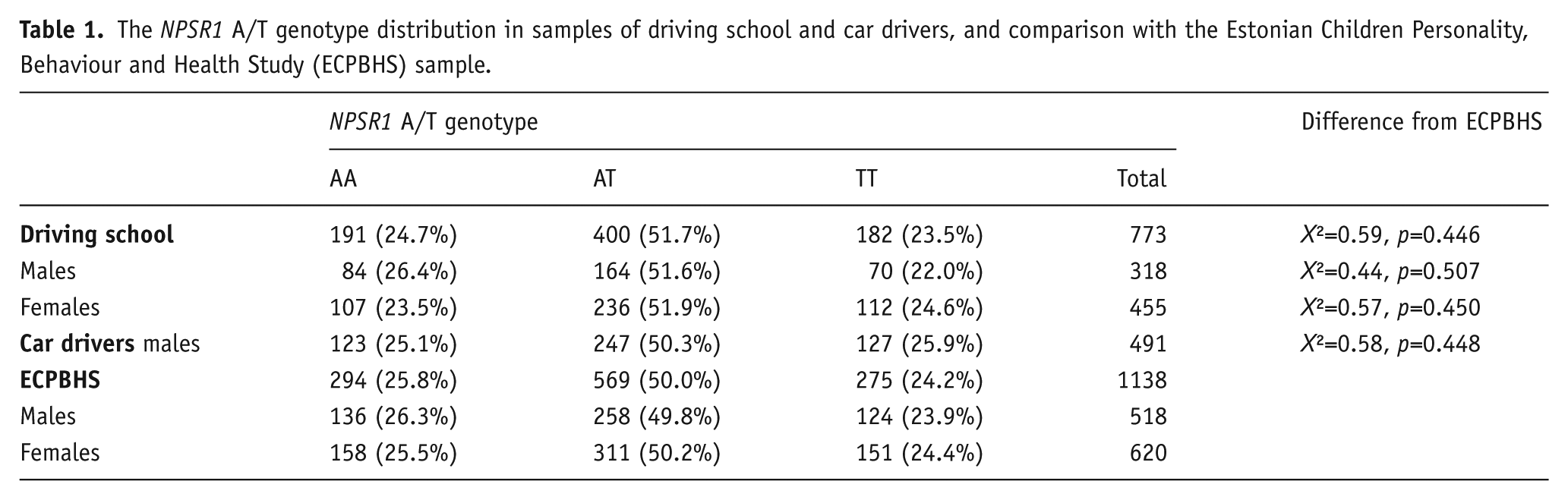
DNA was isolated from venous blood. NPSR1 rs324981 was genotyped by routine polymer chain reaction followed by restriction enzyme digest and gel electrophoresis as previously described (Domschke et al., 2011). Genotype frequencies were in Hardy–Weinberg equilibrium. The NPSR1 A/T polymorphism was genotyped in 773 subjects in the driving school sample and 491 subjects in the car drivers sample (Table 1). The distribution of NPSR1 rs324981 genotype was similar to the distribution found previously in a sample Estonian Children Personality, Behaviour and Health Study (ECPBHS) representative of the Estonian population (Laas et al., 2014c).
The NPSR1 A/T genotype distribution in samples of driving school and car drivers, and comparison with the Estonian Children Personality, Behaviour and Health Study (ECPBHS) sample.
Data analysis
Differences in NPSR1 rs324981 genotype distribution between samples were calculated by Pearson’s Chi Square tests. We used analysis of covariance to reveal differences in impulsivity and ADHD symptom raw scores by NPSR1 genotype. Model assumptions were checked by the Levene’s test of equality of error variances and standardized residual plots. Effect sizes were expressed as Cohen’s d. Age was used as a covariate because it was reversely associated (Pearson correlation) with outcome variables. Analyses were also done separately for males and females as sex differences had been reported previously (Okamura et al., 2007; Dannlowski et al., 2011; Domschke et al., 2011; Laas et al., 2014a; Laas et al., 2014b; Laas et al., 2014c). Based on previous studies on impulsivity and preliminary analysis (Figure 1), T-allele carriers were compared with AA homozygotes. To detect one score difference in impulsivity subscales the sufficient power was 0.96 for the the sample of car drivers, and 0.8 for the subsample of males of driving school.
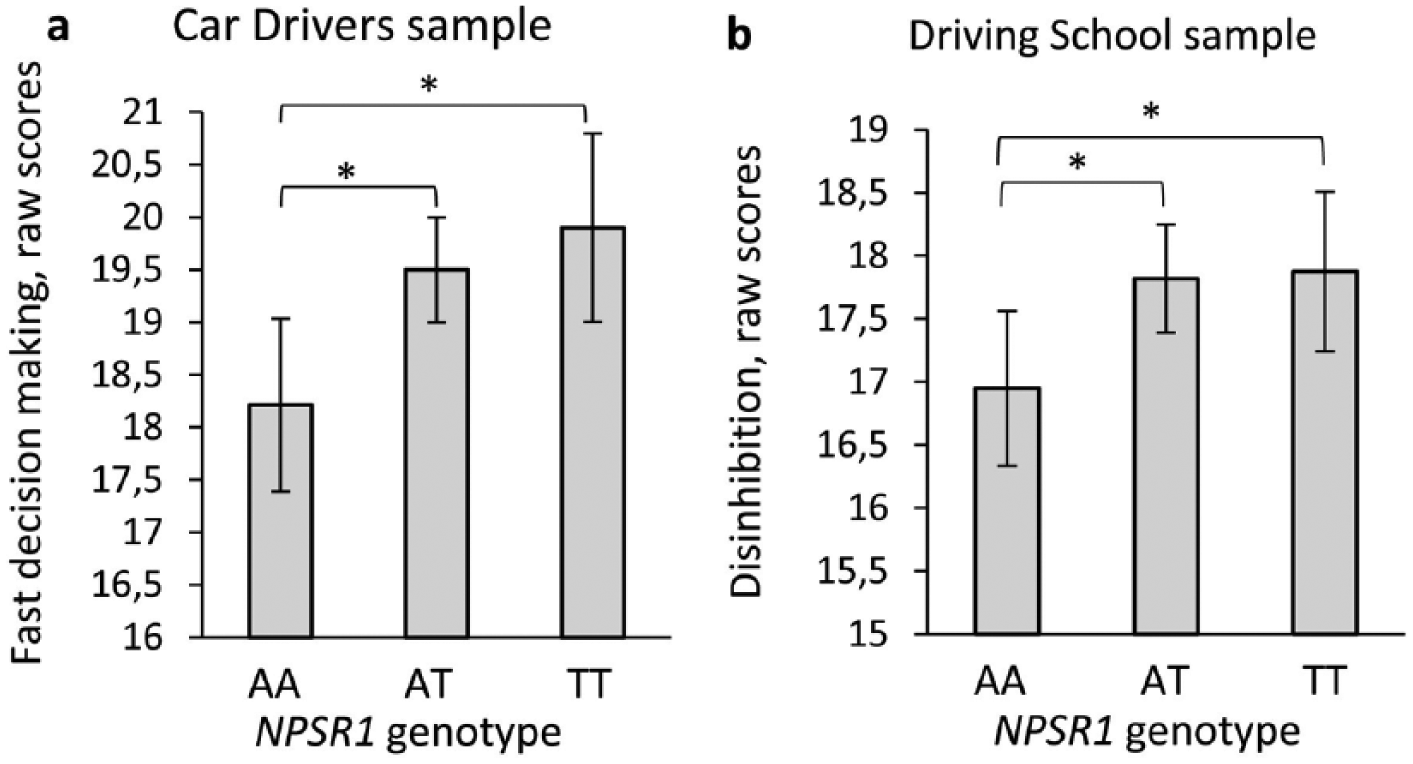
NPSR1 A/T genotype effects on impulsivity in two samples. (a) NPSR1 effect on Fast Decision Making in car drivers sample, F(2, 479)=4.76, p=0.009; (b) NPSR1 effect on Disinhibition in driving school sample, F(2, 760)=3.03, p=0.049. Whiskers indicate 95% confidence intervals.
Results
NPSR1 A/T polymorphism, impulsivity and ADHD-related symptoms
In general, males had significantly higher scores of adaptive impulsivity compared to females (not shown). No significant sex difference in BIS, maladaptive impulsivity and ADHD-related symptoms were detected.
The NPSR1 A/T polymorphism was associated with both impulsivity and ADHD symptom-related measures. In both samples, male T-allele carriers had higher scores of impulsivity (Table 2). However, in car drivers, T-allele carriers had higher adaptive impulsivity; whereas in driving school males, T-allele carriers had higher maladaptive impulsivity. It may be relevant that differences in maladaptive impulsivity scores were larger between males of the two samples as compared to differences between males and females within the same sample. When the males of the two samples were combined, T-allele carriers exhibited higher scores of disinhibition, fast decision making, adaptive impulsivity and the total AMIS scale. The effect of the T-allele on disinhibition remained significant in the pooled sample containing the data of all the subjects, including females.
Main statistics of NPSR1 A/T genotype effects on impulsivity (group means and standard errors), covariate age. Bold figures indicate significant difference.
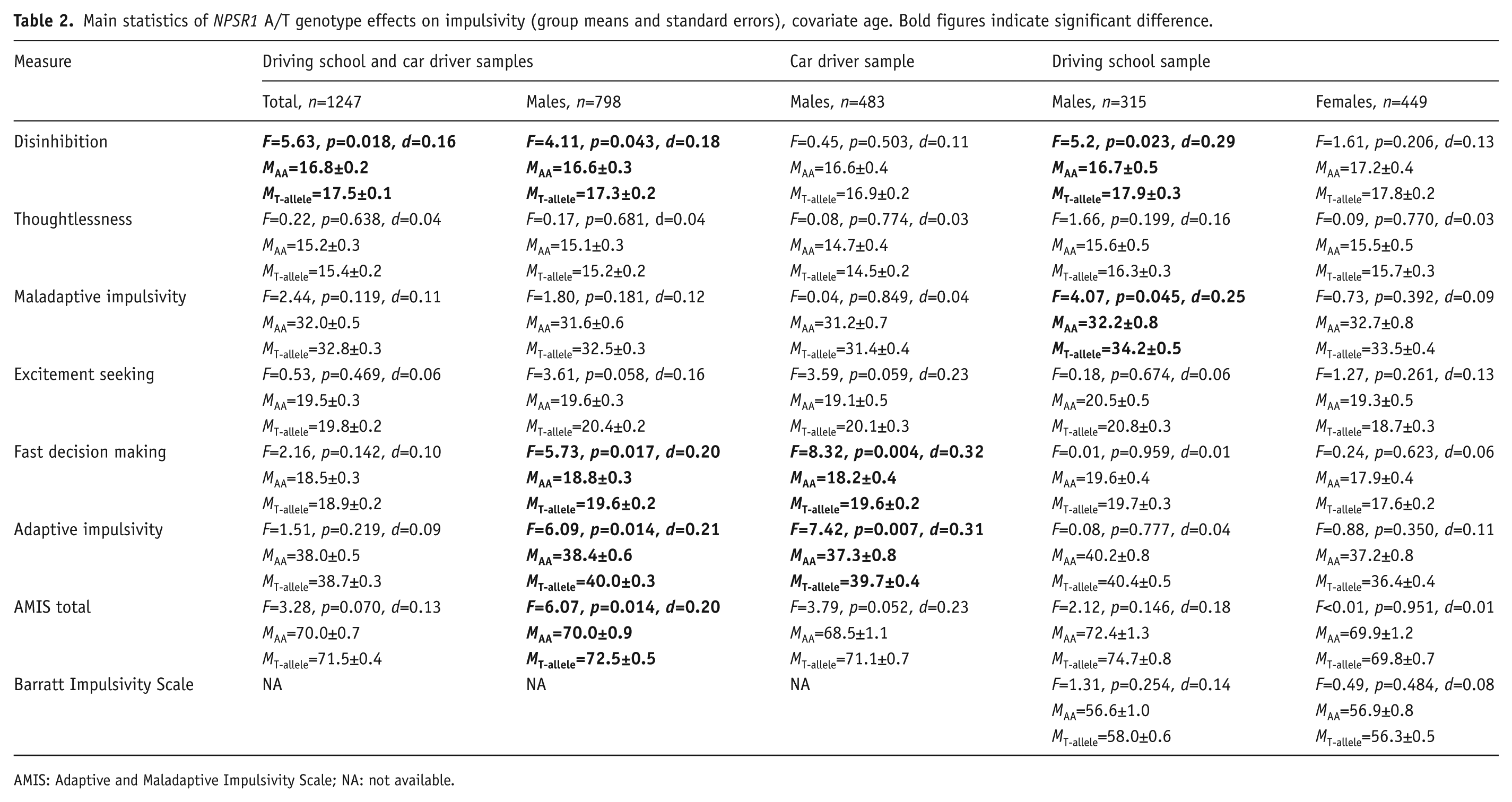
AMIS: Adaptive and Maladaptive Impulsivity Scale; NA: not available.
Subjects with the T-allele also reported more ADHD-related symptoms: scores of both the Hyperactivity subscale and the total ADHD score were higher in T-allele carriers (Table 3). No significant NPSR1 × sex interaction effects were found; the effect originated from both sexes.The differences in scores with alpha level less than 0.05 ranged from 0.7–1.4 with the effect sizes from 0.16–0.32.
Main statistics of NPSR1 A/T genotype effects on attention-deficit/hyperactivity disorder (ADHD) scale (group means and standard errors) in driving school sample, covariate age. Bold figures indicate significant difference.
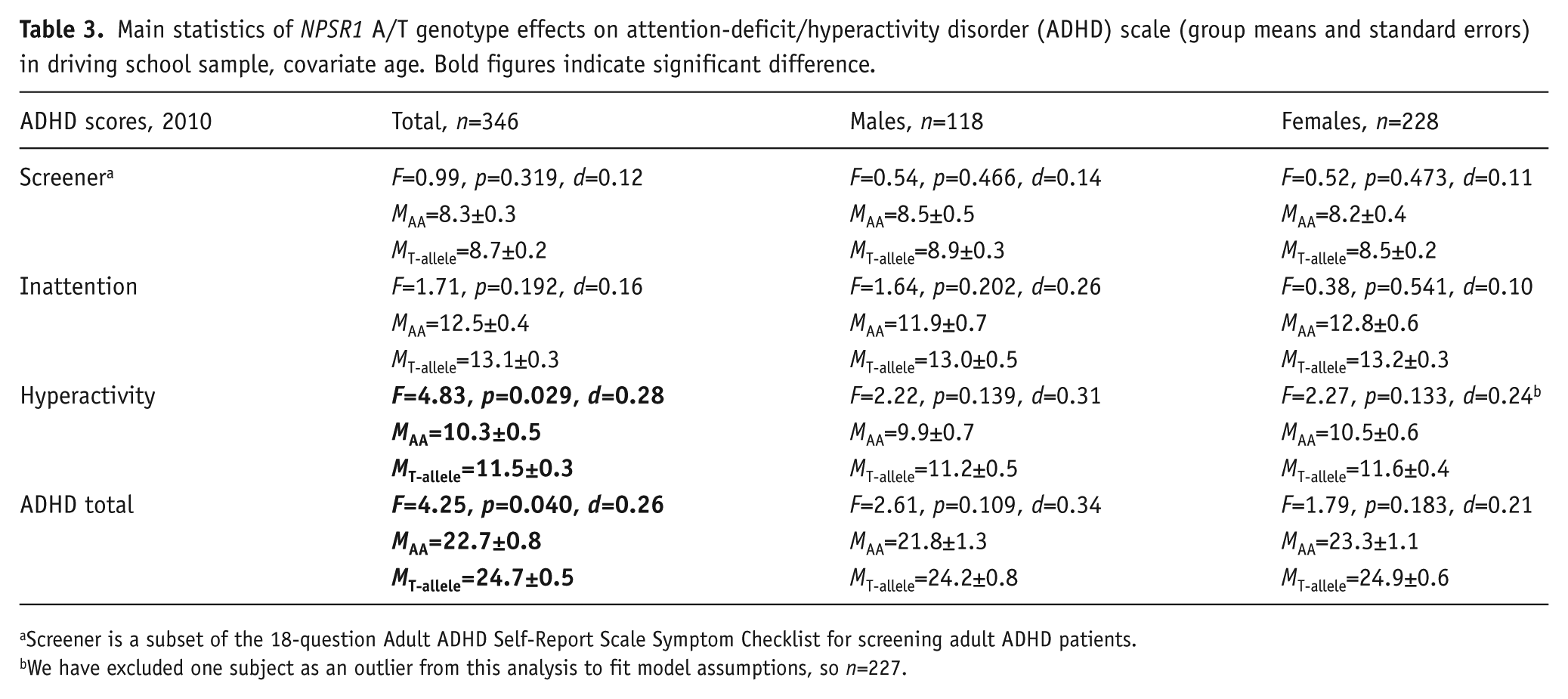
Screener is a subset of the 18-question Adult ADHD Self-Report Scale Symptom Checklist for screening adult ADHD patients.
We have excluded one subject as an outlier from this analysis to fit model assumptions, so n=227.
Discussion
Our results in two independent samples of adult male and female subjects provide further supportive evidence for the association of the NPSR1 A/T polymorphism with impulsivity and hyperactivity. The T-allele carriers, and males in particular, have higher scores of both impulsivity and ADHD-related symptoms. This falls in line with the findings in our previous study in a population-representative sample of adolescents and young adults (Laas et al., 2014c) where TT homozygous males displayed the highest level of ADHD-related symptoms. The distribution of NPSR1 A/T polymorphism in these three population-derived non-clinical samples was similar. These findings are consistent with both animal and human studies where indices of higher NPS-ergic activity are associated, in general, with higher arousal and activity levels (Domschke et al., 2011; Pulga et al., 2012; Reinscheid, 2008; Rizzi et al., 2008; Smith et al., 2006; Xu et al., 2004).
Recently, Beiderbeck et al. (2014) reported that NPS had anti-aggressive effects in male rats independent of both anxiolysis and general social behaviour. Hence it is obvious that, while aggression and anxiety have common denominators, they can be independently regulated (reviewed by Neumann et al., 2010), and the NPS-ergic neurotransmission is rather what makes them distinct. So how would the higher impulsivity/hyperactivity in the T-allele (that corresponds to more efficient NPS neurotransmission) carriers fit with low anxiety and low aggressiveness, given that impulsivity is often a predictor of aggressive behaviour? There is as yet no mechanistic explanation at the level of neural pathways but the consistent findings on the role of NPS receptor function in impulsivity but not aggression suggest that studies on chemical neuroanatomy of NPS may help to reveal why some impulsive subjects behave aggressively while others do not.
While in general the association of the NPSR1 genotype and certain impulsivity measures and ADHD symptoms was corroborated, this study also revealed distinct genotype associations with aspects of impulsivity when comparing the two samples. According to the AMIS scale that enables to differentiate between adaptive and maladaptive types of impulsivity (Dickman, 1990; Paaver et al., 2006), the effects of the T-allele differed in the samples: In car drivers, the T-allele carriers had higher adaptive impulsivity; but in the driving school sample, the T-allele carriers had higher maladaptive impulsivity. The reasons behind this distinction remain ambiguous for the time being but may have to do with inherent differences between samples, and gene-environment interactions (G×E). Regarding the sample formation, while subjects of both car drivers and driving school samples were population derived, the participation rate in each was 30–40%, allowing for important dissimilarities in response bias especially as the recruitment of the former group occurred on average five years earlier. Second, responders may differ if invited by phone call (car drivers) or in direct contact during a lesson (driving school).
Furthermore, car drivers were on average twelve years older than the driving school subjects. Apart from simple age difference effect this also facilitates distinct G×E effects. Interaction effects of genotype and birth cohort can occur, being probably mediated by changes in environment when a society is in transition (Vaht et al., 2014). However, in this study, we had no self-reports of life events or family environment to test for the quantifiable G×E effects, so such an interpretation remains speculative.
As a limitation, it should be made clear that statistical significance of most of the group differences would not withstand the conservative correction for multiple testing. However, we had drawn the hypotheses based on our previous findings (Laas et al., 2014c); in addition, all the results point to the same direction of the T-allele being associated with increased hyperactivity and impulsivity, this also being in line with animal studies (e.g. Rizzi et al., 2008; Xu et al., 2004). Another limitation of the study is the use of self-reports, but the results fall in line with our previous data based on teacher’s ratings of hyperactivity symptoms.
In conclusion, evidence is accumulating that NPSR1 A/T T-allele carriers, especially males, have higher impulsivity and hyperactivity. This suggests that higher signalling efficiency via the NPS receptor may serve as a future drug target in impulsivity-related disorders.
Footnotes
Acknowledgements
The authors are grateful to the EPSTB study participants and the whole study team.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Estonian Ministry of Education and Science Project IUT20-40; Estonian Road Administration; European Regional Development Fund ERC Program TerVE project VIGA 3.2.1002.11-0002 and the EC FP7 project Aggressotype (FP7-Health-2013-Innovation-1 602805); the German Research Foundation, DFG (SFB TRR 58, A05, B06, C02, and Z02 to KPL and AR, RTG 1256, to AR and KPL); and the Bundesministerium für Bildung und Forschung (BMBF) grant (DZHI, project 01EO1004, to AR and KPL).