
Abstract
The serotonin-6 (5-HT6) receptor is the most recently discovered serotonin receptor, and it represents an increasingly promising target for improving cognition in both normal and disease states. Recently, a new selective 5-HT6 receptor agonist, 2-(5 chloro-2-methyl-1H-indol-3-yl)-N,N-dimethylethanamine (ST1936), with nanomolar affinity for 5-HT6 receptors was described. We performed in-vivo electrophysiological studies to investigate the physiological role of 5-HT6 receptors in the control of the function of the substantia nigra pars compacta (SNc) and ventral tegmental area (VTA). Extracellular single-unit recordings were performed from putative dopamine-containing neurons in the SNc and VTA of anesthetised rats. In the SNc, acute systemic administration of ST1936 had no effects on basal firing activity of these dopamine neurons; however, in the VTA, ST1936 induced either dose-related increases (45% of cells) or decreases in basal activity of these dopaminergic neurons. Local application of ST1936 into the VTA caused excitation in all of the dopamine neurons, but had no effects on non-dopamine VTA neurons. Both effects of systemic and microiontophoretic ST1936 were completely reversed by the potent and selective 5-HT6 receptor antagonist 5-chloro-N-(4-methoxy-3-piperazin-1-ylphenyl)-3-methyl-2- benzothiophene-sulfonamide (SB271046). Systemic application of another 5-HT6 agonist, 2-(1-{6-chloroimidazo[2,1-b] [1,3]thiazole-5-sulfonyl}-1H-indol-3-yl)ethan-1-amine (WAY-181187), induced dose-dependent inhibition of these VTA dopaminergic neurons. ST1936 and WAY-181187 appear to have different effects on these VTA dopaminergic neurons, potentially due to different mechanisms of action or to the complexity of 5-HT6 receptor functions. Our data demonstrate the need for further investigations into the use of 5-HT6 receptor agonists to control cognitive disfunction, such as in schizophrenia and depression.
Keywords
Introduction
The serotonin-6 (5-HT6) receptor is one of the most recently discovered of the serotonin receptors, and it was first identified and sequenced in 1993; it is one of the three serotonin receptors that are positively coupled to the Gs protein, with the induction of cAMP production (Monsma et al., 1993; Plassat et al., 1993; Ruat et al., 1993). In the past decade, this receptor has become an increasingly promising target for the improvement of cognition in both normal and disease states (Mitchell and Neumayer, 2005). Indeed, molecular biology studies in adult animals have demonstrated the highest 5-HT6 receptor expression in the striatum, nucleus accumbens, olfactory tubercle, and cortex, with moderate expression in the amygdala, hypothalamus, thalamus, cerebellum, and hippocampus, but not in cell bodies of the raphe nuclei, which suggests a postsynaptic role for this receptor (Boess et al., 1998; Gérard et al., 1997; Yoshioka et al., 1998). Interestingly, the distribution of 5-HT6 receptor mRNA in human brain parallels that of rat brain (Kohen et al., 1996). Additionally, 5-HT6 receptors are localised on striatal γ-aminobutyric acid (GABA)-ergic interneurons, which also receive dopamine (DA) input (Ward and Dorsa, 1996). Therefore, the relative abundance of 5-HT6 receptor-like immunoreactivity in extrapyramidal and limbic areas, together with the finding that some atypical antipsychotic drugs and antidepressant agents are relatively potent 5-HT6 receptor antagonists (Monsma et al., 1993; Roth et al., 1994), suggest that the 5-HT6 receptor participates in serotonergic control of motor function, and modulation of DA-dependent behaviour, and thus of psychiatric diseases such as schizophrenia and depression (Gérard et al., 1997; Reavill and Rogers, 2001).
However, the function of the 5-HT6 receptor remains unclear, due to limited knowledge of their transduction mechanisms, lack of full centrally acting agonists, and inconsistencies in the pharmacological and neurochemical effects of their antagonists (Borsini et al., 2011; Di Matteo et al., 2008; Marazziti et al., 2011). DA-ergic neurons in the ventral tegmental area (VTA) showed decreased spontaneous activity after acute administration of the 5-HT6 antagonist 5-chloro-N-(4-methoxy-3- piperazin-1-ylphenyl)-3-methyl-2-benzothiophene-sulfonamide (SB271046; Minabe et al., 2004). However, chronic administration (21 days) of SB270146 did not alter the firing patterns of these VTA neurons. In contrast, substantia nigra pars compacta (SNc) neurons showed no changes in spontaneous firing with acute administration of SB271046, but demonstrated an increase in firing following chronic SB271046 administration. Thus, it appears that 5-HT6 blockade has less effect on DA regulation in the nigrostriatal system than the mesocortical system, due to differences in 5-HT6 receptor expression and/or localisation, and 5-HT-ergic tone (Minabe et al., 2004).
However, contrasting data have been obtained in neurochemical studies. For example, systemic administration of selective 5-HT6 receptor antagonists increased dialysate levels of DA, but not of 5-HT, in the rat prelimbic/infralimbic subregion of the prefrontal cortex (Lacroix et al., 2004) and hippocampus (Li et al., 2007). On the contrary, other studies have failed to demonstrate direct effects of 5-HT6 antagonism on cortical (Dawson and Li, 2003; Dawson et al., 2000, 2001; Li et al., 2007) and hippocampal (Dawson et al., 2001) DA efflux. Furthermore, in other DA-ergic regions, such as the nucleus accumbens and striatum, the selective blockade of 5-HT6 receptors had no effect on basal DA release (Dawson and Li, 2003; Dawson et al. 2000, 2001).
Based on the above considerations, we investigated the physiological effects of the new full 5-HT6 receptor agonist 2-(5 chloro-2-methyl-1H-indol-3-yl)-N,N-dimethylethanamine (ST1936; Riccioni et al., 2011; Valentini et al., 2011) on the electrical activity of putative DA neurons in the midbrain, using in-vivo single-cell recording. Although ST1936 is a highly selective agonist of 5-HT6 receptors (Ki=13, 168 and 245 nM, for human 5-HT6, 5-HT7 and 5-HT2B receptors, respectively), it also shows moderate affinity (Ki=300 nM) for human α2 adrenergic receptors (Riccioni et al., 2011; Valentini et al., 2011). Thus we also evaluated the effects of 5-HT6 receptor blockade on the ST1936-induced changes using the potent and selective 5-HT6 receptor antagonist SB271046 (Bromidge et al., 1999; Miguel-Hidalgo, 2001). Finally, the effects of systemic 5-HT6 agonist 2-(1-{6-chloroimidazo[2,1-b] [1,3]thiazole-5-sulfonyl}-1H-indol-3-yl)ethan-1-amine (WAY-181187) (Cole et al., 2007; Schechter et al., 2008) were compared to those of ST1936 in the VTA.
Materials and methods
Animals
Male Sprague-Dawley rats (Charles River, Calco, Italy) that weighed 340–380 g were used. The rats were kept at constant room temperature (21±1°C) and relative humidity (60±5%) under a regular light/dark schedule (light, 08:00–20:00 hours). Food and water were supplied ad libidum. The procedures that involved the rats and their care were conducted in conformity with the institutional guidelines, which are in compliance with national (D.L. n. 116, G.U., suppl. 40, 18 February 1992) and international laws and policies (EEC Council Directive 86/609, OJ L 358,1, 12 December 1987; National Institutes of Health Guide for the Care and Use of Laboratory Animals, NIH Publication N. 85-23, 1985; Giles, 1987). All efforts were made to minimise the number of animals used and their suffering.
Single-cell recording procedures
The rats were anesthetised with chloral hydrate (400 mg/kg i.p.) and mounted on a stereotaxic instrument (SR-6; Narishige, Tokyo, Japan). Supplemental doses of anaesthetic were administered via a lateral tail vein cannula. Throughout the experiments, the body temperature of the rats was maintained at 36–37°C, using a thermostatically regulated heating pad.
Relative to the interaural line, the coordinates for placement of the recording electrodes in the VTA were: anterior, 2.7–3.4 mm; lateral, 0.1–0.5 mm; and ventral, 7–8 mm, to the level of exposed tissue. For the SNc, the coordinates were: anterior, 2.7–3.4 mm; lateral, 1.8–2.2 mm; and ventral, 6.5–7.5 mm (Paxinos and Watson, 1986). Extracellular recordings were performed using either single or multi-barrel micropipettes. For the study of systemic drug effects on the SNc and VTA putative DA-ergic neurons, the single-barrel micropipettes used measured 2–3 µm at the tip, and were filled with 2% Pontamine Sky Blue dye in 2 M NaCl (in vitro resistance, 4–7 MΩ). For the study of local drug effects on putative DA and non-DA spontaneously active VTA neurons, four-barrel or five-barrel homemade micropipettes were used.
Putative DA neurons were identified by their location, waveform, firing rate, and pattern, which included: spontaneous (basal) slow firing rate (2–10 Hz), in regular or irregular patterns, including bursts (i.e. series of 2–10 spikes of decreasing amplitude and increasing duration, and interspike interval of 80–160 ms within a burst); biphasic (positive-negative) action potentials, typically with a ‘notch’ in the rising phase and a prominent negative component; and action potential duration >2.2 ms overall. The spikes typically varied in shape within a train from a single neuron, due to interactions between the slow depolarisation of the preceding spike and the prominent inhibitory synaptic input of biphasic or triphasic action potentials with a duration of ≥1.1 ms (measured from spike initiation to the maximal negative phase of the action potential), and to inhibition by systemic administration of low doses of DA or DA receptor agonists (e.g. apomorphine i.v.) for nigrostriatal or mesolimbic DA neurons (Bunney et al. 1973; Grace and Bunney, 1980; Ungless and Grace, 2012; Wang 1981). Although DA neurons represent the majority within the VTA, at around 70%, there are subregional variations, with around 30% GABA-ergic neurons and 2–3% glutamatergic neurons also in the VTA (Ungless and Grace, 2012). Non-DA neurons (which were presumed to be GABA-ergic) were identified on the basis of established electrophysiological characteristics: short-duration (<0.7 ms), biphasic (positive/negative) or triphasic (positive/negative/positive) action potentials of large amplitude (1.0–1.5 mV); and baseline firing rates between 10 spikes/s and 40 spikes/s, with nonbursting activity (Steffensen et al., 1998; Zhang et al., 1993).
The electrical signals of the spike activities were passed through a high-impedance amplifier, the output of which entered an analogue oscilloscope, with an audio monitor and window discriminator. The unit activity was then converted into an integrated histogram by rate averaging, and displayed as spikes per 10 s interval. After each experiment, the recording site was marked by the ejection of Pontamine Sky Blue dye from the electrode using a 20 μA current for 10 min. The rat brains were removed and placed in 10% buffered formalin for two days, prior to histological examination. Frozen sections were cut at 40 μm intervals and stained with neutral red. Microscopic examination of the sections was carried out to verify that the electrode tip had been in the VTA or the SNc. Only recordings of neurons into the VTA and SNc were considered for the data analysis.
Drug administration protocols
Four distinct experimental groups of rats were used for the systemic acute study of ST1936, as the following treatments:
Group 1: Vehicle of ST1936 (0.9% saline)+vehicle of SB271046 (see below): control.
Group 2: Vehicle of ST1936+SB271046.
Group 3: ST1936+vehicle of SB271046.
Group 4: ST1936+SB271046.
ST1936 was freshly diluted in physiological saline (0.9% NaCl) (Valentini et al., 2011), and systemically injected via a lateral tail vein (i.v.) every 2 min at increasing cumulative doses (50–1600 μg/kg for the SNc; 50–400 μg/kg for the VTA). The effects of ST1936 on the basal firing activity of putative DA neurons were recorded in both brain areas.
The vehicle for the potent and selective 5-HT6 receptor antagonist SB271046 was 0.9% NaCl containing 8% (w/v) hydroxypropyl-ß-cyclodextrin and 25 mM citric acid (henceforth referred to as ‘vehicle’). SB271046 was administered at 100 and 200 μg/kg, i.v. either after saline (control) or after the increasing cumulative doses of ST1936, to determine the specificity of ST1936 as a selective 5-HT6 agonist under our experimental conditions. The doses of ST1936 and SB271046 were chosen on the basis of preliminary experiments.
The new 5-HT6 receptor agonist WAY-181187 was dissolved in saline (Schechter et al., 2008) and given systemically (50–1600 μg/kg, i.v.) for the VTA, to compare the effects of the two 5-HT6 receptor agonists (ST1936, WAY-181187) on these DA neurons.
Control rats were treated with repeated injections of saline and/or vehicle of SB271046 every 2 min. Apomorphine (10–30 μg/kg i.v.; in saline) was given at the end of each experiment to confirm the DA-ergic identity of the neuron recorded. Only one cell per animal was studied. Each vehicle and drug application was given as a volume of 100 μL.
In a final series of experiments, the effects of local administration of both ST1936 and SB271046 in VTA DA and non-DA neurons was evaluated using multibarrel micropipettes, by microiontophoresis carried out in drug-naïve rats. Four-barrel homemade micropipettes were pulled to an optimal wide tip angle, and mechanically bevelled under microscopic control to a final tip diameter of 4–6 μm. The central barrel was filled with 2% Pontamine Sky Blue dye in 2 M NaCl, (in vitro resistance, 4–7 MΩ) and was used for the recording, while one of the side barrels was filled with 2 M NaCl and used for continuous automatic current balancing. The remaining barrels were filled with ST1936 (40 mM, pH 4.5; in water), and DA hydrochloride (100 mM, pH 4; in water), for the study of local drug effects on spontaneously active DA and non-DA VTA neurons. ST1936 was applied by microiontophoresis for 60 s using ejection currents of +5, +10 and +15 nA, while a retention current of 10 nA was used to prevent leakage between ejection periods (60 s). To determine the specificity of ST1936 as a selective 5-HT6 agonist under our experimental conditions, five-barrel micropipettes were used, with SB271046 (20 mM, in 10% dimethyl sulfoxide in water) administered locally by micropressure ejection at 12 p.s.i., at 1–2 min before and during application of ST1936. It is important to note that microiontophoresis of SB271046 was not possible to perform due to its low solubility in water. DA was ejected (+20 nA, 60 s) at the end of each experiment, to confirm the DA-ergic identity of the neurons recorded.
Data analysis and statistics
Data acquisition and analysis were carried out using an 83386-based PC and an integrated software package for electrophysiology (RISI; Symbolic Logic, Dallas, Texas, USA). Dose-response curves were constructed by comparing the mean firing rates of 300–500 consecutive spikes, starting immediately after the injection of each drug, with the basal firing rate. Burst analysis of DA-ergic neurons was performed using the RISI program running on a PC computer. A total of 300–500 consecutive spikes were recorded for each neuron before and at the peak of the drug effects. When present, burst firing was detected using an algorithm similar to that previously described by Grace and Bunney (1984): bursts were defined in the standard fashion, starting with an interspike interval of 80 ms, and ending with the concurrence of two spikes with an interspike interval of 160 ms. The drug-induced changes in the neuronal firing rates were analysed by two-way analysis of variance (ANOVA) with one factor repetition, followed by Fisher’s protected least significance difference (PLSD) post-hoc tests, to allow multiple comparisons between groups. The percentage changes in the firing rates of spontaneously active (basal) DA and non-DA neurons produced by the iontophoretic application of ST1936 were calculated by subtracting the mean numbers of action potentials recorded during the periods of iontophoresis (ejection time, 60 s) from those obtained in the same periods prior to microiontophoresis with ST1936 (retention time, 60 s), and then dividing these by the latter and multiplying by 100. The data regarding the effects of ST1936 were subjected to ONE sample Students’ t-tests for comparisons between the mean effects of each ST1936 current ejection (+5 to +15 nA) and the basal value that was considered as 100%. Two-way ANOVA with one factor repetition was used for the analyses between groups. The value of p<0.05 was regarded as significant. All of the statistical analyses were performed with Stat-View version 5.0.1 (SAS Institute, Cary, North Carolina, USA).
Drugs
ST1936 and WAY-181187 were synthesised in the Sigma-Tau Chemistry Department (Pomezia, Italy). SB271046 was from Tocris (Bristol, UK), and 2-hydroxypropyl-ß-cyclodextrin and DA hydrochloride were from Sigma (St Louis, Missouri, USA).
Results
Effects of systemic administration of increasing cumulative doses of ST1936 on the basal firing rate of SNc DA-ergic neurons
Systemic administration of ST1936 (50-1600 μg/kg i.v.) had no effects on the basal firing activity of DA-ergic neurons in the SNc, compared to the control group (n=6; two-way ANOVA with one factor repetition; F1,11=0.06; p=0.8; n=7) (Figure 1).
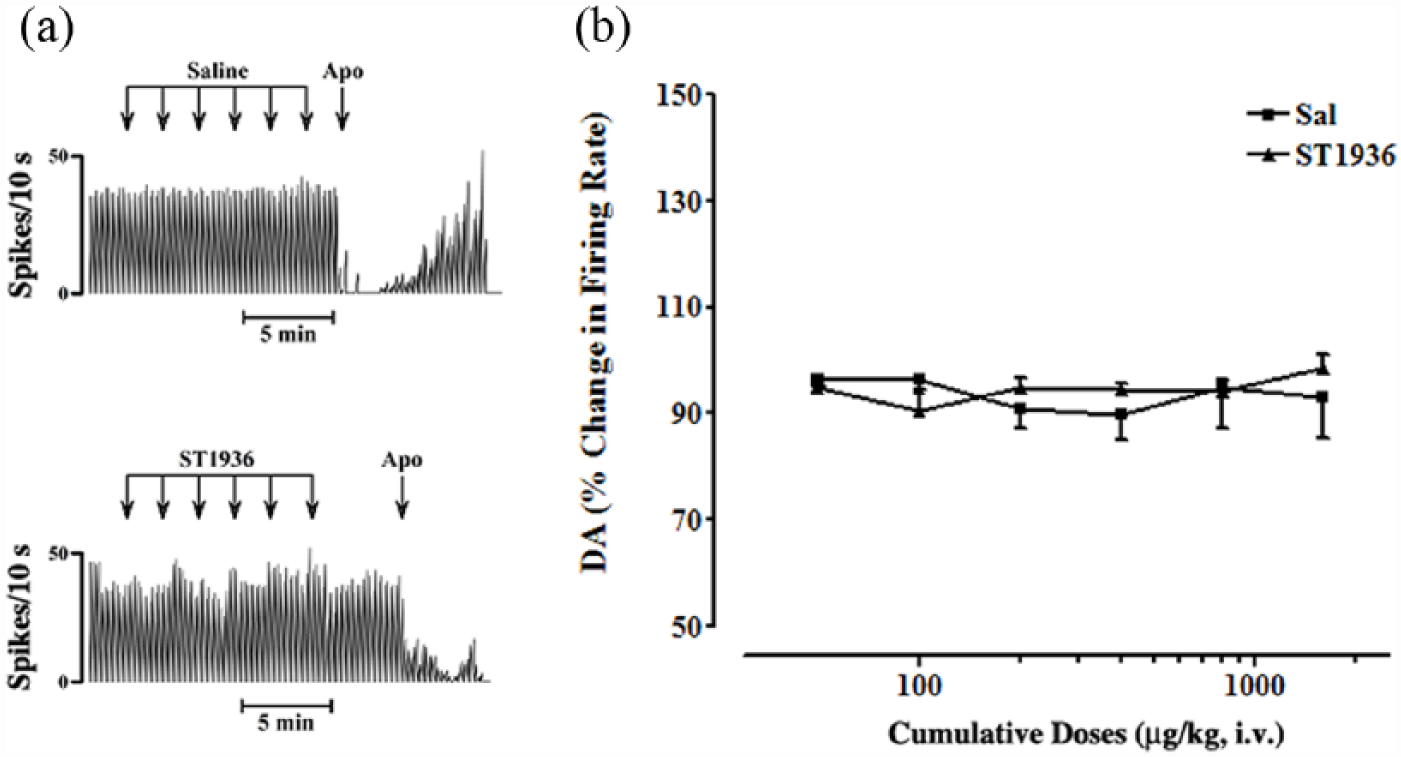
Effects of systemic administration of ST1936 on the basal firing rate of substantia nigra pars compacta (SNc) dopamine (DA)-ergic neurons. (a) Representative rate histograms showing the effects elicited by i.v. administration of saline (Sal) (control; arrows, 100 µL at each) and of cumulative doses of ST1936 (arrows, 50, 100, 200, 400, 800, 1600 µg/kg, at each). Apo, apomorphine (10–30 μg/kg i.v.). (b) Cumulative dose-response curves showing the mean percentage changes (±standard error of the mean (SEM)) in basal firing rate of SNc DA-ergic neurons, as elicited by administration of Sal and ST1936. Statistical analysis showed no significant difference between rats treated with Sal (control group, n=6) and with ST1936 (two-way analysis of variance (ANOVA) with one factor repetition; F1,11=0.06; p=0.8; n=7).
Effects of systemic administration of increasing cumulative doses of ST1936 on the basal firing rate of putative VTA DA-ergic neurons
The effects of systemic administration of ST1936 were recorded on a total of 20 putative DA-ergic neurons in the VTA. For the basal firing activity of these putative VTA DA-ergic neurons, as compared to controls (saline+vehicle; n=10; Figures 2 and 3), the systemic administration of increasing cumulative doses of ST1936 from 50–400 µg/kg (i.v.) induced either dose-related increases, in nine out of the 20 neurons (+48±6.5% at 400 µg/kg, i.v.; two-way ANOVA with one factor repetition; F1,16=138.71; p<0.0001; Figure 2), or dose-related decreases, in 11 out of the 20 neurons (–25±4.7% at 400 µg/kg, i.v.; two-way ANOVA with one factor repetition; F1,19=25.14; p<0.0001; Figure 3). This systemic administration of ST1936 did not result in any significant changes in the basal firing patterns (e.g. bursts) of these putative VTA DA-ergic neurons (data not shown). During these procedures, there were both the absence in many neurons of firing bursts and the presence of diverging baseline values and responses to the drugs used in the present study.
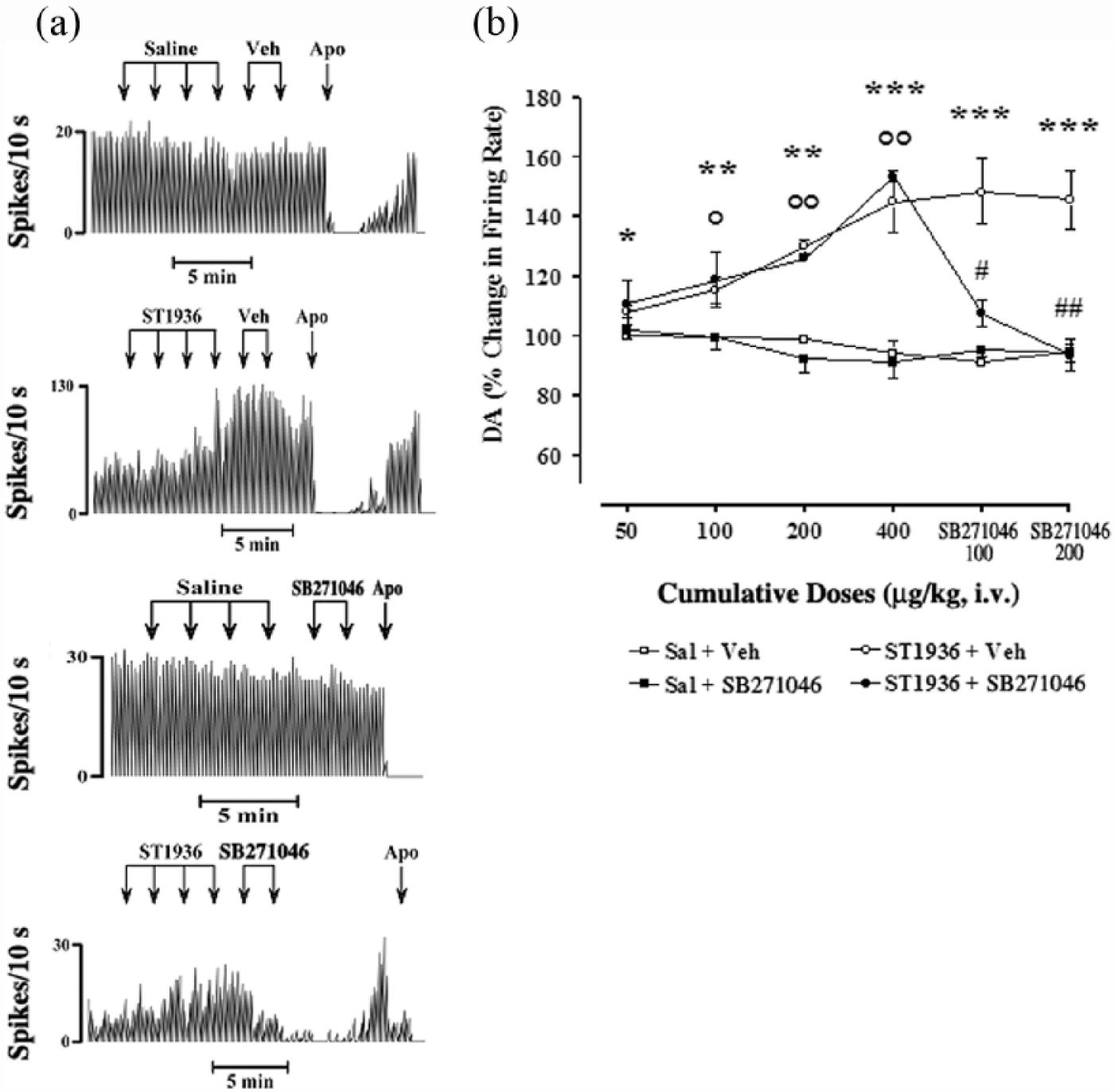
Effects of systemic administration of ST1936 and SB271046 on the basal firing rate of putative ventral tegmental area (VTA) dopamine (DA) neurons, where ST1936 induced a dose-related increase (in nine out of 20 neurons). (a) Representative rate histograms showing the effects elicited by i.v. administration of saline (Sal) and vehicle (Veh) of SB271046 (controls; arrows, 100 µL, at each) and of cumulative doses of ST1936 (arrows, 50, 100, 200, 400 µg/kg) and SB271046 (arrows, 100, 200 µg/kg). Apo: apomorphine (10–30 μg/kg i.v.). (b) Cumulative dose-response curves showing the mean percentage changes ±standard error of the mean (SEM) in basal firing rate of putative VTA DA neurons elicited by administration of the vehicles, ST1936 and SB271046. Statistical analysis showed a significant effect of ST1936 (50–400 µg/kg, i.v.) (two-way analysis of variance (ANOVA) with one factor repetition; F1,16=138.7; p<0.0001; n=9) compared with the control group (Sal+Veh and Sal+SB271046) (n=10) *p<0.05, **p<0.01, ***p<0.001, for ST1936+Veh vs control group (Fisher’s protected least significance difference (PLSD) post-hoc test). SB271046 (100, 200 µg/kg, i.v.) following Sal did not show any significant effects (two-way ANOVA with one factor repetition; F1,8=0.04; p=0.8; n=5, Sal+Veh vs Sal+SB271046). SB271046 (100, 200 µg/kg, i.v.) reversed the ST1936-induced increase in basal firing rate (two-way ANOVA with one factor repetition; F1,6=11.08; p<0.01; n=3. #p<0.05, ##p<0.01 ST1936+SB271046 vs ST1936+Veh); °p<0.01, °°p<0.001 ST1936+SB271046 vs Sal+SB271046 (Fisher’s PLSD post-hoc test).
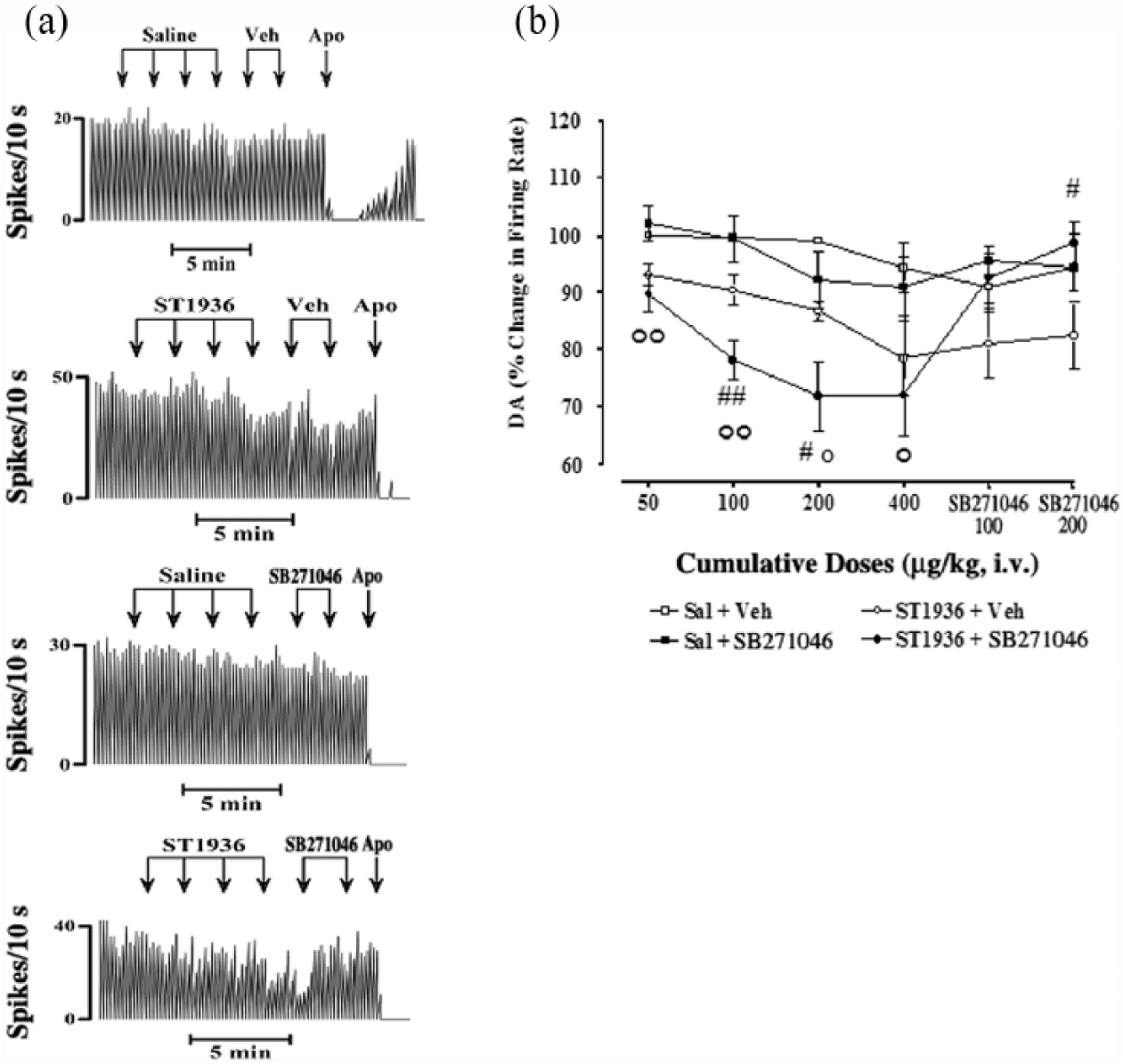
Effects of systemic administration of ST1936 and SB271046 on the basal firing rate of putative ventral tegmental area (VTA) dopamine (DA) neurons, where ST1936 induced a dose-related decrease (in 11 out of 20 neurons). (a) Representative rate histograms showing the effects elicited by i.v. administration of saline (Sal) and vehicle (Veh) of SB271046 (controls; arrows, 100 µL, at each) and of cumulative doses of ST1936 (arrows, 50, 100, 200, 400 µg/kg) and SB271046 (arrows, 100, 200 µg/kg). Apo: apomorphine (10–30 μg/kg i.v.). (b) Cumulative dose-response curves showing the mean percentage changes (±standard error of the mean (SEM)) in basal firing rate of putative VTA DA neurons elicited by administration of the vehicles, ST1936 and SB271046. Statistical analysis showed significant effects of ST1936 (50–400 µg/kg, i.v.) (two-way analysis of variance (ANOVA) with one factor repetition; F1,19=25.1; p<0.0001; n=11) compared with the control group (Sal+Veh and Sal+SB271046) (n=10). Moreover, only ST1936+SB271046 (n=6) reached statistical significance vs Sal+SB271046 (n=5) (two-way ANOVA with one factor repetition; F1,9=13.5; p<0.005) (°p<0.01, °°p<0.001 ST1936+SB271046 vs Sal+SB271046, Fisher’s PLSD post-hoc test). SB271046 (100, 200 µg/kg, i.v.) following Sal did not show any significant effects (two-way ANOVA with one factor repetition; F1,8=0.04; p=0.8; n=5, Sal+Veh vs Sal+SB271046). SB271046 (100, 200 µg/kg, i.v.) reversed the ST1936-induced decrease in basal firing rate (two-way ANOVA with one factor repetition; F1,9=3.7; p=0.08; n=6), which reached statistical significance only for 200 µg/kg SB271046 (#p<0.05, ST1936+SB271046 vs ST1936+Veh, Fisher’s PLSD post-hoc test). ST1936+SB271046 was statistically different vs ST1936+Veh at 100 and 200 µg/kg SB271046 (two-way ANOVA with one factor repetition; F1,9=4.04; p=0.07; #p<0.05, ##p<0.01, Fisher’s PLSD post-hoc test).
SB271046 blockade of the effects of systemic administration of ST1936 on the basal firing rate of putative VTA DA-ergic neurons
The potent and selective 5-HT6 receptor antagonist SB271046 was administered 2 min after administration of the increasing cumulative doses of ST1936 (50–400 µg/kg), to assess its specificity as a selective 5-HT6 agonist under our experimental conditions. SB271046 (100, 200 µg/kg, i.v.) showed no significant effects on the basal firing rate by itself (two-way ANOVA with one factor repetition; F1,8=0.04; p=0.8; n=5, saline+vehicle vs saline+SB271046). However, SB271046 (100, 200 µg/kg, i.v.) completely reversed the ST1936-induced increases in the basal firing rates (two-way ANOVA with one factor repetition; F1,6=11.08; p<0.01; n=3, ST1936+vehicle vs ST1936+SB271046, Fisher’s PLSD post-hoc test) (Figure 2), at all doses, and also significantly reversed the ST1936-induced decreases in basal firing rates (two-way ANOVA with one factor repetition; F1,9=3.7; p=0.08; n=6), which here reached statistical significance only for 200 µg/kg SB271046 (#p<0.05, ST1936+vehicle vs ST1936+SB271046, Fisher’s PLSD post-hoc test). It is important to note that the dose-response curves of ST1936+vehicle did not reach statistical significance F1,9=4.2; p=0.07; n=5 (two-way ANOVA with one factor repetition; Figure 3). The vehicle of SB271046 did not modify the firing rate induced by ST1936 (Figures 2, 3).
Effects of microiontophoretic application of ST1936 on the basal firing rate of VTA DA-ergic neurons
ST1936 was also ejected locally into the VTA. This iontophoresis of ST1936 produced a current-dependent increase (+5 to +15 nA; higher ejection currents produced disturbances in the electrical activity and loss of the VTA DA neurons) in the basal firing rates of these spontaneously active DA cells. Statistical analysis indicated significant current-dependent effects: +31±7.8% change in the basal firing rate at +5 nA (t=4; **p<0.0013); +41±8.7% at +10 nA (t=4.7; **p<0.0003) and +56±13% at +15 nA (t=4.3; **p<0.0007) (one sample Students’ t-tests; n=15; Figure 4).
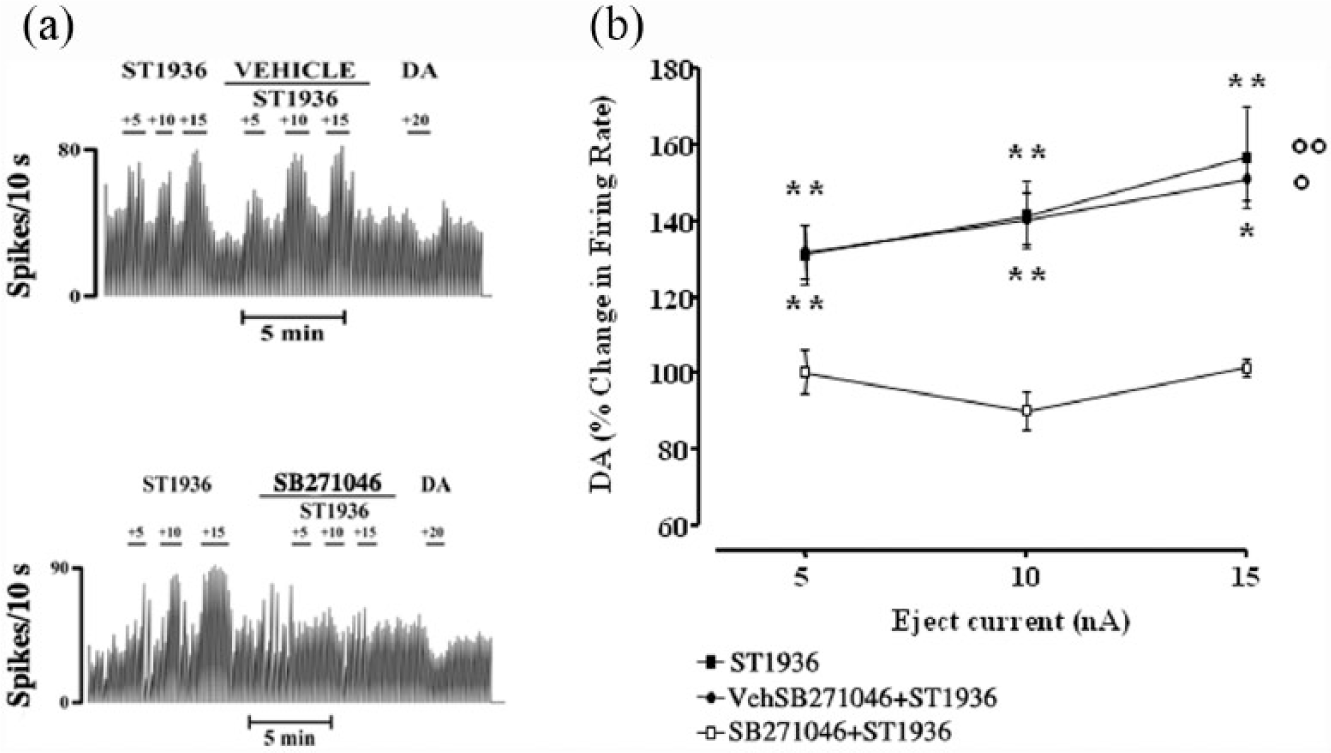
Effects of local application of ST1936 and SB271046 on the basal firing rate of putative ventral tegmental area (VTA) dopamine (DA)-ergic neurons. (a) Representative rate histograms showing excitatory effects elicited by microiontophoretically applied ST1936. Local co-application of SB271046 counteracted the excitatory effects elicited by microiontophoretically applied ST1936. Numbers above each short bar indicate the ejection currents, in nA. (b) Current-response curves showing the mean percentage changes (±standard error of the mean (SEM)) in the basal firing rate of the VTA DA neurons elicited by local administration of ST1936 alone (n=15) and with the vehicle (Veh) of SB271046 (n=4), and ST1936+SB271046 (n=4). Local application of the SB271046 serotonin-6 (5-HT6) antagonist significantly counteracted the excitatory effects on the basal firing rate of the VTA DA-ergic neurons following the microiontophoresis of ST1936 (treatment by ST1936, F2,20=5.6; p<0.01; n=4; two-way analysis of variance (ANOVA) with one factor repetition; °°p<0.003 ST1936 vs ST1936+SB271046, Fisher’s protected least significance difference (PLSD)). The Veh (10% dimethyl sulfoxide) used to dissolve SB271046 did not modify the effects of ST1936 alone (n=4; p=0.9; ST1936 vs ST1936+Veh; °p<0.02, Fisher’s PLSD).
Local application of SB271046 by micropressure ejection prevents the effects elicited by microiontophoretically applied ST1936 on the basal firing rate of VTA DA-ergic neurons
To determine the specificity of ST1936 as a selective 5-HT6 agonist under our experimental conditions, SB271046 (20 mM) was injected locally into the VTA by micropressure ejection, 1–2 min before and during ST1936 administration. This local application of the 5-HT6 antagonist SB271046 significantly counteracted the excitatory (treatment by ST1936, F2,20=5.6; p<0.01; n=4; two-way ANOVA with one factor repetition) effects exerted by microiontophoresis of ST1936 on the basal firing rate of these VTA DA-ergic neurons (Figure 4; °°p<0.003 ST1936 vs ST1936+SB271046, Fisher’s PLSD). The vehicle (here as 10% dimethyl sulfoxide; referred to as ‘vehicle’) used to dissolve SB271046 did not modify the effects of ST1936 (n=4; Figure 4; p=0.9, ST1936 vs ST1936+vehicle; °p<0.02, ST1936+SB271046 vs ST1936+vehicle, Fisher’s PLSD).
Effects of microiontophoretic application of ST1936 on the basal firing rate of VTA non-DA-ergic neurons
The iontophoresis of ST1936 had no effects on the basal firing rates of spontaneously active non-DA neurons in the VTA (t=0.48; p=0.6, one sample Students’ t-tests; Figure 5).
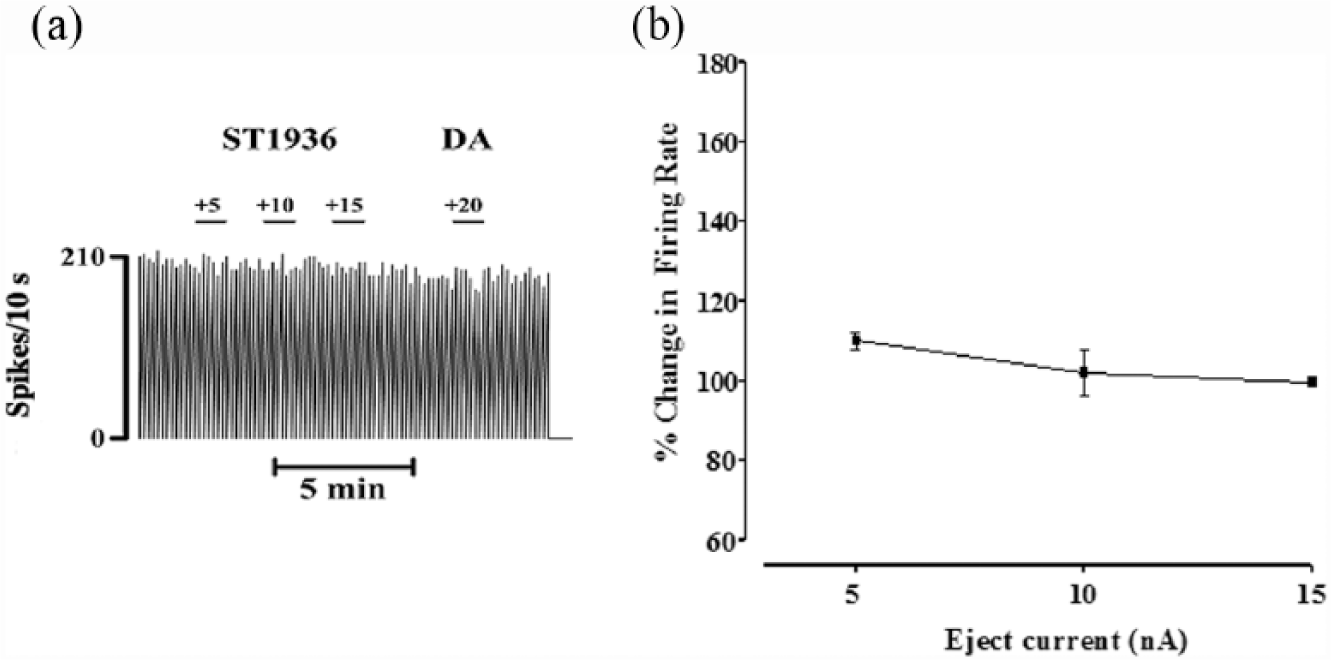
Effects of local application of ST1936 on the basal firing rate of ventral tegmental area (VTA) non-dopamine (DA)-ergic neurons. (a) Representative rate histograms showing the null effect of microiontophoretically applied ST1936. Numbers above each short bar indicate the ejection currents, in nA. (b) Current-response curves showing the mean percentage changes (±standard error of the mean (SEM)) in basal firing rates elicited by local administration of ST1936 (t=2.011; p=0.06, one sample Students’ t-tests).
Effects of systemic administration of increasing cumulative doses of WAY-181187 on the basal firing rate of VTA DA-ergic neurons
WAY-181187 is a new 5-HT6 receptor agonist, and it was administered systemically by the same route as ST1936, to compare the effects of these two specific 5-HT6 receptor agonists on these putative VTA DA-ergic neurons. Acute systemic treatment with WAY-181187 (50–1600 μg/kg, i.v.; n=5) induced dose-related inhibition of the basal firing activity of these DA neurons in the VTA (Figure 6). This reached a maximal effect at the cumulative dose of 1600 μg/kg, which reduced the activity of these putative DA-ergic neurons by 37.3%, with respect to the basal firing rate. At 400 μg/kg, both WAY-181187 (–27.7±5%) and ST1936 (–25±5%) showed similar effects. Statistical analysis revealed a significant effect of WAY-181187 (two-way ANOVA with one factor repetition; F1,7=10.94; p<0.013; n=5) compared to saline (control group, n=4; Figure 6).
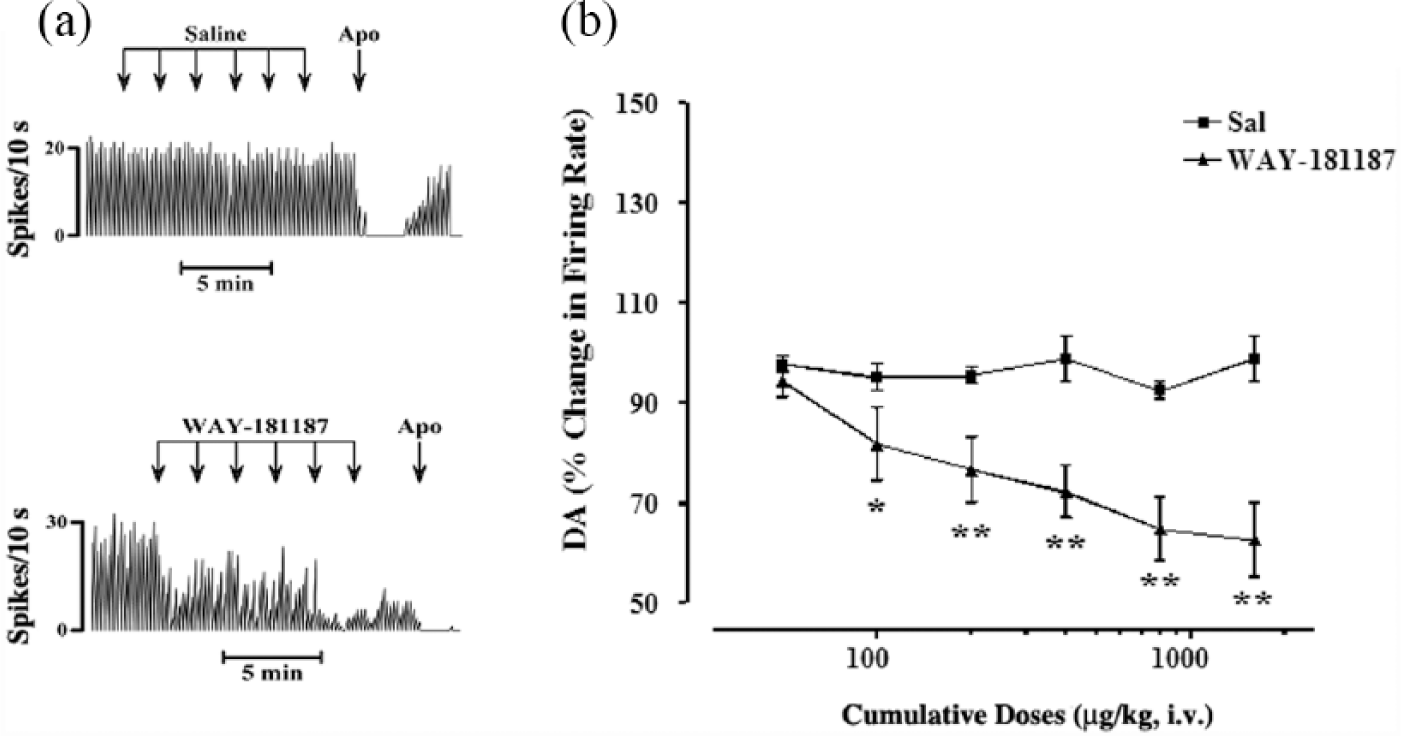
Effects of systemic administration of WAY-181187 on the basal firing rate of ventral tegmental area (VTA) dopamine (DA)-ergic neurons. (a) Representative rate histograms showing the effects elicited by i.v. administration of saline (Sal) (control; arrows, 100 µL, at each) and of cumulative doses of WAY-181187 (arrows, 50, 100, 200, 400, 800, 1600 µg/kg). Apo: apomorphine (10–30 μg/kg i.v.). (b) Cumulative dose-response curves showing the mean percentage changes (±standard error of the mean (SEM)) in basal firing rate of VTA DA-ergic neurons elicited by administration of WAY-181187. Statistical analysis shows significant inhibition of the basal firing rate after WAY-181187 (two-way analysis of variance (ANOVA) with one factor repetition; F1,5=7.8; p<0.0001; n=5), with respect to the control group (n=4); *p<0.05, **p<0.01, Fisher’s protected least significance difference (PLSD) post-hoc tests.
Discussion
The present study was undertaken with the aim to determine the role of 5-HT6 receptors in the regulation of neuronal function of the rat midbrain DA-ergic systems. Using in-vivo electrophysiological techniques, we have shown that acute intravenous administration of ST1936, a new 5-HT6 agonist (Riccioni et al., 2011), can either increase or decrease the basal firing rates of putative DA-containing neurons in the VTA of anaesthetised rats. Moreover, ST1936 did not affect the basal firing activity of DA-ergic neurons in the SNc. Previous studies also described no effects by 5-HT6 ligands in the nigrostriatal system (Dawson et al., 2000; 2001; Minabe et al., 2004; Schechter et al., 2008). ST1936 appears to be relatively selective for 5-HT6 receptors, although it has shown affinity also for 5-HT2B, 5-HT1A, 5-HT7 receptor and α-adrenergic receptors when tested in a broad cross-reactivity panel that comprised G-protein-coupled receptors, ion channel binding sites, enzymes, and transporters (Riccioni et al., 2011; Valentini et al., 2011). Nevertheless, the excitatory effects of ST1936 on these VTA putative DA-ergic neurons was completely reversed by treatment with SB271046, a potent 5-HT6 receptor antagonist, thus indicating an effect mediated by stimulation of 5-HT6 receptors. In agreement with this, systemic administration of ST1936 showed a dose-dependent increase in the levels of dialysate DA and noradrenaline in the nucleus accumbens shell and prefrontal cortex, and to a lesser extent in the nucleus accumbens core (Valentini et al., 2011). Furthermore, ST1936 was shown to induce conditioned place-preference and conditioned saccharin aversion, and to be self-administered i.v. by rats, which indicated the DA-stimulant properties of ST1936 in the nucleus accumbens shell (Valentini et al., 2011).
Only the higher dose of SB271046 significantly reversed the inhibitory effects of ST1936, which indicates partial involvement of 5-HT6 receptors in this action. A previous report showed that DA-ergic neurons in the VTA have decreased spontaneous activity after acute administration of SB271046 (Minabe et al., 2004), although that study used different doses and routes of SB271046 administration (1, 3 and 10 mg/kg, per os), compared to the doses of SB271046 used in the present study (100, 200 µg/kg, i.v.). Consistent with the view that midbrain DA cells are not homogeneous, there have been recent findings that showed that the specific molecular and physiological properties of midbrain DA cells are associated with the target structures into which they project (Lammel et al., 2008; Margolis et al., 2006, 2008). A subgroup of ‘unconventional’ DA neurons with a high frequency of firing (>10 Hz) and a low DA reuptake capacity (i.e. low DA transporter/ tyrosine hydroxylase expression ratio) is located in the medial posterior VTA, and these project into the medial prefrontal cortex, and the nucleus accumbens core and medial shell (Lammel et al., 2008). In contrast, ‘conventional’ DA neurons with a low frequency of firing (<10 Hz) are located in the lateral VTA and SNc, and these project into the nucleus accumbens lateral shell and the dorsal striatum, respectively (Lammel et al., 2008). In the present study, we observed that slow and fast firing DA neurons (2–9 Hz) share these non-uniform responses after systemic administration of ST1936. Therefore, we cannot correlate the different increase/decrease responses observed after this systemic administration of ST1936 with distinct populations of DA neurons in the VTA (e.g. parabrachial/paranigral). However, there is considerable confusion over the extent to which 5-HT6 receptors can affect DA transmission, and particularly cortical DA release. The picture of 5-HT6 receptors is more complex than expected, as their interactions with 5-HT6 ligands might also depend on their neuroanatomical region. Thus, regional brain differences might also then account for some of the inconsistencies that have been seen in particular in microdialysis studies, as indicated in the Introduction.
WAY-181187 is a new 5-HT6 receptor agonist, and it produced significant dose-related decreases in the basal firing rates of these VTA DA-ergic neurons in the present study, as seen previously for cortical DA and 5-HT levels (Schechter et al., 2008). Indeed, mesencephalic DA neurons that project into the limbic and cortical areas have a rich and complex organisation, with subcomponents sparsely located not only in the VTA, but also in the dorsal tier of the SNc (Björklund and Dunnett, 2007). As the release of DA derives from a set of synaptic terminals around the dialysis membrane, the differences in DA release seen in the medial prefrontal cortex and the nucleus accumbens by Valentini and colleagues (2011) might be due to the combined firing activity of cells after stimulation of 5-HT6 receptors by systemic ST1936 not only in the VTA, but also in other brain compartments. Interestingly, two subpopulations of VTA DA principal neurons with different firing patterns and frequencies have been reported to have different localisations into the VTA and distinct projections into the prefrontal cortex and nucleus accumbens (Chiodo et al., 1984; Koyama et al., 2005; Neuhoff et al., 2002). In this respect, in the present study, systemic administration of WAY-181187 resulted in a decrease in the basal firing rate of putative VTA DA-ergic neurons, whereas ST1936 showed both inhibitory and excitatory effects. These differences might be due to different pharmacological mechanisms of ST1936 and WAY-181187. Indeed, Riccioni et al. (2011) recently showed that ST1936 activates both cAMP and Ca2+ responses in HEK/HA–5-HT6 receptor cells transfected with Gα15. In contrast, WAY-181187 is a full agonist for the cAMP response, but only a partial agonist of the Ca2+ response (Codony et al., 2010), as also seen in a GαS-based assay (Dupuis et al., 2008).
Local administration of ST1936 in the VTA of anaesthetised rats elicited a current-dependent increase in the basal firing rate of the VTA DA neurons through direct stimulation of the 5-HT6 receptors present on these DA cells, as demonstrated by the concomitant administration of SB271046 that strongly reversed the excitatory activity of ST1936 on these VTA DA-ergic cells. Moreover, ST1936 was devoid of any effects on VTA non-DA neurons. The mechanism by which this systemic administration of ST1936 can elicit either stimulatory or inhibitory effects on these putative VTA DA cells is still unclear. Indirect effects mediated by cAMP or Ca2+ release, or interactions with other cell types, might explain these differences. On the other hand, a possible role of GABA extra-VTA interneurons should be considered in this study for these inhibitory versus direct excitatory effects of ST1936. Indeed, neurochemical and localisation studies have clearly demonstrated a central role for the 5-HT6 receptor in the modulation of GABA-ergic neurotransmission across a wide range of brain structures and networks (Dawson, 2011). Moreover, Schechter et al. (2008) observed dose-related increases in extracellular levels of GABA in several brain areas of the rat after administration of WAY-181187. Indeed, WAY-181187 also enhanced inhibitory postsynaptic currents in the hippocampus of rats, which represents a direct correlate of enhanced GABA-ergic function (West et al., 2009). However, the inhibitory effects seen after systemic ST1936 might also be due to a decrease in the excitatory input to the VTA induced by 5-HT6 from corticostriatal glutamatergic transmission via GABA-ergic extra-VTA neurons (Tassone et al., 2011).
As the local application of ST1936 increases this basal firing activity, while its systemic application either increased or decreased it, this suggests that the increase in the basal firing rate that was obtained following systemic administration of ST1936 should have acted on same site that was activated locally, and it is possible that this is the effect of 5-HT6 receptor agonists on DA neurons. In the case of the decrease in the basal firing rate, the systemic administration of ST1936 might have acted on different sites (i.e. other than local sites) through the GABA neurons that control these DA neurons.
Some previous studies have shown that 5-HT6 agonists can increase DA and noradrenaline levels in postsynaptic areas, while other studies have shown an enhancement of DA by 5-HT6 antagonists, as previous discussed. In conclusion, this study shows that activation of 5-HT6 receptors by the new full agonist ST1936 has either indirect, and less marked and selective, inhibitory effects, or direct selective excitatory dose-related effects on the basal firing rates of these putative VTA DA-ergic neurons, without affecting their firing pattern (e.g. bursts). As disturbances in mesocorticolimbic DA function are involved in the pathophysiology of schizophrenia and depression (Brown and Gershon, 1993; Fibiger, 1995), and as 5-HT6 antagonists might have a role in enhancing cognition (Meffre et al., 2012), these data open new possibilities for the use of 5-HT6 receptor ligands as therapeutic agents for these diseases. However, a lot of work remains to be done before a full understanding of the role of 5-HT6 receptors is obtained. In this regard, it would also be relevant to test ST1936 in chronic studies.
Footnotes
Acknowledgements
This paper is dedicated to Ennio Esposito, a distinguished scientist and expert in neurophysiology and neurobiology, who is well known for his studies on serotoninergic and dopaminergic receptors. Ennio Esposito died on 23 October 2011. The authors gratefully acknowledge Christopher Berrie for English revision of the manuscript, and Massimo Pierucci and Elena Fontana for the figures.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by Sigma-Tau, Industrie Farmaceutiche Riunite SpA, Pomezia, Italy, and that except for income received by their primary employer, no compensation was received for this study.