
Abstract
Recent evidence supports the value of endophenotypes and genome-wide association studies in psychiatric genetics, and their importance for dissecting the neural pathways and molecular mechanisms of complex neuropsychiatric disorders. Continuing this important discussion, here we outline three new mechanisms by which novel classes of genes may facilitate CNS pathogenesis without directly worsening its individual ‘established’ endophenotypes. These putative genetic mechanisms can apply to other human disorders in general, and may also be used for designing novel effective CNS drug treatments.
Keywords
Introduction: endophenotypes and brain disorders
Representing complex and polygenic maladies, human neuropsychiatric disorders have a multifactorial nature and poorly understood pathobiology (Kalueff et al., 2014; Nestler and Hyman, 2010; Tsankova et al., 2007). Their genetic architectonics also remains unclear (Fears et al., 2014; Gaugler et al., 2014; Harrison, 2015). Addressing this complexity 40 years ago, Gottesman and Shields introduced the concept of endophenotypes (Glahn et al., 2014; Gould and Gottesman, 2006)—measurable and heritable behavioral or physiological biomarkers positioned between the genes and the phenotypes/syndromes (Figure 1). Endophenotypes are conceptually similar, but not identical, to “intermediate phenotypes”, which represent biological phenotypes that are closer to the molecular effects of risk genes than the clinical symptoms; see detailed discussion in Rasetti and Weinberger (2011).
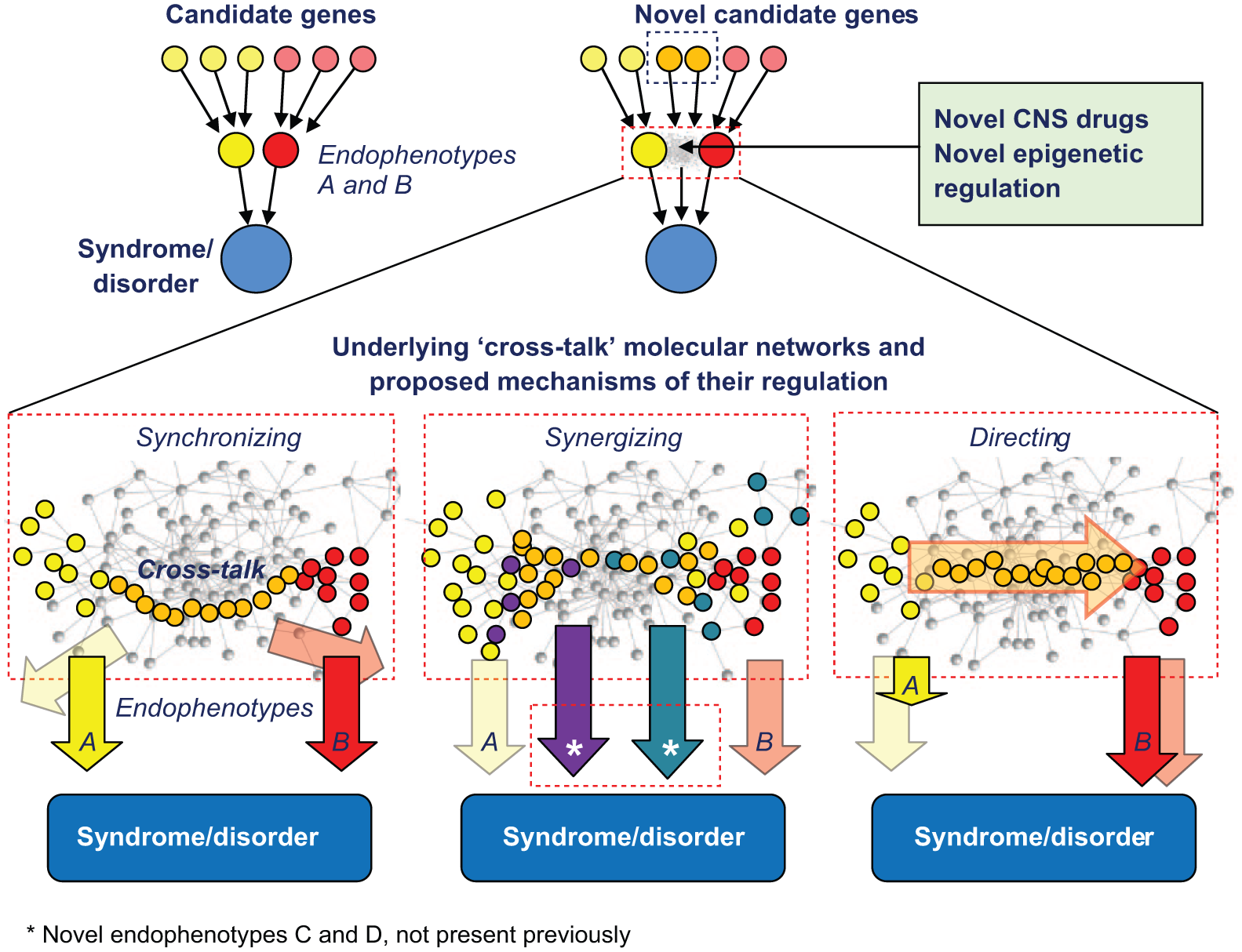
Potential novel genetic mechanisms of brain disorders (see text for details). “Synchronizing” mechanisms, encoded by specific subsets of novel putative central nervous system (CNS) genes, can temporarily organize/align several disordered endophenotypes (e.g. A and B) by synchronizing their underlying molecular pathways, collectively rendering the CNS less resistant to a coordinated set of simultaneous pathogenic cascades. “Synergizing” mechanisms, encoded by another subset of putative novel CNS genes, can have an additive effect on disease pathogenesis, triggering the emergence of principally new (previously not expressed) endophenotypes, C and D (*), without worsening the original endophenotypes, A and B. Finally, “directing” mechanisms of genetic regulation, controlled by their own specific putative genes, may guide the phenotypic transitions within the common pathogenetic process (e.g. causing shifts in anxiety with low mood > anxious depression > major depression), thereby changing the current dynamic combination of disordered endophenotypes (A > B) worsening them individually per se.
While some studies question the utility of endophenotypes (e.g. (Iacono et al., 2014)), such analyses appear to be underpowered for gene discovery, and focus on a set of specific (e.g. electrophysiological) measures without drawing on all conventional, well-studied endophenotypes for neuropsychiatric illness. In contrast, numerous other studies have successfully applied endophenotype strategies to implicate candidate disordered genes and/or central nervous system (CNS) pathways—in both clinical studies (Blasi et al., 2013; Dick et al., 2006; Glahn et al., 2012; Glahn et al., 2014; Hasler and Northoff, 2011; Hibar et al., 2015; Waldman, 2005) and pre-clinical animal models (de Mooij-van Malsen et al., 2015; Heinzmann et al., 2014; Kalueff et al., 2010). The value of endophenotype-based analyses has also been recognized as an essential part of today’s neuroinformatics, computational psychiatry (Wang and Krystal, 2014), neuroimaging (Hibar et al., 2015; Lisiecka et al., 2015; Mathew et al., 2014; Rasetti and Weinberger, 2011), cognitive neuroscience (Riesel et al., 2015; Scoriels et al., 2015), psychiatric genetics (Hibar et al., 2015), and other areas of translational biological psychiatry (Braff, 2015).
Genome-wide association studies (GWAS) represent a powerful tool for gene discovery (Nuechterlein et al., 2015). Exploring endophenotypes and GWAS in psychiatric genetics, recent evidence demonstrates their importance for dissecting neural molecular mechanisms of complex behavioral disorders (Flint and Munafo, 2013, 2014), as well as for identifying and validating novel drug targets (Braff, 2015; Harrison, 2015). However, the goal of psychiatric genetics—to link candidate genes to disordered phenotypes—is a challenging task, because complex neuropsychiatric disorders are polygenic and typically represent clusters of pathogenetically overlapping/interlinked endophenotypes (Figure 1) (Kalueff et al., 2014, 2015a). Furthermore, some of the “hidden/missing” heritability may be due to genetic variance (e.g. tandem repeat polymorphisms) not captured by GWAS that assesses single-nucleotide polymorphisms (SNP) (Hannan, 2010). The genetics (heritability) of complex syndromes may also be stronger than genetic associations of its individual endophenotypes (Farmer et al., 1987), raising the possibility of additional genetic mechanisms that may underlie complex CNS traits.
Continuing this important discussion, here we outline three new mechanisms by which novel putative “interlinking” (“interplay”) genes may facilitate the progression of neuropsychiatric diseases without directly worsening their individual “established” endophenotypes. Summarized in Figure 1, these putative mechanisms may explain how a substantial portion of genetic factors of CNS disorders, remaining undetected by traditional GWAS approaches, can contribute to disease pathogenesis, its risks, and therapies.
Targeting neuropsychiatric spectra
Our present approach simultaneously applies two key paradigms—systems neuroscience and neuropsychiatric spectra (Adam, 2013)—to the clinically relevant overlap between brain disorders (Consortium, 2013; Lee et al., 2013). Complementing the traditional views on the genetics of complex behavior, we have recently postulated novel CNS genes that may regulate pathogenetic linkages among endophenotypes by integrating their molecular pathways (Kalueff et al. 2014). A principal distinction of these putative genes (vs traditional candidate genes) is in their anticipated strategic “hub” location at the intersection of endophenotype-specific molecular networks, and the lack of direct effects on (and association with) traditional established endophenotypes (Kalueff et al., 2014, 2015b) (Figure 1). Thus, while contributing to overall CNS pathogenesis by facilitating the cross-talk between disordered endophenotypes, such novel genes are unassociated with ordinary endophenotypes, thereby remaining undetectable by conventional GWAS analyses (Kalueff et al., 2014, 2015a), and even potentially contributing to their reported failures (e.g. Iacono et al., 2014). Developing this concept further, here we summarize different mechanisms by which such novel putative class of CNS genes may facilitate the neuropsychiatric disease progression without directly affecting its individual endophenotypes.
Potential novel mechanisms of genetic regulation
One of the simplest potential mechanisms of endophenotype interplay may involve novel genes responsible for synchronizing several disordered endophenotypes (Figure 1). Such synchronizing, without enhancing pathological behaviors per se, may render the CNS less resilient by challenging it via different mechanisms simultaneously, thereby escalating the overall disorder progression and reducing brain resilience. For example, simultaneously-occurring social deficits and repetitive behaviors would make autism pathogenesis and its clinical picture objectively worse, compared to clinical forms of autism, where social deficits and behavioral perseverations of the same amplitude occur at different times/phases (i.e. with a lesser “synchrony”). The same logic may be applied to any other psychiatric disorder which includes more than one disordered domain.
Another potential mechanism of CNS complex genetic regulation may involve synergizing several disordered endophenotypes, where one pathogenetic pathway could reciprocally increase the probability of another disordered process (Figure 1). Collectively, this may worsen the neuropsychiatric disorder outcome by inducing principally novel negative effects (e.g. causing treatment resistance, more frequent recurrence, and/or comorbidities not present previously). An example of such interactions may be the possible link between low locomotor activity, low mood, anhedonia, and suicidal ideation in depressed patients. For example, drug-evoked reduction of motor retardation (or hypoactivity, a common endophenotype of depression) during antidepressant therapy may result in overall energizing of such patients, thereby triggering their suicidal behavior (otherwise suppressed by low energy/activity) (Leon et al., 2014; Morrison and Schwartz, 2014). Thus, such objective reduction of some disordered endophenotypes may, in principle, lead to the emergence of principally new symptoms in patients. In other words, although the original aberrant behaviors per se may not get worse, their combination can now have an additive effect and trigger novel pathogenic processes and/or vulnerabilities not overtly expressed previously. Clearly, further clinical attention to this putative “synergizing” mechanism and its genetics may generate more evidence of such type of modulation of neuropsychiatric pathogenesis.
Finally, the third potential novel mechanism of genetic regulation of complex CNS traits may include pathways that specifically direct the overall trajectory of the neuropsychiatric disorder progression from one endophenotype (e.g. anxiety) to another (e.g. depressed/low mood, Figure 1). Such trajectories have already been well-documented in longitudinal clinical studies (Merikangas et al., 2003; Purcell et al., 2015), and their further genetic parsing is important (also see discussion of distinct pathogenic trajectories of depression-like states in animal models in Kalueff et al. (2015b)). Here, we recognize that activation of novel putative “interplay” pathways may trigger phenotypic shifts (A > B) within the existing disorder. By leading to gradual replacement of one endophenotype with another, such mechanisms may eventually lead to a more serious, treatment-resistant, and debilitating syndrome. Again, genetic determinants underlying such “directing” mechanisms would mostly remain invisible to GWAS because they are unassociated with the severity of endophenotypes (e.g. final clinical manifestations) of the disorder.
Furthermore, the role of environmental and epigenetic factors in CNS pathogenesis is widely recognized (Caspi and Moffitt, 2006; Tsankova et al., 2007). Therefore, it is possible to expect complex gene×environment interactions and epigenetic regulation, not only for “traditional” endophenotype-associated genes, but also for the proposed novel genetic mechanisms (Figure 1) that specifically modulate the endophenotypes’ interplay as a core part of the disease development.
Concluding remarks
In general, mounting clinical and preclinical evidence supports the important role of “hub” genes and the endophenotypes’ interplay in CNS pathogenesis; see (Kalueff et al., 2015b) for a recent comprehensive review. For example, aberrant neural synchrony has been implicated in a schizophrenia-related rodent behavioral model (Lee et al., 2014), raising the possibility that the synchronization of endophenotypes may itself be one pathogenetic mechanism in the brain, with its own unique genetic determinants (note that electrophysiological synchrony is used here as a mechanic analogy, rather than as a direct example of a broader “brain endophenoytes’ synchrony” concept proposed here). Likewise, autism susceptibility candidate 2 (AUTS2) gene, while implicated in autism spectrum disorder, stimulates other brain genes involved in neurodevelopment (Gao et al., 2014), thereby suggesting major modulation of other CNS disorders, including those not directly linked to autism.
Examining neuropsychiatric spectra, the complexity of genetic dissection of anxiety comorbid with depression has recently been emphasized (Flint et al., 2014). For instance, mapping depression and anxiety in the same subjects has been correctly suggested to distinguish genetic loci that contribute specifically to one or the other disorder, and also identify loci that affect both (Flint et al., 2014). Likewise, discussing the complex genetics of schizophrenia and the criteria relevant to prioritizing drug targets based on genetic findings, the most important criterion listed recently is “the gene contains a causal variant unequivocally associated with the disorder” (Harrison, 2015). Here, we suggest that this may not always be the case, and that disordered genes may not always be directly linked to the disorder. The latter two examples are particularly important to illustrate the point we are making. Although the proposed endophenotype-centered analysis may reveal important information about the genetics of the two affective disorders, it, by default, would still miss critical disorder-related “interlinking” genes. In other words, genes responsible for synchronizing, synergizing, and/or directing the anxiety–depression pathogenic trajectory would remain undetectable by GWAS because they, by definition, do not worsen the traditional “anxiety” or “depression” endophenotypes.
Complementing these approaches, and logically drawing from Aristotle’s famous notion that “the whole is more than the sum of its parts”, we argue for more comprehensive genetic analyses that widen the spectrum of genetic mechanisms (Figure 1) potentially implicated in the regulation of complex CNS traits (Kalueff et al., 2015b). For example, applied to complex neuropsychiatric disorders, recent sophisticated omics-based analyses of clinical samples (Chang et al., 2015; Li et al., 2014) and preclinical data (Heinzmann et al., 2014; Malki et al., 2013, 2014) may become a particularly powerful tool in dissecting such additional layers of their genetic architectonics. Specifically, focusing on networks of molecules interacting with protein products of known/established endophenotype-specific genes, may help “connect the dots” and reveal novel cross-talk mechanisms (Kalueff et al., 2014). As these cross-talk pathways are likely to be encoded by “interlinking” genes (Figure 1), such omics-based bionetwork analyses can help identify these novel putative classes of genes. Accordingly, once novel cross-talk genes and their specific molecular pathways are established, novel drugs may be discovered by targeting the proposed synchronizing, synergizing, and directing mechanisms (Figure 1) of genetic regulation of complex CNS traits. Furthermore, it is also possible that the same “interlinking” genes may affect several proposed mechanisms in combination (e.g. simultaneously synchronizing and directing), thereby adding another layer of complexity to genetic regulation of CNS traits and the search for novel therapeutic drug targets.
Notably, as the “synchronizing” and “synergizing” mechanisms proposed here may at first glance appear similar to the well-established pleiotropic and epistatic genetic interactions (Kalueff et al., 2015b), this aspect merits further scrutiny. We posit that the proposed regulation via the “interlinking” genes differs markedly from pleiotropy and epistasis, because such novel putative genes do not affect disordered endophenotypes per se (Kalueff et al., 2015b). For example, they cannot make anhedonia and low mood worse, but can make them better correlated and integrated, thereby rendering the entire depressive pathogenesis more severe, better orchestrated, and more debilitating (Kalueff et al., 2015b). Pleiotropy occurs when a candidate gene causes multiple deficits in various systems, simultaneously affecting several, seemingly unrelated neural endophenotypes (Kalueff et al., 2015b). While pleotropic genes exert shared influence on several endophenotypes, the proposed interlinking genes are associated not with endophenotypes per se, but with the interconnectedness (pathogenetic linkage) between them (Kalueff et al., 2015b). Likewise, epistasis represents gene×gene (G×G) interactions, when the effects of one gene depend on the presence of one or more other genes (Kalueff et al., 2015b). Because epistatic G×G interactions directly affect the traditional endophenotypes, they would differ from the proposed “interlinking” genes, which are not associated with endophenotypes, and only regulate their overall pathogenetic overlap (Kalueff et al., 2015b). Again, while pleoitropic (and, eventually, epistatic) genes may possibly be detected by GWAS, the proposed synchronizing and synergizing “interlinking” genes would remain insensitive to such genetic analyses, therefore meriting further investigation.
Finally, from the systems biology standpoint, as proposed recently (Kalueff et al., 2015a), the disease-promoting “bad” interlinking genes (Figure 1) may be countered by another class of putative “interlinking” CNS genes (termed “angel genes”). We suggest that these putative genes may exert opposite, protective effects by activating (via similar cross-talk mechanisms) the brain resilience pathways (Kalueff et al., 2015a). Thus, such “angel” CNS interlinking genes may reduce the progression of the disease, not by directly inhibiting disordered pathways and symptoms, but indirectly—either by uncoupling the overlap among these pathogenic pathways (Figure 1), or by mobilizing/integrating the brain resilience pathways (Kalueff et al., 2015a). We posit that activation of CNS resilience pathways by such putative “angel” genes may be mediated in a similar manner (i.e. by synchronizing, synergizing, and directing), as discussed above for their “bad” interplay counterparts (Kalueff et al., 2015a) (Figure 2). In addition, a separate subset of “angel” genes (tentatively called “cherubim genes”) may also reduce pathogenesis via a different putative mechanism—by specifically inhibiting the activity of pathogenic networks of “bad” interlinking genes (Mechanism 3 in Figure 2) (Kalueff et al., 2015a, 2015b). Again, it is possible that some interlinking “angel” genes may simultaneously modulate several different mechanisms proposed here, further contributing to the growing complexity of genetics of CNS traits. Respectively, novel drugs to treat CNS disorders can be developed based, not only on inhibiting putative pathogenic interlinking genes (Kalueff et al., 2014), but also by activating the protective “angel” interplay genes and their respective biomolecular networks (Figure 2) (Kalueff et al., 2015a).
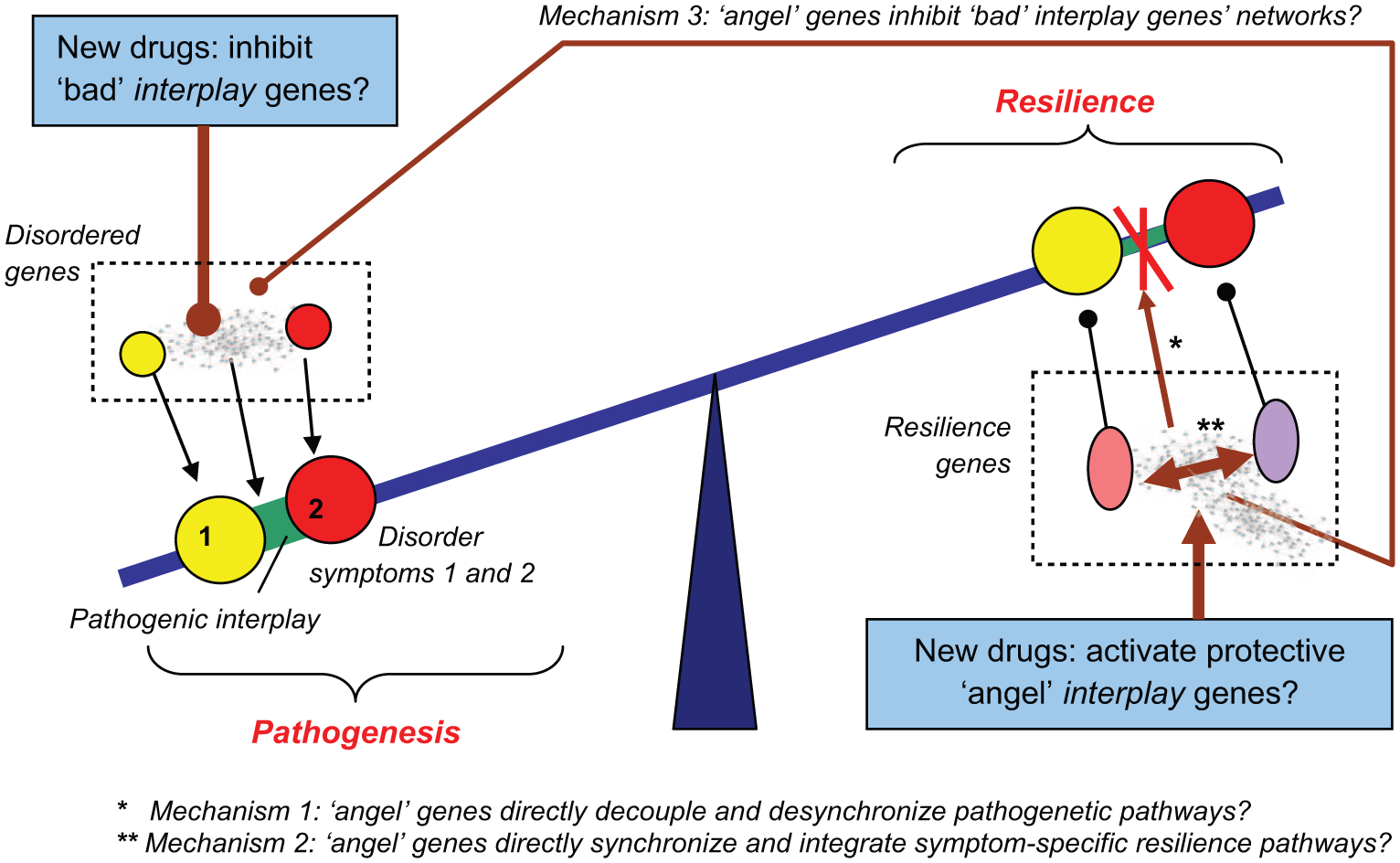
Potential complex genetic and epigenetic regulation of brain disorders bidirectionally controlled by proposed “bad” disease-contributing interlinking (interplay) genes, and putative protective “angel” interplay genes that modulate CNS resilience pathways. Note that “angel” genes may directly decouple/disintegrate disordered pathways (Mechanism 1) or activate specific cross-talk mechanisms, “synchronizing”, “synergizing”, and “directing” of CNS pathways underlying brain resilience (Mechanism 2). In addition, a separate sub-class of “angel” interplay genes (“cherubim genes”) may exist to specifically inhibit the activity of networks of “bad” interplay genes discussed in Figure 1 (Mechanism 3). Sharp arrows denote activation, dull arrows denote inhibition.
Moreover, there are no reasons to limit these novel proposed pathogenic and resilience genetic mechanisms to CNS disorders only, and the same basic principles may apply to many other human disorders in general. As already mentioned, we also recognize the important role of epigenetic regulation of brain genes (de Mooij-van Malsen et al., 2015; Petronis, 2010; Tsankova et al., 2007). Therefore, it is possible that epigenetic regulation of such novel putative “interlinking” genes can also be critical for mechanisms discussed here (Figure 2).
Footnotes
Acknowledgements
The authors thank Professor Irving I. Gottesman (University of Minnesota, USA) for his comments on this manuscript, and for sharing many valuable insights into endophenotypes and their developing utility in psychiatric genetics. We also thank Drs Anthony Hannan and David Goldman for their helpful comments and feedback.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study is supported by the ZENEREI Institute (AVK and AMS) and the Guangdong Ocean University (AVK, CS) Intramural Research Programs.