
Abstract
A recent hypothesis, based on electrophysiological and behavioural findings, suggests that ethanol simultaneously exerts opposed effects on the activity of dopamine (DA) neurons in the ventral tegmental area (VTA) through two parallel mechanisms, one promoting and the other reducing the GABA release onto VTA DA neurons. In this sense, the activating effects are mediated by salsolinol, a metabolite of ethanol, acting on the μ-opioid receptors (MORs) located in VTA GABA neurons. The inhibitory effects are, however, triggered by the non-metabolized fraction of ethanol which would cause the GABAA receptors-mediated inhibition of VTA DA neurons. Since both trends tend to offset each other, only the use of appropriate pharmacological tools allows analysis of this phenomenon in depth. Herein, we present new behavioural findings supporting this hypothesis. Motor activity was evaluated in rats after intra-VTA administration of ethanol 35 nmol, an apparently ineffective dose, 24 h after the irreversible blockade of MORs in the VTA with β-FNA. Our results showed that this pre-treatment turned the initially ineffective ethanol dose into a depressant one, confirming that the activating effect of ethanol can be selectively suppressed without affecting the depressant effects mediated by the non-biotransformed fraction of ethanol.
Introduction
The effect of ethanol on the activity of dopamine (DA) neurons in the posterior ventral tegmental area (VTA) seems to be complex and difficult to predict. Recent behavioural and electrophysiological evidence has allowed us to better understand factors involved in the mechanisms by which ethanol modifies the activity of VTA DA neurons. An interesting working hypothesis, proposed in our previous paper, is that ethanol dually modulates the activity of the VTA DA neurons. Following this idea, ethanol simultaneously exerts opposing effects on the activity of DA neurons through two parallel mechanisms, one promoting and the other reducing GABA release onto VTA DA neurons (Marti-Prats et al., 2013). Although both effects derive from the administered ethanol, different compounds are, in fact, ultimately responsible for these responses.
The underlying mechanism involved in the reduction of GABA release and, consequently, the activation of VTA DA neurons, points to the activation of the μ-opioid receptors (MORs). The involvement of MORs in alcohol effects was explored long ago and has been recognized (see Nutt, 2014 for a recent review). But there is some controversy on the mechanism by which the activation of MORs is produced after ethanol administration. A hypothesis that has received significant experimental support in recent years is based on the assumption that the products resulting from local ethanol metabolism directly interact with the MORs located in VTA GABA neurons. This mechanism is similar to that proposed by Johnson and North in their seminal work on the effects of opioids on the VTA DA neuron activity (Johnson and North, 1992). In the VTA, as in other parts of the brain (Hipolito et al., 2012), racemic (R and S) salsolinol is formed by non-enzymatic Pictet–Spengler condensation of DA with acetaldehyde (Haber et al., 1996; Melis et al., 2015; Rommelspacher et al., 1995), the main metabolite of ethanol. Salsolinol may disinhibit VTA dopamine neurons, via direct interaction with MORs located on VTA GABA neurons. Several findings support this assertion. First, the structure of salsolinol is similar to that of morphine (Davis and Walsh, 1970); second, salsolinol can bind to opioid receptors and produces opioid-like effects (Fertel et al., 1980; Lucchi et al., 1982); and, third, as MOR agonists do, salsolinol can increase both the nucleus accumbens DA extracellular levels and the motor activity of rats after intra-VTA administration, effects that can be blocked by MOR antagonists (Hipolito et al., 2010). Recent electrophysiological (Foddai et al., 2004; Melis et al., 2007, 2015; Xie et al., 2012), neurochemical (Deehan et al., 2012) and behavioural (Hipolito et al., 2010, 2011; Marti-Prats et al., 2010, 2013; Sanchez-Catalan et al., 2009) evidence has given robustness to that view. Concretely, our recent demonstration that salsolinol is able to dose-dependently and reversibly increase the ongoing firing of VTA DA neurons through MORs (Xie et al., 2012) can be considered an important step forward in demonstrating the direct interaction of salsolinol with these receptors.
On the other hand, the mechanism causing the increase of GABA release and, therefore, the inhibition of VTA DA neuron activity seems to depend on the non-metabolized fraction of ethanol, acting through GABAA receptors (Marti-Prats et al., 2013). Interestingly, electrophysiological experiments have shown that exposure of midbrain slices to ethanol in the presence of saturating concentrations of the MORs agonist DAMGO was accompanied by an increase in spontaneous inhibitory postsynaptic current frequency in VTA DA neurons (Xiao and Ye, 2008). This finding, obtained under specific pharmacological conditions, suggested the possible existence of a parallel mechanism through which ethanol would increase the GABA release from presynaptic terminals and, consequently, could reduce VTA DA neuron activity. This phenomenon was electrophysiologically confirmed and explored in more detail by another research group (Theile et al., 2008, 2009, 2011) and, recently, by our group using a behavioural approach (Marti-Prats et al., 2013). As we suggested, the non-metabolized fraction of ethanol could be responsible for this GABA release and, therefore, for the inhibition of VTA DA neurons.
The net effect produced by ethanol on the VTA DA neuron activity is clearly dependent on the administered dose of the drug. Hence, we have reported that the intra-VTA administration of ethanol 35 nmol does not modify the motor activity, whereas doses of 75 or 150 nmol produce an intense activation (Sanchez-Catalan et al., 2009). As hypothesized (Marti-Prats et al., 2013), the absence of effects on behaviour is not due to the lack of efficacy derived from very low local levels of ethanol, as at first glance it might be thought. On the contrary, as we demonstrated, even at this low ethanol dosage, both salsolinol and the non-metabolized fraction of ethanol are interacting in an efficient manner with their respective molecular targets; the absence of observable effects is due to the fact that their corresponding effects offset each other.
The use of appropriate pharmacological tools was crucial to enable us to analyse these two simultaneous and opposing tendencies in a detailed manner. Concretely, we employed catalase inhibitors (to reduce acetaldehyde formation and, therefore, salsolinol production) and acetaldehyde sequestering agents (to avoid the condensation of acetaldehyde with dopamine and, hence, the formation of salsolinol) to suppress selectively or diminish the activating effects of ethanol without suppressing the inhibitory effect mediated by the non-metabolized fraction of the drug. Nonetheless, and according to our hypothesis, an alternative strategy to selectively suppress the ethanol-derived activation of the VTA DA neurons is possible.
In the present study, this alternative strategy to suppress selectively the activating effect of ethanol without affecting the inhibitory action is explored. If our hypothesis on the dual effects of ethanol on VTA DA neurons (Marti-Prats et al., 2013) is correct, the intra-VTA administration of ethanol in animals subjected to local MOR blockade should lead to the depression of the activity of the VTA DA neurons and, therefore, to the reduction of animal motor activity.
Materials and methods
Animals
Male albino Wistar rats (300–340 g at the time of surgery) were used for our experiments (n=32). They were housed in plastic cages (48 × 38 × 21 cm3) in groups of four to six with controlled humidity and temperature (22°C), a 12:12-h light/dark cycle (on 08:00, off 20:00), and free access to food and water. All the procedures were carried out in strict accordance with the EEC Council Directive 86/609, Spanish law (RD 53/2013) and animal protection policies. Experiments were approved by the Animal Care Committee of Faculty of Pharmacy at Universitat de València, Spain.
Drugs and chemicals
Ethanol was purchased from Scharlau (Madrid, Spain). β-funaltrexamine (β-FNA; an irreversible antagonist of the μ-opioid receptors) was obtained from Tocris (Bristol, UK). Stock solutions of β-FNA were prepared by dissolving the compound in the correct volume of distilled water. Aliquots of these solutions were then kept frozen at −20°C until use. Prior to use, aliquots of the stock solutions were conveniently diluted with artificial cerebrospinal fluid (aCSF)/ascorbate solution to the appropriate concentration (Sanchez-Catalan et al., 2009). The aCSF/ascorbate solution consisted of 120.0 mM NaCl, 4.8 mM KCl, 1.2 mM KH2PO4, 1.2 mM MgSO4, 25.0 mM NaHCO3, 1.2 mM CaCl2, 100 mM D-glucose, and 0.2 mM ascorbate and pH was adjusted to 6.5. Ethanol was also freshly dissolved in aCSF solution prior to intra-VTA administration. All the other reagents used were of the highest commercially available grade.
Surgery and post-surgical care
Rats were anaesthetized with ketamine/xylazine (95 mg/kg of ketamine and 10 mg/kg of xylazine, ip) and placed in a stereotaxic apparatus (Stoelting, USA). An incision (8–10 mm) was made in the skin above the skull and the wound margin was infiltrated with lidocaine (3%). Three holes were drilled: two for the skull screws and the other for the guide cannulae (Plastics One, USA). Each animal was implanted unilaterally with one 28-gauge guide cannula aimed at 1.0 mm above the posterior VTA. The coordinates relating to bregma and skull surface (Paxinos and Watson, 2007) were as follows: A/P −6.0 mm; L ±1.9 mm; D/V −7.8 mm. Cannulae were angled towards the midline at 10° from the vertical (all the measurements in the dorsal-ventral plane refer to distances along the track at 10° from the vertical). Cannulae assemblies were secured in place with dental cement. A stainless steel stylet (33-gauge), extending 1.0 mm beyond the tip of the guide cannula, was put in place at the time of surgery and removed at the time of testing. Following surgery, rats were housed in individual rectangular plastic cages (47 × 22 × 15 cm3, located side by side in order to prevent the influence of chronic stress on performance due to isolation) with free access to food and water for at least 7 days. These cages were used as test cages on the day of the experiment.
Drug microinjection procedures
All the intra-VTA drug microinjections were carried out with 33-gauge stainless steel injectors, extending 1.0 mm below the tip of the guide cannulae. Injectors were attached to a 25 μL Hamilton syringe by using PE-10 tubing. Microinjections were carried out using a syringe pump (Kd Scientific) which was programmed to deliver a total volume of 200 nL in 20 s (flow rate of 0.6 μL/min) with the exception of the β-FNA injections in which the syringe pump was programmed to deliver a total volume of 300 nL in 2 min (flow rate of 0.15 μL/min). Following the infusion, the injector remained in place for 1 min to allow the diffusion of the drugs, and then it was removed, the stylet was replaced, and the locomotor activity was registered when appropriate. All the injections were carried out in the experimental room.
Handling and test procedure
Three days after surgery, animals were taken from the colony, brought to the experimental room, and handled for 5 min/day until the experimental day. During this phase, animals became accustomed to the experimenter, the experimental room, and to the injection procedure with a total of four to seven sessions to decrease the activational effects of the manipulations taking place during the injection process, as well as the novelty-activating effects of the testing room. Tests were performed 7–10 days after surgery. On the day of the experiment, rats were again taken from the colony room and brought to the experimental room 20–30 min prior to the start of the session, in the same rectangular cages in which the animals were housed. After this initial period, experiments started according to the protocol described in the drug microinjection procedures. All the experiments were recorded by a digital video camera and the distance travelled every 2-min was analysed until a total period of 20 min had been reached and for the total distance covered (both in cm) along the 20-min period using the Raddot program (Universitat de València, Spain). For the experiments, the experimental room was illuminated with soft white light.
Experimental design
The experiment was conveniently designed to determine whether the irreversible blockade of the μ-opioid receptors with β-FNA in VTA would be able to modify the response of animals to an intra-VTA injection of ethanol 35 nmol. This low dose of ethanol is unable to produce any significant change on the motor activity of naïve animals, that is, animals that have not previously received any pharmacological agent (Marti-Prats et al., 2013). Animals were randomly assigned to one of the four experimental groups used (n=8/group) that received two intra-VTA injections separated each by 24 h. The first injection (pre-treatment phase) consisted of the intra-VTA administration of aCSF or β-FNA (2.5 nmol). The second injection (treatment phase) consisted of the administration of aCSF or ethanol (35 nmol) and the subsequent analysis of the motor activity of the animals. Therefore, the experimental groups planned for this study were named: aCSF + aCSF, aCSF + ethanol, β-FNA + aCSF and β-FNA + ethanol.
In order to reduce to the minimum the number of rats used in our experiment and considering our previous experience in the intra-VTA injection of drugs, we decided not to include any additional experimental group devoted to analysing the site specificity for our drug treatments and pre-treatments.
Histology
At the end of the experiments, rats were anaesthetized using sodium pentobarbital. Brains were frozen and 40 µm-thick coronal sections were obtained using a cryostat. Sections were used for the verification of the cannulae placements of all the datasets presented in this study. To this end, coronal sections of the VTA area were stained according to the cresyl violet protocol. Then, the location of the cannula tips was carefully examined by a researcher, who was unaware of the experimental condition of the animals, using optical microscopy.
Statistical methods
A two-way ANOVA (2 × 2) with interaction was used to analyse the distance travelled by animals across the 20-min sessions. Factors analysed were: pre-treatment (aCSF or β-FNA) and treatment (aCSF or ethanol). Post-hoc comparisons were made using an adjusted Bonferroni’s test. At each of the 2-min intervals, distances travelled by animals were compared by means of one-way ANOVA, the experimental group being the between-groups factor analysed. The level of significance was always p=0.05. All the analyses were done using SPSS, v. 15.0.
Results
Of the 32 animals initially included in the experiment, 29 showed a correct position of the cannula, according to the current definition for the boundary between the anterior and posterior VTA. This limit is established around −5.5 mm from the bregma (Ikemoto, 2007; Sanchez-Catalan et al., 2014) (Figure 1(a)). Only two animals of the β-FNA + ethanol group and one of the β-FNA + aCSF group were rejected because of incorrect placements. The great majority of the injection sites (25 of 32) were centred at −5.88 to −6.72 mm posterior to the bregma, which corresponds to the posterior VTA. Data from four animals that showed injection sites at −5.64 mm, that is, slightly caudal to the −5.5 mm limit, were also included in our analysis. Six animals showing injection sites in the most caudal part of the VTA (from −6.36 to −6.72 mm), a region that overlaps with the anterior part of the tail of the VTA (tVTA) (Kaufling et al., 2009, 2010; Sanchez-Catalan et al., 2014), were also included in our analysis.
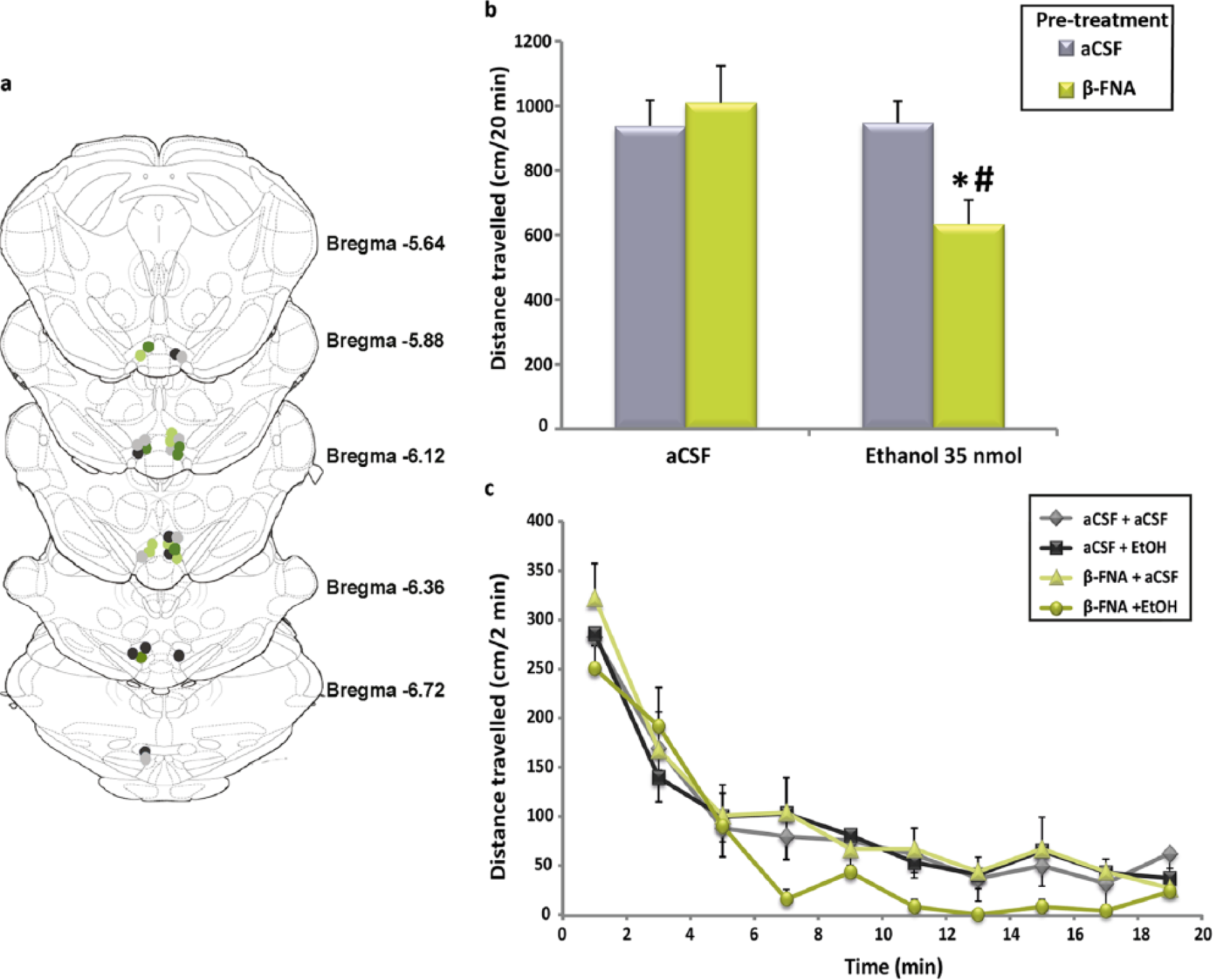
The mean values of the distance travelled by animals in each experimental group along 20 min obtained in this experiment are represented in Figure 1(b). As can be seen, animals pre-treated with β-FNA and treated 24 h later with 35 nmol of ethanol showed an apparent reduction in their motor activities relative to those measured in the other three experimental groups. The two-way ANOVA confirmed the existence of significant differences for both treatment (F(1,25)=4.594; p=0.042) and the pre-treatment × treatment interaction (F(1,25)=5.103; p=0.033). Post-hoc comparisons revealed that the mean distance travelled by animals receiving β-FNA + ethanol was significantly lower than those obtained in the other three experimental groups. These results indicate that MOR blockade with the selective antagonist β-FNA is able to efficiently and selectively suppress the activating effect of ethanol. In Figure 1(c), the distance travelled by animals in each of the 10 2-min intervals of the experimental session is shown. As can be appreciated, the activity of the animals of the β-FNA + ethanol group was lower than that observed in the animals belonging to the other three experimental groups. This reduction extended from the 6–8-min period to the end of the experiment. Statistical comparison did not detect any significant difference between groups at any specific time interval. However, the cumulative effect of these small and non-significant motor reductions could explain the significant decrease in the motor activity detected for this experimental group along the 20-min experimental period.
Discussion
In our previous work (Marti-Prats et al., 2013) we hypothesized that ethanol directly administered into the posterior VTA gives rise to two simultaneous and opposing effects on motor behaviour in rats, reflecting the two ethanol-derived opposing effects on VTA DA neuron activity (Theile et al., 2008, 2009, 2011; Xiao and Ye, 2008). When ethanol is locally microinjected at low doses (35 nmol in our experiments), both effects, the activating and the depressing one, tend to offset each other, thus preventing the observation of any behavioural change. This lack of effect is usually considered a logical result, compatible with the U inverted-shape of the dose-response curve for ethanol, and attributable to the existence of very low tissue levels of ethanol in the brain region studied. However, our data clearly show that this approach is flawed.
To unravel the “non-observable” effects of ethanol, it is necessary to suppress selectively either the depression effects derived from the non-metabolized fraction of ethanol or, alternatively, the activating effects derived from the products of the local ethanol metabolism. Concretely, the strategy followed in this latter case has consisted of applying two different pharmacological pre-treatments to animals aimed at preventing salsolinol production: (i) selective blockers of acetaldehyde production or (ii) acetaldehyde sequestering agents to inactivate this metabolite chemically (Marti-Prats et al., 2013). The rationale for the use of these pre-treatments comes from electrophysiological, neurochemical and behavioural studies. Previous findings showed that the activating effects of ethanol could be selectively suppressed with the use of ethanol metabolism inhibitors, such as sodium azide (Arizzi-LaFrance et al., 2006; Melis et al., 2007), a potent inhibitor of catalase (DeMaster et al., 1985; Gill et al., 1992; Zimatkin and Lindros, 1996; Zimatkin et al., 2006), or using D-penicillamine, a β-mercapto-α-amino acid that efficiently reacts with acetaldehyde forming inactive adducts (Cohen et al., 2000; Nagasawa et al., 1980). This compound has an unusual ability to block the activating effects of ethanol (Enrico et al., 2009; Font et al., 2005; Marti-Prats et al., 2010). Using either of these two classes of pre-treatments, it was possible to transform the 35 nmol dose (initially ineffective) into a motor-depressing dose, demonstrating the ability of ethanol to exert opposing effects on behaviour simultaneously. At this point, an important question emerges: could the depressing effect observed after blocking the metabolism of ethanol be a non-desired effect derived from the alteration of the normal balance between ethanol metabolites and the non-biotransformed fraction of ethanol? This is an intriguing issue since the blockade of the acetaldehyde production may have affected the normal balance between unaltered and biotransformed fractions of ethanol, displacing it towards the unaltered fraction. In other words, could the inhibitory effects be falsely enhanced by pre-treatments?
In the present experiments, we have re-evaluated our previous results by employing a more direct strategy to suppress selectively the salsolinol-derived activation of animals without affecting the normal balance between ethanol metabolites and the non-metabolized fraction of ethanol. Since the activating effects of ethanol are dependent, in accordance to our working hypothesis, on the direct interaction of the metabolite salsolinol with MORs located in the soma of VTA GABA neurons (Xie et al., 2012), it seems logical to assume that the local and specific blockade of these receptors should produce behavioural consequences similar to those observed after inhibition of catalase or sequestration of acetaldehyde with D-penicillamine. This is, in fact, what we have demonstrated in the present study. Our results show that the irreversible blockade of MORs by means of β-FNA intra-VTA injected is able to selectively suppress the activating component of the ethanol effects, unmasking the motor depression derived from the non-biotransformed fraction of ethanol. One important subject to be considered is related to the use of β-FNA as a MOR antagonist. It is plausible to assume that an irreversible blockade of MORs could change baseline activity levels within the VTA and, additionally, induce homeostatic changes to compensate for the persistent MOR blockade (for example, increasing membrane expression of receptors or increasing the local release of other neurotransmitters and modulators). Furthermore, β-FNA has acute agonist properties at the kappa opioid receptor (Ward et al., 1982) which also could induce long-lasting changes through kinase pathways in the VTA function, including the GABA function (Graziane et al., 2013; Polter and Kauer, 2014). Nonetheless, under our experimental conditions, intra-VTA administration of β-FNA did not modify motor activities in animals the β-FNA + aCSF group in relation to those observed in the aCSF + aCSF-treated animals. We reported similar results in previous papers (Hipolito et al., 2010; Sanchez-Catalan et al., 2009). All in all, these data allow us to rule out the influence of these non-desired effects of β-FNA on our results.
The present data confirm and extend our suggestion that ethanol exerts two opposing and simultaneous effects on VTA DA neuron activity but, at the same time, they pose new questions. For example, at present it is not clear if this inhibiting effect of motor activity described for ethanol after intra-VTA administration to animals subjected to MOR blockade can be reproduced with drug doses lower than 35 nmol. In other words, it might be interesting to delineate the dose-effect curve for these depressing effects of ethanol in animals pre-treated with β-FNA. Another intriguing question is related to the impact that the MOR blockade in specific brain regions could have on the motor activity of rats receiving ethanol by the systemic route. Future experiments should explore both issues.
On the other hand, six animals in our experiment showed injection sites in the zone in which tVTA and posterior VTA overlapp (Figure 1(a)). Therefore, we cannot rule out that the observed effects in these animals are partly due to changes in the activity of tVTA neurons. Anatomically, the tVTA is positioned caudally beyond the VTA, although its most rostral portion overlaps with the posterior VTA (Bourdy and Barrot, 2012; Kaufling et al., 2009; Sanchez-Catalan et al., 2014). The tVTA is mostly composed of GABA neurons and exerts a major inhibitory control on the activity of dopamine cells (Jalabert et al., 2011). Neurons in tVTA show strong MOR immunostaining (Jalabert et al., 2011), and agonists for these receptors inhibit tVTA cells (Lecca et al., 2011; Matsui and Williams, 2011). Behavioural studies have also shown that MOR agonists are self-administered in the tVTA and induce a conditioned place preference when injected into this region (Jhou et al., 2012). Hence, it is plausible to assume that our treatments and pre-treatments may have affected the tVTA GABA neuron function. Unfortunately, due to the small number of animals showing injection points in this area we cannot draw any definitive conclusion.
In our opinion, the present behavioural findings are relevant to disentangle the mechanism of action of ethanol, but we are aware that more direct and detailed experiments are necessary to fully understand these phenomena. VTA cells seem to exhibit regional differences in response to different drugs of abuse including opioids (Ford et al., 2006; Margolis et al., 2008) and ethanol (Mrejeru et al., 2015). Consequently, the use of analysis techniques more precise than the behavioural ones (such as in vitro electrophysiological recordings) will be necessary to explore the specific role of salsolinol and the non-biotransformed fraction of ethanol on the activity of the different subpopulations of midbrain DA neurons in detail.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by grants from Ajudes per a Accions Especials d’Investigació, Universitat de València (UV-INV-AE13-136776) (Spain). LM-P is recipient of a predoctoral fellowship from the “Atracció de Talent” program, Universitat de València.