
Abstract
Childhood/adolescent antipsychotic drug (APD) use is exponentially increasing worldwide, despite limited knowledge of the long-term effects of early APD treatment. Whilst investigations have found that early treatment has resulted in some alterations to dopamine and serotonin neurotransmission systems (essential to APD efficacy), there have only been limited studies into potential long-term behavioural changes. This study, using an animal model for childhood/adolescent APD treatment, investigated the long-term effects of aripiprazole, olanzapine and risperidone on adult behaviours of male and female rats. Open-field/holeboard, elevated plus maze (EPM), social interaction and forced swim (FS) tests were then conducted in adult rats. Our results indicated that in the male cohort, early risperidone and olanzapine treatment elicited long-term hyper-locomotor effects (open-field/holeboard and FS tests), whilst a decrease in depressive-like behaviour (in FS test) was observed in response to olanzapine treatment. Furthermore, anxiolytic-like behaviours were found following testing in the open-field/holeboard and EPM in response to all three drug treatments. Effects in the female cohort, however, were to a far lesser extent, with behavioural attributes indicative of an increased depressive-like behaviour and hypo-locomotor activity exhibited in the FS test following early risperidone and olanzapine treatment. These results suggest that various APDs have different long-term effects on the behaviours of adult rats.
Introduction
The use of antipsychotic drugs (APDs) in children and adolescents is rapidly increasing worldwide, despite serious limitation in the safety and efficacy of APD use in these population groups (Alexander et al., 2011; Hoekstra, 2014; Olfson et al., 2010, 2012; Rani et al., 2008; Seida et al., 2012; Varley and McClellan, 2009). Whilst first-generation APDs (e.g. haloperidol and chlorpromazine) are used less frequently in children and adolescence due to their serious extrapyramidal side-effects, second-generation APDs, including aripiprazole, olanzapine and risperidone, are commonly prescribed (mostly off-label) for treating various childhood disorders; from mental illnesses including anxiety, depression and child-onset schizophrenia (Memarzia et al., 2014; Olfson et al., 2010; Schneider et al., 2014), to various behavioural disorders (Haw and Stubbs, 2007; Sharma and Shaw, 2012; Vitiello et al., 2009). Whilst the therapeutic effects of APDs are predominantly based on their antagonist or partial agonist mechanisms of action on the dopamine (DA) D2 and serotonin (5-HT) 5-HT1A and 5-HT2A/2C receptors (Grace et al., 2007; Kegeles et al., 2010; Meltzer, 2002; Meltzer et al., 2003; Milstein et al., 2013; Purves-Tyson et al., 2012), the dopaminergic and serotonergic neurotransmitter (NT) systems are also heavily involved in multiple neurodevelopmental processes during the childhood/adolescent period (Andersen, 2003; Frost and Cadet, 2000; Klomp et al., 2012; Levitt et al., 1997; Milstein et al., 2013). There is, therefore, the potential for APD use in such a critical neurodevelopmental time period to carry a substantial risk of long-term behavioural pathologies, due to their potent actions on the still developing DA and 5-HT NT systems (Andersen, 2003, 2005; Andersen and Navalta, 2004; Memarzia et al., 2014; Vinish et al., 2012).
Although current clinical research into the use of APDs in the adolescent population over both short term periods of between one and two months, and longer-term periods of up to six months has found some benefits in the treatment of some mental illness symptomology (Kumra et al., 2008; Stigler et al., 2004; Zuddas et al., 2011), there is limited data on the potential for long-term alterations to adult behaviours (Milstein et al., 2013; Shu et al., 2014; Varela et al., 2014). Although the effects of childhood/adolescent APD use on adult behaviours is unknown in a clinical setting, current animal studies have shown that early drug treatment for a period of up to four weeks can result in various significant alterations in the dopaminergic and serotonergic pathways in the brain long-term (Maciag et al., 2006; Memarzia et al., 2014; Milstein et al., 2013; Moran-Gates et al., 2006; Varela et al., 2014; Vinish et al. 2012), including permanent changes in the distribution of NT receptors and dendritic architecture (Maciag et al., 2006; Milstein et al., 2013; Moran-Gates et al., 2006), with minimal evidence of the direct effects on behaviour. Alterations to the production, transport and binding of DA and 5-HT in the cortical and striatal brain regions have already been directly linked to changes in behavioural attributes, including locomotor activity levels, along with depressive-like and anxiety-like behaviours. The present study, therefore, investigated the long-term effects of APD treatment during the critical neurodevelopmental window of childhood/adolescence, on behavioural attributes in adult rats, including locomotor activity, depressive-like, anxiety-like, exploratory and social behaviours.
Materials and methods
Animals and housing
Timed pregnant Sprague–Dawley rats were obtained at gestation day 16 from the Animal Resource Centre (Perth, Australia). They were housed in individual cages under environmentally controlled conditions (22°C, light cycle from 07:00h to 19:00h and dark cycle from 19:00h to 07:00h), and allowed ad libitum access to water and standard laboratory chow diet (3.9 kcal/g: 10% fat, 74% carbohydrate, 16% protein). Day of birth was considered postnatal day (PD) 0. Pups were sexed on PD14, and then 96 Sprague–Dawley rats (48 male and 48 female) were weaned on PD20 and housed in individual cages. All experimental procedures were approved by the Animal Ethics Committee, University of Wollongong, NSW, Australia (AE 12/20), and complied with Australian Code of Practice for the Care and Use of Animals for Scientific Purpose (2004).
Drug treatment groups
Before the drug treatment commenced, the rats were trained for self-administration by feeding them cookie dough (0.3 g) without drugs two times per day for PD18–PD21. Animals were then assigned randomly to one of four experimental groups per gender on PD21 (n=12/group): (1) aripiprazole (Otsuka, Japan), (2) olanzapine (Eli Lilly, USA), (3) risperidone (Apotex, Canada) or (4) control (vehicle). The drug treatment period from PD22–PD50 in the rats was carried out at the equivalent time of the childhood/periadolescent phase in humans (Andersen, 2003). A staggered drug treatment pattern, where lower APD dosages are slowly increased to a final dosage amount, was used to mimic a clinical setting (Taylor et al., 2009). The APD doses were initiated on PD22 at 0.2 mg/kg three times per day for aripiprazole, 0.25 mg/kg three times per day for olanzapine and 0.05 mg/kg three times per day for risperidone, and then increased in three steps over the first seven days of the four week treatment period, to achieve a final dose on PD28 of 1 mg/kg three times per day for aripiprazole, 1 mg/kg three times per day for olanzapine and 0.3 mg/kg, three times per day for risperidone. The proposed dosages are within the recommended dosage ranges for the psychiatric treatment of paediatric patients, based on the body surface area formula for dosage translation between humans and rats in the FDA guideline for clinical trials (Food and Drug Administration, 2005; Reagan-Shaw et al., 2008; Taylor et al., 2009; Zuddas et al., 2011). It has been previously reported that, at these used dosages, aripiprazole drug treatment reaches above 90% DA D2 receptor occupancy rates in the rat brain (Wadenberg, 2007), while olanzapine and risperidone reach 65%–80% DA D2 receptor occupancy (Kapur et al., 2003; Natesan et al., 2006). These dosages have also been shown to be physiologically and behaviourally effective in our laboratory, with similar dosages seen to induce weight gain and changes in hypothalamic neuropeptide Y expression in adolescent rats (Lian et al., 2015), whilst alterations to both DA and 5-HT receptor binding has been reported in adult rats (Han et al., 2009; Weston-Green et al., 2011). The 0.3 g dry cookie dough pellets were fed to the rats three times per day (07:00h, 14:00h and 22:00h; with 8±1 hour intervals) over the four week treatment period. Rats were observed throughout the treatment to ensure that they completely consumed the cookie dough pellet. The rats in the control group received an equivalent pellet without the drug.
Behavioural tests
All behavioural tests were carried out between 09:00h and 17:00h. All protocols for the behavioural tests used in the present study were adapted from previous studies (Burne et al., 2004; du Bois et al., 2008). Behavioural tests were carried out over a four week period from PD72 to PD94 (the equivalent of adulthood) in the following sequence; open-field/holeboard test, elevated plus maze (EPM) test, social interaction test, and forced swim (FS) test, as described below.
Open-field/holeboard test
The open-field/holeboard test was modified from previous designs and carried out on PD72 to examine exploratory behaviour and locomotor activity (Burne et al., 2004; du Bois et al., 2008). A single rat was placed in a black square arena (60 cm × 60 cm × 40 cm), with a floorboard insert placed 5 cm above the base, with four holes, 5 cm in diameter, each situated 10cm in from each of the corners. The light intensity was 30 lx across the entire arena. Rat behaviour was recorded with a centrally located video camera for 30 minutes. Locomotor activity was later analysed using video-tracking software (Noldus Information Technology, Wageningen, The Netherlands). The frequency of head dips were scored manually.
Elevated plus maze
The EPM test was modified from previous designs and carried out on PD79, to examine anxiety-like behaviours and locomotor activity (Burne et al., 2004; du Bois et al., 2008; Mällo et al., 2007; Wigger and Neumann, 1999). It consisted of two open arms (50 cm × 7 cm) and two closed arms (50 cm × 7 cm × 30 cm) with an open roof, arranged around a central platform (7 cm × 7 cm). Like-arms opposed each other across the central platform. The maze was a wooden structure, painted with a black acrylic, and elevated 60 cm above the floor. The light intensity was set at 100 lx at the open arms and central platform. A single rat was placed on the central platform facing an open arm and rat behaviour was recorded for seven minutes with a centrally located video camera. We measured the number of open and closed arm entries, and the duration of time spent in the open and closed arms, central platform and ‘open part’ (open arms plus central platform). We also measured the number of arm changes as a measure of general locomotor activity on the maze. Data was scored manually, the criterion for entry into the open/closed arms was whole body and four paws entry; a rat was considered to be in the central platform zone if its head and front paws were in the central platform, with its body positioned in the closed arm.
Social interaction
The social interaction test was modified from previous designs and was carried out on PD86 to examine social and anxiolytic behaviours (Burne et al., 2004; du Bois et al., 2008). It was performed in a black circular arena (90 cm diameter × 30 cm high walls) made of black acrylic, with light intensity at 30 lx across the entire arena. Two unfamiliar rats of a similar size, of the same sex and from the same treatment group, were placed on opposing sides of the arena and allowed to move around freely, and behaviour was recorded for seven minutes with a centrally located video camera. All rats were only tested once. Behaviour was analysed via the use of video-tracking software (Noldus Information Technology) looking at time spent within a given proximity (set at 20 cm – average body length) of one another and mean distance between each other, and data gathered from each Social Interaction test involving two rats was analysed as a single data point. No aggressive behaviours were observed using this procedure.
Forced swim test
The FS test was modified from previous designs and was carried out on PD93 and PD94 to examine depressive-like behaviours and locomotor activity (Burne et al., 2004; du Bois et al., 2008). A blue opaque cylinder (30 cm × 50 cm) was filled to a predetermined water level (30°C), dependent upon the body length of each individual rat. Rats were measured from the tip of the snout to the base of the tail, and water depth was then calculated by multiplying this value by 1.25. Rats were tested over two days, at a light intensity of 1000 lx (fluorescence strip lighting-stress conditions). On the first day, the rats were placed in the water for a 10-minute period, then removed and dried with towels and placed in a cage with paper towels for 20 minutes prior to being returned to their home cages. Water was changed between rats. On the second day, the rats were placed in the cylinder for a period of five minutes. Locomotor activity was recorded on the second day using a centrally located video camera. We manually evaluated the time spent climbing (defined as vertical movements of the forepaws, usually up against the cylinder), swimming (movement throughout the cylinder, usually in a horizontal direction), and floating (immobile – an absence of movement except for that required to keep its head above water).
Statistical analysis
All collected data were analysed using SPSS software (Windows version 19.0, SPSS Inc., Chicago, USA). The Kolmogorov–Smirnov test was used to examine the distribution of data from all experiments. All normally distributed data from male and female rats were also analysed by two-way analysis of variance (ANOVA) (gender×treatment). Then data from males or females were analysed separately by one-way ANOVA, followed by post-hoc Tukey tests for multiple comparisons between the treatment groups. Due to the interrelationship of activity levels, we also utilized an analysis of covariance (ANCOVA) test with total distance travelled (cm) obtained in the open field/hole board test as the covariate factor to further analyse data from the EPM and FS tests. Data that was not normally distributed was analysed via the non-parametric Mann–Whitney U-test. All data were expressed as mean ± standard error of the mean (±SEM), and statistical significance was accepted when p⩽0.05.
Results
Following a childhood/adolescent treatment of aripiprazole, olanzapine, risperidone or vehicle for four weeks (PD22–PD50), a series of four behavioural tests, including the open-field/holeboard, EPM, social interaction and FS tests, were conducted over a four week period (PD72–PD94).
Open field/holeboard
Two-way ANOVAs (gender×treatment) revealed significant effects of gender on average velocity (F1,90=49.780, p<0.001), total distance moved (F1,90=32.838, p<0.001) and rearing (F1,90=17.028, p<0.001), along with a significant interaction between the two factors on total distance moved (F3,90=2.772, p<0.05). Analyses revealed that female rats had a higher locomotor activity level compared to male rats (p<0.05) and, furthermore, male and female rats were also found to respond differently to early APD treatment (Table 1). In male rats, a significant effect of treatment on average velocity (F3,46=4.473, p<0.01), total distance moved (F3,46=5.115, p<0.01) and a trend to significance with rearing (F3,45=2.685, p=0.059) were found. Early risperidone treatment increased total distance moved (+32%, p<0.01), average velocity (+33%, p<0.01) and rearing (+49%, p<0.05) (Table 1). These increases in average velocity, total distance moved and rearing indicated that male rats treated with risperidone in the childhood/adolescent period (PD22–PD50) exhibit hyper-locomotor activity levels following open-field/holeboard behavioural testing in adulthood (PD72) (Figure 1). However, no alterations in locomotor activity were observed in the female treatment groups, indicating that early APD treatment has a stronger long-term effect on the male cohort.
Behaviour of male and female Sprague–Dawley rats with chronic treatment of aripiprazole (1.0 mg/kg, t.i.d.), olanzapine (1.0 mg/kg, t.i.d.), risperidone (0.3 mg/kg, t.i.d.) in the open-field/holeboard tests.
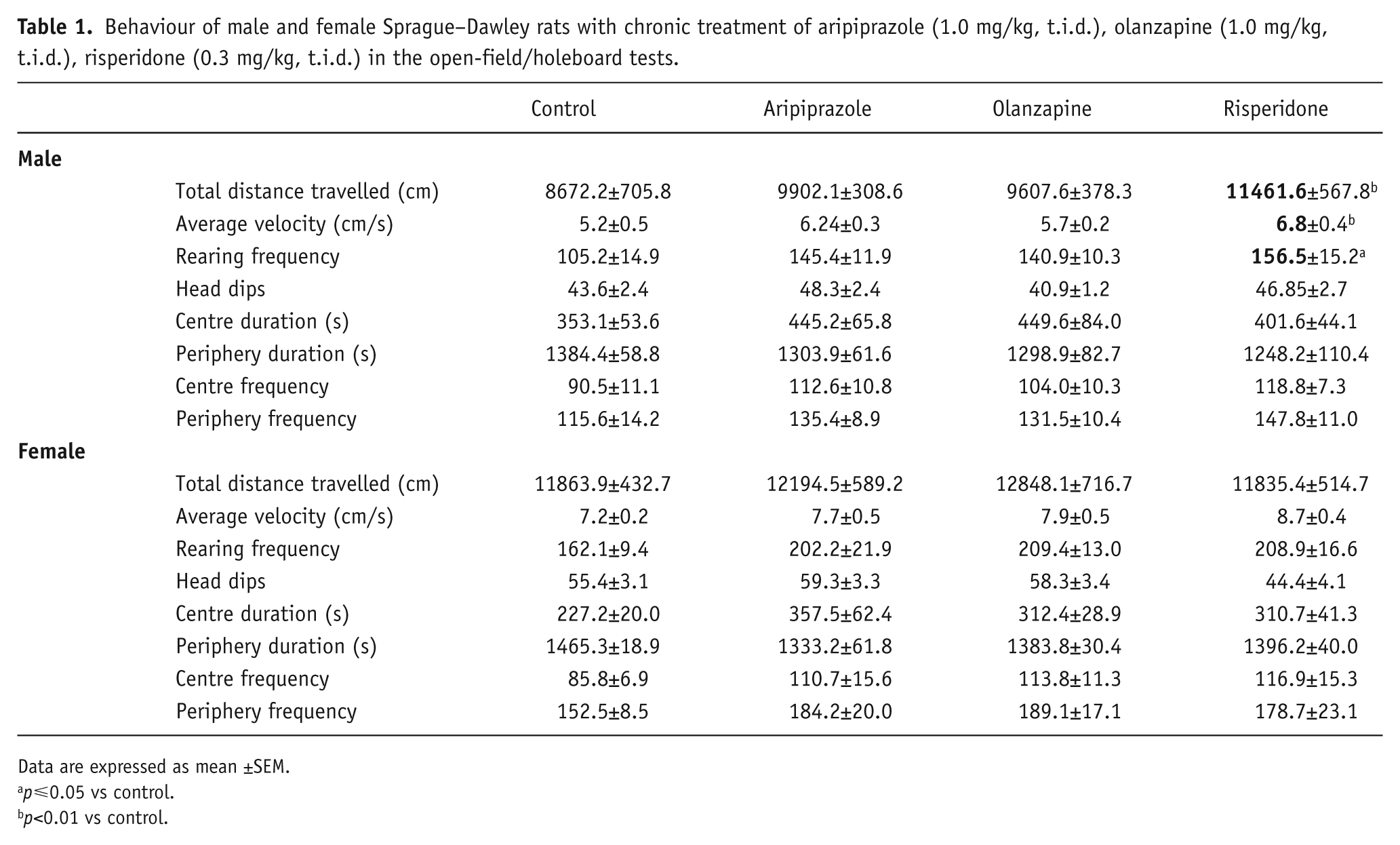
Data are expressed as mean ±SEM.
p⩽0.05 vs control.
p<0.01 vs control.
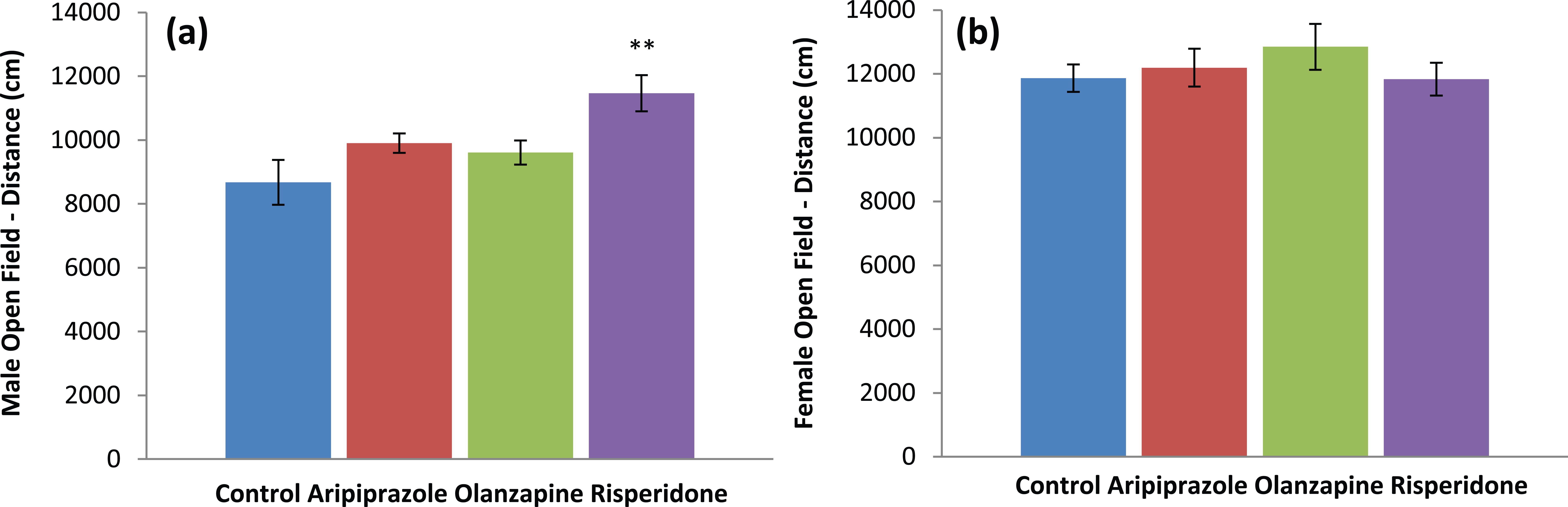
Behavioural effects – activity levels: (a) male open field/holeboard, distance travelled (cm); (b) female open field/holeboard, distance travelled (cm) in Sprague–Dawley rats treated chronically with aripiprazole (1.0 mg/kg, t.i.d), olanzapine (1.0 mg/kg, t.i.d), risperidone (0.3 mg/kg, t.i.d) or control (vehicle). Data expressed as mean ±SEM. **p<0.01 vs control. t.i.d.: three times daily.
Elevated plus maze
Analyses of EPM behavioural data once again revealed that male and female rats were found to respond differently to early APD treatment (Table 2). Two-way ANOVA (gender×treatment) revealed significant interaction between treatment and gender on centre duration (F3,85=4.023, p<0.02) and closed arm duration (F3,85=5.149, p<0.01). In male rats, analysis via ANCOVA of closed arm changes data found a significant effect of total distance travelled (F1,46=8.403, p<0.01), whilst one-way ANOVA found that early APD treatment significantly affected centre duration (F3,45=8.924, p<0.001), closed arm duration (F3,45=7.303, p<0.001) and open part duration (F3,45=7.303, p<0.001) (Table 2). Early aripiprazole (+26%, p<0.001), olanzapine (+27%, p<0.05) and risperidone (+14%, p<0.01) treatment also increased centre duration compared to the control group. Furthermore, early aripiprazole (+53%, p<0.001) and risperidone (+30%, p=0.05) treatment was found to significantly increase time spent in the open part (open arms + centre platform), with a similar trend also found in the early olanzapine (+29%, p=0.082) treatment group compared to the control. There were reduced closed arm durations following the early aripiprazole (-26%, p<0.001) and risperidone (-14%, p=0.05) treatments, whilst the early olanzapine (-14%, p=0.082) treatment resulted in a similar trend (Table 2) (Figure 2). However, there were no significant differences in open arm entries or closed arm entries. In female rats, the early aripiprazole treatment resulted in a trend to a significant increase in closed arm duration (+57%, p=0.056). These results indicated that early APD treatment with aripiprazole, olanzapine and risperidone in the male rodent model all subsequently led to the expression of anxiolytic-like behaviours in adulthood. In the female model, although no significant effects of early APD treatment on anxiety-like behaviours were found, analysis via ANCOVA found a trend to significant effects of total distance travelled on closed arm changes (F1,42=3.077, p=.088), indicating again that early APD treatment has stronger long-term effects on the male cohort.
Behaviour of male and female Sprague–Dawley rats with chronic treatment of aripiprazole (1.0 mg/kg, t.i.d.), olanzapine (1.0 mg/kg, t.i.d.), risperidone (0.3 mg/kg, t.i.d.) in the elevated plus maze (EPM) tests.
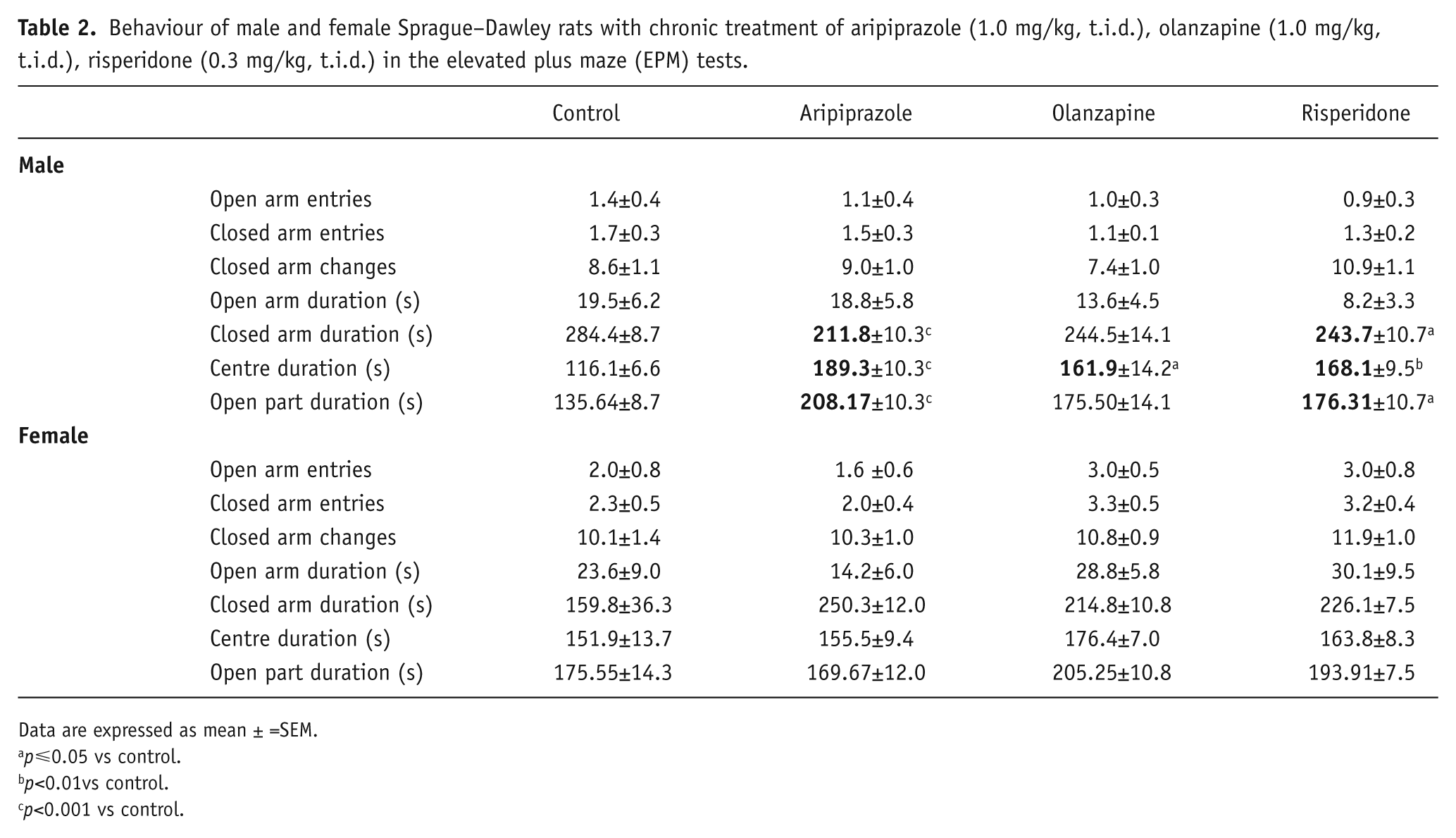
Data are expressed as mean ± =SEM.
p⩽0.05 vs control.
p<0.01vs control.
p<0.001 vs control.
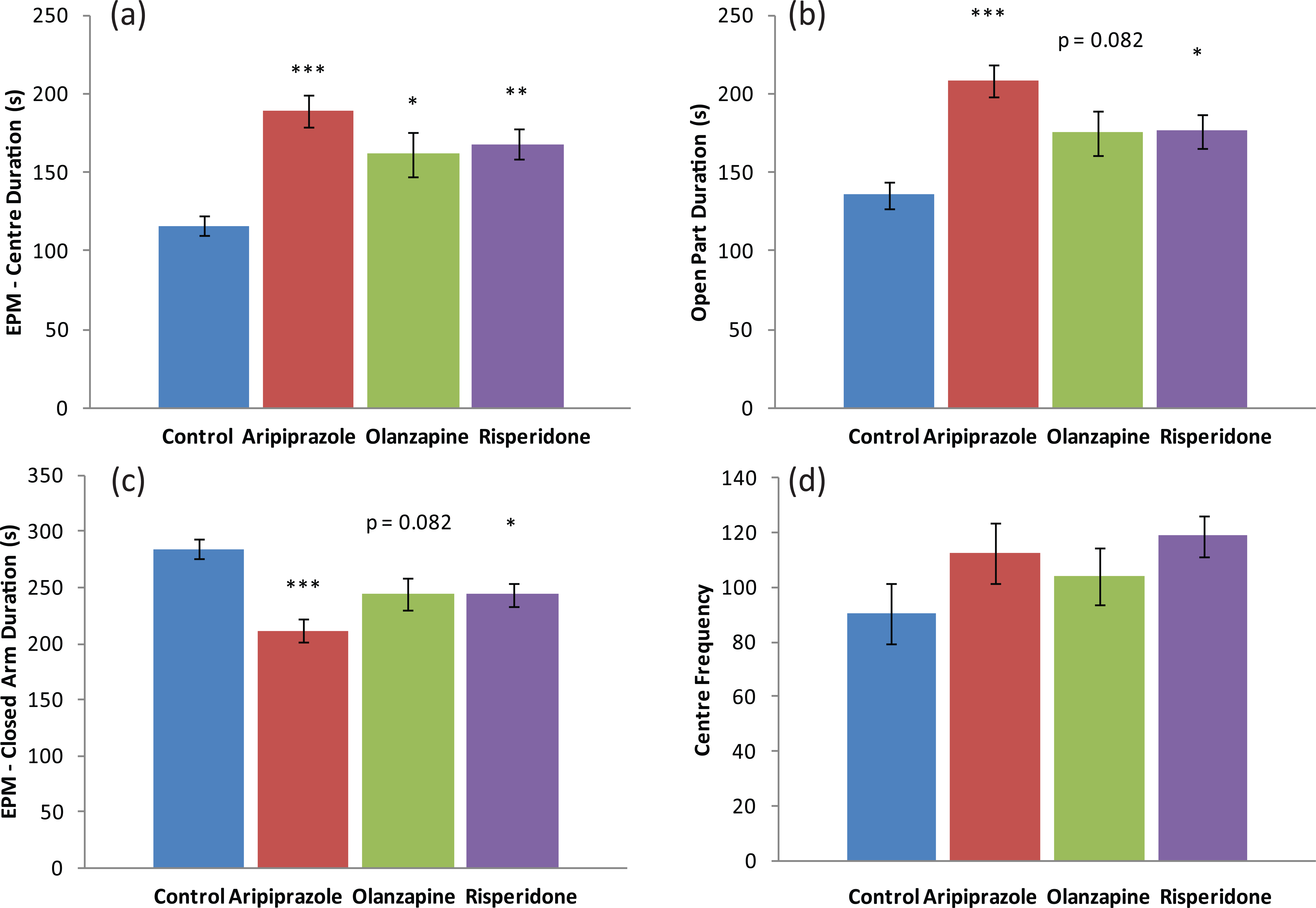
Behavioural effects – anxiety: (a) elevated plus maze (EPM) test centre duration (s); (b) EPM test open part duration (s); (c) EPM test closed arm duration (s); (d) open field/holeboard centre frequency, in male Sprague–Dawley rats treated chronically with aripiprazole (1.0 mg/kg, t.i.d), olanzapine (1.0 mg/kg, t.i.d), risperidone (0.3 mg/kg, t.i.d) or control (vehicle). Data expressed as mean ±SEM. *p<0.05, **p<0.01, ***p<0.001 vs control. t.i.d.: three times daily.
Social interaction
On PD86, all treatment groups and both genders underwent social interaction testing to examine behavioural parameters, including anxiety and social behaviours. Analysis found no significant differences between the control and drug treatment groups across all of the measured parameters (Table 3). The results indicate that childhood/adolescent APD treatment in both male and female rats had no significant effects on the adult social behaviours examined by the social interaction tests, when compared to the control group.
Behaviour of male and female Sprague–Dawley rats with chronic treatment of aripiprazole (1.0 mg/kg, t.i.d.), olanzapine (1.0 mg/kg, t.i.d.), risperidone (0.3 mg/kg, t.i.d.) in the social interaction tests.
Data are expressed as mean ±SEM.
Forced swim test
Analyses again demonstrated that male and female rats responded differently to early APD treatment (Table 4). Two-way ANOVA (gender× treatment) revealed significant effects of gender on climbing duration (F1,86=60.582, p<0.001), swimming duration (F1,86=67.758, p<0.001) and floating duration (F1,86=19.833, p<0.001), along with a significant interaction between the two factors on climbing duration (F3,86=6.929, p<0.001), swimming duration (F3,86=5.880, p<0.01) and floating duration (F3,86=7.989, p<0.001). In male rats, analysis via ANCOVA found a trend to significant effect of total distance travelled on swimming duration (F1,46=3.165, p=0.083) and a trend to significant effect of treatment on climbing duration (F3,46=3.012, p<0.05), whilst analysis via one-way ANOVA found that early APD treatment caused a trend towards significance on floating duration (F3,45=2.720, p=0.056). Early olanzapine drug treatment increased climbing duration (+87%, p<0.05) and decreased floating duration (-21%, p<0.05) compared with the control (Table 4). However, similar effects were not observed in any of the other male drug treatment groups (both p>0.05). In female rats, analysis of data via ANCOVA found a significant effect of both total distance travelled (F1,43=4.839, p<0.05) and treatment (F3,43=3.477, p<0.05) on swimming duration, whilst analysis of climbing duration also found a significant effect of treatment (F3,43=5.019, p<0.01). Furthermore, one-way ANOVA of floating duration also found a significant effect of early APD treatment (F3,46=4.428, p<0.01) (Table 4). Early olanzapine drug treatment decreased climbing duration (-55%, p<0.05), and increased floating duration (+17%, p<0.05) when compared to the control group. Furthermore, early risperidone drug treatment was found to significantly decrease swimming duration (-50%, p<0.05) and climbing duration (-47%, p=0.05) and increase floating duration (+20%, p<0.05) when compared to the control (Table 4). These results indicate that early APD treatment has opposing effects on locomotor activity and depressive-like behaviour in adult male and female rats, indicating again that early APD treatment causes a gender-specific response in behavioural activity (Figure 3). Furthermore, the early aripiprazole treatment was not found to change locomotor activity, nor depressive-like behaviour in either the male or female cohorts.
Behaviour of male and female Sprague–Dawley rats treated with chronic treatment of aripiprazole (1.0 mg/kg, t.i.d.), olanzapine (1.0 mg/kg, t.i.d.), risperidone (0.3 mg/kg, t.i.d.) in the forced swim (FS) tests.
Data are expressed as mean ± SEM.
p⩽0.05 vs control.
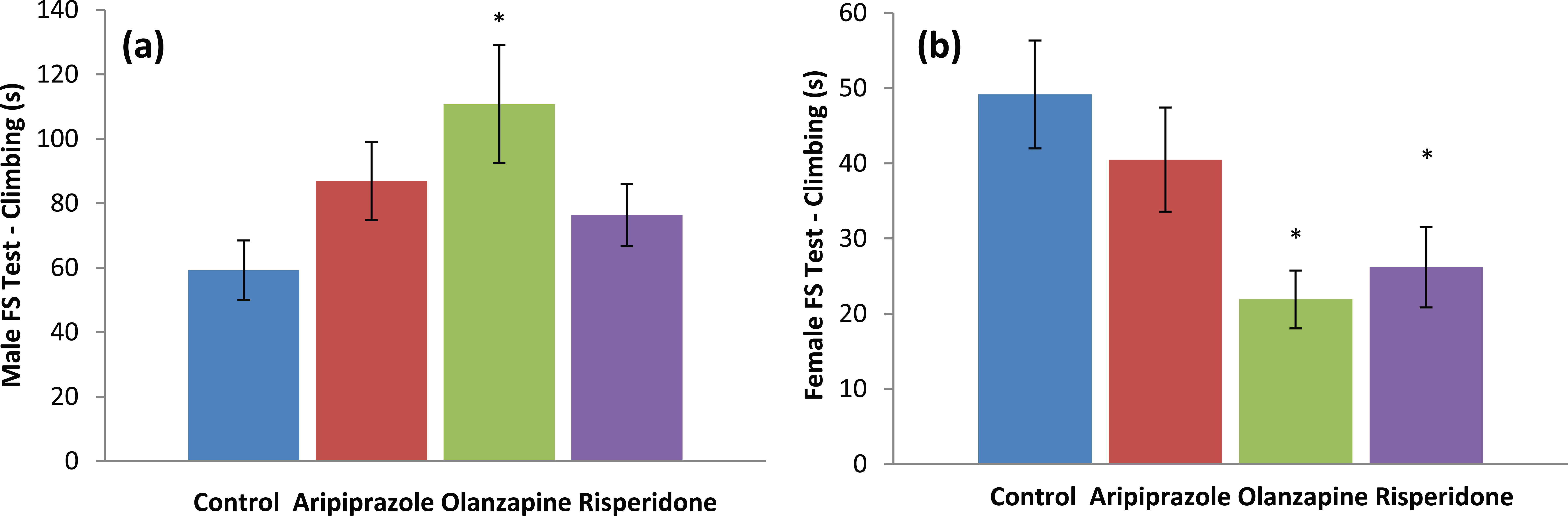
Behavioural effects – depressive: (a) male forced swim (FS) test climbing (s); (b) female FS test climbing (s) in Sprague–Dawley rats treated chronically with aripiprazole (1.0 mg/kg, t.i.d), olanzapine (1.0 mg/kg, t.i.d), risperidone (0.3 mg/kg, t.i.d) or control (vehicle). Data expressed as mean ±SEM. *p<0.05 vs control. Abbreviations: FS: forced swim; t.i.d.: three times daily.
Discussion
The present study investigated the effects of childhood/adolescent treatment with aripiprazole, olanzapine and risperidone on long-term (adult) behavioural attributes in both male and female rats. Our results provide evidence that early APD treatment causes long-term changes to locomotor, depressive-like and anxiety-like behaviours, and demonstrate that there are gender differences in both locomotor activity and depressive-like behaviours in response to early APD treatment.
Our study found significant increases in numerous parameters indicative of hyper-locomotor activity in the male rodent model between the drug treatment and control groups, across both the open-field/holeboard and FS behavioural tests (Figure 1). Rats that received early risperidone treatment showed increased velocity, distance and rearing in the open-field/holeboard test, whilst rats that received early olanzapine treatment showed increased time spent climbing and decreased time spent floating when compared to the control. In addition, locomotor activity of the male rat early aripiprazole drug treatment group was unchanged compared to the control. Although increased rearing behaviour has previously been correlated to vertical activity and exploratory/vigilant behavioural attributes (Brenes Sáenz et al., 2006; du Bois et al., 2008), no alterations in the number of head dips, also an indicator of exploratory behavior, were observed. This therefore indicates the differences in rearing activity observed may potentially be due to alterations in activity levels alone.
Thus far, animal studies investigating the effects of early, short-term treatment with APDs on long-term locomotor activity have found differing results (Milstein et al., 2013; Shu et al., 2014; Varela et al., 2014). Milstein and colleagues (Milstein et al., 2013) investigated the effects of early olanzapine treatment on locomotor activity in adulthood, with no changes to numerous parameters of the open-field test observed. Similarly, the present study found increases in the activity levels of olanzapine-treated animals in the FS test but not in the open-field/holeboard test, potentially due to the increased stress levels induced by the FS testing procedure. In addition, the investigation by Shu et al. (2014) found that repeated early treatment with olanzapine and clozapine induced a persistent inhibition of PCP-induced hyper-locomotion in adulthood, a test commonly used to investigate motor activity levels. However, they reported that on ‘rehabituation’ days in adulthood (when rats were totally drug free during the long-term drug treatment period), visible increases in activity levels were observed. It is possible that this is due to a drug withdrawal-induced behavioural hypersensitivity, and that the long-term antagonistic actions of drugs, such as the administration of olanzapine and risperidone in adolescence, potentially cause the changes in activity levels observed in the present study. Furthermore, concurrent with the findings of our study, numerous studies have also found that aripiprazole treatment does not cause any impairment to motor activity levels (Biojone et al., 2011; Leite et al., 2008).
Along with increased locomotor activity, the aforementioned changes to behavioural parameters in the FS test was also indicative of changes to depressive-like behaviour in both the male and female rodent models of the present study. In the male cohort, rats that received early olanzapine treatment showed increased time spent climbing and decreased time spent floating, indicative of a decrease in depressive-like behaviour (Costa et al., 2013). Conversely, in the female cohort, significant decreases in time spent climbing and significant increases in time spent floating were observed in both early olanzapine and risperidone drug treatments, indicative of an increase in depressive-like behaviour (Costa et al., 2013).
Previous animal studies looking at the effects of APD treatment on depressive-like behaviour in the FS test has found similar gender-dependent results to those in the present study (Sharma et al., 2015; Weiner et al., 2003). Weiner and colleagues (Weiner et al., 2003) found that acute treatment of adult male rodents with olanzapine decreased immobility time (time spent floating) in response to the APD treatment, similar to the results found in the present study. Additionally, increases in immobility time were found by Sharma and colleagues (Sharma et al., 2015) following long-term olanzapine treatment in adult female rodents, observations that were also found in the present study.
The differences in the olanzapine, risperidone and aripiprazole receptor binding profiles, receptor occupancy levels and mechanisms of action, in particular on the DA and 5-HT receptors, may result in the locomotor activity and depressive-like behavioural differences across these different drug treatment groups. With high-affinity, high receptor occupancy, partial agonist mechanism of action known to have the beneficial effects of normalizing NT activity (Natesan et al., 2006), aripiprazole (a partial D2 agonist) may not cause the long-term alterations to the DA NT system seen with adolescent exposure to the potent antagonistic effects of APDs, including olanzapine and risperidone, and subsequently no changes to locomotor activity. Although changes to locomotor activity-related behavioural parameters were exhibited in the holeboard and FS tests, similar changes were not found in the EPM. One possible explanation for this is the added parameter of height present in the EPM that was not present in the other behavioural tests. The effects of this increased height also potentially affect the number of open-arm entries/duration, and thus the anxiety-like behavior, as examined below. This leads to the possibility that previously documented differences in height and bright-space anxiety (Komada et al., 2008) effect the level of locomotor activity in the EPM test.
Analysis of behavioural data from the male cohort of the present study also found changes to behavioural parameters indicative of anxiolytic behaviour. Compared to the control group, long-term increases in time spent in the open-part (open-arm plus centre duration), and corresponding decreases in time spent in the closed-arms of the EPM, were observed in all three APD treatment groups, parameters known to be indicative of anxiolytic behaviour (Komada et al., 2008; Mällo et al., 2007; Tsujimura et al., 2008; Walf and Frye, 2007). However, segmenting the open-part data into time spent on the open-arm and centre platform individually found a significant increase in time spent on the centre platform only, with no differences between drug treatment groups in time spent on the open arms. In addition, a trend to significant increase in centre frequency in the open field/holeboard test following early risperidone treatment was also found. Thus, in the present study, adolescent APD treatment may potentially cause anxiolytic effects specifically towards bright spaces (due to the differences in light intensity of the open part (100 lx) and closed-arms (darkness)) when compared to the control (Figure 2), with no differences in height-anxiety between the treatment groups and the control being found at the 60 cm elevation. Further behavioural testing, such as the light/dark transition test, is required, however, to clarify any different aspects in anxiety-like behaviour. However, previous animal studies examining the effects of APD treatment on anxiety-like behaviour have found differing results across treatment groups. Whilst studies in adult rats over a short-term/acute period have found that the partial agonist actions of aripiprazole induced a similar increase in time spent in the open-arms and a decrease in time spent in the closed-arms of the elevated maze (indicative of anxiolytic effects) (Biojone et al., 2011), studies investigating the effects of treatment with risperidone and haloperidol demonstrated anxiogenic effects, opposite to the effects seen in the present study (Karl et al., 2006).
The discrepancy between the results of our study and previous ones may be due to both the differences in time between treatment and behavioural testing and the age of the animals undergoing treatment. Firstly, it is important to consider that, whilst previous studies have investigated the immediate effects of short-term/acute or chronic treatment in adult rats on anxiety-related behaviours in adulthood, our study investigated the long-term effects of early (childhood/adolescent) APD treatment on adult behaviour. Hence, changes to NT systems, including the DA and 5-HT systems targeted by APDs, may have occurred following APD withdrawal in the time period between drug treatment and behavioural testing in the present study, resulting in the differences in both depressive-like and anxiety-like behaviours. Furthermore, the current study used a young animal treatment model compared to the adult model of previous studies. This difference in treatment age may be critical to the different results observed, with the antagonistic actions of olanzapine treatment during the critical neurodevelopmental period previously found resulting in long-term alterations to NT functioning, including the DA and 5-HT NT systems (Milstein et al., 2013; Vinish et al., 2012).
Surprisingly, our study also found gender-specific behavioural effects in response to early APD treatment. Firstly, no anxiety-related changes were observed in adult female rats, whilst the alterations to locomotor activity were found to be both dissimilar and to a far lesser extent to those of the male cohort. Further comparison of activity related changes also identified that the significant gender-based difference in total distance moved (~33% greater for females) was almost as strong as the effect of risperidone drug treatment in the male cohort (~26% greater for risperidone treatment), reported previously. Also, whilst behavioural observations indicative of changes in depressive-like behaviour were also seen in the adult female rats FS test, early treatment with olanzapine and risperidone were found to induce an increase in depressive-like behaviour in adulthood, an opposite effect to that seen in the male cohort. Analysis of the FS test data revealed decreases in time spent climbing (s; olanzapine treatment) and swimming (s; risperidone treatment), and subsequent increases in time spent floating (s; olanzapine and risperidone treatment). There could be numerous reasons for the differential effects in locomotor, anxiety and depressive-related behaviour observed between the genders. Firstly, gender-based differences in the neurodevelopment of the DA NT system are well-postulated, and may affect the behavioural differences observed in the present study. During the developmental phase, studies have found both regional and gender-based differences in the development of the DA NT system, including differences in striatal DA D1 and D2 receptor overproduction and elimination, DA D1 receptor densities in the nucleus accumbens, grey and white matter overproduction and elimination, and rates of myelination (Andersen, 2003; Sinclair et al., 2014). These gender-based differences in the development of the DA NT system, combined with early APD treatment during this neurodevelopmental window, may explain the observed gender differences in the predominately DA-related locomotor activity changes of the present study.
Furthermore, the sex hormones testosterone and estrogen may also contribute to the gender differences in the behavioural results observed in the present study. During adolescence, testosterone and estrogen influence brain maturation/development in dopaminergic regions including the striatum and prefrontal cortex, and also have a significant role in shaping the dopaminergic signal (Andersen, 2003; Purves-Tyson et al., 2012; Sinclair et al., 2014). Previous animal studies have found that augmenting both testosterone and estrogen levels has resulted in altered DA neurotransmission, including DA synthesis, DA receptor mRNA expression and DA transporter levels, as well as changes in DA-related behaviours such as locomotor activity (Purves-Tyson et al., 2012; Sinclair et al., 2014). Additionally, some ‘neuro-protective’ effects of estrogen on DA and 5-HT NT system-related behaviours have been found, with estrogen exhibiting an ability to inhibit the DA D2 and 5-HT1A receptor-induced mediated behavioural changes in sensorimotor gating/information processing, which are deficient in people with mental illnesses (Dunlop and Nemeroff, 2007; Gogos et al., 2012). This ability of estrogen to inhibit both DA and 5-HT related changes may account for the differences in the predominately 5-HT-related anxiety, along with the DA and 5-HT related depressive-like behaviours observed between genders; however, further research in this area is needed.
It has been postulated that the physiology behind early insult drug induced hyper-locomotor effects may be due to a drug-treatment-induced long-term receptor hypersensitivity (Samaha et al., 2007; Shu et al., 2014). This phenomenon reports that continuous APD treatment (with high affinities for DA receptors) may cause an up-regulation of DA receptors, and thus, upon APD withdrawal, enhance dopaminergic signalling in brain regions including the nucleus accumbens, hippocampus and striatum, previously observed in animals exhibiting hyper-locomotor activity (Beaulieu and Gainetdinov, 2011; Kelly et al., 1975; Kusljic et al., 2003; Miyakawa et al., 2003; Seo et al., 2008). The potent antagonist actions of chronic risperidone and olanzapine APD treatment on the DA D2 receptors during the critical childhood/adolescent neurodevelopmental period in the present study may have resulted in a long-term DA receptor and consequent DA synthesis up-regulation, enhancing the dopaminergic signal along these brain regions previously heavily implicated in locomotor activity.
Furthermore, the antagonistic actions of olanzapine and risperidone on the 5-HT2A and 5-HT2C receptors resulting in decreases in 5-HT projections may also result in similar long-term alterations to dopaminergic signalling and locomotor activity. Previous studies have found strong links between the antagonism of 5-HT2 receptors and enhanced locomotor activity (Kusljic et al., 2003; Seo et al., 2008), potentially due to the negative correlation between 5-HT and DA functioning. In such cases, a deficiency in 5-HT projections leads to a disinhibition of DA signalling, and thus enhanced DA signal (Kusljic et al., 2003; Seo et al., 2008). Similarly, the antagonistic actions of olanzapine and risperidone on the 5-HT2A, 5-HT2C and DA D2 receptors, resulting in the aforementioned long-term alterations in signalling, have also been correlated to anxiolytic and decreased depressive-like behaviours (Andersen and Navalta, 2004; Biojone et al., 2011; De Oliveira et al., 2009; Dunlop and Nemeroff, 2007; Karl et al., 2006; Mora et al., 1997; Seo et al., 2008). Previous studies have demonstrated that repeated antagonism of the 5-HT2 receptors (both 5-HT2A and 5-HT2C) has been found to result in a down regulation of both the number and sensitivity of 5-HT2 receptors (Mora et al., 1997), resulting in a long-term deficiency in serotonergic signalling, correlated with anxiolytic- and decreased depressive-like behavioural effects. Long-term APD use, resulting in the up-regulation and hypersensitivity of DA receptors and thus enhanced dopaminergic signaling, may also play a role, with enhanced DA signalling in brain regions, including the ventral tegmental area, prefrontal cortex and nucleus accumbens, correlated with the anxiolytic- and decreased depressive-like behavioural effects exhibited in the present study (Biojone et al., 2011; De Oliveira et al., 2009; Karl et al., 2006).
Finally, despite a high affinity partial agonist mechanism of action on the DA D2 and 5-HT1A receptors (Shapiro et al., 2003), adolescent aripiprazole APD treatment was also found to elicit anxiolytic-like effects in adult rats. Such effects on anxiety, with no differences in locomotor activity or depressive-like behaviours, suggests a potential vital role of the 5-HT2A and 5-HT2C receptors in anxiety-like behaviour alone, as aripiprazole is known to have an antagonistic mechanism of action on the former, and a slightly lower affinity for the latter 5-HT receptor subtypes (Shapiro et al., 2003). Since olanzapine and risperidone also have similar antagonistic and agonistic mechanisms of action on the 5-HT2A and 5-HT2C receptors, respectively, and exhibit similar effects, this may provide further evidence for the potentially crucial role that mediating these individual receptor subtypes may have in regulating anxiety, although further studies investigating this are necessary.
With clear long-term depressive-like, anxiety behavioural alterations found in the present study, minor alterations to current methods, along with additional investigations, will provide further insight into the effects of childhood/adolescent APD use on behavioural attributes. Firstly, altering the social interaction protocol in order to pair naïve age- and sex-matched animals may reveal some potential differences in social behaviour that the current method of sex and treatment-matched animals may have occluded. Secondly, additional behavioural testing investigating the effects of early APD use on other behavioural parameters, including cognitive abilities, would be helpful in assessing any potential issues with short- and long-term memory in the adult rat, whilst also investigating the effects of other commonly-used APDs, including clozapine, would also increase the scope of the current investigation. Furthermore, it may also be of benefit for supplementary investigations to be completed on the effects of early APD treatment in disease animal models of mental illness, such as schizophrenia. Whilst the use of a standard rat strain in the present study is appropriate and clinically relevant, the inability to investigate, in a single animal model, the multiple illnesses (such as schizophrenia, bipolar disorder, autism, ADHD, Tourette’s disorder, anxiety disorder, borderline personality disorder and even hemiplegia) that APDs are used to treat (Memarzia et al. 2014; Olfson et al., 2010; Schneider et al., 2014), and the reported inadvertent exposure of healthy adolescent brains to APDs due to difficulties in diagnosis at a young age (Andersen and Navalta, 2011; Zuddas et al., 2011), increasing the scope of the study to investigate the long-term effects that early APD use has on individual illnesses will no doubt be beneficial.
In conclusion, this study indicates that early APD treatment results in long-term changes to locomotor, depressive-like and anxiety-like behaviours in adult rats. In addition, behavioural differences between genders were also observed in response to early APD treatment. Potential detrimental long-term behavioural changes following early APD treatment may outweigh the initial benefits of childhood/adolescent APD prescription and use. Whilst previous clinical studies have shown gender-based differences in APD treatment effects (Ratzoni et al., 2002), our study indicates that these differential effects may not only extend to behavioural attributes, but also into adulthood. Hence, careful consideration of both the pharmacology of the APD being prescribed, along with the gender of the child, may be crucial to reduce any potential long-term effects, including alterations to activity, anxiety or depressive-like behaviours. Further research is required in order to explore the potential physiological reasons behind the behavioural differences found across both treatment groups and genders, which have potentially resulted in the observed effects. Investigations into potential alterations to both the DA and 5-HT NT systems in key brain areas would provide further evidence of these potential detrimental long-term neurotransmitter alterations.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by an Australian NHMRC project Grant (APP1008473) to C Deng. M De Santis was supported by Australian Rotary Health in the form of an Ian Scott PhD Scholarship.