
Abstract
Neurotensin is a tridecapeptide originally identified in extracts of bovine hypothalamus. This peptide has a close anatomical and functional relationship with the mesocorticolimbic and nigrostriatal dopamine system. Neural circuits containing neurotensin were originally proposed to play a role in the mechanism of action of antipsychotic agents. Additionally, neurotensin-containing pathways were demonstrated to mediate some of the rewarding and/or sensitizing properties of drugs of abuse.
This review attempts to contribute to the understanding of the role of neurotensin and its receptors in drug abuse. In particular, we will summarize the potential relevance of neurotensin, its related compounds and neurotensin receptors in substance use disorders, with a focus on the preclinical research.
Keywords
Introduction
Neurotensin (NT), a tridecapeptide originally identified in extracts of bovine hypothalami by Carraway and Leeman (1973), is widely distributed in the central nervous system (CNS) and in the periphery, including the gut, pancreas and adrenal glands (Carraway and Leeman, 1976; Kitabgi et al., 1976). The biological actions of NT are initiated by binding to three different receptor subtypes: neurotensin receptor subtype 1 (NTS1), NTS2 and NTS3 (Geisler et al., 2006; Mazella and Vincent, 2006). NTS1 and NTS2 receptors belong to the family of G-protein-coupled receptors (Pelaprat, 2006). These NT receptor subtypes differ in their affinity for NT and their sensitivity to levocabastine, an antihistaminic compound.
The NTS1 receptor is levocabastine-insensitive, with a high affinity for NT; whereas the NTS2 receptor is levocabastine-sensitive, with a low affinity for NT (St-Gelais et al., 2006; Vincent et al., 1999). The NTS1 receptor is the best characterized NT receptor subtype. It is functionally coupled to the phospholipase C and the inositol phosphate signaling cascade, but its activation has been also associated with cyclic guanosine monophosphate, cyclic adenosine monophosphate and arachidonic acid production, along with mitogen-activated protein (MAP) kinase phosphorylation. The transduction mechanisms of the NTS2 receptor remain a matter of controversy (Mazella and Vincent, 2006). NT3 is structurally unrelated to these receptors, belongs to the family of sorting receptors and modulates NT intracellular signaling processes (Mazella and Vincent, 2006).
By interacting with its receptors, NT is known to exert several effects in mammals, including analgesia, hypothermia, neuroendocrine control of thyroid hormones, regulation of blood pressure and body weight homeostasis. NT is also deeply involved in immunity and inflammation; but its true role in these events still remains to be elucidated (Katsanos et al., 2008), as well as its relevance in cancer development and progression (Myers et al., 2009). When injected systemically, NT decreases blood pressure, gastric motility, gastric acid secretion and induces hyperglycemia.
The differential regional distribution of NT in the brain, the presence of NT receptors in different brain areas (such as cortical, striatal and limbic regions), the sodium- and calcium-dependent NT release and the involvement of NT in several electrophysiological and behavioral responses (Carraway and Leeman, 1973; Kitabgi et al., 1989; Saint-Gelais et al., 2006; Tanganelli et al., 1994; Vincent et al., 1999), suggest that this peptide acts as a neurotransmitter or neuromodulator in the mammalian CNS. In this context, the effects of NT include the well-documented interaction of the peptide with dopaminergic (DAergic) systems (Geisler al., 2006; Kitabgi et al., 1989; Rostène et al., 1992; Von Euler and Fuxe, 1987). This is mainly due to the ‘antagonistic’ action of the activated NTS1 on the dopamine (DA) D2 receptor recognition and signaling, via an intra-membrane NTS1/DA D2 receptor-receptor interaction (Antonelli et al., 2007b; Fuxe et al., 1992; Von Euler, 1991; Von Euler and Fuxe, 1987; Von Euler et al., 1991). In particular, pioneer binding studies demonstrated that, by means of an intra-membrane NTS1/DA D2 receptor-receptor interaction, NT reduced the affinity of the DA D2 receptor agonist (Agnati et al., 1983). More recently, it has been reported that the dynamic changes in DA D2 receptor signaling produced by NT receptor agonists may involve antagonistic allosteric receptor-receptor interactions in the NTS1/D2 receptor heteromers at the plasma membrane level (cAMP response element-binding protein (CREB) pathway) and synergistic interactions in PKC activation at the cytoplasmic level (mitogen-activated protein kinases (MAPK) pathway). The literature on NTS1/DA D2 receptor interactions and the possible relevance of this heterodimer in the CNS were recently reviewed (Ferraro et al., 2014; Tanganelli et al., 2012).
NT also appears to modulate the activity of other neurotransmitter circuits that are innervated by the DA system. Neural circuits containing NT were originally proposed to play a role in the mechanism of action of antipsychotic agents (Boules et al., 2007; Boules et al., 2014; Kinkead and Nemeroff, 2004; Nemeroff, 1980). In particular, the interaction between NT and DA has been implicated in the pathogenesis and treatment of schizophrenia (Boules et al., 2014; Caceda et al., 2006; LaCrosse and Olive, 2013; Nemeroff et al., 1983; Tanganelli et al., 2012). In fact, it has been reported that NT and the NT agonists possess neuroleptic-like properties in DA-mediated animal models of psychosis, such as amphetamine-induced locomotor activity, apomorphine-induced climbing and drug-induced disruption of pre-pulse inhibition (PPI) (Boules et al., 2001; Kalivas et al., 1983; 1984; Nemeroff et al., 1983; Shilling et al., 2003).
Besides DA systems, growing evidence suggests that NT may also play an important role in the modulation of aminoacidergic transmission in the basal ganglia and cerebral cortex (Antonelli et al., 2007a; Ferraro et al., 2008, 2009). In particular, NT amplifies glutamate transmission and enhances glutamate-induced excitotoxicity. The hypothesis was therefore introduced that NT may be involved in ischemic brain damage and neurodegenerative disorders (Antonelli et al., 2007b, 2008; Ferraro et al., 2009).
Several reviews on the relevance of NT in schizophrenia and neurodegenerative disorders have been published (Antonelli 2007b; Boules et al., 2014; Ferraro et al., 2008; 2009; Kinkead and Nemeroff, 2004; LaCrosse and Olive, 2013; Saint-Gelais et al., 2006; Tanganelli et al., 2012), underlining the evidence that the NT receptor agonists or antagonists may represent novel antipsychotic and neuroprotective drugs, respectively. In contrast, the important role played by central NT receptor mechanisms in drug addiction has only partially been reviewed (Boules et al., 2014; Ferraro et al., 2007), which is also true regarding their relevance for drug abuse treatment. This is probably due to the complex and partially controversial results emerging from the studies in this field. They demonstrate that the effect of NT depends on many parameters, such as: the dose used, the injection procedure and the NT receptor reached, as well as the experimental model or species used. Nevertheless, the possible implication of NT in drug abuse (Saint-Gelais et al., 2006) is supported by substantial experimental evidence and the similarities between certain NT- and psychostimulant drug-induced effects.
This review attempts to contribute to the understanding of the role of NT and its receptors in drug abuse, by also discussing the possible relevance of NTS1/DA D2 receptor heterocomplexes. To this purpose, the potential relevance of NT, its related compounds and NT receptors in substance use disorders will be summarized, focusing on preclinical research.
Neurotensin in the central nervous system
Biochemistry and localization
Within the CNS, NT is synthesized, stored at specific synapses and asynaptic varicosities and, under appropriate conditions, it may be released or co-released with classical neurotransmitters. NT and its structurally related hexapeptide neuromedin N (NN) are products of the same larger precursor, whose cDNA was cloned from bovine brain in 1987 (Dobner et al., 1987; Minamino et al., 1988). The precursor molecule, a highly conserved polypeptide of 169 or 170 amino acids in length, contains one copy each of NT and NN near the C-terminus; and it undergoes a differential tissue-specific cleavage at its four dibasic sites, by proprotein convertases (PCs). Therefore, pro-NT/NN may be processed to generate different sets of peptides. Four biologically active products of pro-NT/NN processing have been described: NT, NN, large NT and large NN (Kitabgi, 2010). In the brain, pro-NT/NN processing mainly depends on PC2 activity and leads to high amounts of NT and NN, and small quantities of large NT and large NN (Kitabgi, 2010). Using radioimmunoassay techniques, it was demonstrated that the regional distribution of NT and NN in brain tissues is, generally, the same (Kitabgi et al., 1992); however, marked differences in the ratio of NT over NN have been observed in different brain areas, with NT being generally more abundant in the DAergic regions such as the substantia nigra, pars compacta and ventral tegmental area (VTA).
Once processed as an active peptide in neurons, NT is stored in dense core vesicles and released in a calcium-dependent manner (Iversen et al., 1978). The physiological inactivation of NT is operated by endopeptidases (EPs) belonging to the family of metallopeptidases, which act on primary cleavage sites in the peptide sequence: Arg8-Arg9, Pro10-Tyr11 and Tyr11-Ile12 bonds. Three EPs are responsible for NT degradation: EP 24.11, EP 24.15 and in particular, EP 24.16, which is ubiquitously expressed (Kitabgi, 2006). Other exo- and endopeptidases further degrade the breakdown products generated by these metallopeptidases. Another mechanism that produces an inactivation of NT transmission is the process of NT internalization (Mazella and Vincent, 2006).
NT pathways
Several NT-containing neuronal circuits have been described in the rat. Among others (Saint-Gelais et al., 2006), these include the neurons projecting from:
The amygdala to the striae terminalis, the substantia nigra, pars compacta, the substantia nigra, pars reticulata and to the ventromedial nucleus of the hypothalamus;
The hippocampus through the cingulate cortex to the frontal cortex;
The hypothalamus, the ventromedial ventral pallidum, the dorsal raphe nucleus and the diagonal band of Broca to the VTA; and
The cells in the VTA to the nucleus accumbens (NAC), the amygdala, and especially the prefrontal cortex (PFC), from the striatum to the substantia nigra, pars reticulata.
High levels of the peptide are present in the hypothalamus, amygdala, bed nucleus of the striae terminalis, lateral septum, NAC, caudate-putamen and VTA (Geisler et al., 2006; Tyler-McMahon et al., 2000). Such a distribution of the peptide generally matches the distribution of the NT receptors in the brain (Geisler et al., 2006; Vincent et al., 1999).
NT and DA signaling: Focus on mesolimbic transmission
Berger et al. (1992) demonstrated that in the rat, but not in primates, NT is in part co-localized with DA in mesocortical neurons, but not in nigrostriatal and mesolimbic DA neurons, thus suggesting that NT might play a special role in the regulation of mesocortical DA transmission (Kalivas and Miller, 1984; Von Euler et al., 1990); however, the class of mesocortical DA projections that are not co-localized with NT in rat are particularly developed in human, as are the NT projections to the limbic system. These findings strengthen the role of NT corticolimbic innervations in primate brain. The mixed NT/DA neurons project to the PFC, entorhinal cortex, NAC, basolateral nucleus of the amygdala and lateral septum (Figure 1). NT/DA projections mostly overlap mesocorticolimbic DA projections with the exception of the central nucleus of the amygdala and the NAC core, where there are no mixed projections (Binder et al., 2001). NT fibers innervate the VTA and possibly originate from the rostral lateral septum, the pre-optic area and the lateral hypothalamus (Binder et al., 2001).
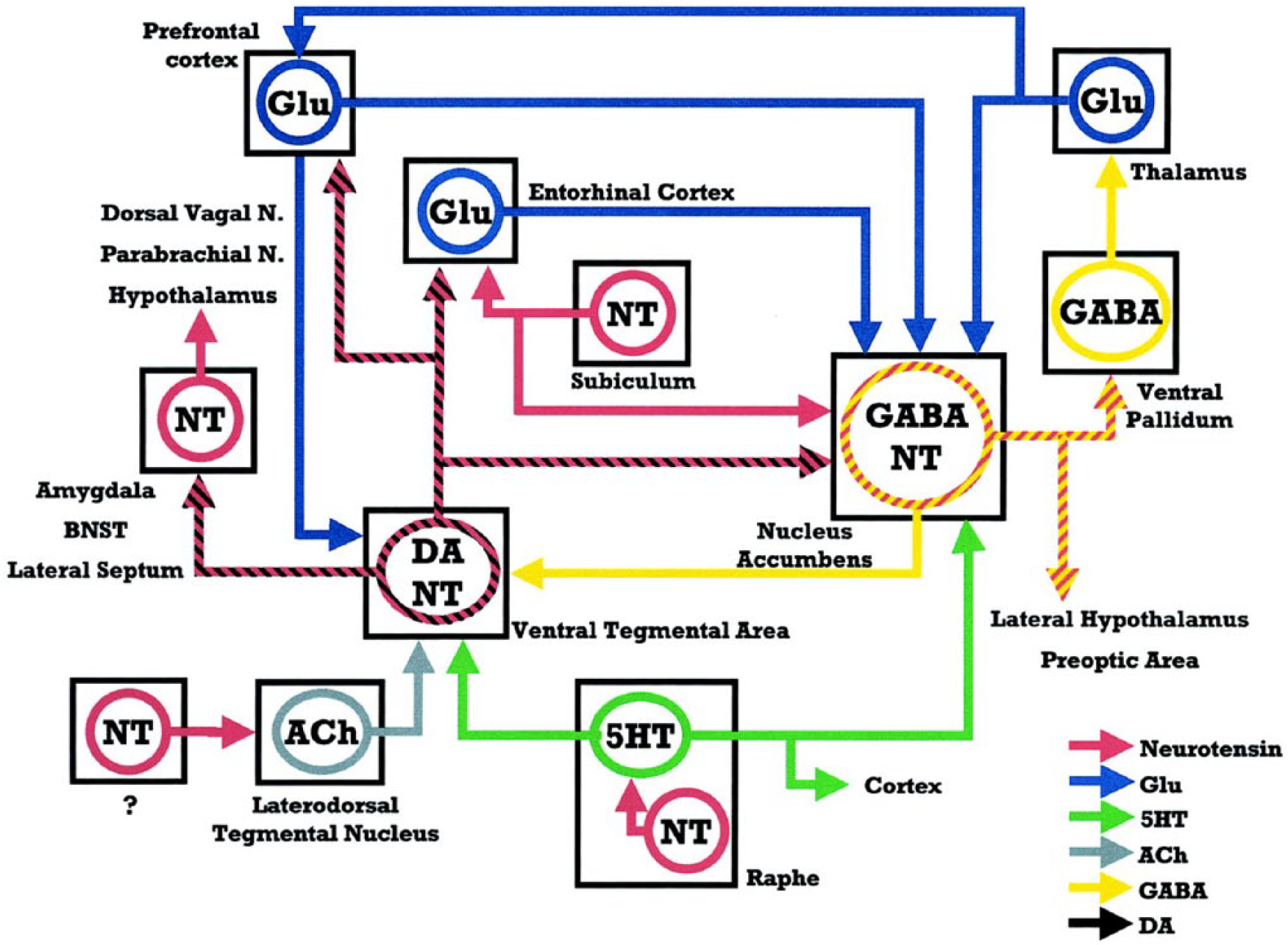
Schematic representation of direct and indirect associations between NT projections and the mesocorticolimbic DA system. The cross-hatched lines represent projections colocalizing either the NT and DA (black and red), or NT and GABA (yellow and red).
The activation of somatodendritic NTS1 receptors increases the firing rate of mesolimbic and mesocortical DAergic neurons (Werkman et al., 2000), most likely by increasing intracellular Ca2+ and reducing K+ conductances. NT administered to the VTA acutely excites DA neurons (Shi and Bunney, 1990; Werkman et al., 2000) and induces increased turnover and extracellular concentration of DA in the NAC (Kalivas and Duffy, 1990; Steinberg et al., 1995). Concerning the NAC, NT receptors are co-localized with postjunctional DA receptors on glutamate terminals and on soma-dendrites of striato-pallidal GABA neurons, although this matter is still controversial and inconsistent (Ferraro et al., 2007). Graphic representation can be seen in Figure 2.
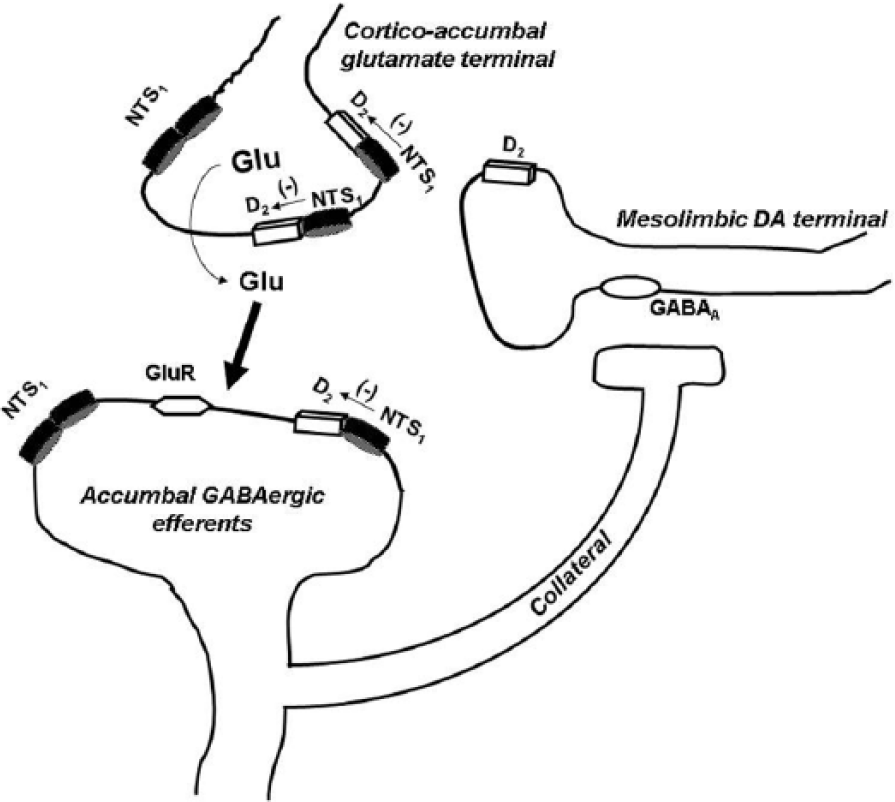
Schematic representation of the main location of the NTS1/D2 heteromers mediating the NTS1/D2 receptor-receptor interactions in the control of the ventral striato-pallidal GABA pathway from the nucleus accumbens. In the nucleus accumbens, there exists a dominance of the post-junctional antagonistic NTS1/D2 receptor-receptor interactions on the cortico-accumbens glutamate terminals over the weak antagonistic NTS1/D2 receptor-receptor interaction on the accumbens DA terminals (not shown). The intra-accumbens perfusion of NT antagonizes the inhibitory dopamine D2 tone on glutamatergic terminals via a NTS1/D2 receptor-receptor interaction, thus inducing an enhancement of glutamate outflow. Such an increase can then activate the inhibitory GABAergic signaling that could be responsible, via a collateral, for the significant reduction of DA release observed in the nucleus accumbens (Tanganelli et al., 1994); however, a direct involvement of NTS1 located on GABAergic neurons (Binder et al., 2001) cannot be excluded. The two D2/NTS1 heteromers on the glutamate terminals are to show that this is their major location, since only few are found on the striato-pallidal GABA neurons. NTS1 alone without D2 could be a monomer or a homomer, which is true also for D2 when not present in a heteromer.
Interestingly, NT was reported to either increase or decrease NAC DA transmission, depending on the dose (Boules et al., 2014; Ferraro et al., 2007). It was suggested that intra-NAC NT application preferentially modulates prejunctional DAergic transmission, mainly via indirect mechanisms involving other neuronal systems, rather than through a direct activation of the few NTS1 receptors located on the NAC DAergic terminals. In particular, it seems likely that the peptide, by activating NTS1 receptors mainly located on NAC glutamate terminals, induces an enhancement of extracellular DA levels. One of the possible mechanisms underlying this effect may be an indirect inhibitory action of the peptide on glutamate terminal DA D2 receptors (Agnati et al., 1983; Fuxe et al., 1992). The extracellular increase in NAC glutamate levels can then activate the inhibitory GABAergic signaling of dendrites and collaterals of the ventral striato-pallidal GABA pathway, which could be responsible for the significant reduction of extracellular DA levels observed in the NAC, as seen in Figure 2 (Tanganelli et al., 1994). This mechanism could be involved in the neuroleptic-like action of NT. In contrast to the NAC, NT increases DAergic signaling in the dorsal striatum, mainly via the activation of a relatively high density of NTS1 receptors located on striatal DA terminals involving inhibition of the DA D2 autoreceptors (Li et al., 1995; Tanganelli et al., 1994).
Neurotensin and substance use disorders
Substance use disorder refers to alcohol/drug abuse or alcohol/drug dependence (American Psychiatric Association, 2013). Impairments due to drugs of abuse start in the brain reward circuits, including the mesolimbic DA system (as all the addictive substances enhance DAergic transmission); while long-term drug intake leads to dysfunctions of the brain regions involved in learning and memory, habit-forming learning and inhibitory control, being under control of the glutamatergic or GABAergic neurotransmissions (Tzschentke and Schmidt, 2003). Wide NT distribution in the brain and its localization to DAergic rich areas, as well as its direct or indirect modulation of glutamatergic and GABAergic transmission, speak for the interaction of NT with drugs of abuse belonging to different chemical classes. Below, we describe and discuss the preclinical behavioral evidence (based on acute and repeated drug treatments) on the role of NT in chemical addictions.
Neurotensin and drug of abuse-evoked hyperlocomotion
Locomotor hyperactivity mainly depends on the stimulation of the meso-NAC DAergic neurons, projecting from the VTA to NAC, which constitutes the so-called ‘reward’ pathway (Filip and Siwanowicz, 2001; Hedou et al., 1999). Because locomotor hyperactivity was proposed as an index of the stimulatory effects of drugs of abuse (Phillips and Shen, 1996; Wise and Bozarth, 1987) that use the same neuroanatomical pathway to enhance their rewarding properties, the effects of NT on spontaneous and drug-induced locomotion will be summarized in the present section.
Several studies have focused on the investigation of the behavioral effects induced by acute NT administration, leading to somewhat controversial conclusions (Table 1). Thus, pioneering behavioral studies demonstrate that microinjection of NT into the NAC leads to a reduction of locomotion (Kalivas et al., 1984; Meisenberg and Simmons, 1985; Tanganelli et al., 2012). Conversely, the non-peptide NTS1 receptor antagonist SR48692 reduces both haloperidol-induced hypolocomotion and haloperidol reversal of amphetamine-induced hyperlocomotion. These findings point to the inhibitory role of NT on NTS1 receptors in locomotor activity.
Neurotensin and drug of abuse-evoked hyperlocomotion.
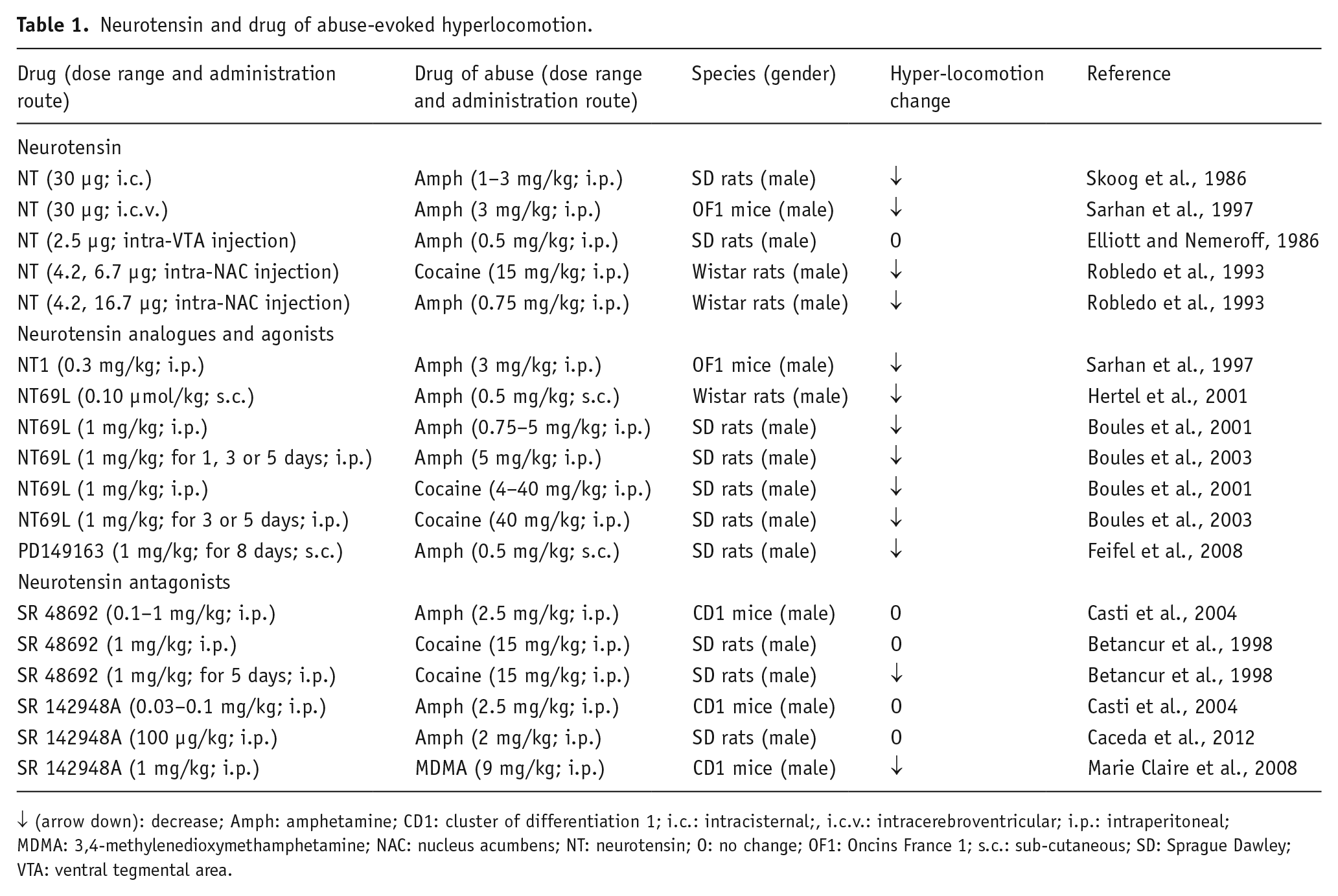
↓ (arrow down): decrease; Amph: amphetamine; CD1: cluster of differentiation 1; i.c.: intracisternal;, i.c.v.: intracerebroventricular; i.p.: intraperitoneal; MDMA: 3,4-methylenedioxymethamphetamine; NAC: nucleus acumbens; NT: neurotensin; O: no change; OF1: Oncins France 1; s.c.: sub-cutaneous; SD: Sprague Dawley; VTA: ventral tegmental area.
More recently, the involvement of NT signaling in the psychostimulant properties of drugs of abuse was extensively explored (Table 1). The intracerebroventricular (i.c.v.) or intracisternal (i.c.) administration of NT attenuated amphetamine-induced hyperlocomotion and reward behaviors induced by cocaine (Sarhan et al., 1997). Furthermore, intra-NAC injection of the peptide significantly decreased cocaine-induced and, at a higher dose, amphetamine-elicited hyperlocomotion. This suggested the ability of the peptide to modulate the psychostimulant-induced locomotor activation through an interaction with the mesolimbic DA system (Robledo et al., 1993; Sarhan et al., 1997). In line with the above observations, the acute systemic administration of the brain-penetrating NT analogue NT69L (N-methyl-l-Arg, L-Lys, L-Pro, L-neo-Trp, L-tert-Leu, L-Leu) reduced spontaneous locomotor activity, as well as the hyperlocomotion caused by both amphetamine and cocaine; however, the inhibitory effect of NT69L was lost, following repeated drug administration, due to a desensitization of NT receptors (Boules et al., 2001, 2003; Hertel et al., 2001).
Other investigators demonstrated that the i.p. injections of NTS1, a brain-penetrable NT-related peptide, decreases the amphetamine-stimulated hyperlocomotion (Sarhan et al., 1997); whereas the systemic administration of PD149163, a selective and potent NTS1 receptor agonist, counteracted the locomotor effects of amphetamine in both acute and subchronic regimens (Feifel et al., 2008). In contrast to the above-cited literature, other studies suggest that some behavioral effects of NT are similar to those induced by peripheral administration of psychomotor stimulants (Boules et al., 2013; Richelson et al., 2003); however, such excitatory locomotor effects are associated with results obtained with intra-VTA NT microinjection, and are abolished by both i.c.v. injection of haloperidol and the destruction of the meso-NAC DAergic pathway (Kalivas and Taylor, 1985; Kalivas et al., 1983). Thus, it was proposed that NT stimulates locomotor activity through activation of DA neurons, when it is injected at the level of the VTA DAergic cell bodies (Kalivas and Duffy, 1990; Kalivas et al., 1983). Instead, the peptide inhibits locomotion through a post-synaptic modulation of neuronal systems regulated by NAC DA terminals (Kalivas et al., 1984). The latter effects involve an inhibitory action exerted by NTS1 receptor activation on D2 receptor function, through a post-junctional NT-D2 receptor-receptor interaction (Fuxe et al., 1992); while in the VTA, the existence of NTS1-D2 autoreceptor complexes on DA nerve cells has been proposed (Borroto-Escuela et al., 2013). NTS1 receptors inhibit DA D2 autoreceptor signaling in such heterocomplexes; and thereby, increase the firing in the DA neurons (Ferraro et al., 2014).
Other studies investigated the effects of NT receptor antagonists, such as SR48692 and SR142948A, on locomotor hyperactivity after acute treatment with drugs of abuse. SR48692 is widely used as a NTS1 receptor antagonist, as it displays higher affinity for NTS1 than for NTS2 receptors (Boules et al., 2006). Some authors also report that SR48692 acts as an agonist at NTS2 receptors (Botto et al., 1997; Richard et al., 2001; Vita et al., 1998). SR142948A is considered a non-selective NT receptor antagonist (Bose et al., 2015); however, Vita et al. (1998) provided evidence that SR142948A acts as an agonist, not an antagonist, at NTS2 receptors. Neither SR48692 nor SR142948A had any effect on mouse spontaneous motor activity or amphetamine- and cocaine-elicited hyperlocomotion (Betancur et al., 1998; Caceda et al., 2012; Casti et al., 2004). This suggests that endogenous NT does not modulate spontaneous and drug-induced locomotion. On the other hand, repeated systemic injection of SR48692 reduced the behavioral response to acute cocaine (Betancur et al., 1998); and the acute systemic injection of SR142948A blocked the hyperlocomotion elicited by 3,4-methylenedioxymethamphetamine (MDMA), providing evidence for the possible involvement of the neuropeptide in the behavioral activation induced by some psychostimulants through the release of NT at the level of the DA cell bodies in the ventral midbrain.
In conclusion, the above results led us to hypothesize that NT analogs or selective NTS1 receptor agonists that cross the brain blood barrier after their systemic administration might be useful, clinically in modulating hyperactivity and certain behavioral responses to drugs of abuse (Boules et al., 2006; Vadnie et al., 2014); however, some of these effects seem to be reachable also by using NT receptor antagonists (Betancur et al., 1998) and other studies argue against the involvement of endogenous NT signaling in the hyperlocomotion elicited by psychostimulant drugs (Hall et al., 2012). It seems possible that this could be due to the use of unselective NT receptor ligands, the different neuronal localization of NT receptors in several brain areas and the heteromeric receptor complexes, in which NT receptor protomers participate, that under different addiction phases may evoke an opposite action; however, further and more extensive investigations are necessary to possibly clarify the role of the peptide in drug of abuse-evoked locomotor behaviors.
Neurotensin and drug of abuse-evoked sensitization
Repeated, intermittent administrations of drugs of abuse induce a strong enhancement in locomotor stimulation that may last for a long time. This phenomenon, also called locomotor sensitization or behavioral sensitization, has been suggested to predict the addictive property of a drug combined with forms of neuronal adaptations linked to an enhancement of the reinforcing and motivational aspects of drugs of abuse. Two separate temporal domains of drug-induced sensitization within neuronal networks have been identified and termed initiation and expression. The initiation of behavioral sensitization to psychostimulants is operationally defined as the transient sequence of cellular and molecular events precipitated by psychostimulant administration, which ultimately leads to enduring changes in neural function.
Expression is defined as the enduring neural alterations arising from the initiation process that directly mediate the sensitized behavioral response (Robinson and Berridge, 1993; Steketee and Kalivas, 2011). Initiation and expression of locomotor sensitization are reported to have distinct neurochemical mechanisms, and different brain structures are involved in the two temporal phases: initiation of behavioral sensitization to psychostimulants is mostly related to DAergic and glutamatergic VTA transmission, while the neuronal changes associated with sensitization expression are mainly localized among the interconnection between VTA, NAC, PFC and amygdala (the so-called ‘motive circuit’), which leads to a drug-induced increase in DA and glutamate release in the NAC. Moreover, the neuronal adaptations involved in the expression of behavioral sensitization to different drugs of abuse are shown to be distinct in the motive circuit (i.e. cocaine-elicited sensitization appears to involve more descending corticofugal excitatory efferents than amphetamine) and result in an altered balance between the interconnections of several neurotransmitter systems (Pierce and Kalivas, 1997). Numerous studies have evaluated the possible involvement of NT in psychostimulant sensitization, giving mixed results (Tables 2(a), 2(b) and 2(c)).
Neurotensin and drug of abuse-evoked sensitization development.
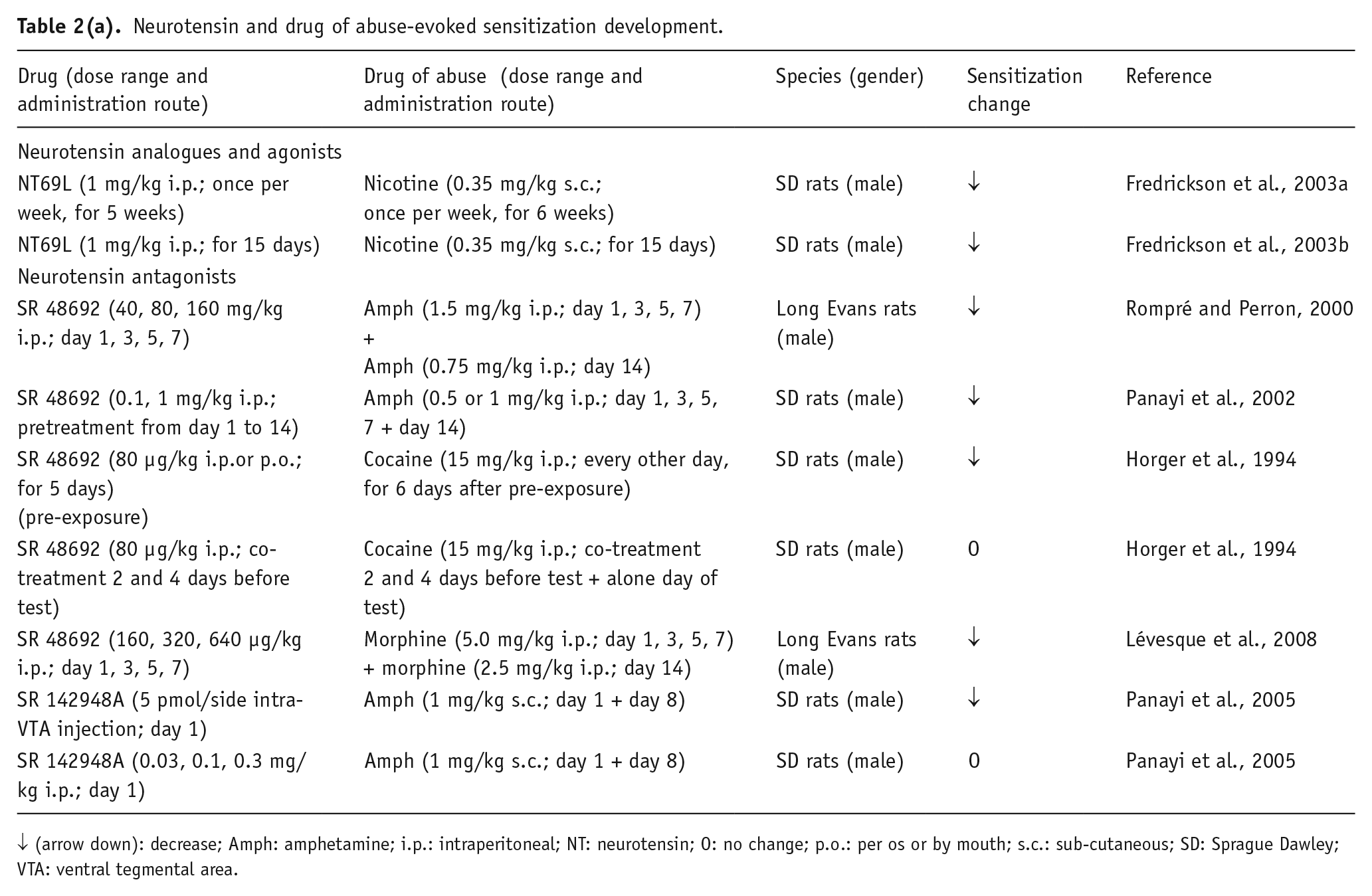
↓ (arrow down): decrease; Amph: amphetamine; i.p.: intraperitoneal; NT: neurotensin; O: no change; p.o.: per os or by mouth; s.c.: sub-cutaneous; SD: Sprague Dawley; VTA: ventral tegmental area.
Neurotensin and drug of abuse-evoked sensitization expression.

↓ (arrow down): decrease; Amph: amphetamine; i.p.: intraperitoneal; NT: neurotensin; O: no change; s.c.: sub-cutaneous; SD: Sprague Dawley; VTA: ventral tegmental area.
Neurotensin and drug of abuse-evoked sensitization, cross-sensitization.
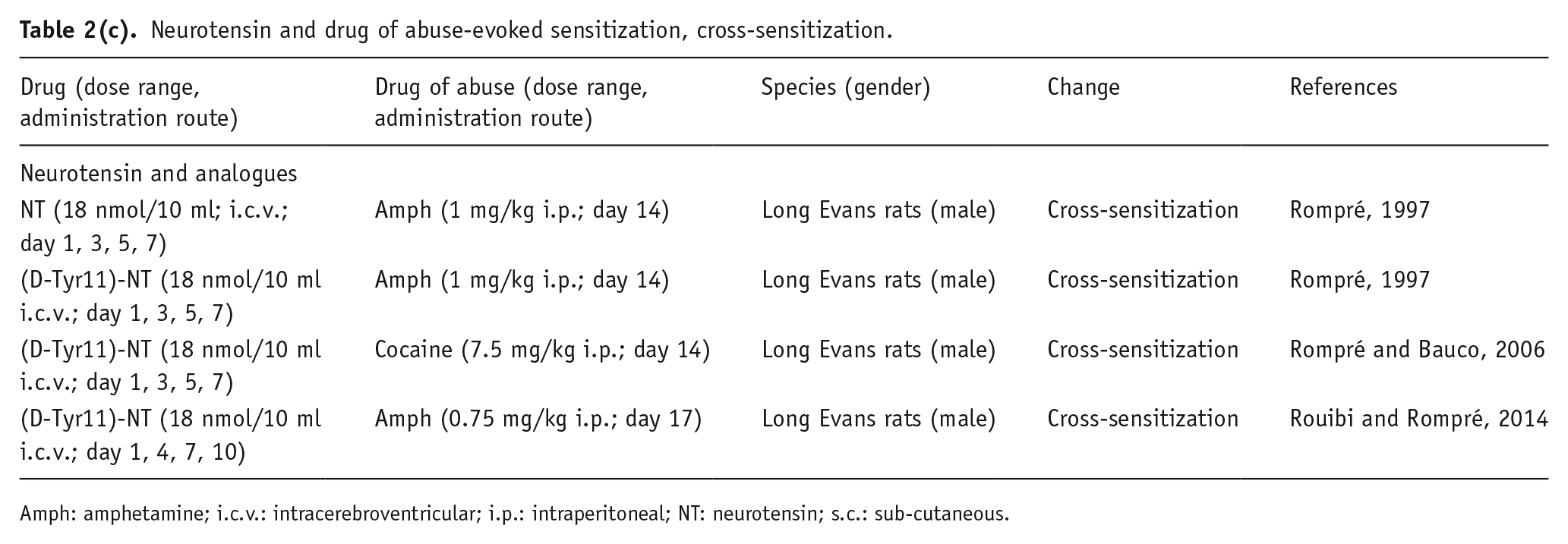
Amph: amphetamine; i.c.v.: intracerebroventricular; i.p.: intraperitoneal; NT: neurotensin; s.c.: sub-cutaneous.
Development of sensitization
The first evidence indicated that chronic pre-exposure to the NTS1 receptor antagonist SR48692 (without cocaine injections) before the development of cocaine sensitization reduced the latter phenomenon, whereas the same drug given daily before cocaine injections had no effect on cocaine sensitization (Betancur et al., 1998; Horger et al., 1994). On the other hand, SR48692 dose-dependently attenuated and/or prevented the development of sensitization to amphetamine (Panayi et al., 2002; Rompré and Perron, 2000), pointing to the role of NAC NTS1 receptors in attenuating the sensitizing locomotor actions of such drugs of abuse. Interestingly, the other NTS1/2 receptor antagonist, SR142948A, acutely administered with amphetamine, failed to effect the acquisition of drug sensitization (Panayi et al., 2005). These discrepancies have been proposed to be due to the different drug administration routes, the drug treatment schedule or to other methodological variables (Panayi et al., 2005; Rompré and Perron, 2000). Also, differences in their ability to counteract DA D2 postjunctional versus autoreceptor signaling in heterocomplexes should be considered, as well as the NT receptor antagonist selectivity toward NTS1 or NTS1/2 receptors.
Based on these data, it can be speculated that NT might act on some neural mechanisms that support the neuroadaptations underlying psychostimulant drug sensitization. To support this hypothesis, it was reported that NT is released in the VTA after amphetamine injection; and that it contributes, although not sufficiently, to the development of amphetamine behavioral sensitization (Panayi et al., 2005). An increase in ventral midbrain DA release seems to be critical for this NT action. In fact, cocaine and amphetamine sensitization are augmented by compounds that increase extracellular catecholamine levels (Kalivas and Weber, 1988); and the stimulation of ventral midbrain NT receptors enhances neuronal DA firing and release (Kalivas and Duffy, 1990; Kalivas and Taylor, 1985). It is also possible that NT might produce psychostimulant sensitization by activating its receptors in the medial PFC, a brain area receiving NT afferents (Fatigati et al., 2000; Petrie et al., 2005; Rompré et al., 1998; Sesack and Pickel, 1992).
Although the available literature was focused on NT receptor antagonism in the development of drug sensitization, limited findings indicate the effects of NT receptor agonists on this phenomenon. Thus in rats, NT69L significantly reduced the initiation of amphetamine- and cocaine-induced behavioral sensitization (Fredrickson et al., 2003a, 2014). A local (intra-ventral pallidum) injection of the biologically active NT(8–13) fragments before systemic cocaine injection did not alter locomotor responses to a cocaine challenge, reducing the significance of the endogenous pallidal release of NT for the development of cocaine sensitization (Torregrossa and Kalivas, 2008).
Expression of sensitization
Opposite to the controversial data obtained in studies on sensitization development, the results concerning the role of NT in the expression of psychostimulant drug sensitization consistently provided evidence that the blockade of NT receptors decreasesthis process. In fact, the acute administration of SR48692, priorto amphetamine challenge doses, prevented the expression of amphetamine behavioral sensitization in rats that were pre-exposed to the psychostimulant (Costa et al., 2001). Furthermore, given daily after the amphetamine exposure period necessary to sensitize the animals to the behavioral effects of the drug, SR48692 reverted the expression of amphetamine-induced sensitization (Costa et al., 2007). The same antagonist, chronically administered after a cocaine regimen during the withdrawal period, was effective in attenuating the expression of cocaine-elicited sensitization (Felszeghy et al., 2007).
When injected into the VTA, the other NT receptor antagonist, SR142948A, failed to alter the expression of amphetamine sensitization; however, this effect is in line with the accepted notion that the VTA by itself is not an anatomical substrate for expression of drug sensitization (Panayi et al., 2005). Even though most studies focus on the expression of behavioral sensitization to psychostimulant drugs mainly considered for the role of DA transmission, the imbalance between some neurotransmitter pathways, such as the glutamatergic and GABAergic ones, could contribute to this process (Pierce and Kalivas, 1997; Vanderschuren and Kalivas, 2000).
NT receptors are localized in most of the structures of the motive circuit; and that neuropeptide is known to interact with neuronal pathways involved in drug addiction (Liang et al., 2008; Picciotto and Corrigall, 2002). Thus, the NT-induced modulation of different neurotransmitter systems in these brain regions might be involved in the role that the peptide plays in the expression of psychostimulant sensitization; however, it is worth noting that systemic administration of both NT receptor agonists and antagonists have been reported to produce an attenuation of the expression of drug-elicited sensitization. This issue needs, therefore, to be deeply investigated by future studies, to better clarify the mechanisms through which the different NT receptor ligands exert these effects.
Finally, cross-sensitization between NT and psychostimulants has been observed. Specifically, it was shown that repeated, intermittent i.c.v. injections of NT in rats produce sensitization to the behavioral stimulant effect of systemic amphetamine, while the more potent NT analogue D-Tyr[11]NT, repeatedly administered by i.c.v., increases both cocaine and amphetamine sensitization (Rompré, 1997; Rompré and Bauco, 2006). This action, at least in the case of amphetamine sensitization, was demonstrated to be either context-dependent or context-independent, based upon the pattern of locomotor activity (Rouibi and Rompré, 2014); and to be prevented by excitotoxic lesions of the PFC (Blackburn et al., 2004). Thus, given the long-lasting changes in responsiveness to psychostimulant drugs after repeated activation of the NT systems, it was reasonable to conclude that NT could act in the same neural mechanisms that induce some neuroadaptations related to drug sensitization; however, further investigation is required to better understand the exact role, the anatomical sites, and the functional and molecular mechanisms through which NT mediates these effects.
Neurotensin and drug of abuse reward
Some evidence for the rewarding properties of NT and/or its ability to affect the reinforcing behaviors induced by drugs of abuse has been provided by studies using different experimental procedures, such as drug self-administration, drug preference and conditioned place preference (CPP) in rodents (Table 3 and Table 4).
NT and drug of abuse-evoked CPP.
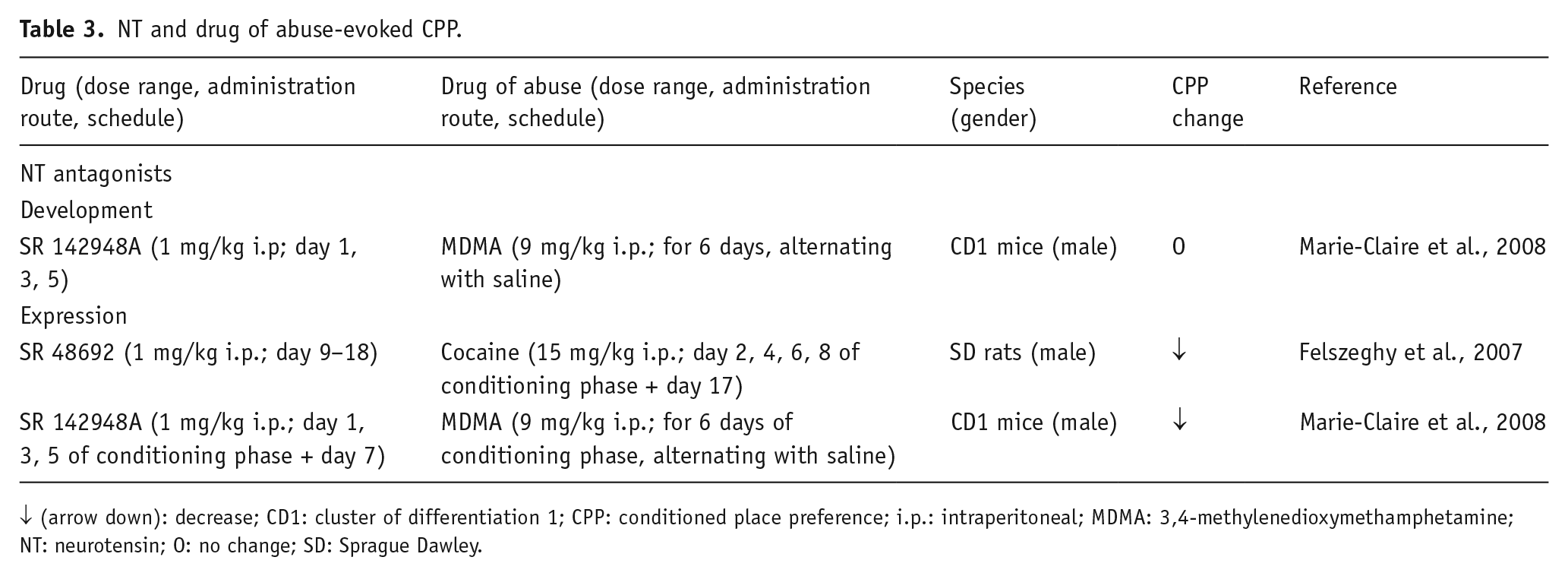
↓ (arrow down): decrease; CD1: cluster of differentiation 1; CPP: conditioned place preference; i.p.: intraperitoneal; MDMA: 3,4-methylenedioxymethamphetamine; NT: neurotensin; O: no change; SD: Sprague Dawley.
Neurotensin and drug of abuse-evoked self-administration.
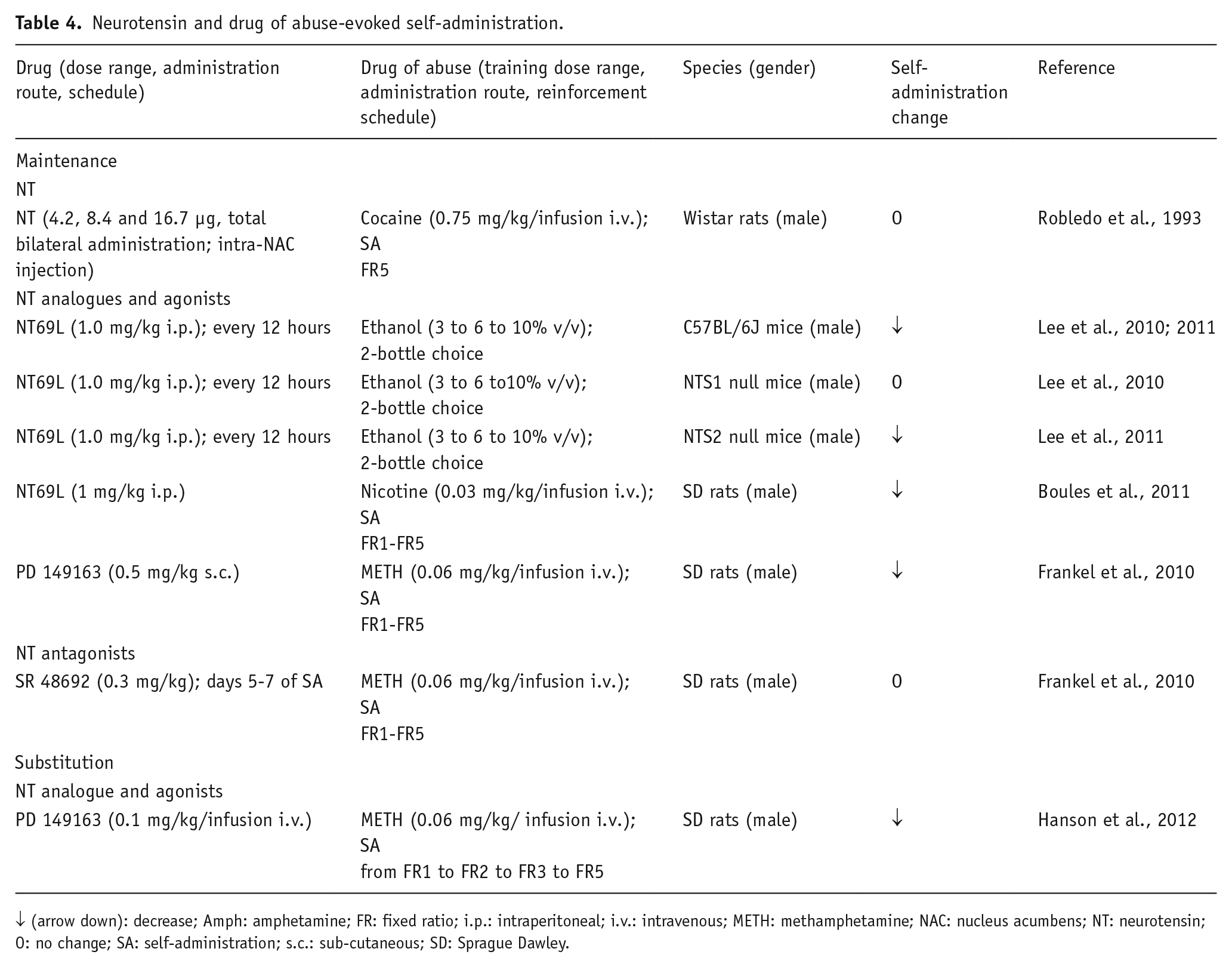
↓ (arrow down): decrease; Amph: amphetamine; FR: fixed ratio; i.p.: intraperitoneal; i.v.: intravenous; METH: methamphetamine; NAC: nucleus acumbens; NT: neurotensin; O: no change; SA: self-administration; s.c.: sub-cutaneous; SD: Sprague Dawley.
Drug self-administration
Early investigations using the self-administration paradigm demonstrated that rats performed operant tasks to obtain intra-VTA infusions of NT, thus suggesting that the peptide by itself exerts a primary positive reinforcement in this brain area (Glimcher et al., 1984, 1987). These findings are in line with previous results obtained in CPP experiments; however, when NT was injected into the NAC immediately before cocaine SA in the maintenance phase, no effect of the peptide on cocaine self-infusions was found, whereas the administration of the peptide in the same region attenuated the drug-elicited locomotor activity (Robledo et al., 1993). It is worth noting that, in the latter study, NT was administered into the NAC core that could be more involved in motor than in motivational aspects. In fact, the projections from the NAC core mainly innervate the ventromedial central pallidum, which participates in the indirect pathway to the subthalamic nucleus and the substantia nigra, pars reticulata. Instead, the projections from the NAC shell reach the dorsolateral ventral pallidum, which innervates the medio-dorsal thalamic nucleus, sending efferents to the prefrontal cortex (Fuxe et al., 2008), VTA and the lateral hypothalamus (Heimer et al., 1991; Kalivas and Miller, 1984; Zahm and Heimer, 1988, 1990). The microinjection of NT(8–13) into the ventral pallidum did not affect cocaine self-administration (Torregrossa and Kalivas, 2008).
On the other hand, the NT receptor agonist PD149163 blocked intravenous (i.v.) mephedrone self-administration (German et al., 2014) or i.v. methamphetamine (METH) self-injection in rats (Frankel et al., 2011; Hanson et al., 2012). Of note, in a substitution test of METH SA, the PD149163 was not self-administered by the animals, which excludes rewarding properties (Hanson et al., 2012). These authors proposed that self-administration extinction is associated with increases in NT release involving a DA D2 receptor mechanism in the NAC (Antonelli et al., 2007b; Borroto-Escuela et al., 2013).
Taken together, the data from combination and substitution SA paradigms provide evidence for the involvement of NT signaling in regulating psychostimulant consumption and suggest that the activation of NT receptors generally suppresses operant behaviors linked with drugs’ SA.
It is well-known that the mesolimbic DA system, where NT also exists in its mesocortical component (Saint-Gelais et al., 2006; Von Euler et al., 1990) is the main anatomical substrate for the rewarding effects of drugs of abuse (McBride et al., 1999). NT is associated with inhibitory feedback actions on basal ganglia and limbic DA pathways, and elevated extracellular NT levels have been reported in both the NAC and dorsal striatum, after treatment with high doses of psychostimulants (German et al., 2014; Hanson et al., 2012). The mechanisms underlying the ability of NT receptor agonists to reduce the reinforcing effects in i.v. self-administration and two drugs choice experiment seem complex, but are possibly related to the antagonistic interaction of the NTS1 receptor with the DA D2 receptor, reducing post-junctional DA D2 receptor signaling.
Based on results of the above-cited SA studies, which were focused mainly on the effects of psychostimulants, it seems likely that receptor-receptor interactions in the NAC NTS1-D2 heteroreceptor complexes contribute to counteract the reward properties of psychostimulants. Thus, an increased activity in the ventral striato-pallidal GABA neurons is obtained, which represents an anti-reward system.
Conditioned place preference
CPP is widely used to study the motivational and reinforcing effects of both natural stimuli and drugs of abuse (Tzschentke, 2007). This paradigm is based upon the acquisition of preference for neutral environmental stimuli (conditioned or secondary reward) that were previously combined with a drug administration (primary reward). The CPP comprises an acquisition phase and an expression phase, in which drug-free animals are tested for their preference for the environment previously paired with the drug. It was reported that the acquisition and expression of this incentive learning are mediated by different neurochemical mechanisms. For example, the neurons involved in the expression and acquisition of CPP in the case of amphetamine and morphine are anatomically distinct, at least within the NAC (Fenu et al., 2006; Marie-Claire et al., 2008; Sellings and Clarke, 2003).
An early study reported that intra-VTA NT was associated with CPP acquisition and expression, thus suggesting that the peptide acts as a primary reinforcement in rats (Glimcher et al., 1984). More recently, it was demonstrated that NT displays positive reinforcing actions, when microinjected into the central nucleus of the amygdala and ventral pallidum, and that these effects were mediated by NTS1 receptors, because they were prevented by prior administration of selective receptor antagonists (Laszlo et al., 2010; Ollmann et al., 2015).
Other studies investigated the role of NT in drug of abuse-induced CPP. Pre-treatment with the NTS1/2 receptor blocker SR142948A did not modify the development of, but suppressed the expression of, MDMA-elicited CPP in mice (Marie-Claire et al., 2008); while chronic treatment with the more selective NTS1 receptor antagonist SR48692 blocked cocaine-induced CPP in rats (Felszeghy et al., 2007). These findings suggested that endogenous NT might participate in some behaviors elicited by drugs of abuse and it appears to be mainly involved in the expression of psychostimulant-induced CPP. Interestingly, the mouse striatal NTS1 receptor mRNA levels were up-regulated when the MDMA-induced CPP expression was tested. Based on these results, it was proposed that NTS1 receptors may be involved in the behavioral consequences induced by the conditioned environmental reward, without directly mediating the reinforcing actions of drugs of abuse. Thus, NT could be a neural target for reward expectation (Marie-Claire et al., 2008); however, the rewarding properties of cocaine, as explored in the CPP paradigm, using NT knockout (KO) mice, are reported to be similar to those observed in wild-type (WT) animals, indicating that endogenous NT is not required for the cocaine-induced CPP and for the rewarding effect of the drug (Hall et al., 2012). Nevertheless, the authors did not exclude that NT could be involved in other aspects of cocaine-elicited CPP, such as the retention of CPP following cocaine conditioning (agreeing with Felszeghy et al., 2007) not explored in NT KO mice.
Drug preference
The drug preference procedures have been mainly used to assess the involvement of NT in ethanol-mediated physiological and behavioral changes (Erwin and Su, 1989; Erwin et al., 1997; Luttinger et al., 1982; Widdowson, 1987); however, the functional role of NT and its receptors in ethanol preference has not been sufficiently established. The activation of NTS1 receptor signaling appears to be inversely correlated with ethanol preference (Table 5). In fact, the systemic administration of the NT analogue NT69L significantly reduced ethanol preference, in both WT and NTS2 receptor KO mice, in a 2-bottle choice experiment, while the NTS1 KO mice were insensitive to this effect (Lee et al., 2010, 2011). In line with these findings, rats that are alcohol-preferring show a lower concentration of PFC NT, in comparison with non-alcohol-preferring rats (Ehlers et al., 1999). In addition, either NTS1 or NTS2 KO mice display similar elevated ethanol intake, when compared to WT animals. It has been proposed that NTS1 receptors are involved in the effect of lower, ataxic doses of ethanol, whereas NTS2 receptors might be responsible for the effect of higher, hypnotic doses of ethanol (Lee et al., 2011). Concerning the possible mechanism of action, NT69L might prevent ethanol consumption through the modulation of both DAergic and glutamatergic systems that are implicated in ethanol addiction (Li et al., 2011). Taken together, these data suggest that there is a potential therapeutic use of NT analogues in alcohol use disorder.
Neurotensin and drug preference.
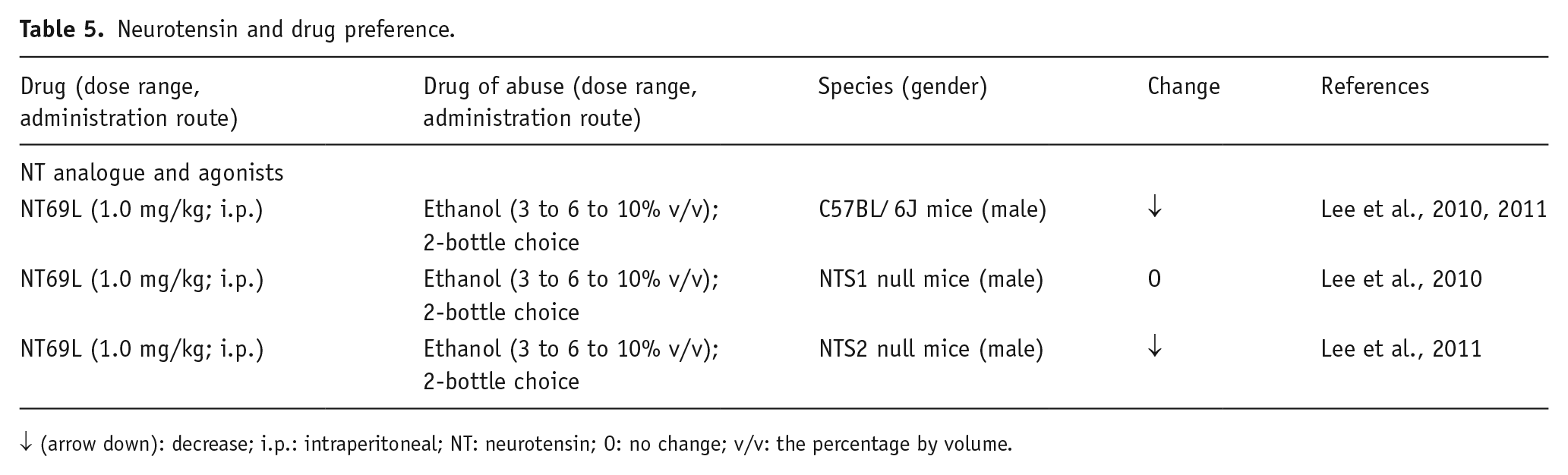
↓ (arrow down): decrease; i.p.: intraperitoneal; NT: neurotensin; O: no change; v/v: the percentage by volume.
Neurotensin and self-stimulation
Reward stimuli include, among others, electrical stimulations of some brain areas. In the paradigm of brain stimulation reward (BSR), rats rapidly learn an operant task, in order to electrically stimulate their own brain in specific sites that are mostly under control of DA neurotransmission. Consistent data show that BSR is modified by the administration of drugs of abuse, whose mechanisms of action were known to be related to a modulation of CNS DA transmission. Experiments using the self-stimulation procedure indicated that NT is involved in the control of behaviors motivated by positive reinforcement (Kempadoo et al., 2013; Rompré et al., 1992, 1995; Rompré and Boye, 1993). The evidence is supported by consistent data that indicate the anatomical and functional interactions between the DA brain reward system and the NT pathways seen in Figure 1 (Binder et al., 2001; Boules et al., 2013; Geisler et al., 2006; Saint-Gelais et al., 2006).
The microinjection of NT into the ventral mesencephalic region, produced a dose-dependent reduction in the stimulation frequency necessary to sustain the threshold levels of responding for BSR. The activation of ventral mesencephalon NT receptors induced DA neuron firing and the neurotransmitter release in limbic terminal fields; thus providing evidence that NT enhances reward-related activity in DA cells (Rompré et al., 1992), potentially through antagonistic NTS1-DA D2 autoreceptor interactions. Moreover, an augmentation of BRS is found when the peptide is i.c.v. injected, and this effect is associated with a significant time-dependent decrease in frequency threshold, in rats. These results provide evidence for psychostimulant-like effects of centrally-administered NT.
On the other hand, i.c.v. administration of the peptide was also found to suppress the maximal rate of response that is a typical action of neuroleptics. These results can be explained by the fact that i.c.v.-injected NT can also reach NAC and produce a reduction of postjunctional DA D2 receptor signaling, possibly via a NTS1-D2 receptor-receptor interaction (Borroto-Escuela et al., 2013). Of note, at a low dose NT tended to decrease the threshold; whereas at a higher dose, even a small increase was found immediately after the i.c.v. NT injection that might reach the NAC. Opposite effects of different doses of NT were also observed on spontaneous locomotion (Nouel et al. 1990), which may also be explained according to this proposal.
Recent work by Kempadoo et al. (2013) also reported that NTS1 receptors alter intracranial self-stimulation from the lateral hypothalamus to VTA in mice, as the selective antagonist SR48692 was found to decrease the amount of reinforced behavior, although maintaining lower levels of stimulation-seeking. The proposed mechanism involves NT-mediated enhancement of reward-related behavior via excitatory glutamate transmission on midbrain DA neurons, as well as by NT-mediated reduction in the D2 autoreceptor function in the VTA. Considering this evidence, it seems likely that the NT receptor antagonists might counteract human forms of pathological reward-seeking, among other mechanisms, through an action on the NTS1-D2 autoreceptor complex in the VTA DA neurons.
Neurotensin and drug-seeking behavior
Recently, the role of NT in drug-seeking behavior and, specifically, in a relapse to drug-seeking following a prolonged drug-free period, has been investigated by a few authors (Table 6). In experimental models of drug-seeking, the animals were trained to self-administer a drug (reinforce); then the reinforcement was removed to extinguish this behavior, and finally, the drug-seeking was restored by the presentation of a stressor, a drug-associated cue (sound and/or light), or the drug itself (Torregrossa and Kalivas, 2008).
Neurotensin and reinstatement of drug-seeking behavior.
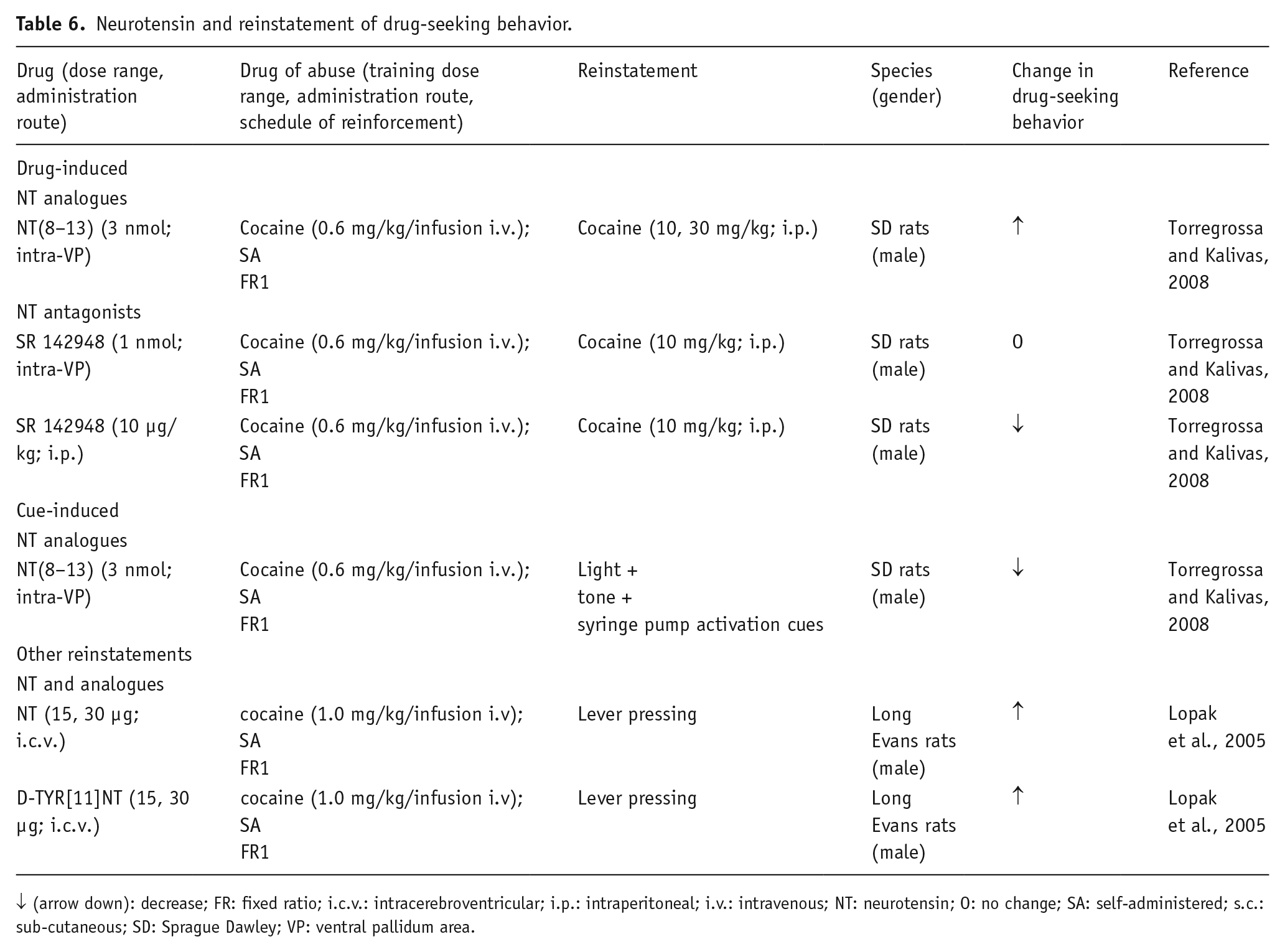
↓ (arrow down): decrease; FR: fixed ratio; i.c.v.: intracerebroventricular; i.p.: intraperitoneal; i.v.: intravenous; NT: neurotensin; O: no change; SA: self-administered; s.c.: sub-cutaneous; SD: Sprague Dawley; VP: ventral pallidum area.
Using this paradigm, both NT and its analogue D-TYR[11]NT, injected i.c.v. prior to the reinstatement test in rats, were found to produce a robust reinstatement of cocaine-seeking behavior; and this effect was attenuated by a prior injection of a DA D1/D5 receptor antagonist. Interestingly, the same local pre-treatment with D-TYR[11]NT did not affect the reinstatement of sucrose-seeking in the rat.
The lack of efficacy of NT in non-drug (sucrose) reinstatement, in comparison to the cocaine-reinstatement, suggested the hypothesis that during the psychostimulant SA, certain neuronal adaptations in some brain regions lead NT to enhance the vulnerability to relapse, after a drug-free period (Lopak and Erb, 2005). Other data demonstrated that the active NT fragment NT (8–13), injected into the intra-ventral pallidum before the reinstatement test for cocaine-seeking, inhibited the cue-induced reinstatement (Torregrossa and Kalivas, 2008). This is likely related to NT fragment-induced GABA release from the striato-pallidal GABA terminals, via antagonistic NTS1-D2 interactions in those terminals. The same authors showed an unexpected augmentation of cocaine-primed reinstatement (Torregrossa and Kalivas, 2008). This profile of action is surprising, because both the cue- and the cocaine-induced reinstatements are mediated by ventral pallidum neurotransmission. At present, it is difficult to explain these results. It may be speculated that cocaine treatment has reorganized the postjunctional NTS1-DA D2 complexes of the striato-pallidal GABA neurons into producing facilitatory NTS1-DA D2 receptor-receptor interactions, which can explain the augmentation of the cocaine-induced reinstatement. Finally, only one dose of NT (8–13) was tested in the reinstatement experiments; thus, it cannot be excluded that other doses of the NT analogue might have different effects, so further experiments will be necessary to clarify this possibility. It is worth noting that when given systemically, but not when directly administered into the VP, the NTS1/2 receptor antagonist SR142948 blocked the cocaine-induced reinstatement, excluding the significance of ventral pallidum NT receptors to this behavioral effect (Torregrossa and Kalivas, 2008).
Taken together, these data indicated that NT systems seem to play a role in drug-seeking behavior, but additional studies are requested to deeply investigate the involvement of the neuropeptide in this model of drug relapse, and the exact mechanisms underlying the brain interactions between NT and other neurotransmitter systems during drug-induced seeking behavior.
Neurotensin and drug withdrawal
Limited data show that NT may alter withdrawal symptoms following chronic exposure to some drugs of abuse. Thus, the NT analog NT(8–13) decreased behavioral signs of the withdrawal syndrome in rats that were repeatedly treated over 5 days with increasing doses of morphine, followed by the opioid receptor antagonist naloxone. Such behavioral reductions were due to NT(8–13) restoration of decreased DA and serotonin turnover in brain areas (Konstatinopolskii et al., 2013). Another report showed no effects of NT (i.c.) on ethanol-withdrawal tremor and on audiogenic-seizure susceptibility in ethanol (Frye et al., 1981). The lack of effects during withdrawal from chronic ethanol on NT-like immunoreactivity (Wachi et al., 1996) and on NT binding sites (Campbell and Erwin, 1993) supports the above negative behavioral data.
On the other hand, 14 days of repeated cocaine injections and its early (24 hours) or late (8 days) withdrawal enhanced NT-like immunoreactivity in the substantia nigra; however, the degree of this increase was dependent on the mode of drug administration: Subcutaneous injection versus continuous minipumps (Cain et al., 1993). Changes after cocaine injections were also reported in rat brains processed for a NT receptor autoradiography, with significant changes of binding in subcortical areas and the PFC (Pilotte et al., 1991). Up till now, there has been no behavioral proof for NT-cocaine (or other drugs of abuse) interactions during drug withdrawal.
Conclusions
The mesolimbic and mesocortical DA pathways are highly implicated in the psychomotor stimulant and reinforcing effects of drugs of abuse. The above-reported studies strongly suggest that NT signaling is one of the specific neurochemical mechanisms for the modulation of the rewarding activity of cocaine and other drugs of abuse. Nevertheless, controversial results exist. At the present, the precise molecular mechanisms by which NT exerts its role in drug addiction remain to be elucidated. Furthermore, it is quite surprisingly the total lack of notion to date about the potential role of the peptide in stress-induced reinstatement of drug-seeking behavior, a very important aspect of relapse that is due to the recognized role of NT in stress regulation.
In contrast to what is the case after treatment with typical antipsychotic drugs, the psychostimulants enhance NT levels, mainly in subpopulations of the striato-nigral GABA pathways (Merchant, 1994) that are regulated primarily by the DA D1 receptors, although increases in NT levels in other brain regions have been also described. An increase in NT levels and the consequent NTS1 receptor stimulation appear to be essential for c-fos expression; and thus, activation of subpopulations of the striato-nigral GABA pathway (Fadel et al., 2006) that may be part of the reward pathways, especially when originating from the NAC. Furthermore, it could be possible that the psychostimulant-induced NT levels in the cell body-dendritic regions of these nerve cells would allow the peptide, through extrasynaptic volume transmission, to diffuse and reach the glutamate and DA terminals. Here, the NT peptides would inhibit DA D2 receptor signaling, via the ‘antagonistic’ NTS1-DA D2 receptor-receptor interaction in the striatal and accumbal glutamate and DA (only dorsal striatal) terminals, leading to increased DA and glutamate release. This increased synaptic glutamate and extra-synaptic DA release may enhance activation of accumbal-VTA GABA pathways that are participating in the reward circuits that are activated by drugs of abuse; however, released NT in the NAC, via NTS1-DA D2 receptor-receptor interactions in the GABA neurons and their glutamate afferents, which project from the NAC to the ventral pallidum, can also activate the anti-reward circuit. In other words, the NT-induced alterations in the balance of activity in the reward and the anti-reward systems will determine how NT mechanisms contribute to counteracting the development of psychostimulant sensitization and possibly, cocaine use disorder.
NTS1 receptors are also localized in the VTA, where their activation may antagonize DA D2 autoreceptor functions and increase glutamate drive, thus enhancing drug abuse by increasing activity in the meso-limbic DA neurons mediating reward. Enhanced NTS1-NMDA receptor interactions in the VTA DA cells also lead to an increased activation of the mesolimbic DA reward neurons.
Based on the current preclinical research, there is no doubt that NT and its heteroreceptor complexes play a significant role in the modulation of substance use disorder. Further research is highly warranted, to understand the mechanisms involved in the interactions between NT, its heteroreceptor complexes including the NTS1-D2 heterocomplex and drugs of abuse, in the brain circuits of reward and anti-reward. Finally, it should be considered that only male animals were used in the previous studies that aimed to investigate the effects of NT in drug addiction. For these animal models to be most relevant to drug abuse in humans, the effects of NT compounds in female rodents will also need to be investigated.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was partly supported by the Swedish Research Council (grant number 04X-715 to KF).