
Abstract
Background:
Deficits in impulse control feature in many psychiatric conditions including bipolar disorder, suicidality and addictions. Lithium lowers impulsivity in clinical populations and decreases pathological gambling in experimental medicine studies, but suffers from adverse effects, poor compliance and a low therapeutic index.
Aims:
Recently we identified that the neuroprotective agent ebselen, which is reportedly safe in humans, inhibited inositol monophosphatase (IMPase), a candidate lithium mechanism. Ebselen also reduced 5-HT receptor (5-HT2A) function which predicts impulsivity lowering properties. Here we investigated the effect of ebselen in rat models of impulsive behaviour.
Methods:
Ebselen was tested in two models of impulsivity with human analogues: the five-choice serial reaction time task (5-CSRTT) and rodent gambling task (rGT). The main outcome measures were premature responses (5-CSRTT and rGT) and choice behaviour (rGT), which model motor impulsivity and choice impulsivity, respectively.
Results:
At doses that decreased 5-HT2A receptor function (DOI-induced wet dog shakes), ebselen decreased premature responding in the 5-CSRTT both in the absence and presence of cocaine. The 5-HT2A receptor antagonist MDL 100,907 also reduced premature responding in the 5-CSRTT although not in the presence of cocaine. In the rGT ebselen showed a tendency to reduce premature responding but had no effect on choice behaviour.
Conclusions:
These findings suggest that ebselen preferentially reduces motor impulsivity over choice impulsivity, and that inhibition of 5-HT2A receptor function is a contributing mechanism. Collectively, these data support the repurposing of ebselen as an anti-impulsive treatment and fast-tracking to clinical trials in patient groups characterised by poor impulse control.
Introduction
Deficits in impulse control are a prominent feature of diverse psychiatric illnesses including bipolar depression, attention deficit hyperactivity disorder, addictions and the diagnostic category of impulse conduct disorders. Impulsivity is also strongly associated with suicidality, self-harm, aggression and pathological gambling, and a significant predictor of relapse and worse clinical outcomes in alcohol and substance dependence (Rogers et al., 2010). Drug therapies for impulse control are few, and those available are poorly tolerated as exemplified by lithium. Lithium is the mainstay therapy for bipolar depression, reduces risk of suicide (Baldessarini et al., 2006), and is reported effective in clinical trials of pathological gambling (Hollander et al., 2005; Rogers and Goodwin, 2005). In accordance with these effects in humans, lithium reduced impulsivity in rodent models (Halcomb et al., 2013; Ohmura et al., 2012). However, lithium can cause a range of disabling side-effects including renal damage (McKnight et al., 2012) and has a low therapeutic index and low compliance as a consequence. These adverse effects limit the high therapeutic potential of lithium as an impulsivity lowering agent, yet the clinical need for such an agent is significant.
A leading candidate mechanism underlying lithium’s therapeutic action is inhibition of inositol monophosphatase (IMPase), resulting in reduced transmitter signalling via the phosphoinositide (PI) pathway (Agam et al., 2009; Berridge et al., 1989; Sade et al., 2016). However, previous attempts to develop IMPase inhibitors have been hampered by poor brain penetrance (Atack, 1997). Recently, we ‘re-profiled’ a library of clinically safe and bioavailable compounds, and identified a potent IMPase inhibitor, ebselen (Singh et al., 2013). Ebselen inhibited IMPase in cultured cells expressing either mouse or human forms of the enzyme, as well as in mouse brain in vivo, thereby suggesting good bioavailability (Singh et al., 2016). Ebselen is an organoselenium compound with antioxidant properties that was originally developed for the treatment of stroke. While it progressed to clinical trials in this and other disorders associated with ischaemic mechanisms (Parnham and Sies, 2013), it has not yet been marketed.
Ebselen has lithium-like effects in neuropharmacological studies in animals (Singh et al., 2013; Antoniadou et al., 2018), some of which are predictive of an impulsivity-lowering agent. In particular, as expected of an IMPase inhibitor, ebselen reduced functions mediated by the PI pathway-coupled 5-HT2A receptor in a mouse model (5-HT2 agonist-induced head-twitch reponse; Singh et al., 2013; Antoniadou et al., 2018) as previously observed for lithium (Goodwin et al., 1986). The 5-HT2A receptor is strongly linked to impulse control (Fink et al., 2015), and 5-HT2A receptor blockade is associated with decreased impulsivity in both animals and humans. For example in the five-choice serial reaction time task (5-CSRTT) in rodents, 5-HT2A receptor antagonists including the selective agent MDL 100,907 decreased premature responding, which is a widely used measure of motor impulsivity (Fletcher et al., 2007; Higgins et al., 2003; Winstanley et al., 2004). MDL 100,907 also decreased premature responding in a rodent gambling task (rGT) although it did not affect choice impulsivity (Adams et al., 2017). Whilst these data await detailed investigation in humans, the non-selective 5-HT2A receptor antagonist quetiapine decreased measures of impulsivity in healthy volunteers and patients with bipolar depression (Rock et al., 2013; Van den Eynde et al., 2008).
Here we tested the hypothesis that ebselen would lower impulsivity in animals using two 5-HT-sensitive rodent models, the 5-CSRTT (Bari et al., 2008; Robbins, 2002) and rGT (Zeeb et al., 2009). MDL 100,907 was used as a reference compound in the 5-CSRTT.
Materials and methods
Animals
For impulsivity testing, rats (male Long Evans; Charles River, Saint-Constant, QC, Canada) were pair-housed (12 h reverse light–dark cycle; lights off at 07:00 hours) and acclimatised to the animal facility over at least 2 weeks during which rats were handled briefly each day. Food was then restricted and progressively decreased to 14 g/rat/day prior to habituation to operant boxes and training. For measurement of 5-HT2A function, rats (male Lister Hooded; Harlan, Bicester, UK) were housed in fours (12 h light–dark cycle; lights on at 07:00 hours) and acclimatised to the animal facility as above. Experiments followed the principles of the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines and were approved by local ethical committees and followed national guidelines for animal welfare (Canadian Council of Animal Care, UK Animals (Scientific Procedures) Act 1986).
Experimental procedures
Five-choice serial reaction time task
The 5-CSRTT was performed as described previously (Bari et al., 2008; Robbins, 2002). Briefly, 14 rats were trained to respond to brief visual cues presented in one of five target areas with an inter-trial interval of 5 s. On the day of drug/vehicle testing, the inter-trial interval was lengthened from 5 s to 7 s to cause a reliable increase in premature responding. Three doses of ebselen (2, 5 and 10 mg/kg) or vehicle were administered i.p. (1 h prior to testing) in a pseudorandom order in a Latin square design. Rats were tested 5 days a week with the treatment day falling on the third day. The lengthened inter-trial interval was used only on the drug day to ensure that rats did not habituate to the lengthened inter-trial interval and to provide pre- and post-drug data. One week following ebselen treatment, three doses of MDL 100,907 (0.01, 0.03 and 0.1 mg/kg i.p.) were tested in the same rats, again using a shift from 5s to 7s inter-trial interval length on the day of drug administration. The Latin square design for the 5-CSRTT (and rGT, see below) was counter-balanced based for performance in the last training day of each task. A baseline session was performed both the day before and the day after drug administration to ensure stable off-drug performance.
To assess the effect of ebselen on stimulant-induced impulsive responding, ebselen (10 mg/kg i.p.) or vehicle was administered 1 h prior to cocaine (15 mg/kg i.p.; Fletcher et al., 2011) that was injected 10 min before testing (following a Latin square design). The inter-trial interval for these sessions was kept at 5 s. The same cohort of 14 rats were used for this, although two rats were excluded from these experiments due to increasing levels of omissions across testing days. This experiment was then repeated in the same 12 rats but with 0.03 mg/kg i.p. MDL 100,907 in place of the ebselen.
Rodent gambling task
The rGT (uncued version) was performed as described previously (Zeeb et al., 2009). In short, a separate cohort of 21 rats was trained to respond to brief visual cues in 5-CSRTT boxes. Once reliable responding was established, different reward contingencies were associated with four nose-poke apertures (the central aperture was not used in this task). For each aperture choice, the reward and punishment size, and probability of outcome, differed between the choices. When no reward was delivered, a fixed-length time-out was delivered.
The length of this time-out varied between choices, with the longest time-out being associated with the largest, least likely reward. The choice P1 resulted in 1 × 45-mg sucrose pellet on 90% of trials and a 5 s time-out 10% of the time. The P2 choice resulted in 2 × 45-mg sucrose pellets 80% of the time and a 10-s time-out on the remaining 20% of occasions. Similarly, P3 yielded either 3 × 45-mg sucrose pellets 50% of the time or a 30-s time-out 50% of the time. Finally, the P4 choice resulted in 4 × 45-mg sucrose pellets 40% of the time, but 60% of these choices triggered a 40-s time-out period. These contingencies were designed such that choice of options paired with smaller per trial gains and lower penalties ultimately resulted in the most reward over time, with P2 representing the most optimal choice. Once stable performance was achieved, rats were administered ebselen (2, 5 and 10 mg/kg i.p.) or vehicle in a Latin square design.
DOI-induced wet dog shakes
The effect of ebselen on 5-HT2A receptor function was tested in rats by measuring ‘wet dog shakes’ induced by the 5-HT2A agonist 2,5-dimethoxy-4-iodoamphetamine (DOI, e.g. Willins and Meltzer, 1997). Ebselen (2, 5 or 10 mg/kg i.p.) or vehicle was given 1 h before DOI (1 mg/kg i.p.) or vehicle, during which time the locomotor activity of the rats was monitored using a beam break system (PAS home cage system, San Diego Instruments, San Diego, CA, USA). Wet dog shakes were counted during 20 min beginning 5 min after injection of DOI. The group sizes were slightly uneven (ebselen 2 mg/kg n = 9, 5 mg/kg n = 15 and 10 mg/kg n = 6 and vehicle n = 14) as they reflect a combination of data from pilot and confirmatory experiments (cohort was included as a factor and found not to alter the results).
Drugs
Ebselen, DOI and cocaine were obtained from Sigma Aldrich, and MDL 100,907 was obtained from Tocris. Ebselen was dissolved to 0.5 M in dimethyl sulfoxide (DMSO) and made up to injection volume (8 mL/kg) using 4% cyclodextrin. Cocaine and DOI were dissolved in 0.9% saline. MDL 100,907 was dissolved in 0.9% saline and the pH adjusted to 6.25 using 0.1 M NaOH and 0.1 M HCl.
Data analysis
The principal measure of motor impulsivity in both the 5-CSRTT and rGT was premature responding and the a priori prediction was that this would be reduced by ebselen. The principal measure of choice impulsivity (cost/benefit decision making) was choice preference in the rGT. Other behavioural variables were accuracy, omissions, latency to make choice responses, latency to collect rewards and total number of trials completed.
For both the 5-CSRTT and rGT change from baseline values are reported. Baseline values have been taken into account in other studies (e.g. Barnes et al., 2018; Grottick and Higgins, 2000). For example, Δ% premature responses was calculated as the percentage of trials in which a premature response was made during the drug test day minus the percentage of trials in which a premature response was made on the previous drug-free day, i.e. Δ% premature responses = % premature on drug day − % premature on baseline day. This increased the power of the analysis as it controlled for a large between-subject variance in baseline performance. Raw data for the premature responses are given in Supplementary Information.
For experiments on the effect of ebselen or MDL 100,907 alone, data were analysed with repeated measures ANOVA with dose as a within-subjects factor. For experiments involving cocaine, treatment with ebselen and cocaine were used as separate repeated measures in a nested design. For experiments investigating the effect of ebselen on DOI-induced wet dog shakes, data were analysed by one-way ANOVA with ebselen dose as the between-subjects factor and then compared with vehicle. Post hoc analysis was carried out using planned comparisons with Student’s paired t-tests, corrected for multiple testing. Data are presented as mean ± SEM values. All statistical analyses were conducted using SPSS software (version 22) with p < 0.05 being considered statistically significant.
Results
Effect of ebselen on premature responding in the 5-CSRTT
Rats had the high and stable level of performance required for inclusion in the 5-CSRTT, aside from the increasing rate in omissions of two rats noted in Methods. Thus, inclusion of treatment order as a factor in the analysis of premature responding data revealed no significant effect (F(3,10) = 0.159, p = 0.922 for ebselen study, F(3,10) = 2.368, p = 0.132 for MDL 100,907 study), thereby ruling out habituation to inter-trial interval manipulation. Similarly, comparison of pre-drug day baseline data revealed no long-term drug effects or alterations in performance (F(3,39) = 1.456, p = 0.242).
In the 5-CSRTT, rats administered ebselen (2, 5 and 10 mg/kg i.p.) demonstrated an overall decrease in premature responding compared with vehicle-injected controls (F(3,39) = 2.870, p = 0.049; Figure 1(a)). Pairwise comparisons found that this effect of ebselen was statistically significant at both 5 mg/kg (p = 0.011) and 10 mg/kg (p = 0.032) but not 2 mg/kg ebselen (p = 0.158). In comparison, ebselen had no significant effect on omissions (F(3,39) = 0.890, p = 0.455; Figure 1) or accuracy (F(3,39) = 0.393, p = 0.759; Figure 1) or any other behavioural variables measured (reward latency F(3,39) = 0.880, p = 0.460; correct latency F(3,39) = 0.329, p = 0.805; trails completed F(3,39) = 1.656, p = 0.192).
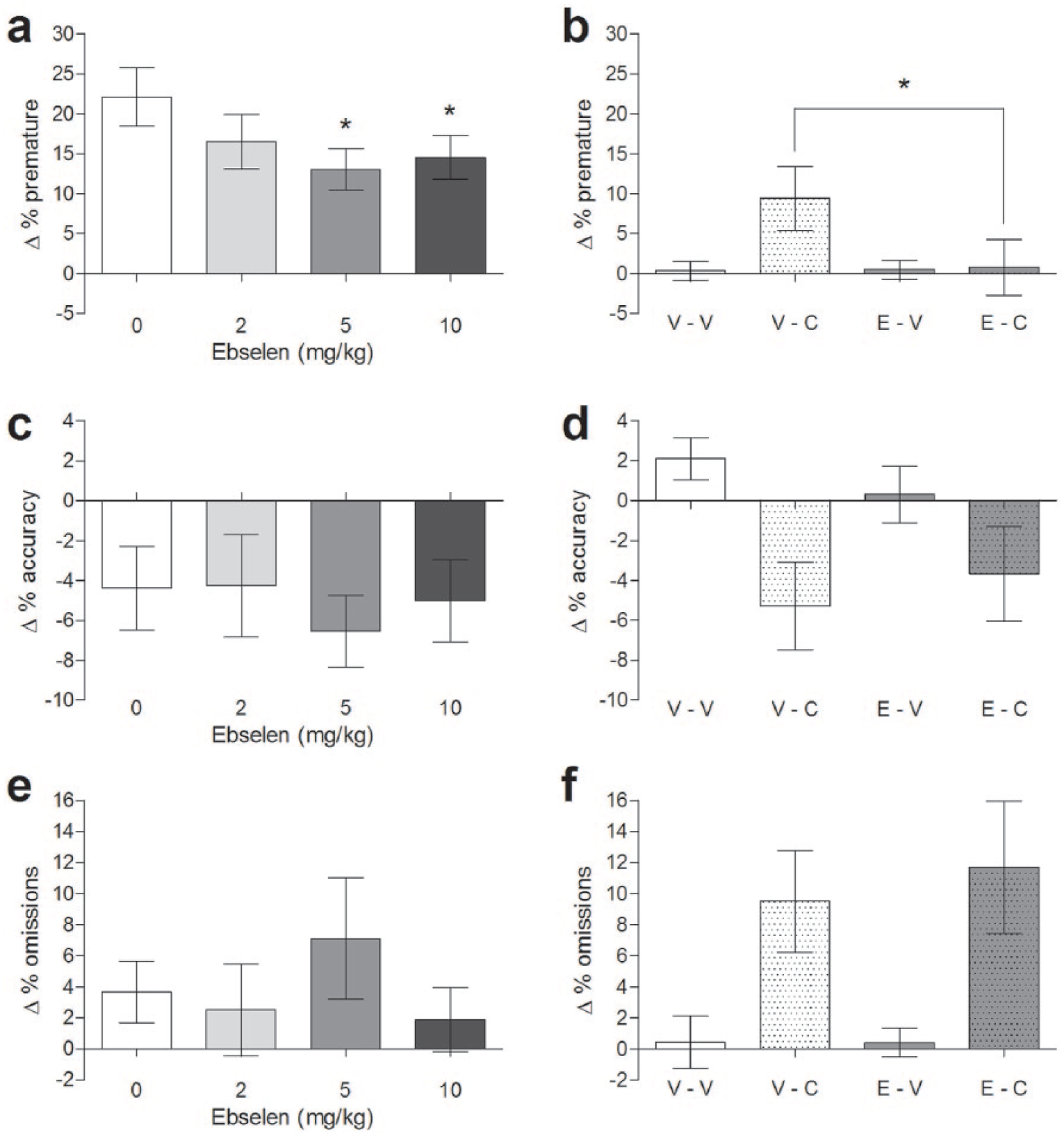
Effect of different doses of ebselen in the five-choice serial reaction time task under conditions of either a long inter-trial interval (left column, n = 14) or a cocaine (15 mg/kg) challenge (right column, n = 12). Parameters reported are premature responding (a, b), accuracy (c, d) and omissions (e, f). Ebselen was administered at the doses indicated (10 mg/kg in the cocaine experiment). V-V: vehicle–vehicle; V-C: vehicle plus cocaine; E-V: ebselen plus vehicle; E-C: ebselen plus cocaine. *p < 0.05 in post hoc paired t-test compared with vehicle. Mean ± SEM values are shown.
Further 5-CSRTT experiments tested the effect of ebselen (10 mg/kg i.p.) when premature responding was elevated by the administration of cocaine (15 mg/kg i.p.). Only the vehicle–cocaine group show a significant increase in premature responses on the test day (vehicle–cocaine t(11) = 2.357, p = 0.038; vehicle–vehicle t(11) = 0.265, p = 0.796; ebselen–vehicle t(11) = 0.417, p = 0.685; ebselen–cocaine t(11) = 0.223, p = 0.827). In these experiments ebselen had an overall effect on premature responses (F(1,11) = 5.442, p = 0.040) that interacted with cocaine (ebselen × cocaine interaction F(1,11) = 5.998, p = 0.032; Figure 1(b)). Pairwise comparisons revealed decreased premature responding in animals administered ebselen plus cocaine compared with cocaine plus vehicle (p = 0.023) such that premature responses made following ebselen plus cocaine did not differ from vehicle–vehicle treatment (p = 0.906).
In comparison, the decrease in accuracy (Figure 1(d)) and increase in omission rates (Figure 1(f)) induced by cocaine were not altered by ebselen (accuracy: effect of ebselen F(1,11) = 0.002, p = 0.969; ebselen × cocaine interaction F(1,11) = 1.302, p = 0.278; omissions: effect of ebselen F(1,11) = 0.787, p = 0.394; ebselen × cocaine interaction F(1,11) = 0.579, p = 0.463).
For other parameters, no changes were detected in number of completed trials (effect of ebselen F(1,11) = 0.682, p = 0.427; effect of cocaine F(1,11) = 1.477, p = 0.250; ebselen × cocaine interaction F(1,11) = 0.569, p = 0.467), correct response latency (effect of ebselen F(1,11) = 2.264, p = 0.160; effect of cocaine F(1,11) = 0.217, p = 0.650; ebselen × cocaine interaction F(1,11) = 0.061, p = 0.810) or reward collection latency (effect of ebselen F(1,11) = 2.782, p = 0.124; effect of cocaine F(1,11) = 0.648, p = 0.438; ebselen × cocaine interaction F(1,11) = 0.364, p = 0.559).
Effect of ebselen on premature responding and choice behaviour in the rGT
In the rGT, rats administered ebselen (2, 5 and 10 mg/kg i.p.) tended to make fewer premature responses although in the overall analysis this was of borderline statistical significance (F(3,60) = 2.286, p = 0.088; Figure 2(a)). However, given our a priori prediction that ebselen would decrease premature responding, further investigation using planned pairwise comparisons revealed that decreases in premature responses by ebselen were statistically significant at 10 mg/kg i.p. (p = 0.028), with non-significant trend effects at 2 mg/kg i.p. (p = 0.052) and 5 mg/kg i.p. (p = 0.077).
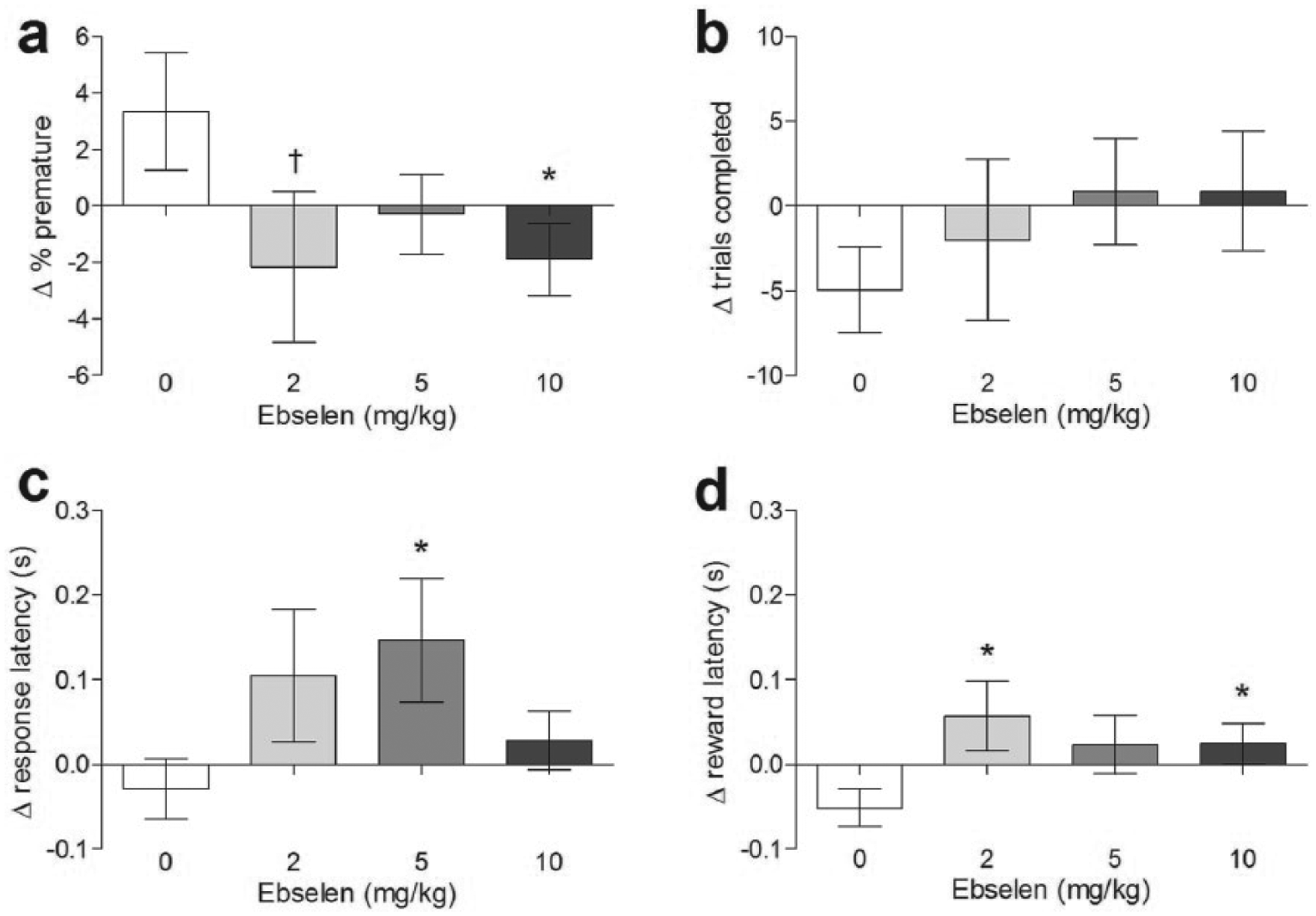
Effect of differing doses of ebselen in the rodent gambling task. Parameters were premature responding (a), trials completed (b), and response (c) and reward (d) latencies. *p < 0.05, †p = 0.052 in post hoc paired t-test compared with vehicle. Mean ± SEM values are shown, n = 21.
In contrast, ebselen had no effect on choice behaviour (dose × choice interaction F(9,180) = 0.712, p = 0.698; Figure 3) at doses that reduced premature responding. Ebselen also had no effect on the number of trials completed (F(3,60) = 0.716, p = 0.546; Figure 2(b)), but tended to increase the latency to collect rewards (F(3,60) = 2.575, p = 0.062), and make a choice (F(3,60) = 2.262, p = 0.090). Further pairwise comparisons showed that ebselen increased the latency to both collect rewards (2 mg/kg p = 0.024; 5 mg/kg p = 0.069; 10 mg/kg p = 0.005; Figure 2(c)) and make choice responses (2 mg/kg p = 0.056; 5 mg/kg p = 0.028; 10 mg/kg p = 0.246; Figure 2(d)).
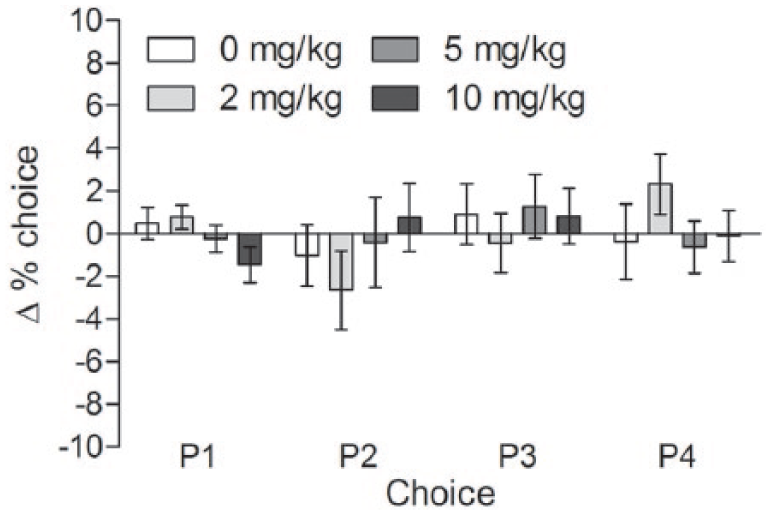
Effect of differing doses of ebselen on choice behaviour in the rGT. The choices available on each trial (P1–P4) are named according to the number of reward pellets delivered on a proportion of the choices made, with P2 being the optimal choice and P4 being the choice of highest risk. See Materials and Methods for further detail. Mean ± SEM values are shown, n = 21 per group.
Effect of ebselen on DOI-induced wet dog shakes
Ebselen decreased the number of DOI-induced wet dog shakes in a dose range similar to that which reduced premature responding (2–10 mg/kg i.p.; F(3,40) = 4.131, p = 0.012; Figure 4(a)), and this effect was significant at 5 mg/kg (p = 0.002) and 10 mg/kg (p = 0.049) but not 2 mg/kg (p = 0.403). To ensure this effect did not represent a generalised decrease in locomotor activity, we analysed the activity of the rats in the hour after ebselen administration but before the rats received DOI (Figure 4(b)). Ebselen had no overall effect on locomotor activity (F(3,40) = 0.206, p = 0.892), even at the highest dose used (p = 0.457). Thus, ebselen decreased premature responding at doses that inhibited 5-HT2A receptor function.
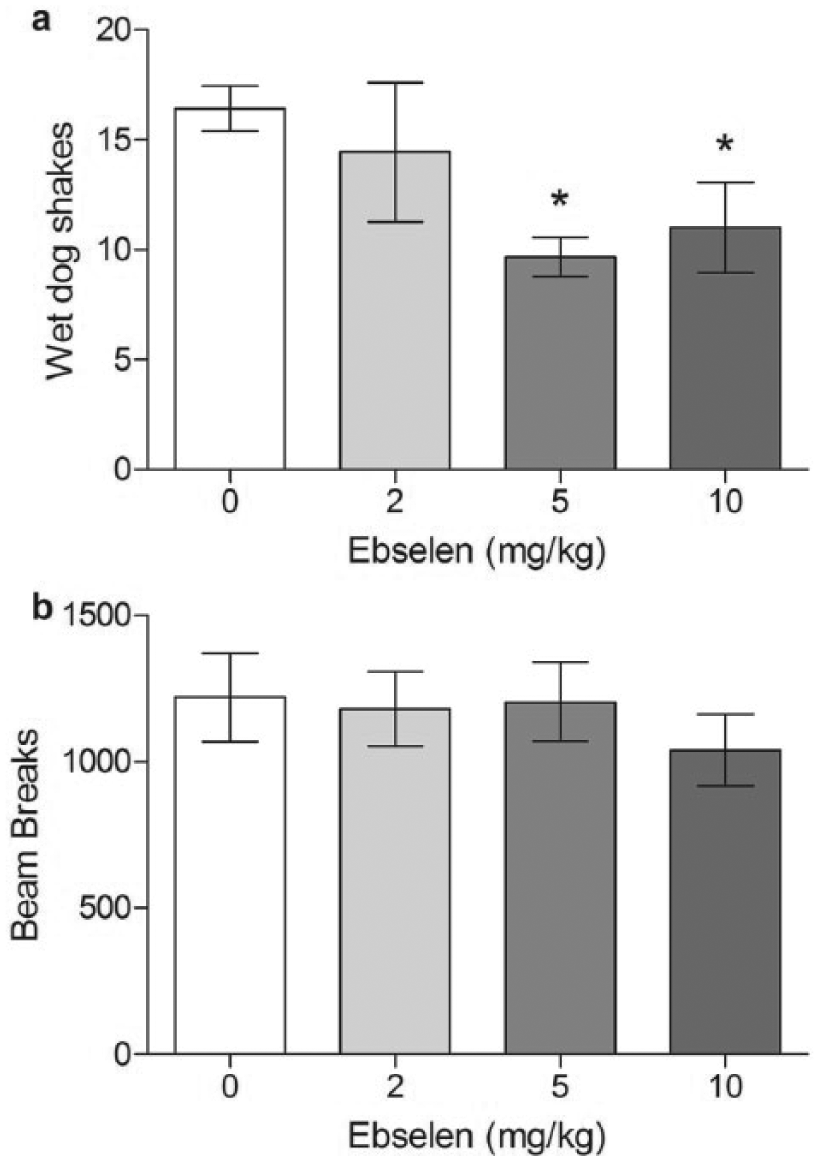
Effect of ebselen on wet dog shakes induced in rats by the 5-HT2A agonist DOI (1 mg/kg) (a) and locomotor activity (beam breaks) over the first 60 min following ebselen injection (b). *p < 0.05 in post hoc unpaired t-test compared with vehicle. Vehicle group n = 14, 2 mg/kg ebselen n = 9, 5 mg/kg group n = 15, 10 mg/kg ebselen group n = 6. Mean ± SEM values are shown.
Effect of MDL 100,907 on premature responding in the 5-CSRTT
As with ebselen, the selective 5-HT2A receptor antagonist MDL 100,907 (0.01, 0.03 and 0.1 mg/kg i.p.) decreased premature responding in the 5-CSRTT when compared with vehicle-injected controls (F(3,39) =5.870, p = 0.002). In pairwise comparisons, this effect was statistically significant at all doses (vehicle versus 0.01 mg/kg p=0.008; vehicle versus 0.03 mg/kg p = 0.009; vehicle versus 0.1 mg/kg p = 0.001; Figure 5(a)). MDL 100,907, unlike ebselen, also increased response accuracy (Figure 5(c); F(3,39) =3.078, p=0.039), and this effect was significantly different from vehicle treatment at doses of 0.01 mg/kg (p = 0.020) and 0.1 mg/kg (p = 0.027), but was only a trend effect at 0.03 mg/kg (p = 0.052). In comparison, MDL 100,907 had no effect on the number of omissions (Figure 5(e); F(3,39) = 0.308, p = 0.819), trials completed (F(3,39) = 0.234, p = 0.872), correct latency (F(3,39) = 1.134, p = 0.347) or reward latency (F(3,39) = 1.766, p = 0.170).
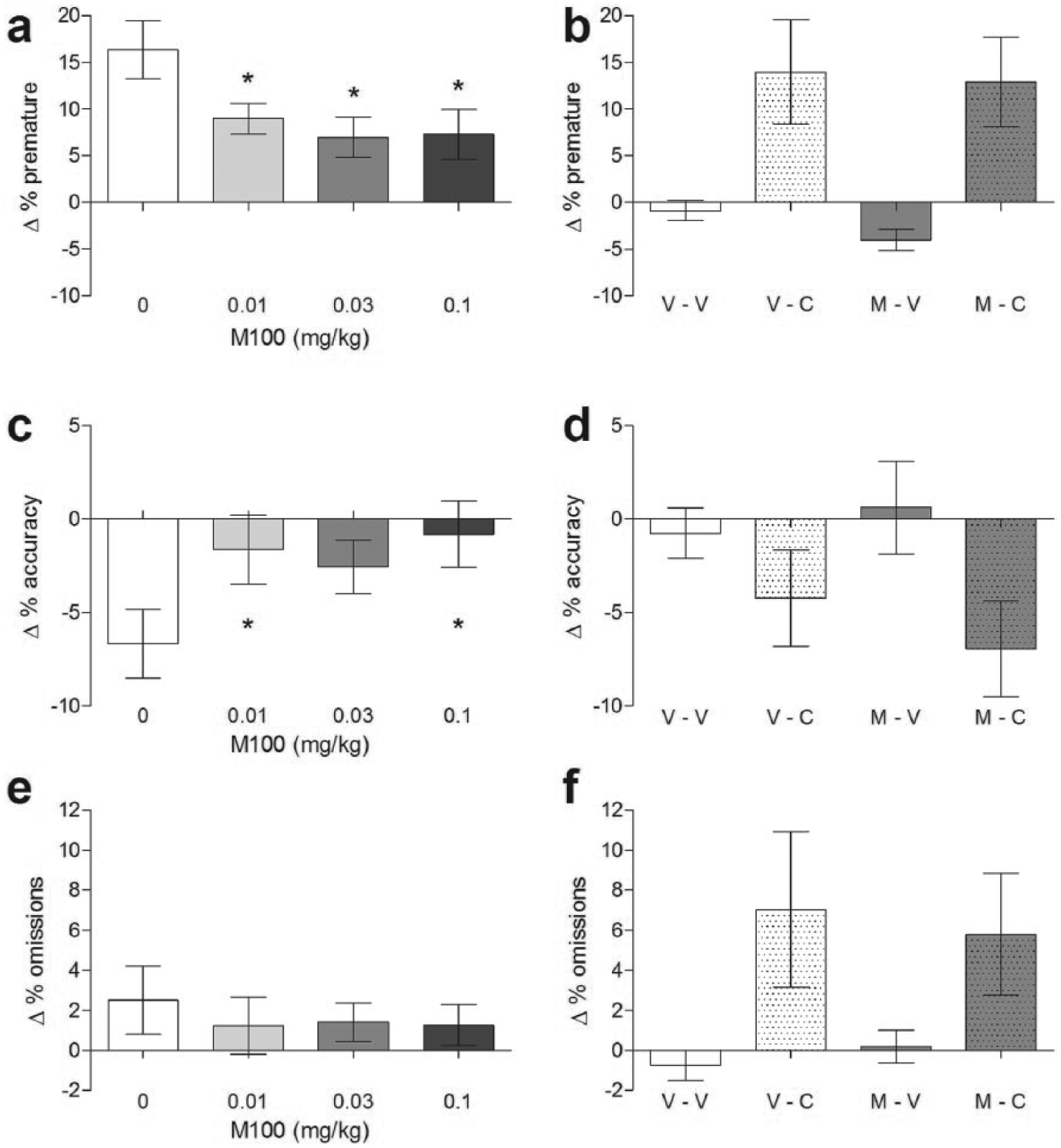
Effect of MDL100,907 in the 5-CSRTT under conditions of either a long inter-trial interval (left column, n = 14) or a cocaine (10 mg/kg) challenge (right column, n = 12). Parameters reported are premature responding (a, b), accuracy (c, d) and omissions (e, f). MDL100,907 was administered at the doses indicated (0.03 mg/kg in the cocaine experiment). V-V: vehicle–vehicle; V-C: vehicle plus cocaine; M-V: MDL100,907 plus vehicle; M-C: MDL100,907 plus cocaine. *p < 0.05 in post hoc paired t-test compared with vehicle. Mean ± SEM values are shown.
In contrast to ebselen, at the dose tested (0.03 mg/kg) MDL 100,907 did not reduce cocaine-induced increases in premature responding in the 5-CSRTT (overall effect of MDL 100,907 F(1,11) = 1.277, p = 0.282; overall effect of cocaine F(1,11) = 10.627, p = 0.008; MDL 100,907 × cocaine interaction F(1,11) = 0.196, p = 0.667; Figure 5(b)).
There was a trend for cocaine to increase the number of omissions (F(1,11) = 4.292, p = 0.063; Figure 5(f)) and this was not altered by MDL 100,907 treatment (overall effect of MDL 100,907 F(1,11) = 0.009, p = 0.925; MDL 100,907 × cocaine interaction F(1,11) = 0.422, p = 0.529). No change was seen in the number of trials completed (effect of MDL 100,907 F(1,11) = 0.137, p = 0.719; effect of cocaine F(1,11) = 3.195, p = 0.101; MDL 100,907 × cocaine interaction F(1,11) = 0.137, p = 0.719) or in accuracy (effect of MDL 100,907 F(1,11) = 0.120, p = 0.735; effect of cocaine F(1,11) = 2.687, p = 0.129; MDL 100,907 × cocaine interaction F(1,11) = 1.373, p = 0.266; Figure 5(d)). Cocaine shortened response latency (F(1,11) = 7.537, p = 0.019) and there was a trend for MDL 100,907 to lengthen it (F(1,11) = 3.968, p = 0.072) but these did not interact (F(1,11) = 0.047, p = 0.832).
Discussion
A decrease in impulsivity is likely to be crucial to lithium’s therapeutic actions. Here we investigated whether the lithium-mimetic ebselen (Singh et al., 2013) could improve impulse control in two rat models, the 5-CSRTT and rGT. Ebselen reduced premature responding, a commonly accepted measure of motor impulsivity in both models although this finding was more robust in the 5-CSRTT than the rGT. This impulsivity lowering effect of ebselen was behaviourally selective in that it was observed at doses that did not alter other variables such as attentional accuracy or omission rates. Ebselen also reduced heightened premature responding in the presence of cocaine, suggesting that the drug retains efficacy even under conditions of high levels of impulsivity. Finally, in the rGT ebselen had no effect on choice behaviour. The latter finding supports the idea that ebselen preferentially reduces motor impulsivity versus choice impulsivity, at least as assessed under the current conditions. Overall, these data demonstrate that ebselen exerts impulsivity lowering effects and support the repurposing of this neuroprotective agent for the management of disorders associated with loss of impulse control.
Our previous studies (Singh et al., 2013; Antoniadou et al., 2018) identified that ebselen had various neuropharmacological actions similar to those reported for lithium in preclinical models, and the present study extends this to impulse control. Thus, in the 5-CSRTT, the inhibition of premature responding by ebselen was observed at more than one dose, and under conditions of elevated premature responding as induced by cocaine. These data accord with previous findings that lithium decreased premature responding in a three-choice version of the 5-CSRTT (Ohmura et al., 2012) as well as the 5-CSRTT itself (Adams and Winstanley, unpublished observations). In the 5-CSRTT, ebselen reduced premature responding at doses that had no significant effect on other parameters such as accuracy or number of trials completed. Moreover, ebselen did not reduce spontaneous locomotor activity in the open field. Together these findings suggest that ebselen did not reduce premature responding through a non-specific impairment of task performance.
We investigated the effect of ebselen in the rGT principally to assess its effects on choice behaviour for which the task is well optimised. While ebselen did not alter this choice behaviour (see below), the drug decreased premature responding in this task although the effect was of borderline statistical significance and thereby less clear-cut than in the 5-CSRTT. It is possible that the effect of ebselen on premature responding in the rGT could be enhanced by increasing the inter-trial interval, as performed in the 5-CSRTT. However, the impact of manipulating inter-trial interval on choice behaviour is little explored and would need to be investigated prior to any further testing of ebselen on premature responding in this model.
In the rGT, ebselen also increased the time taken to collect rewards and make choice responses. One interpretation of this finding is that ebselen decreases premature responding not simply as a result of greater inhibitory control, but also through increasing patience and/or promoting waiting for rewards. It is also plausible that ebselen decreases sensitivity to positive reinforcement. In support of these ideas, ebselen is known to alter 5-HT function (see below) and recently changes in 5-HT have been linked to both waiting for rewards and altered reward sensitivity (Fonseca et al., 2015; Line et al., 2014; Miyazaki et al., 2014). However, ebselen did not alter reward latencies in the 5-CSRTT. On the other hand, the different reward sizes available during the rGT might better probe the reward sensitivity than the 5-CSRTT in which rewards are small and not variable.
The present observation that ebselen inhibited 5-HT2A receptor function in rats, as indicated by the reduction in DOI-induced wet dog shakes, is in accordance with our previous findings on DOI-induced head-twitch responses in mice (Singh et al., 2013; Antoniadou et al., 2018). Several lines of evidence support the idea that this pharmacological effect may contribute to the reduction in impulsivity induced by ebselen. First, the selective 5-HT2A receptor antagonist MDL 100,907, like ebselen, reduced premature responding in the 5-CSRTT as observed here and in earlier studies (Winstanley et al., 2004). Second, 5-HT2A receptor agonist administration increased premature responding, and elevated 5-HT2A receptor expression is associated with impulsive traits in rodents (Fink et al., 2015). Third, like ebselen, lithium reduced 5-HT2A receptor function (Goodwin et al., 1986) and also inhibited premature responding (see above). Finally, the 5-HT2A receptor is coupled to the PI signalling pathway and therefore likely sensitive to IMPase inhibition by ebselen and lithium.
However, while inhibition of 5-HT2A receptor function may contribute to the impulsivity lowering action of ebselen, other mechanisms may also be involved. Thus, the current experiments detected differences in the actions of ebselen and MDL 100,907. For example, in contrast to ebselen, MDL 100,907 did not reduce the increase in premature responding in the 5-CSRTT after cocaine (although note that a previous study found MDL 100,907 to reduce cocaine-induced premature responding, albeit at >10 times higher dose than that used here; Fletcher et al., 2011). Also, ebselen tended to decrease premature responding in the current uncued version of the rGT whereas recently it was found that MDL 100,907 had no significant effect in this model (although it did reduce premature responding in the cued version; Adams et al., 2017).
In addition to impacting signalling via 5-HT2A receptors, ebselen likely decreases the function of 5-HT2C and 5-HT2B receptors as these are also Gq-coupled to the PI pathway. Since both 5-HT2C and 5-HT2B receptors have been implicated in the regulation of impulsivity (Bevilacqua et al., 2010; Winstanley et al., 2004), they may contribute to ebselen’s activity in the 5-CSRTT and rGT. However, blockade and/or loss of function of 5-HT2C and 5-HT2B receptors is associated with increased impulsivity, and specifically increased premature responding in the 5-CSRTT and rGT in the case of 5-HT2C receptor antagonists (Adams et al., 2017; Winstanley et al., 2004). Thus, rather than contributing to the impulsivity lowering effect of ebselen, these actions are likely to work in the opposite direction, although there is a wealth of literature showing that mixed 5-HT2A/B/C receptor antagonists reduce impulsivity, including for example ketanserin (Fletcher et al., 2007; Passetti et al., 2003; Talpos et al., 2006). This suggests that 5-HT2A-mediated alterations to behaviour typically dominate when using mixed-class 5-HT2 receptor ligands and inhibitors.
Inhibition of IMPase also likely reduces the functionality of other Gq-coupled receptors including mGluR5 glutamate receptors and α1-adrenoceptors. The latter receptors have also been linked to the regulation of impulse control, although typically it is increased activity at these receptors that is associated with improved impulse control. Thus, the positive allosteric modulator of mGluR5 receptors ADX47273 reduced premature responding in the 5-CSRTT at doses that were not confounded by evidence of disruptive changes in behaviour (Isherwood et al., 2015). This contrasts with the non-specific effects of mGluR5 negative allosteric modulator MPEP (2-methyl-6[phenylethynyl]-pyridine; Quarta et al., 2007). The α1-adrenoceptor agonist phenylephrine also decreased premature responding in the 5-CSRTT, albeit at doses that induced non-specific effects that likely disrupted 5-CSRTT performance (Pattij et al., 2012). This suggests that any inhibitory action of ebselen on the function of these receptors would likely oppose the impulsivity-lowering effect of ebselen rather than contribute to it.
Overall, whilst reduced 5-HT2A receptor function (through IMPase inhibition) seems a likely important contributor to the impulsivity-lowering effect of ebselen, other mechanisms may well be involved. Such mechanisms may extend beyond inhibition of IMPase and Gq-coupled receptors because ebselen has pharmacological effects on other enzymes including inhibition of glutaminase (Thomas et al., 2013). This enzyme plays a key role in converting glutamine to glutamate, and therefore inhibition has the potential to reduce glutamate function and potentially alter impulsivity. Further experimentation (e.g. incorporating genetic knockdown of IMPase) is necessary to clarify the mechanisms underlying ebselen’s effect on impulsivity.
As noted above, in the rGT ebselen did not affect choice behaviour, that is a preference for the delivery of risky high rewards or reliable low rewards. Although this observation requires further exploration, for example through testing the action of ebselen in additional models of impulsive choice such as reward-based delay discounting strategies, it suggests that ebselen may selectively reduce motor impulsivity over choice impulsivity. Impulsivity has long been regarded as a multifaceted, heterogeneous construct (Dalley and Robbins, 2017; Evenden, 1999), and different forms of impulsivity are thought to involve dissociable neurobiological mechanisms. In this context, whereas motor impulsivity is sensitive to 5-HT manipulations as mentioned above, often this is not the case for choice impulsivity (Cardinal, 2006; Homberg, 2012; Pattij and Vanderschuren, 2008; Winstanley et al., 2006). Deficits in impulsive action are common in conditions associated with impulse control disorders, such as bipolar depression (e.g. Fleck et al., 2001), but drugs may be effective as impulsivity lowering agents in clinical practice regardless of their selectivity for a specific impulsivity domain. While atomoexetine has been found to be effective across multiple forms of impulsivity (Robinson et al, 2008), one study has found that atomoxetine selectively reduced motor impulsivity versus choice whereas methylphenidate preferentially reduced choice impulsivity versus motor (Paterson et al., 2012), but both drugs are clinically useful in the management of ADHD.
Consistent with the IMPase inhibition by ebselen detected in preclinical models, recent data show that ebselen decreased brain inositol at a safe and tolerated dose in healthy human volunteers (Singh et al., 2016). The latter findings are evidence that ebselen engages with IMPase in the human brain at a dose that is seemingly without adverse effects. Moreover, in human subjects, ebselen was recently found to have certain lithium-like neuropsychological effects, including a reduction in delay aversion in the Cambridge Gambling Task, which is consistent with improved impulse control (Masaki et al., 2016). The latter findings combined with the current evidence of ebselen having an impulsivity-lowering action in animal models support the fast-tracking of this agent into clinical trials for the management of disorders characterised by poor impulse control such as bipolar disorder, self-harm, aggression, pathological gambling and compulsive drug use.
Supplemental Material
JOP784876_Supplementary_Material – Supplemental material for The putative lithium-mimetic ebselen reduces impulsivity in rodent models
Supplemental material, JOP784876_Supplementary_Material for The putative lithium-mimetic ebselen reduces impulsivity in rodent models by Chris Barkus, Jacqueline-Marie N Ferland, Wendy K Adams, Grant C Churchill, Philip J Cowen, David M Bannerman, Robert D Rogers, Catharine A Winstanley and Trevor Sharp in Journal of Psychopharmacology
Footnotes
Declaration of conflicting interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Some of the authors are listed as co-inventors on patents for the use of ebselen in the control of impulsivity disorders (Barkus, Churchill, Cowen, Bannerman, Rogers, Winstanley and Sharp) and bipolar depression (Churchill and Sharp). Patent rights are assigned to the University of Oxford but the authors will share a percentage of any royalties on the basis of revenue-sharing arrangements between the University and other collaborating institutions.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by a research grant from the Medical Research Council (DPFS grant, MR/L013150/1). CAW also received salary support through the Canadian Institutes for Health Research (CIHR) New Investigator Award program. WA was supported by philanthropic funding provided by Paul and Diane Erickson and the Gillespie family.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.