
Abstract
Background:
Research interest has grown around the potential therapeutic use of cannabidiol in mood-related disorders, due to its anxiolytic and antidepressant-like effects. These have been partially attributed to its action as an allosteric modulator of 5-HTR1A. However, the exact mechanism supporting cannabidiol properties remains unclear.
Aims:
To assess the effects of cannabidiol on different targets of the hypothalamus-pituitary-adrenal axis under baseline and stress conditions.
Methods:
We administered cannabidiol (5 mg/kg, 15 mg/kg or 30 mg/kg, intraperitoneally) or vehicle to male C57BL/6J mice 90 min before single restraint stress exposure (20 min). Using real-time polymerase chain reaction analysis, we measured alterations in the relative gene expression of corticotropin-releasing factor in the paraventricular nucleus, pro-opiomelanocortin in the arcuate nucleus of the hypothalamus, glucocorticoid receptor in the hippocampus, and serotonin 5-HTR1A receptor in the hippocampus and amygdala.
Results:
Under baseline conditions, cannabidiol did not modify any element of the hypothalamus-pituitary-adrenal axis. In contrast, all doses induced alterations in 5-HTR1A in the amygdala and hippocampus. Interestingly, cannabidiol at low (5 mg/kg) and intermediate doses (15 mg/kg) successfully blocked the effects induced by acute stress on corticotropin-releasing factor, pro-opiomelanocortin and glucocorticoid receptor gene expression. Also, restraint stress induced the opposite effects in 5-HTR1A gene expression in the hippocampus and amygdala, an effect not seen in mice treated with cannabidiol at low doses.
Conclusions:
Taken together, these data suggest the ability of cannabidiol to regulate acute stress hypothalamus-pituitary-adrenal axis activation might be explained, at least in part, by its action on 5-HTR1A receptors.
Keywords
Introduction
Cannabidiol (CBD), one of the main compounds present in the plant Cannabis sativa, has recently been proposed as a potential therapeutic drug for the treatment of mood-related disorders (Iseger and Bossong, 2015; Marinho et al., 2015; Shoval et al., 2016). This hypothesis is based on the anxiolytic and antidepressant-like effects observed in the elevated plus maze test (Guimaraes et al., 1990), the forced swimming test (de Mello Schier et al., 2014) and the Vogel conflict test (Moreira et al., 2006), as well as the capacity of CBD to attenuate acute autonomic response to stress in rodents (Resstel et al., 2009). CBD has been reported to act through more than 65 molecular targets (Ibeas Bih et al., 2015). Nevertheless, the exact neurobiological mechanisms underlying its therapeutic properties remain underexplored. Serotonin receptor 1A (5-HTR1A) has long been identified as an important mediator of the stress response due to its distribution throughout the brain, especially in structures traditionally related to stress and anxiety such as the hippocampus (HIPP) and the amygdala (AMY) (Chalmers and Watson, 1991). Growing evidence suggests it appears to play a pivotal role in the anxiolytic- and antidepressant-like effects of CBD (Blessing et al., 2015; Campos et al., 2012; de Mello Schier et al., 2014; Linge et al., 2016).
It is well established that stress exposure triggers a wide range of adaptive physiological and behavioral responses to maintain homeostasis. In this sense, the activation of the hypothalamus-pituitary-adrenal (HPA) axis is a common response to all types of physiological and perceived threats (Rabasa et al., 2015). This activation is meant to be acute or at least of limited duration (Tsigos and Chrousos, 2002); otherwise it leads to a loss of the regulatory negative feedback mechanism, inducing a prolonged increase in the glucocorticoid levels that significantly alters brain neuroplasticity and neurogenesis (Hall et al., 2015). This imbalance would result in a reduction of neuroplasticity, which promotes the development of various mental illnesses, including major depressive disorder, bipolar disorder and anxiety disorder (Merkulov et al., 2017).
To further explore the mechanisms underlying the anxiolytic properties of CBD, in the present study we examine the effects of CBD on the stress HPA axis activation. To this aim, we administered CBD (5 mg/kg, 15 mg/kg and 30 mg/kg, intraperitoneally (i.p.)) to C57BL/6J mice under both physiological conditions and before exposure to 20 min of restraint stress. CBD doses were selected based on published data reporting anxiolytic-like effects (Casarotto et al., 2010; Long et al., 2010). Specific changes in the gene expression of corticotropin-releasing factor (CRF) in the paraventricular nucleus (PVN), glucocorticoid receptor (NR3C1) in the HIPP, proopiomelanocortin (POMC) in the arcuate nucleus (ARC) and 5-HTR1A in the AMY and HIPP were measured by real-time polymerase chain reaction (RT-PCR) under baseline and stress conditions.
Material and methods
Mice
We used 72 6-week-old male C57BL/6J mice, weighing 20–25 g (Charles River, Lille, France): 32 for experiment 1 and 40 for experiment 2. Mice were housed in groups of six per cage (40 cm × 25 cm × 22 cm) under controlled conditions (temperature, 23°C ± 2°C; relative humidity, 60% ± 10%; 12 h light/dark cycle, lights on from 8:00 h to 20:00 h). All the studies were conducted in compliance with the Spanish Royal Decree 1201/2005, Spanish Law 32/2007, and European Union Directive of 22 September 2010 (2010/63/UE) regulating the care of experimental animals, and the ARRIVE guidelines (Kilkenny et al., 2010).
Drugs
CBD (STI Pharmaceuticals, Essex, United Kingdom) was dissolved in ethanol:cremophor:saline (1:1:18) and injected i.p. at the three experimental doses (5 mg/kg, 15 mg/kg and 30 mg/kg in 0.3 mL of solution). The vehicle was a solution of ethanol: cremophor:saline (1:1:18) injected i.p. (0.3 mL). According to (Deiana et al., 2012) cremophor seems to be the most suitable solvent when it comes to achieving high brain concentrations of CBD after intraperitoneal administration in mice. This formulation provides higher brain concentrations than other solvents, suggesting its suitability for obtaining observable effects (Navarrete et al., 2018; Viudez-Martinez et al., 2018a, 2018b).
Behavioral analyses
Experiment 1: to evaluate the effects of CBD on different targets of HPA under baseline conditions
C57BL/6J mice were treated with vehicle, CBD 5 mg/kg (CBD5), CBD 15 mg/kg (CBD15) and CBD 30 mg/kg (CBD30) (n=8 per group). Mice were sacrificed by cervical dislocation 2.5 h following the intervention, and gene expression studies were performed by RT-PCR.
Experiment 2: to evaluate the effects of CBD on different targets of HPA after acute restraint stress exposure
C57BL/6J mice were distributed into five different groups: non-stress exposed (NSE) + vehicle (VEH), stress-exposed + vehicle (SE+VEH), stress-exposed + CBD 5mg/kg (SE+CBD5), stress-exposed + CBD 15mg/kg (SE+CBD15) and stress-exposed + CBD 30mg/kg (SE+CBD30) (n=8 per group). The corresponding treatment was administered 1.5 h before acute restraint stress exposure. This time lapse from treatment administration to experimental exposure was determined according to published data (Deiana et al., 2012).
A modified protocol for restraint stress was employed (Garcia-Gutierrez and Manzanares, 2011). Unanesthetized mice were confined in an acrylic cylindrical tube (inner size 10 cm × 3 cm × 3 cm) for 20 min. After this period of restraint stress, animals were removed from the tubes and housed in their cages. Control animals remained in their home cages until the end of the experiment. Changes in the level of gene expression were studied after killing mice by cervical dislocation and removing brains 2.5 h after the end of the 20 min restraint period, as described elsewhere (Navarrete et al., 2017).
Gene expression studies by RT-PCR
After cervical dislocation, brains were harvested and instantly frozen at −80°C. Brain sections containing the regions of interest were cut (500 μm) according to Paxinos and Franklin (Franklin and Paxinos, 1997), in a cryostat (−10°C), mounted onto slides and immediately stored at −80°C. Sections were microdissected following the method described by Palkovits (1983). Total RNA was obtained from brain micropunches with TRI Reagent extraction reagent (Applied Biosystems, Madrid, Spain) and reverse transcription was carried out. Quantitative analysis of the relative abundance of CRF (Mm01293920_s1) in PVN, NR3C1 (Mm00433832_m1) in the HIPP, POMC (Mm00435874_m1) in the ARC and 5-HTR1A (Mm00434106_s1) in the HIPP and AMY was measured by RT-PCR, performed on the StepOne Sequence Detector System (Applied Biosystems, Madrid, Spain). Life Technologies supplied all reagents and the manufacturer’s protocols were followed. The reference gene used was 18S rRNA (Mm03928990_g1), according to Boda et al. (2009). Data for each target gene were normalized to the endogenous reference gene, and the fold change in target gene expression was determined using the 2-ΔΔCt method (Livak and Schmittgen, 2001).
Blinding, randomization and group size determination
All the experiments were analyzed by observers blinded to treatment. Mice were randomly distributed into the different groups from the beginning of each experiment. Group sizes were determined after performing different power calculations, which showed that 8–12 subjects were needed for each group to obtain a power of more than 80%. To minimize the number of animals employed, we decided to use eight subjects per group.
Statistical analyses
Statistical analyses of gene expression studies were performed using one-way and two-way analysis of variance (ANOVA), followed by the Student-Newman-Keul’s test to compare treatment versus control groups. Statistical analyses were performed with SigmaPlot v11.0 (Systat Software Inc., Chicago, IL, USA) software. Differences were considered significant if the probability of error was less than 5%.
Results
Experiment 1. Dose-response effects of CBD on HPA-axis under baseline conditions: gene expression studies in NSE mice
The administration of CBD (5 mg/kg, 15 mg/kg or 30 mg/kg) did not modify the relative gene expression of CRF in the PVN compared to vehicle-treated mice (Figure 1a) (one-way ANOVA followed by Student–Newman–Keul’s: F(3, 29)= 0.119, p= 0.984). Furthermore, CBD treatment (5 mg/kg, 15 mg/kg or 30 mg/kg) did not produce any significant changes in NR3C1 in the HIPP (Figure 1b) (F(3,29)=1.032, p=0.395) nor in POMC in the ARC under baseline conditions (Figure 1c) (F(3,29)=0.719, p=0.549).
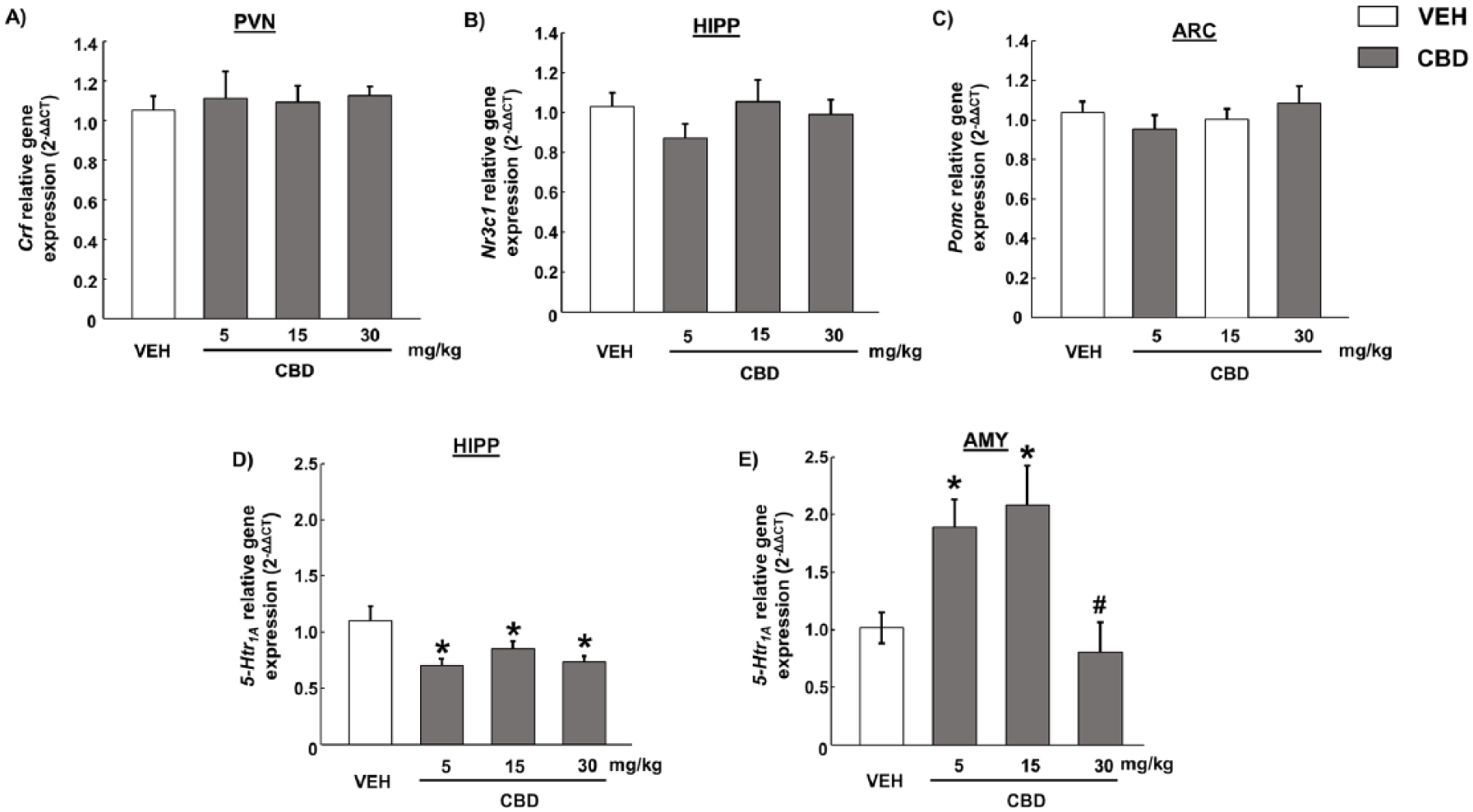
Real-time polymerase chain reaction studies of hypothalamus-pituitary-adrenal axis targets in C57BL/6 J mice treated with cannabidiol (CBD) (5, 15 or 30 mg/kg; intraperitoneally (i.p.)) under physiological conditions. 2-ΔΔCT relative gene expression of (a) corticotropin-releasing factor in paraventricular nucleus, (b) glucocorticoid receptor in the hippocampus (HIPP), (d) proopiomelanocortin in arcuate nucleus, (d) 5-HTR1A in HIPP and (e) 5-HTR1A in amygdala. Columns represent the means and vertical lines ± the standard error of the mean (SEM).
However, CBD significantly reduced 5-HTR1A gene expression in the HIPP at all doses evaluated (Figure 1d) (F(3,29)=4.86, p<0.01) and increased 5-HTR1A gene expression in the AMY at 5 mg/kg and 15 mg/kg (Figure 1e) (F(3,29)=6.129, p<0.01). The 30 mg/kg dose failed to induce any alterations on 5-HTR1A gene expression in the AMY (Figure 1e).
Experiment 2. Dose-response effects of CBD on HPA-axis under stress conditions: gene expression studies in mice exposed to acute restraint stress
Stress exposure significantly increased CRF relative gene expression in PVN (two-way ANOVA followed by Student-Newman-Keul’s test: F(1,39)=17.767, p<0.001) (Figure 2a). The administration of CBD 5 mg/kg and 15 mg/kg (but not CBD 30 mg/kg) blocked this stress-induced augmentation (F(3,39)=17.097, p<0.001) (Figure 2a).
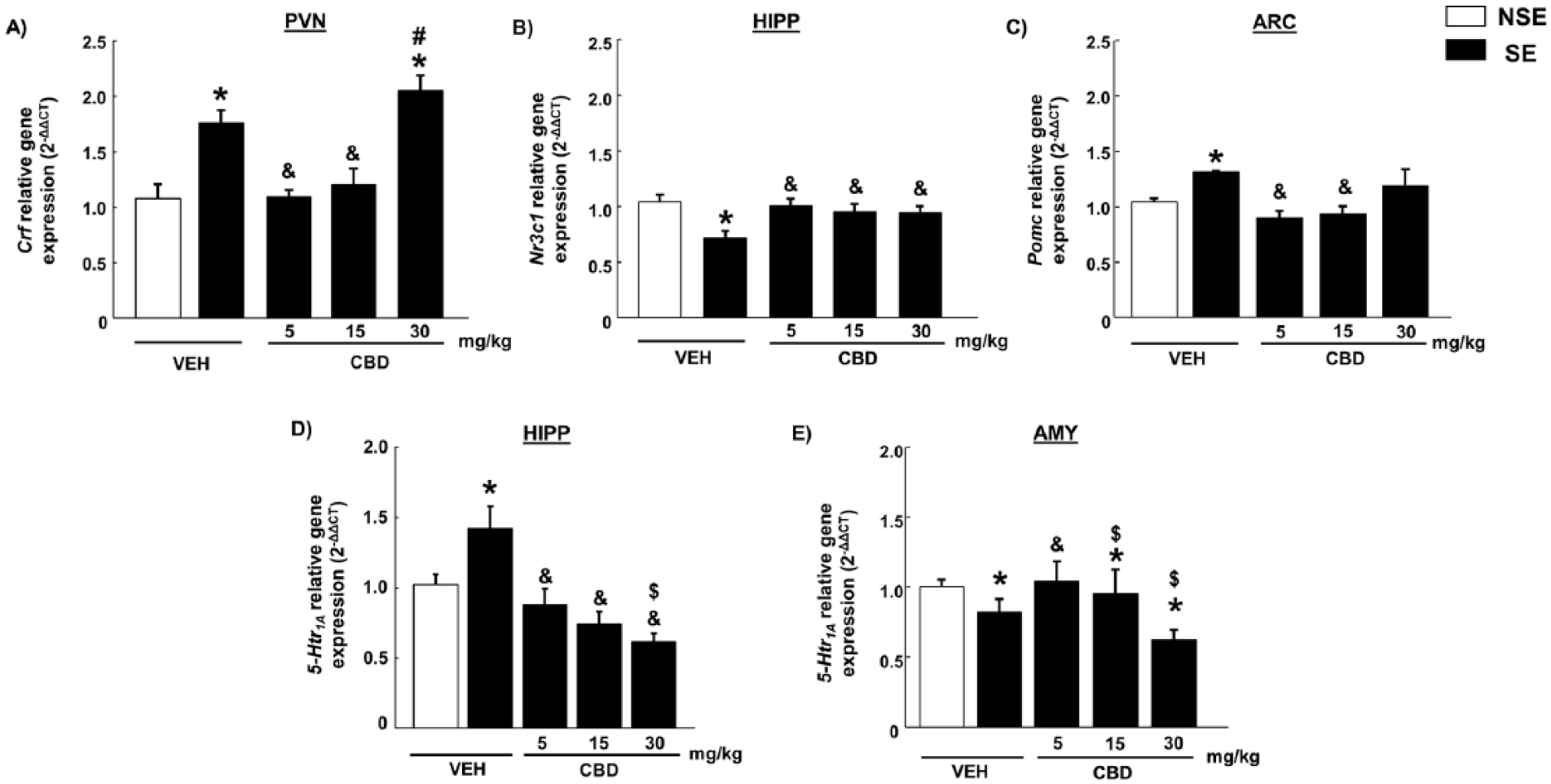
Real-time polymerase chain reaction studies of hypothalamus-pituitary-adrenal targets in C57BL/6 J mice treated with cannabidiol (CBD) (5, 15 or 30 mg/kg; intraperitoneally (i.p.)) and exposed to a model of acute restraint stress. The 2-ΔΔCT relative gene expression of (a) corticotropin-releasing factor in paraventricular nucleus, (b) glucocorticoid receptor in hippocampus (HIPP), (c) proopiomelanocortin in arcuate nucleus, (d) 5-HTR1A in HIPP and (e) 5-HTR1A in amygdala. Columns represent the means and vertical lines ± the standard error of the mean (SEM).
Concurrently, acute restraint incremented POMC gene expression in ARC (F(1,39)=6.47, p<0.05). This increase was not seen in mice treated with CBD 5 mg/kg or 15 mg/kg, yet the highest dose failed to produce any effect (F(3,39)=7.130, p<0.001) (Figure 2c).
Conversely, we observed reduced expression of NR3C1 in the HIPP of mice exposed to restraint stress (F(1,39)=20.155, p<0.001), which was not seen in mice treated with CBD at any dose (F(3,39)=7.294, p<0.001) (Figure 2b).
Furthermore, we also analyzed changes in 5-HTR1A gene expression in the HIPP and AMY. After stress exposure, an increase of 5-HTR1A relative gene expression was observed in the HIPP (F(1,39)=8.589, p<0.05), absent in mice pretreated with CBD 5 mg/kg or 15 mg/kg (F(3,39)=14.383, p<0.001) (Figure 2d). On the contrary, acute restraint stress produced a reduction of 5-HTR1A in the AMY (F(1,39)=5.186, p<0.05) that was only blocked by CBD 5 mg/kg (F(1,39)=5.177, p<0.05) (Figure 2e).
Discussion
The results of the present study suggest CBD may play a relevant role in regulating the HPA axis response to acute stress. This statement is based on the following findings: 1) the administration of CBD (5 mg/kg and 15 mg/kg) blocked the alterations of CRF, POMC, and NR3C1 gene expressions induced by acute restraint stress in the PVN, ARC, and HIPP, respectively; 2) the administration of CBD modified the effects of acute restraint stress on 5-HTR1A gene expression in the HIPP (5 mg/kg, 15 mg/kg, and 30 mg/kg) and AMY (5 mg/kg); and 3) under baseline conditions, CBD did not modify the gene expression of any of the key elements of the HPA axis measured (CRF, POMC, or NR3C1).
Activation of the HPA axis is a critical physiological mechanism for survival after exposure to stressful stimuli. It has long been accepted that this response causes a rise in the CRF secretion to the portal system, which results in a cortisol secretory burst in the general circulation, an augmentation of β-endorphin produced by arcuate POMC neurons (Tsigos and Chrousos, 2002), and a reduction in the expression of glucocorticoid receptors in the hippocampus by negative feedback (Mifsud et al., 2017). The data reported in this study corroborate available evidence, because an increase in the relative gene expression of CRF and POMC in PVN and ARC nuclei, respectively, and a reduction of NR3C1 in HIPP were observed in C57BL/6J mice exposed to acute restraint stress. Interestingly, the administration of CBD blocked all the alterations subsequent to this exposure at low and intermediate doses tested (5 mg/kg and 15 mg/kg). However, the 30 mg/kg dose failed to block the increment of CRF and POMC induced by acute stress. The differences between the dose-response effects of CBD on HPA axis response are in agreement with previous data reporting that the anxiolytic-like effects induced by CBD presented a bell-shaped dose-response curve, which was observable at moderate but not higher doses (Guimaraes et al., 1990). Nevertheless, under baseline conditions, the administration of CBD (5 mg/kg, 15 mg/kg, or 30 mg/kg) did not alter any of these classical targets involved in the HPA axis response to stress. Taken together, these results support the hypothesis that CBD modulates the expression of HPA axis-related genes only under stress conditions.
Considering that 5-HTR1A has long been proposed as the principal serotonin receptor involved in the regulation of stress physiology (Chalmers and Watson, 1991) and one of the main targets for CBD (Blessing et al., 2015; Campos et al., 2012; Ibeas Bih et al., 2015), we also measured 5-HTR1A relative gene expression. Previous studies have proposed that CBD acts as either a 5-HTR1A receptor agonist (Russo et al., 2005) or allosteric modulator (Blessing et al., 2015). The results derived from the present study, in which CBD modified the gene expression of 5-HTR1A in the HIPP and AMY, without exerting any effects on CRF, POMC or NR3C1, suggest that CBD may act as an allosteric modulator. However, further studies, in which hormone levels of corticosterone or ACTH or protein levels of CRF are measured after the administration of CBD, are needed to make a more concise statement about CBD action mechanism.
Additionally, acute stress induced the opposite effects in 5-HTR1A gene expression, increasing it in the HIPP and reducing it in the AMY. These results are in agreement with previous reports (Campos et al., 2012) (for review see Albert, 2012). The differential effects of stress exposure in these brain areas may be related to their divergent raphe nuclei innervation (AMY innervated by dorsal raphe and HIPP innervated by medial raphe) (Kirby et al., 1997). Interestingly, the administration of CBD at low doses blocked the alterations induced by stress on 5-HTR1A, thus supporting the hypothesis that 5-HTR1A plays an important role in the potential utility of CBD for stress-related disorders. Nevertheless, further studies employing selective 5-HTR1A antagonists, such as WAY100635, are needed to determine if the actions of CBD on the HPA axis response to stress involve the participation of this receptor.
In summary, this study reveals the ability of CBD to modulate different HPA axis-related genes, which can contribute to strengthening, at least in part, the potential efficacy of CBD in the treatment of stress-related disorders such as anxiety. The results reported here show for the first time how CBD is able to modulate 5-HTR1A gene expression in two areas closely related with stress response (HIPP and AMY) without altering natural HPA axis elements under baseline conditions. Moreover, the present study also reveals how the administration of low doses of CBD (5 mg/kg) block the alterations induced by stress on HPA-related genes (CRF, POMC, NR3C1, and 5-HTR1A). Nevertheless, further exploration of the CBD action mechanism remains necessary to introduce this drug as a new pharmacological tool for the treatment of stress-related disorders.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by ‘Instituto de Salud Carlos III” (Retics, RD16/0017/0014), ‘Plan Nacional sobre Drogas’ (PNSD 2016/016) and ‘Ministerio de Economía y Competitividad’ (FIS, PI14/00438).