
Abstract
Background:
Alcohol exposure during development has detrimental effects, including a wide range of physical, cognitive and neurobehavioural anomalies known as foetal alcohol spectrum disorders. However, alcohol consumption among pregnant woman is an ongoing latent health problem.
Aim:
In the present study, the effects of trichostatin A (TSA) on emotional and cognitive impairments caused by prenatal and lactational alcohol exposure were assessed. TSA is an inhibitor of class I and II histone deacetylases enzymes (HDAC), and for that, HDAC4 activity was determined. We also evaluated mechanisms underlying the behavioural effects observed, including the expression of brain-derived neurotrophic factor (BDNF) in discrete brain regions and newly differentiated neurons in the dentate gyrus (DG).
Methods:
C57BL/6 female pregnant mice were used, with limited access to a 20% v/v alcohol solution as a procedure to model binge alcohol drinking during gestation and lactation. Male offspring were treated with TSA during the postnatal days (PD28–35) and behaviourally evaluated (PD36–55).
Results:
Early alcohol exposure mice presented increased anxiogenic-like responses and memory deterioration – effects that were partially reversed with TSA. Early alcohol exposure produces a decrease in BDNF levels in the hippocampus (HPC) and prefrontal cortex, a reduction of neurogenesis in the DG and increased activity levels of the HDAC4 in the HPC.
Conclusions:
Such findings support the participation of HDAC enzymes in cognitive and emotional alterations induced by binge alcohol consumption during gestation and lactation and would indicate potential benefits of HDAC inhibitors for some aspects of foetal alcohol spectrum disorders.
Introduction
In Western countries, alcohol is the most commonly consumed drug, frequently used by adolescents and young adults, even during pregnancy, due to a lack of risk perception (European Monitoring Centre for Drugs and Drug Addiction, 2018). Its consumption among pregnant women has increased in the past years (Tsang and Elliott, 2017) and may lead to birth defects, including a wide range of long-lasting physical and neurobehavioural anomalies known as foetal alcohol spectrum disorders (FASD; Crews and Nixon, 2008; Flak et al., 2014; Roozen et al., 2016). Different studies based on physical examinations estimate that the full range of FASDs in the USA and some Western European countries might number as high as 1–5/100 school children (i.e. 1–5% of the population; May et al., 2009, 2014, 2018).
Preclinical studies have consistently established that prenatal alcohol exposure (PAE) caused adverse effects on the development of different brain structures (Eberhart and Parnell, 2016; Guerri et al., 2009; Marquardt and Brigman, 2016; Rojas-Mayorquín et al., 2016). Additionally, PAE has an impact in the development of the central nervous system, disrupting cell interactions, modifying the expression of genes and affecting neurogenesis, myelination and innate immune responses in animal models (Cantacorps et al., 2017; Pascual et al., 2018; Reynolds et al., 2011; Riley et al., 2011; Schneider et al., 2011). The effects on the brain produced by PAE are associated with neuropsychological and cognitive deficits in attention, spatial and visual processing, executive functioning, language and memory (Allan et al., 2014; Comeau et al., 2015; Marquardt et al., 2014; Marquardt and Brigman, 2016; Mattson et al., 2011) and altered responsiveness to stressors (reviewed in Hellemans et al., 2010). Furthermore, in rodents, alcohol exposure during early postnatal development (lactational period) also induces persistent neurobiological alterations, including anxiety-related behaviour and changes in locomotion and motivation (Cantacorps et al., 2017, 2018). Indeed, children whose mothers consumed alcohol reported a variety of cognitive defects, such as impaired social interaction, altered attention and hyperactivity (Clarke and Gibbard, 2003). Together, such data suggest a multifaceted residual ‘footprint’ of neurodevelopmental alcohol exposure that could be relevant for the identification of mechanisms underlying the lifelong persistence of FASD (Guerri et al., 2009; Kleiber et al., 2012).
Recent studies have elucidated that alcohol is capable of acting through epigenetic-mediated mechanisms (Agudelo et al., 2016; Mandal et al., 2017). In the nervous system, one of the best characterised histone modifications is the acetylation of N-terminal lysine residues that shifts the conformation of chromatin into a relaxed state, leading to gene transcription (Renthal et al., 2009). Histone deacetylases enzymes (HDACs) may influence the activities of histone acetyltransferases (HATs) in chromatin remodelling and thereby play an essential role in gene transcription to regulate cell proliferation, migration, apoptosis, angiogenesis and so on (Chater-Diehl et al., 2017; Li and Seto, 2016; Wang et al. 2012). Targeting HDACs using HDAC inhibitors results in neuroprotection and anti-inflammatory and neuro-regenerative effects (Harrison and Dexter 2013; Xu et al. 2011). Therefore, such enzymes have been proposed as potential therapeutic targets for the cognitive deterioration associated with aging and neurodegenerative diseases, and seem to improve the symptoms of various mental diseases, such as post-traumatic stress disorder and depression (Deussing and Jakovcevski, 2017; Matsumoto et al., 2013).
In animal models, several HDAC inhibitors improved test performance related to cognitive abilities (fear conditioning, novel object recognition (NOR), food preference and spatial memory tasks; Bredy et al., 2008; McQuown et al., 2011). Trichostatin A (TSA) is a substance that inhibits both class I and class II HDAC enzymes (Cantley et al., 2017), shifting the balance towards the active transcription of neuronal genes and the amelioration of plasticity and cognitive deficits (Abel and Zukin, 2008). Studies have shown that TSA treatment enhanced contextual and cued fear memory in addition to memory reconsolidation in rodents (Gräff et al., 2013a; Zhong et al., 2015). Furthermore, it reduced alcohol-induced oxidative stress (Agudelo et al., 2011) and lowered alcohol-induced anxiety and alcohol intake by inhibiting HDAC activity (Legastelois et al., 2017). Other studies have also shown acetylation changes associated with the transcription factors involved in learning and memory functions, such as the cyclic adenosine monophosphate response element binding protein (CREB; Barrett and Wood, 2008; Koshibu et al., 2009). Extracellular signal-regulated kinase pathway and CREB activation stimulate the expression of brain-derived neurotrophic factor (BDNF; Barco et al., 2002; Bramham and Messaoudi, 2005). BDNF is one of the most important neurotrophins, activating a range of second messenger pathways that support cell survival, differentiation, synaptic strength and plasticity and dendritic outgrowth (Jia et al., 2008; Kowiański et al., 2018; Minichiello, 2009; Park and Poo, 2013). Alcohol exposure alters CREB function (Dong et al., 2014; Zhang et al., 2018) and BDNF expression (Logrip et al., 2015; Pandey et al., 2015; Tapocik et al., 2014). Moreover, withdrawal from chronic alcohol induced a reduction of BDNF in the central amygdala of rodents, which could be associated with a decrease in MAP kinases, together with a reduction of CREB phosphorylation in this brain structure (Kyzar and Pandey, 2015; Moonat et al., 2010).
In our study, we aimed to assess the effects of a pharmacological treatment involving inhibitors of the HDAC enzymes (TSA) on anxiety-related behaviour alterations, locomotor activity changes and memory impairments caused by prenatal and lactation alcohol exposure (PLAE). Furthermore, we evaluated the mechanisms underlying the behavioural effects observed by analysing molecular factors related to plasticity (pCREB/CREB and BDNF/tropomyosin receptor kinase B (TrkB) expression) in the hippocampus (HPC) and the prefrontal cortex (PFC) – brain areas involved in the processing of the assessed cognitive function – and newly differentiated neuron progenitors in the dentate gyrus (DG). HDAC4 activity was also determined due to its implication in cognition.
Materials and methods
Animals
Male and female C57BL/6 inbred mice aged 10–12 weeks old (Charles River, Barcelona, Spain) were delivered to our animal facility (UBIOMEX, PRBB) to be used as breeders. Mice were housed in couples within standard cages in controlled laboratory conditions at a stable temperature of 21±1°C and 55±10% humidity. The mice were allowed to acclimatise to the new environmental conditions for at least one week prior to experimentation. All tests took place during the first few hours of the dark phase of a reversed light/dark cycle (lights off at 08:00 h and on at 20:00 h). Following successful mating, pregnant females (N=30) were observed daily for parturition. For each litter, the date of birth was designated as postnatal day (PD) 0. Pups remained with their mothers for 21 days and were then weaned (PD21). After weaning, a total of 80 male offspring were housed in groups of four. Food and water were available ad libitum, except when water was substituted for alcohol in accordance with the drinking in the dark (DID) test and during behavioural testing of the offspring. All procedures were conducted in compliance with the guidelines of the European Communities Directive 86/609/EEC regulating animal research and were approved by the local ethics committee (CEEA-PRBB).
Drugs
Ethyl alcohol was purchased from Merck Chemicals (Darmstadt, Germany) and diluted in tap water to obtain a 20% (v/v) alcohol solution.
Animals received either vehicle dimethyl sulfoxide (DMSO; Sigma–Aldrich, Steinheim, Germany) or TSA (Sigma–Aldrich). The stock solution of TSA was diluted with 20% DMSO and saline (0.9% w/v) at the time of administration to reach the required volume (Sharma et al., 2015). TSA (1 mg/kg, 0.5 mg/kg) or vehicle (DMSO in saline, 20% dilution) was injected intraperitoneally once daily during PD28–35. The dose of TSA was selected on the basis of previous studies in rodents (Fleiss et al., 2012).
DID test
This procedure was conducted as previously reported, with minor modifications (Cantacorps et al., 2017; Rhodes et al., 2005), commencing two days after mating. Pregnant females were randomly assigned to two groups: alcohol exposed and water exposed (control). Briefly, food was removed, and the water bottles were replaced with 10 mL graduated cylinders fitted with sipper tubes containing either 20% (v/v) alcohol in tap water or only tap water three hours after the lighting was deactivated. Following a two-hour access period, individual intake was recorded, and food and water bottles were returned to the home cage. During this time, mice were individually housed, and each corresponding male breeding pair was removed from the home cage for the DID procedure. The procedure was repeated on days 2–4, and fresh fluids were provided each day (from Tuesday to Thursday). On day 5 (Friday), alcohol or water cylinders were left for four hours, and fluid intake was recorded. Two empty control cages (water and alcohol) were placed in the rack to measure general liquid loss (leakage/evaporation), and drip values were subtracted from the drinking values. Fluid intake (g/kg of body weight) was calculated from average two-day body-weight values, as dams were weighed at two-day intervals (Mondays and Wednesdays). The procedure was maintained throughout the three-week gestation and three-week lactation periods (gestation day (GD) 1 to PD21). Food and water consumption during ad libitum periods was measured in both pregnant and nursing female mice. In addition, Cantacorps et al. (2017) measured blood alcohol concentrations (BACs) in dams after the last gestational binge-like session (day 12 of the test), and after the last binge drinking session during lactation (day 24 of the procedure), reaching the intoxication levels as defined by the NIAAA (2016) of approximately 80 mg/dL. In the present study, we followed the same procedure, and dams achieved similar levels of alcohol consumption (mL) and levels of alcohol intake (g/kg) as those obtained by Cantacorps et al. (2017), reaching even higher alcohol consumption levels during several days of the procedure. Therefore, we assume that the blood levels of female mice reached levels of intoxication.
Blood alcohol determinations in offspring
Tail blood samples were obtained from the male offspring during lactational period on two occasions (PD15 and PD21) and immediately after the four-hour period of maternal alcohol consumption. BACs were measured using an EnzyChrom™ Ethanol Assay Kit (Bioassay Systems, Hayward, CA).
Experimental design
A schematic description of the procedure of each experiment is provided in Table 1. Animals were randomly separated into six groups after DID test exposure: group 1, control+vehicle; group 2, control+TSA 0.5 mg/kg; group 3, control+TSA 1 mg/kg; group 4, PLAE+vehicle; group 5, PLAE+TSA 0.5 mg/kg; and group 6 PLAE+TSA 1 mg/kg. Mice were used for behavioural assays from PD36 to PD58, and 24 hours after the final test, brains were dissected for biochemical analyses.
Experimental design.
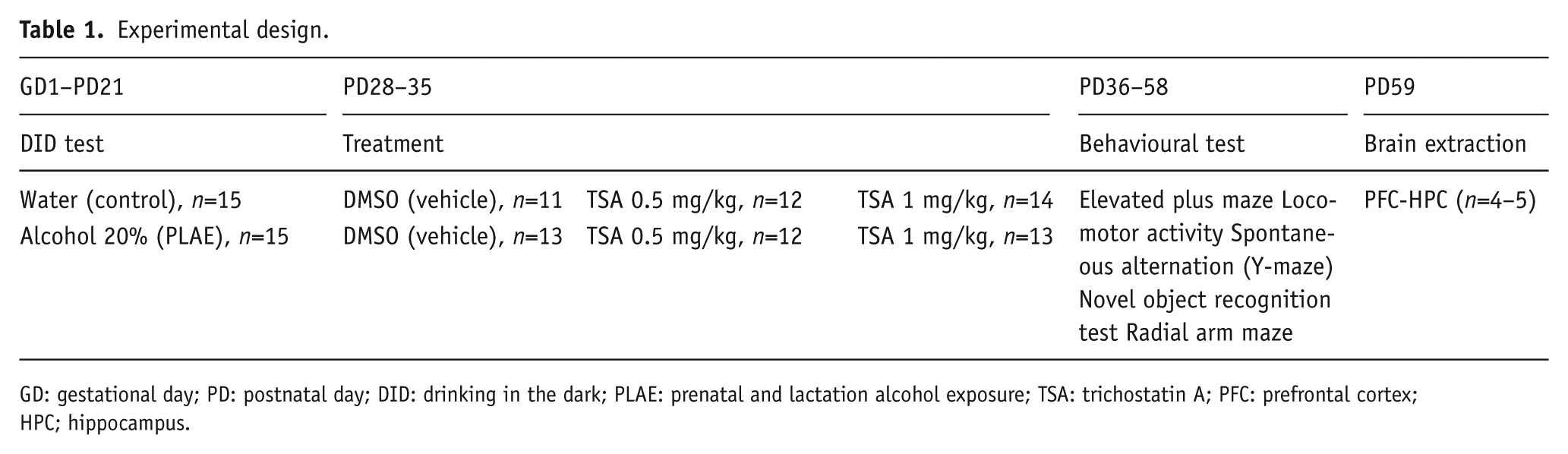
GD: gestational day; PD: postnatal day; DID: drinking in the dark; PLAE: prenatal and lactation alcohol exposure; TSA: trichostatin A; PFC: prefrontal cortex; HPC; hippocampus.
Elevated plus maze
On PD36, each mouse was exposed to the elevated plus maze (EPM; Cantacorps et al., 2018; Pellow and File, 1986; Rodgers et al., 1997). The maze consisted of two open arms (30 cm×5 cm×0.25 cm) and two closed arms (30 cm×5 cm×15 cm), and the junction of the four arms formed a central platform (5 cm×5 cm). The floor and the walls of the closed arms were made of Plexiglas. The open arms had a small edge (0.25 cm) to provide additional grip for the animals. The entire apparatus was elevated 45 cm above floor level. In order to facilitate adaptation, mice were transported to the dimly illuminated laboratory one hour prior to testing. Each mouse was placed in the centre of the maze for five minutes. The percentage of time spent in the open arms and the total number of arm entries were recorded using Smart Software (Panlab s.l.u., Barcelona, Spain).
Locomotor activity
Basal locomotor activity was evaluated in the offspring mice (PD37) in locomotor activity boxes (24 cm×24 cm×24 cm; LE8811 IR; Panlab s.l.u.). Vertical (rearing) and horizontal (deambulation) movements were automatically recorded for 20 minutes, as previously described (Gracia-Rubio et al., 2016).
Spontaneous alternation Y-maze test
Adolescent mice (PD38) were assessed for spatial working memory, as previously reported (Arai et al., 2001; Cantacorps et al., 2017). Briefly, mice were placed in the centre of a Y-shaped maze with two equal arms, each 395 mm long and separated by 120° angles, and were allowed to explore freely for eight minutes. Arm choices were manually recorded. Three consecutive choices of three different arms were counted as an alternation. The score was obtained as the ratio of the total number of alternations by the total number of choices minus 2.
NOR test
Adolescent mice (PD39–42) performed this test in a black open box (24 cm×24 cm×15 cm) using small, non-toxic objects: two plastic boxes and a plastic toy. The task procedure has been described previously (Maccarrone et al., 2002; Pascual et al., 2011) and consists of three phases: habituation, training session (T1) and test session (T2). During the habituation session, mice spent five minutes exploring the open-field arena where the T1 and T2 were performed. In the training session, the mouse was placed in the open-field arena containing two identical sample objects placed in the middle of the testing box for three minutes. After a retention interval of one minute, the animal was returned to the open-field arena with two objects during the test session (three minutes): one object was identical to the sample and the other was novel. The test session was repeated 72 hours after exposure to the familiar object. Object exploration was defined as the orientation of the animal’s snout towards the object within a range of ⩽2 cm from the object. The recognition index was calculated as: DI=(tnovel–tfamiliar)/(tnovel+tfamiliar)×100%, where t is the time each mouse spent exploring an object (recorded using Smart Software; Panlab s.l.u).
Radial arm maze
The radial arm maze (RAM; Panlab s.l.u.) was conducted as previously described, with minor modifications (Moscoso-Castro et al., 2017; Ros-Simó et al., 2013). Briefly, the maze, which was placed on a tripod 100 cm high, consisted of a central hub with eight arms radiating from it. The maze was situated in the middle of a dark room and surrounded by black curtains hanging from the ceiling to the base of the tripod. Three different extra-maze visual cues were fitted to the curtains in order to aid spatial localisation (a pink square, a green rectangle and a white cross). Animals batched for this experiment were food deprived to 80–85% of their baseline body weight with limited access to chow pellets (2–3 g/animal/day) during all the maze procedures (Harvey et al., 2017; Ros-Simó et al., 2013). Previously to RAM sessions, there was a four-day habituation period to the maze (PD43–46). After habituation, RAM training sessions were conducted for 12 consecutive days (PD47–58). During training sessions, food wells at the end of each arm were used to place chocolate pellets (Choco Krispis; Kellogg’s). Animals (PD43–58) were placed in the centre of the maze and were trained to find the food pellets situated in the three randomly selected arms. Each acquisition session consisted of two trials each day, each lasting five minutes or until the mouse found all the pellets. Working memory errors (i.e. entries into baited arms already visited during the same trial) and spatial reference memory errors (i.e. entries into non-baited arms) were recorded. The average value from both daily sessions was calculated for each measure.
Preparation of tissue extract
The mice (n=4–6 in each group) were sacrificed by cervical dislocation 24 hours after the last behavioural test (PD59). PFC and HPC brain areas were dissected following the procedure described by Heffner et al. (1980), and then stored in dry ice at ‒80ºC on an ice-cold plate. The tissue extracts were used to perform Western blot analyses and enzyme-linked immunosorbent assays (ELISA).
Sample preparation and Western blot analyses
First, the PFC and HPC tissues were homogenised in cold lysis buffer (0.05 M Tris-HCl, pH 7.4, 0.15 M NaCl, Triton X-100 1%, Glycerol 10% and EDTA 0.001 M). Homogenates were kept on ice for 30 minutes and centrifuged at 12,000 x g for 15 minutes, and the supernatant was collected to determine the protein concentration. The lysate protein concentration was detected using DC™ Protein Assay (Bio-Rad, Barcelona, Spain). Membranes were incubated overnight at 4ºC with primary antibodies (see Table 2): anti-β-tubulin (anti-mouse, 1:5000; BD Biosciences, Madrid, Spain), anti-phospo CREB (anti-rabbit, 1:1000; Merck Millipore, Madrid, Spain), anti-CREB (anti-rabbit, 1:500, Merck Millipore) and anti-TrkB (anti-rabbit, 1:500; EMD Milipore Corp., Burlington, MA). After washing with TBS/T, blots were incubated with their respective horseradish peroxidase–conjugated antibodies: anti-mouse (1:2500; Rockland, Pottstown, PA) and anti-rabbit (1:2500; Abcam, Cambridge, UK). Blots were developed using the ECL system (ECL Plus; Amersham Biosciences, Little Chalfont, UK). For comparative purposes, control values were normalised to 100%, and the respective protein expression values were adjusted according to the normalisation factor.
Primary antibodies used for the immunohistochemistry and Western blot studies.
ELISA
Sampling and protein extraction
The tissue was homo-genised in cold lysis buffer (137 mM NaCl, 20 mM Tris-HCl, 1% Nonidet P-40, 0.1 g/mL glycerol 10%, 1 mM PMSF, 10 µg/mL leupeptimin, 0.5 mM sodium vanadate, 1 µg/mL pepstatin and dH2O, pH 8.0). Tissue lysates were incubated at 4ºC for 20 minutes. The sample was centrifuged at 9,600 x g for 10 minutes, and then the supernatant was collected. The lysate protein concentration was detected using DC™ Protein Assay (Bio-Rad).
BDNF
BDNF protein levels in brain tissue samples (PFC and HPC) were measured using a commercially available ELISA kit (Promega, Madrid, Spain) according to the manufacturer’s instructions. The ELISA kit used has a wide linear range (7.8–500 pg/mL) and a detection limit, determined in our laboratory, of 15 pg/mL. Standards (eight concentrations) and the assayed brain samples (100 μL aliquots) were run in duplicate. Absorbance was determined at 450 nm using a spectrophotometer.
HDAC4
The total amount of HDAC4 protein activity in the PFC and HPC was measured in offspring mice. Nuclear fractions were isolated, as previously described (Ishida et al., 2002). A colorimetric ELISA assay kit (Epigentek, Farmingdale, NY) was used in accordance with the manufacturer’s instructions. Absorbance was determined using a spectrophotometer at 450 nm. For comparative purposes, control values were normalised to 100%, and the respective activity level values were adjusted according to the normalisation factor.
Immunohistochemistry
The mice (n=4 per group) were anaesthetised with sodium pentobarbital and perfused transcardially through the left ventricle with 100 mL of phosphate-buffered saline (PBS; 0.1 M, pH 7.4) followed by 100 mL of 4% paraformaldehyde-PBS. Brains were extracted, post fixed in the same solution for four hours at room temperature and cryoprotected by immersion in 30% sucrose-PBS at 4ºC. After freezing in dry ice, coronal tissue sections 30 μm thick were obtained using a microtome and stored in 30% sucrose-PBS at ‒20ºC in a six-well dish. An immunohistochemical study of the HPC was performed to measure the doublecortin content (DCX; a marker for newly differentiated and migrating neuroblasts; Brown et al., 2003) in the DG of offspring mice (Luján et al., 2018). Cerebral sections were washed in PBS (0.1 M) and blocked with normal goat serum (NGS; 3%, Triton X-100 0.1% and PBS; Vector Laboratories, Peterborough, UK). The slices were incubated with primary antibody (see Table 2): anti-DCX (anti-rabbit, 1:2000; Abcam) overnight at 4ºC. The sections were then washed with PBS and incubated for two hours with secondary antibody: goat anti-rabbit (1:200; Alexa Fluor®). After labelling, the sections were washed with PBS, and the tissue was dehydrated and mounted onto slides with FluorSave (Calbiochem, San Diego, CA) mounting medium. Images were acquired using a Leica TCS SP5 laser confocal microscope (Leica Microsystems, Wetzlar, Germany). Quantification was carried out by counting the number of DCX-positive cells using ImageJ software (National Institutes of Health, Bethesda, MD).
Statistical analysis
Two-way analysis of variance (ANOVA) with repeated measures was calculated to analyse alcohol intake during the DID test, with group (water–alcohol) as between-subject factors and day as a within-subject factor. One-way ANOVA with repeated measures was used to analyse alcohol intake in the group of dams drinking alcohol during the DID test. Data obtained from behavioural tests and biochemical analyses were evaluated using two-way ANOVA with two factors: group (control–PLAE) and treatment (DMSO (vehicle), TSA 0.5 mg/kg, TSA 1 mg/kg). For the RAM test, three-way ANOVA was used to analyse the data with the variables group, treatment and day (as a within-subject factor). Subsequent post hoc comparisons were calculated using the Bonferroni test. Results are expressed as the mean±standard error of the mean, and statistical significance was set p<0.05.
Results
Maternal alcohol consumption during prenatal and lactation periods
As shown in Figure 1, dams were exposed to alcohol under the DID test procedure from gestational period GD1 to PD21 in order to mimic voluntary binge alcohol drinking. Two-way ANOVA of water and alcohol consumption (mL) during DID testing showed a significant effect of day (F(23, 368)=17.661; p<0.001), group (F(1, 16)=10.334; p<0.005) and an interaction between factors (F(23, 368)=4.378; p=0.05). Bonferroni post hoc comparisons revealed a significant increase in fluid consumption from days 13 to 24 of the test, corresponding to the lactation period (p<0.05). In this period, mice consumed more water than alcohol on day 16 (p<0.05) and days 21–24 of the test (p<0.05; Figure 1(a)).
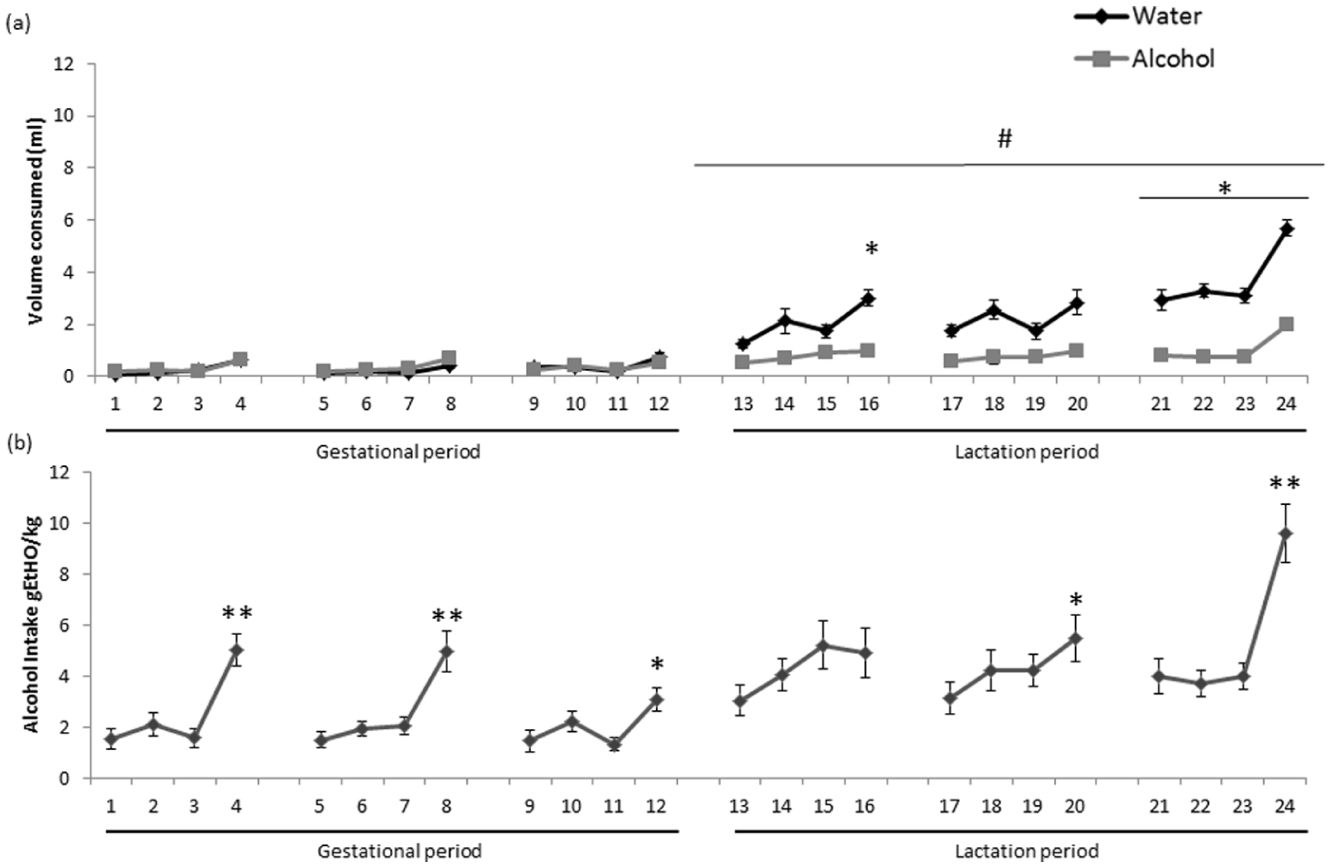
Maternal drinking in the dark (DID) test. Data are expressed as the mean±standard error of the mean (SEM). Alcohol was available two hours per day on days 1–3 and four hours per day on day 4 of the series. (a) Volume of water and alcohol consumed (mL) during the DID test throughout prenatal and lactation periods.
One-way ANOVA with repeated-measures analysis of alcohol intake (g alcohol/kg; Figure 1(b)) showed a significant effect of day (F(23, 414)=8.536; p<0.001). Post hoc comparisons revealed significant differences between the four-hour drinking sessions compared to the two-hour session on days 4, 8 and 24 (p<0.01), and on days 12 and 20 (p<0.05).
Blood alcohol determination in the lactation period
We analysed the BAC in the nursing pups following the EnzyChrom™ Ethanol Assay Kit (Bioassay Systems) calculation. BAC levels for pups receiving alcohol were 14.86±0.82 mg/dL and 19.38±1.59 mg/dL on PD20 and PD24, respectively.
Alcohol-exposed offspring mice present an anxiogenic-like response
Offspring mice were assessed for their anxiety-like behaviour on the EPM (Figure 2). Two-way ANOVA revealed an interaction between the variables group×treatment for the time spent in the open arms (F(2, 72)=4.915; p<0.05) and the percentage of time spent in the open arms (F(2, 72)=5.438; p<0.01). Animals exposed to alcohol during prenatal/lactation periods and treated with vehicle spent less time and a reduced time percentage in the open arms compared to the control group (p<0.01; Figure 2). No significant changes in control groups receiving water were observed. No changes in the number of total entries were found after alcohol exposure or TSA treatment (data not shown).
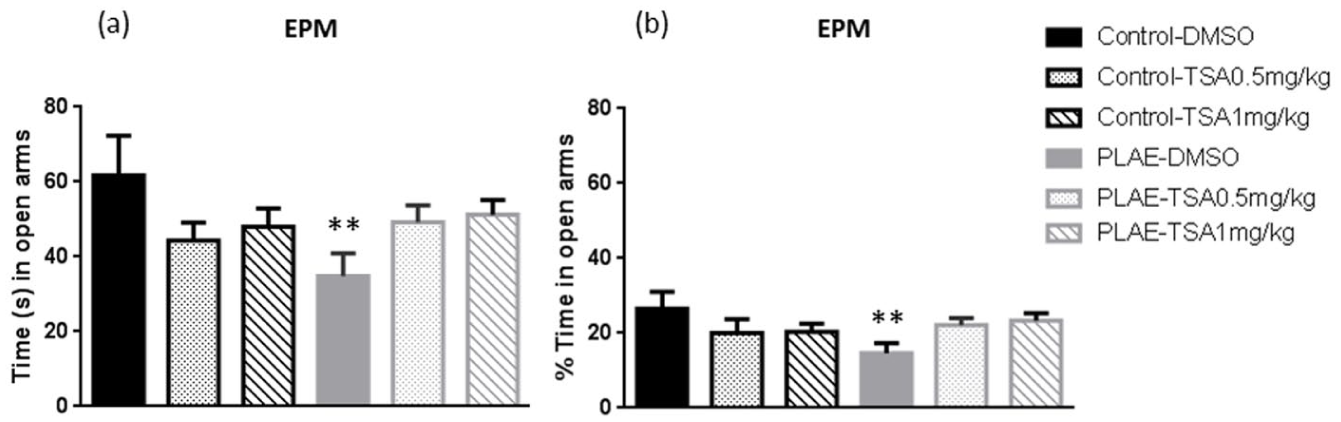
Effects of prenatal and postnatal alcohol exposure in the elevated plus maze. Data are expressed as the mean±SEM. The time spent in open arms (a) and percentage of time spent in open arms (b) was assessed in the offspring.
Maternal alcohol drinking does not affect spontaneous locomotor activity in offspring
The effects of prenatal and lactational alcohol exposure on spontaneous locomotor activity were assessed in the offspring. Two-way ANOVA of horizontal and vertical movements showed no statistical effect (n.s.; data not shown).
Prenatal and postnatal alcohol exposure induces impairments in working memory in adulthood
To assess the prolonged effects of alcohol exposure on spatial working memory, the Y-maze spontaneous alternation test was performed in offspring mice (Figure 3). Two-way ANOVA revealed an effect of group (F(1, 78)=21.853, p<0.001), treatment (F(2, 78)=5.545, p<0.01) and an interaction between factors (F(2, 78)=8.614, p<0.05). PLAE animals performed worse in the Y-maze compared to the control group (p<0.001). In addition, TSA at both doses reversed the deleterious alcohol effects in PLAE mice receiving vehicle (p<0.05; Figure 3).
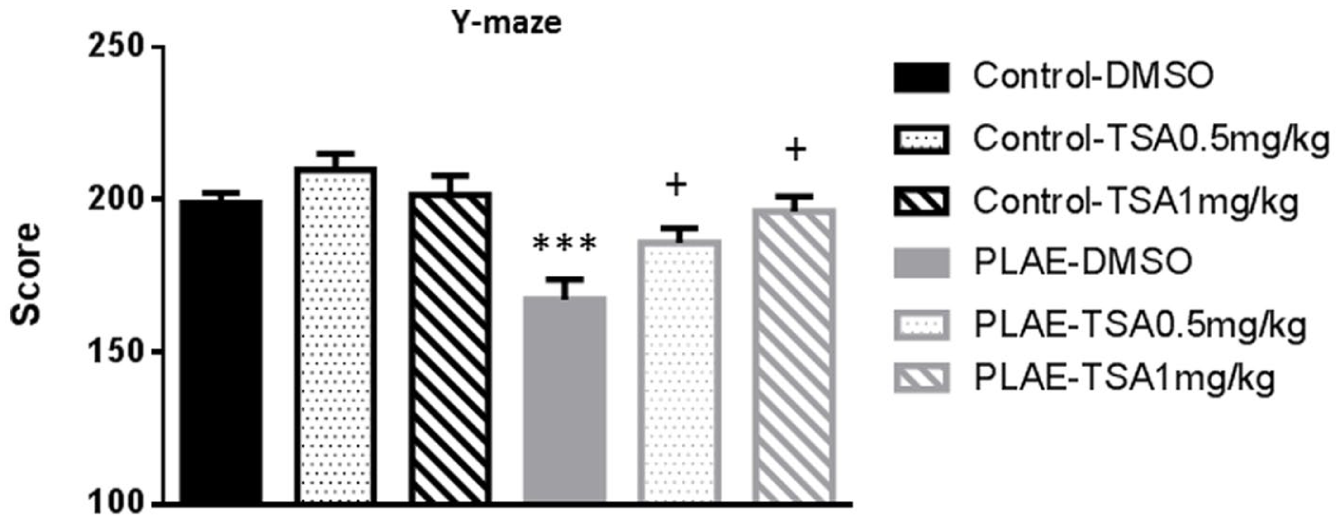
Effects of early alcohol exposure on spontaneous alternation Y-maze test.
Maternal binge-like alcohol drinking impairs HPC memory in the NOR task
Figure 4 shows the results obtained for recognition index in the NOR evaluated one minute after exposure to the familiar object. Two-way ANOVA revealed an effect of group (F(1, 68)=7.598; p<0.01), with no effect of treatment (F(2, 68)=0.028; n.s.) or interaction between factors (F(2, 68)=0.840; n.s.). PLAE mice had lower recognition index scores compared to the control group (p<0.01; Figure 4(a)).
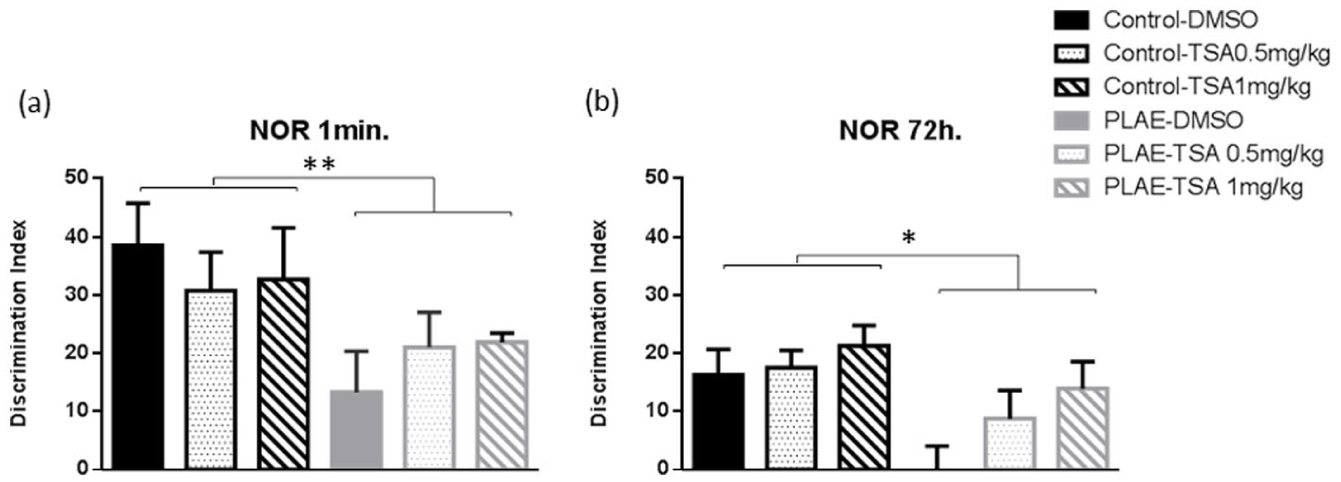
Novel object recognition test in the prenatal and postnatal alcohol/water exposed mice. Data are expressed as the mean±SEM. (a) Novel object recognition test one minute after exposure to the familiar objects. (b) Novel object recognition test 72 hours after exposure to the familiar objects.
When the recognition index was evaluated 72 hours after exposure to the familiar objects, two-way ANOVA revealed significant differences in group (F(1, 66)=5.216; p<0.05) and no effect of treatment (F(2, 66)=2.554; p=0.08) with no observed effect for interaction (F(2, 66)=0.833; n.s.). The animals exposed to alcohol had lower recognition index scores than the control group treated with vehicle (p<0.05; Figure 4(b)).
Maternal binge-like alcohol drinking affects working and spatial memory
Regarding working memory errors (Figure 5(a)–(c)), the three-way ANOVA revealed an effect of group (F(1, 76)=15.3; p<0.001), days (F(11, 836)=18.669; p<0.01), treatment (F(2, 76)=4.951; p<0.01), group×treatment interaction (F(2, 76)=4.108; p<0.05) and group×treatment×days interaction (F(22, 836)=1.692; p<0.05). All groups made fewer errors on days 8–12 compared to days 1–5 (p<0.01; Figure 5(a) and (b)). On day 5, control animals treated with DMSO (p<0.01) made more errors than the group treated with 1 mg/kg of TSA (Figure 5(a)). Furthermore, in PLAE mice, the group of animals treated with DMSO made more errors than the group treated with 0.5 mg/kg of TSA and 1 mg/kg of TSA in days 1 (p<0.05), 6 (p<0.05), 10 (p<0.05) and 12 (p<0.05; Figure 5(b)).
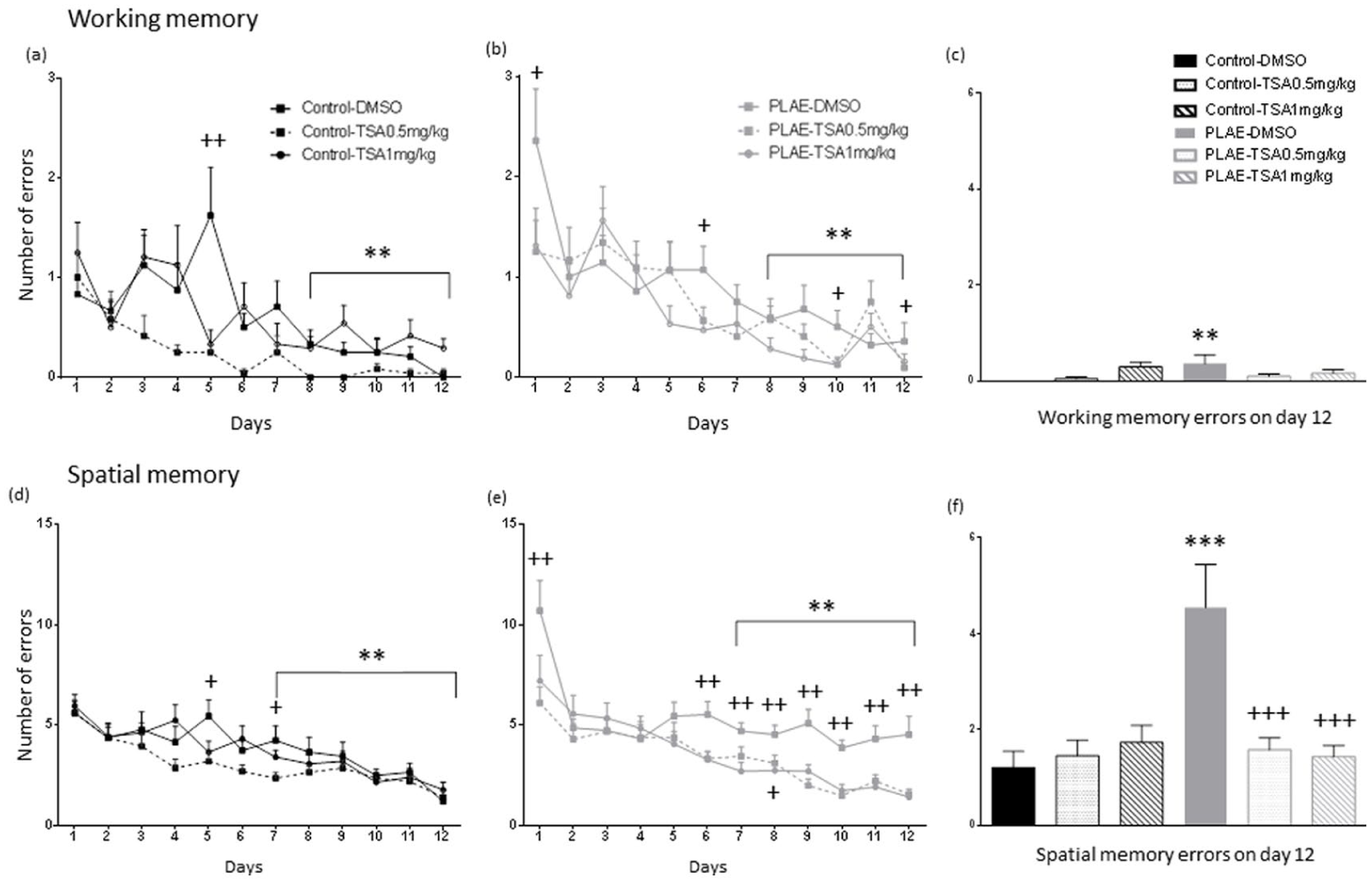
Radial arms maze test in the prenatal and postnatal alcohol/water exposed mice. Data are expressed as the mean±SEM. (a) Control and (b) PLAE working memory errors.
Two-way ANOVA of data for day 12 working memory errors (Figure 5(c)), showed no effect of group (F(1, 76)=1.324; p>0.05), no effect of treatment (F(1, 76)=1.394; p>0.05), with between group×treatment interaction (F(2, 76)=3.228; p<0.05), indicating that PLAE mice treated with vehicle made more errors than the control group (p<0.01).
With regard to spatial memory errors (Figure 5(d)–(f)), the three-way ANOVA revealed an effect of group (F(1, 76)=14.561; p<0.001), days (F(11, 836)=29.793; p<0.01) and treatment (F(2, 76)=16.002; p<0.01), with group×treatment interaction (F(2, 76)=8.978; p<0.001) and group×treatment×days interaction (F(22, 770)=1.69; p<0.05). All group of animals made fewer errors on days 7–12 compared to days 1–4 (p<0.01; Figure 5(d) and (e)). Control animals treated with DMSO made more errors than the group treated with 1 mg/kg of TSA on days 5 and 7 (p<0.05; Figure 5(d)). PLAE mice treated with vehicle made more spatial errors than PLAE animals treated with 0.5 mg/kg of TSA and 1 mg/kg of TSA on days 1 (p<0.01 and p=0.01, respectively), 6 (p<0.01), 7 (p<0.01), 8 (p<0.05 and p<0.01, respectively), 9, 10, 11 and 12 (p<0.01; Figure 5(e)).
Spatial memory errors on day 12 were analysed using a two-way ANOVA (Figure 5(f)). Results showed an effect for group (F(2, 69)=8.070; p<0.01), treatment (F(2, 69)=5.670; p<0.01) and group×treatment interaction (F(2, 69)=3.228; p<0.01). Furthermore, the PLAE group treated with vehicle made more errors than the control group (p<0.001). Therefore, the PLAE group mice treated with vehicle made more errors than PLAE mice treated with TSA at both doses (p<0.001).
Maternal alcohol consumption decreases BDNF levels in offspring
BDNF levels were assessed in the PFC and HPC and showed the effect of early alcohol exposure on neurotrophic factors required for plasticity (Figure 6). In the PFC, two-way ANOVA revealed differences in group (F(1, 24)=7.952; p<0.01), but there were no effects observed in treatment (F(2, 24)= .591; n.s.) or interaction between factors (F(2, 24)=1.306; n.s.). Globally, PLAE animals showed reduced BDNF levels in the PFC compared to the control group (p<0.01; Figure 6(a)).
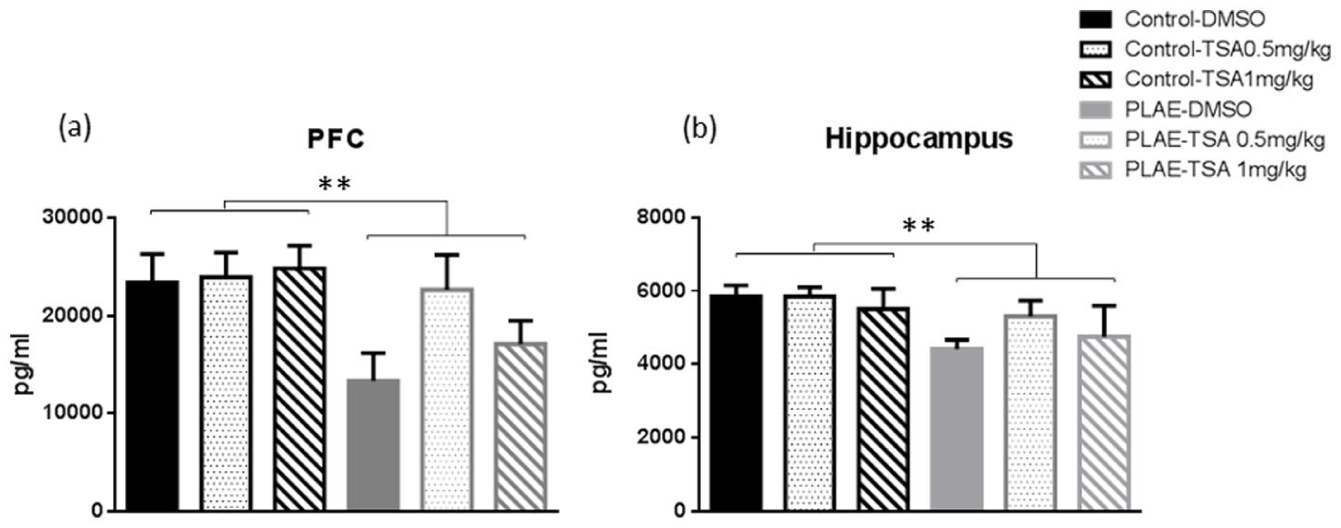
Brain-derived neurotrophic factor (BDNF) levels in the prefrontal cortex (PFC) and hippocampus (HPC) of the prenatal and postnatal alcohol/water-exposed mice. Data are expressed as the mean±SEM. (a) BDNF levels in the PFC.
The analysis of HPC data showed a significant effect for group (F(1, 24)=10.092; p <0.01), but no effect was observed for treatment (F(2, 24)=1.060; n.s.) or interaction between factors (F(2, 24)=0.881; n.s.). Mice exposed to alcohol exhibited decreased BDNF levels in the HPC compared to the control group (p<0.01; Figure 6(b)).
TrkB levels as a consequence of prenatal alcohol/water exposure
TrkB protein expression was assessed in the PFC and HPC of the offspring mice (Figure 7). The effects of early alcohol exposure on the TrkB receptor in the PFC showed no effects for group (F(1, 25)=0.190; n.s.), treatment (F(2, 25)=0.835; n.s.) or interaction between factors (F(2, 25)=0.310; n.s). In addition, in the HPC, no statistical effects were found for group (F(1, 25)=0.010; n.s.), treatment (F(2, 25)=0.399; n.s.) or the interaction between factors (F(2, 25)=2.225; n.s.).
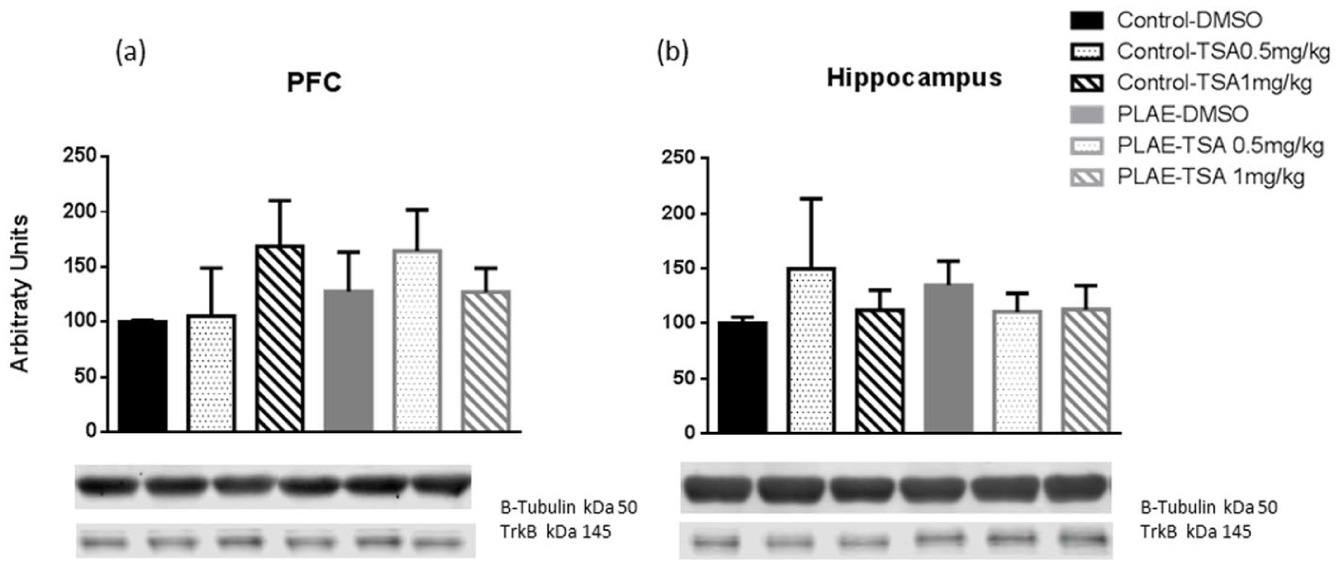
TrkB levels in the PFC and HPC of the prenatal and postnatal alcohol/water exposed mice. Data are expressed as the mean±SEM. (a) TrkB levels in the PFC. (b) TrkB levels in the HPC (n=4–6 per group).
pCREB/CREB protein expression as a consequence of prenatal alcohol/water exposure
pCREB/CREB protein expression was assessed in the PFC and HPC of the offspring mice (Figure 8). No significant effects were observed in the PFC (Figure 8(a)). In the HPC, two-way ANOVA revealed an effect of treatment (F(2, 26)=6.419; p<0.01) but without differences in group (F(2, 26)=0.726; n.s.) or interaction between factors (F(2, 26)=0.085; n.s.). Overall, the main factor of the ANOVA indicated that mice treated with TSA showed a higher protein expression of pCREB/CREB compared to the vehicle group (p<0.01; Figure 8(b)).
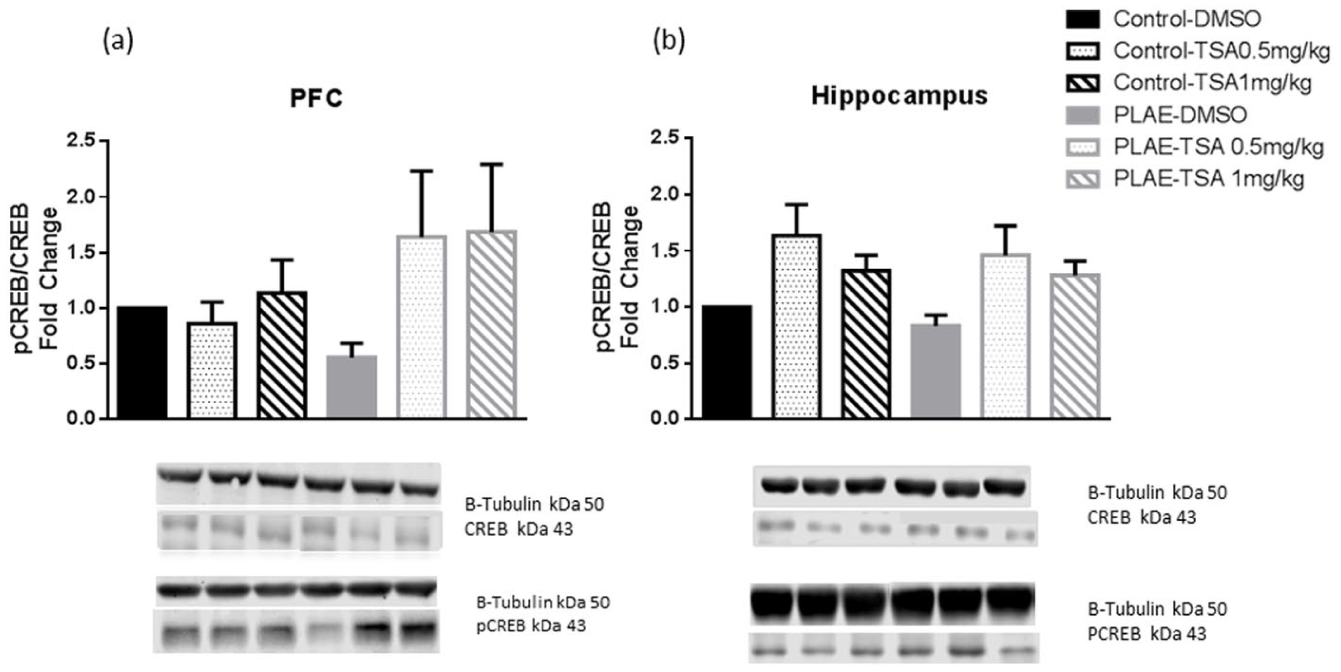
pCREB/CREB levels in the PFC and HPC of the prenatal and postnatal alcohol/water exposed mice. Data are expressed as the mean±SEM. (a) pCREB/CREB levels in the PFC. (b) pCREB/CREB levels in the HPC (n=4–6 per group).
Effects of PAE on HDAC4 levels in the PFC and HPC
HDAC4 levels were assessed in the PFC and HPC of the offspring (Figure 9). Conversely, no changes were observed in the PFC (Figure 9(a)). In the HPC, (Table 3) two-way ANOVA revealed an interaction between treatment×group factors (F(2, 18)=5.377; p<0.05). PLAE animals treated with vehicle presented higher HDAC4 activity compared to the control group (p<0.05). Furthermore, PLAE mice treated with 1 mg/kg of TSA presented lower levels of HDAC4 compared to the PLAE control group (p<0.01; Figure 9(b)).
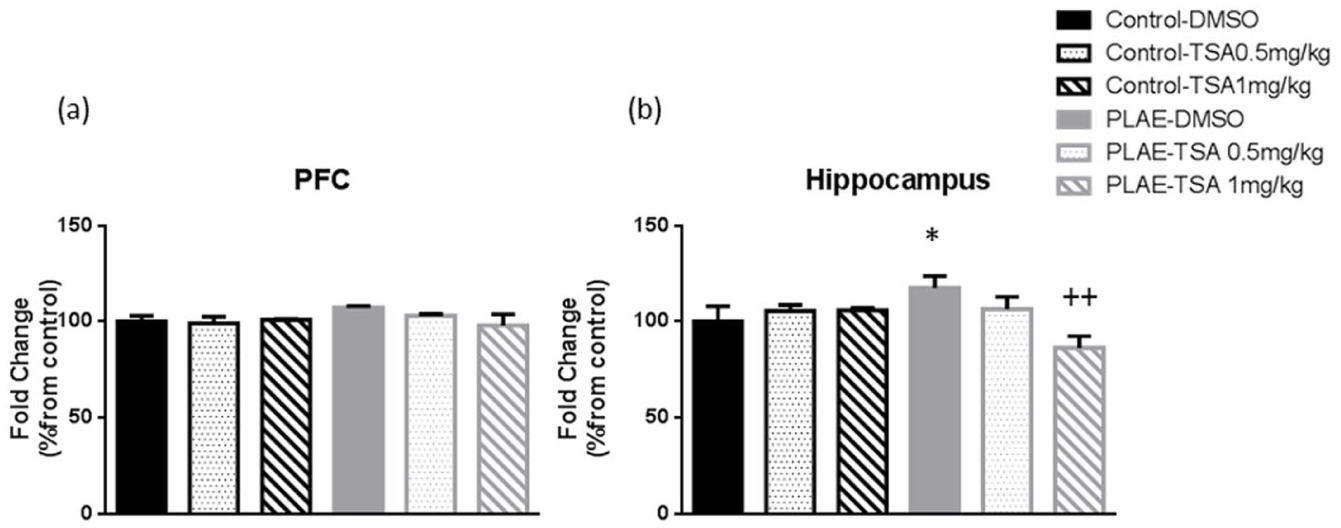
Effects of prenatal and postnatal alcohol/water exposure on HDAC4 levels in the PFC and HPC. Data are expressed as the mean±SEM. (a) HDAC4 levels in the PFC. (b) HDAC 4 levels in the HPC.
Effects of prenatal and postnatal alcohol/water exposure on HDAC4 levels in the hippocampus.

Data are expressed as the mean±SEM (n=4). HDAC 4 levels in the HPC. *p<0.05 PLAE–vehicle versus control–vehicle; ++p<0.01 PLAE–TSA 1 mg/kg versus PLAE–vehicle (Bonferroni post hoc test).
Maternal binge-like alcohol drinking decreases DCX in offspring
A quantitative analysis of DCX was performed in the DG of the HPC. Two-way ANOVA revealed a significant effect of group (F(1, 18)=5.985; p<0.05), no effect of treatment (F(1, 18)=1.265; n.s.) and a group×treatment interaction (F(2, 18)=8.779; p<0.01). PLAE animals presented a decrease in DCX signal in the DG compared to the control group (p<0.05; Figure 10). In addition, the DCX signal cell was reduced in PLAE mice treated with vehicle compared to the control vehicle group (p<0.001). Furthermore, animals exposed to alcohol and treated with TSA presented an increased DCX signal compared to PLAE mice treated with vehicle (p<0.05, TSA 0.5 mg/kg; p<0.01, TSA 1 mg/kg; Figure 10).
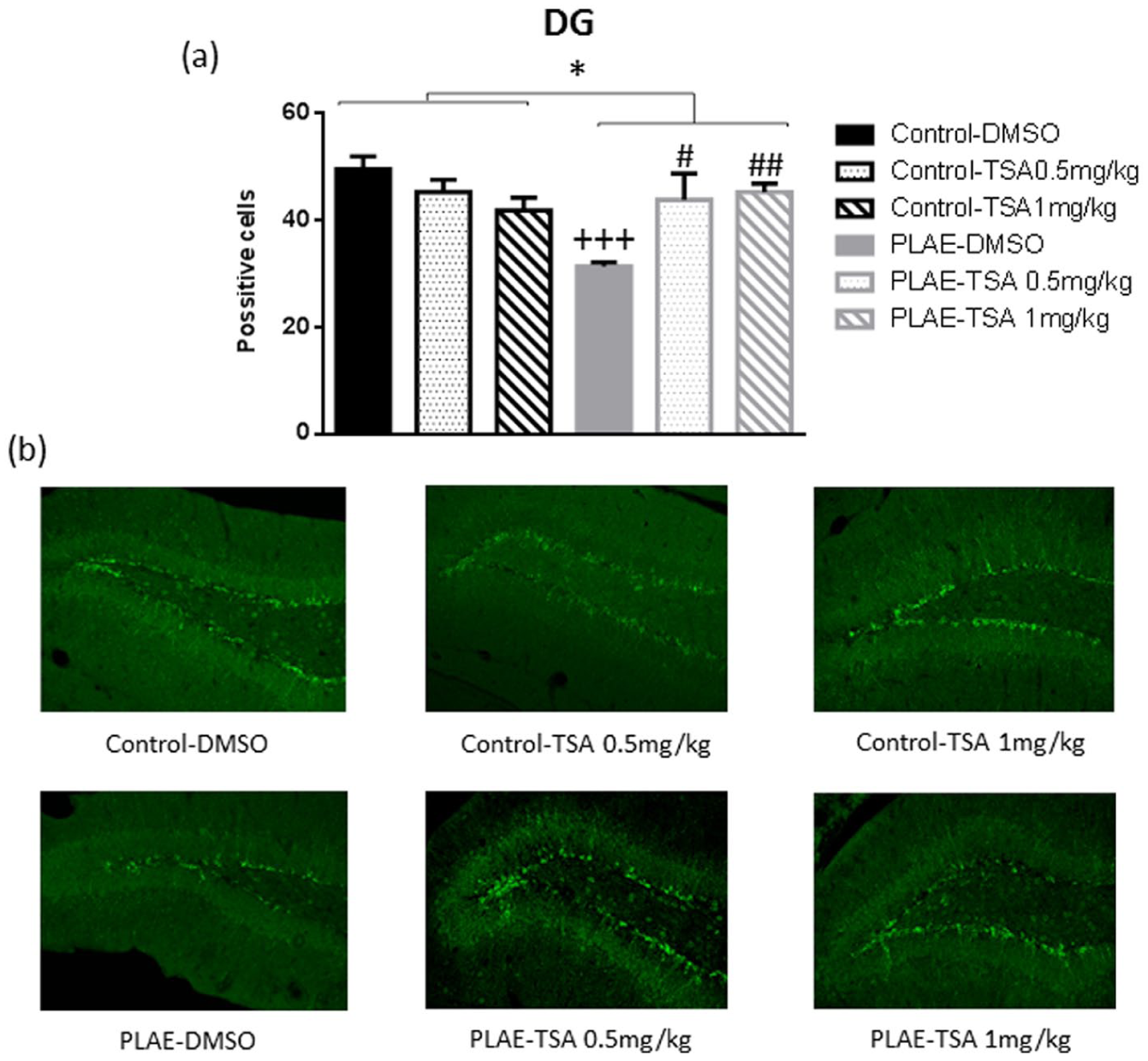
Effects of maternal alcohol binge drinking in neurogenesis (doublecortin content (DCX)). (a) Counts of DCX-positive cells in the dentate gyrus (DG). (b) Representative DG section for DCX immunohistochemistry.
Discussion
Early life experiences can impact adult behaviour, and we propose that such alterations involve epigenetic mechanisms. The present study demonstrates that adolescent mice exposed to PLAE showed behavioural alterations, such as increased anxiogenic-like responses, working and spatial memory impairments and recognition memory deterioration. Most of these effects were subject to significant recovery with TSA treatment. PLAE also produced a decrease in BDNF levels in the HPC and the PFC and reduced neurogenesis (DCX) in the DG. Furthermore, alcohol exposure during the developmental period increased HDAC4 activity in the HPC.
In our study, females were exposed to a voluntary mouse model of binge-like alcohol drinking known as the DID test (Cantacorps et al., 2017, 2018; Rhodes et al., 2005), with a total of six binge alcohol episodes weekly (three-week gestation and three-week lactation). Alcohol exposure during the third trimester equivalent can cause deleterious effects in humans. In fact, the third trimester is associated with a period of rapid neuronal growth and proliferation, referred as the ‘brain growth spurt’. In rodents, the ‘brain growth spurt’ occurs during the first one or two weeks after birth (Barron et al., 2016). We have already found that the effects of alcohol are more detrimental when the mother receives alcohol during both periods – gestation and lactation – thus covering the whole intrauterine and early postnatal development periods in mice (Cantacorps et al., 2017, 2018). Our results showed an increase in alcohol intake during the binge drinking sessions (four-hour access) compared to the three previous days (two-hour access) during weeks 3–6. Additionally, we found an increase in water and alcohol consumption during the lactational period of the DID procedure compared to the prenatal period. This may be due to an increased physiological demand to consume fluid in order to support milk production (Bentley, 1998; Cantacorps et al., 2017).
The levels of alcohol in the pups did not reach levels of intoxication at the particular time points analysed. The blood of the pups was extracted immediately after the four-hour period in which females were exposed to alcohol. Dams at this period were probably focused on drinking alcohol and not on breastfeeding their offspring, and consequently the BAC levels in the offspring did not reach the expected high levels. In spite of these low BAC levels in pups, we observed changes in the behavioural and biochemical parameters, indicating that continued exposure to alcohol produces a negative impact in the nursing pups.
The literature shows that prenatal alcohol consumption produces alterations in the emotional and cognitive behaviour of offspring, and such effects persist throughout life (Cantacorps et al., 2017; Hamilton et al., 2014; Kozanian et al., 2018; Macllvane et al., 2016; Schambra et al., 2017; Thomas et al., 2010). Indeed, the offspring of females exposed to binge alcohol drinking during the developmental period showed an increase in anxiety-related behaviour on the EPM, with no changes in locomotor activity. Such results are consistent with earlier studies reporting an increase in anxiety-related behaviour in PAE animals (Abbott et al., 2016; Cantacorps et al., 2018). Moreover, impairments in recognition memory were reflected in the NOR test, as PLAE mice failed to recognise the new object 72 hours after the first exposure. Similar results were found in working memory (assessed by the Y-maze and RAM), although in these cases, an improvement of the performed tasks was observed in the PLAE mice treated with TSA. On the basis of such findings, we propose that TSA is capable of reversing the impairments and disabilities in cognitive responses caused by PAE. A possible explanation for the deficits observed in PLAE mice may be due to the fact that PAE reduces the size of the anterior HPC in mice (Fish et al., 2018). Moreover, significant thickening of the frontal cortex was also observed in PLAE mice (Abbott et al., 2016). HDAC inhibitors such as TSA were reported to rescue memory deficits in aged mice by increasing long-term potentiation and elevating the level of histone acetylation in the HPC (Francis et al., 2009; Kilgore et al., 2010; Peleg et al., 2010). Furthermore, following the same procedure, a previous study by our group showed no relevant differences in maternal care behaviour in mothers exposed to alcohol or water, suggesting that both groups of dams behaved similarly with regard to the care of the pups (Cantacorps et al., 2019), discarding that the lack of appropriate maternal care could be responsible for the alterations observed in the present study.
Supporting the idea that histone acetylation acts as a molecular memory aid, some research has proposed that the administration of HDAC inhibitors was able to potentiate memory in rodents (Fontano-Lozano et al., 2008; Gräff et al., 2013a, 2013b; Guan et al., 2009; Monsey et al., 2011). Concretely, TSA treatment enhanced contextual and cue fear memory, object recognition memory and memory reconsolidation in rodents (Gräff et al., 2013a). In addition, in different situations such as repeated neonatal exposure to isoflurane, which affects memory formation, Zhong et al. (2015) observed that TSA enhanced contextual fear conditioning. Correspondingly, treatment with TSA (1 mg/kg) attenuated high-fat diet-induced cognitive deficits in mice (Sharma et al., 2015). Similarly, several previous reports (Francis et al., 2009; Itzhak et al., 2012; Kilgore et al., 2010) also demonstrated that memory deficits in some genetic mouse models (i.e. transcriptional coactivator CREB-binding protein mutant mice) were mitigated by HDAC inhibitor treatments, whereas normal cognitive function in wild-type mice was unaffected.
Kim et al. (2012) provided evidence that the specific enzyme HDAC4, grouped in class IIa HDAC, is required for learning and memory and synaptic plasticity in mice. TSA inhibits both class I and class II HDAC enzymes, including HDAC4 (Cantley et al., 2017). Our results showed increased HDAC4 enzyme activity associated with cognitive impairments in the PLAE animals, which were reversed by TSA only in the PLAE group of mice. In contrast with our findings, conditional deletion of HDAC4 leads to impaired learning, memory and continued synaptic plasticity (Kim et al., 2012; Ronan et al., 2013). The expression of truncated HDAC4 allele lacking the deacetylase domain showed similar defects, such as conditional deletion, in transgenic rodents (Sando et al., 2012). The apparent discrepancies with previous findings are probably due to the non-specific targeting of the HDAC enzymes by TSA, thus suggesting that the broad-scale manipulation of class I or class II HDACs has divergent effects on these processes.
Several studies have demonstrated that HDAC inhibitors also improved memory function through the activation of specific genes mediated by the CREB–CREB binding protein transcriptional complex (Green et al., 2008; Haettig et al., 2011; McQuown et al., 2011; Vecsey et al., 2007), which in turn could stimulate the expression of the active form of CREB (phosphorylated CREB). In our study, TSA administration increased the protein expression of pCREB/CREB in the HPC. The increase in the pCREB/CREB ratio might allow neurons to translate extracellular stimulation into prolonged cellular responses, enabling synaptic plasticity and memory consolidation. In this sense, Vecsey et al. (2007) found an enhancement of memory consolidation for contextual fear conditioning induced by intra-HPC injection of TSA.
To explore the molecular mechanisms involved in PLAE cognitive deficits further, we assessed BDNF levels. A decrease in BDNF expression in both structures (PFC and HPC) in animals exposed to alcohol during the developmental period was observed without changes in TrkB expression. This would indicate that the endogenous regulation of the BDNF system can be affected by different conditions, revealing that the system would be partially modified, without showing changes in receptor expression (Ray et al., 2014; Smith et al., 1995; Xu et al., 2004). Previous findings confirm that alcohol consumption during the developmental period alters BDNF protein levels in different brain structures (Barbier et al., 2008; Boschen et al., 2015; Caldwell et al., 2008; Feng et al., 2005; Heaton et al., 2003). The variability of the effects (i.e. increases or decreases) depends on different factors, including alcohol dosage, exposure window and brain region, thus suggesting developmental time-specific vulnerability. Pandey et al. (2015) observed that adolescent intermittent alcohol exposure increased HDAC activity and HDAC2 levels in the amygdala, leading to reduced histone acetylation in synaptic plasticity-related genes such as BDNF in rats. Moreover, TSA was able to reverse these alterations (Pandey et al., 2015, 2017; You et al., 2014).
Previous research has described adolescents as being more sensitive to acute alcohol-induced inhibition of neurogenesis than adults (Crews et al., 2006). In addition, adult binge alcohol exposure decreases neurogenesis during alcohol intoxication, an effect that is reversed during the abstinence period (Crews and Nixon, 2008). Sakharkar et al. (2016) also reported that exposure to intermittent alcohol intake during adolescence decreased neural progenitor proliferation and also reduced the presence of immature neurons in the HPC. Different studies using animal models have reported that intermittent alcohol exposure during adolescence reduces neurogenesis markers, regardless of the animal strain or alcohol exposure paradigm (Briones and Woods, 2013; Broadwater et al., 2014; Ehlers, 2013; Vetreno and Crews, 2015). Accordingly, PLAE mice presented a decrease in the DCX signal in the DG, which was reversed by TSA treatment, thus supporting the anti-inflammatory and neuroprotective effects of TSA in in vitro and in vivo models of different pathologies (Chen et al., 2007; Fleiss et al., 2012; Harrison and Dexter, 2013). BDNF adjusts some aspects of neuronal development and function, including precursor cell proliferation (Davies, 1994). BDNF signalling would seem to play an important role in neurogenesis, and changes in BDNF would therefore have deleterious consequences in the neurogenesis of specific brain regions (Chen et al., 2007; Bath and Lee, 2010; Wei et al., 2015). However, on the basis of our results, we would rule out BDNF as a major role player in the context of TSA’s effects on neurogenesis.
Conclusion
The results from our study would suggest that maternal binge-like alcohol consumption can induce anxiety-like behaviour and memory and learning dysfunction in adolescent offspring. Molecular alterations described in the PFC and HPC of PLAE mice may be associated with the behavioural impairments caused by alcohol exposure. Some of the observed modifications were significantly reversed following treatment with TSA, in particular the reduced neurogenesis in the DG. Our study supports the participation of HDAC enzymes in cognitive and emotional alterations induced by binge alcohol consumption during gestation and lactational periods, and proposes potential benefits of HDAC inhibitors for some aspects of FASD. Pharmacological manipulations acting on epigenetic mechanisms could represent a potential target in the management of FASD, but at the present moment, further investigations are needed.
Footnotes
Acknowledgements
The authors thank Gerald-Patrick Fannon for his English proofreading and editing of the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This work was supported by the Ministerio de Economía y Competitividad (Spanish Ministry of Economy and Competitiveness – grant number SAF2016-75966-R-FEDER), and the Spanish Ministry of Health (Retic-ISCIII-RD16/0017/0010 and PNSD 2018/007). SM-R received a postdoctoral fellowship from the Conselleria d’Educació, Investigació, Cultura i Esport (APOSTD/2017/102), Generalitat Valenciana, Spain. LC received an FPI grant (BES-2014-070657) from the Ministerio de Economía y Competitividad, The Department of Experimental and Health Sciences (UPF), ‘Unidad de Excelencia María de Maeztu’ funded by the MINECO (ref. MDM-2014-0370).