
Abstract
Background:
Psilocybin has shown promise as a treatment for depression but its therapeutic mechanisms are not properly understood. In contrast to the presumed actions of antidepressants, we recently found increased amygdala responsiveness to fearful faces one day after open-label treatment with psilocybin (25 mg) in 19 patients with treatment-resistant depression, which correlated with treatment efficacy.
Aims:
Aiming to further unravel the therapeutic mechanisms of psilocybin, the present study extends this basic activation analysis. We hypothesised changed amygdala functional connectivity, more precisely decreased amygdala-ventromedial prefrontal cortex functional connectivity, during face processing after treatment with psilocybin.
Methods:
Psychophysiological interaction analyses were conducted on functional magnetic resonance imaging data from a classic face/emotion perception task, with the bilateral amygdala and ventromedial prefrontal cortex time-series as physiological regressors. Average parameter estimates (beta weights) of significant clusters were correlated with clinical outcomes at one week.
Results:
Results showed decreased ventromedial prefrontal cortex-right amygdala functional connectivity during face processing post- (versus pre-) treatment; this decrease was associated with levels of rumination at one week. This effect was driven by connectivity changes in response to fearful and neutral (but not happy) faces. Independent whole-brain analyses also revealed a post-treatment increase in functional connectivity between the amygdala and ventromedial prefrontal cortex to occipital-parietal cortices during face processing.
Conclusion:
These results are consistent with the idea that psilocybin therapy revives emotional responsiveness on a neural and psychological level, which may be a key treatment mechanism for psychedelic therapy. Future larger placebo-controlled studies are needed to examine the replicability of the current findings.
Keywords
Introduction
Serotonergic psychedelics (e.g. lysergic acid diethylamide (LSD) and psilocybin) have shown promising results in disorders such as obsessive-compulsive disorder (Moreno et al., 2006), end-of-life psychological distress (Gasser et al., 2014; Griffiths et al., 2016; Grob et al., 2011; Ross et al., 2016), addiction (Bogenschutz et al., 2015; Johnson et al., 2017) and depression (Carhart-Harris et al., 2016a, 2018a; Osorio Fde et al., 2015), when administered under supportive conditions. Our open-label study of psilocybin with psychological support (including one preparation and integration session, as well as support immediately before, during and after the acute drug session) for treatment-resistant depression (TRD) showed reductions in depressive symptoms in all 19 patients from baseline to one week, which was sustained in the majority for at least 3–5 weeks and was predicted by the quality of the acute psychedelic experience (Carhart-Harris et al., 2016a, 2018a; Roseman et al., 2017). Long-term follow-ups at six months revealed that of the nine responders (at five weeks), only three had relapsed, suggesting stable antidepressant effects for some patients (Carhart-Harris et al., 2018a; Watts et al., 2017). Despite accumulating evidence on its efficacy, the therapeutic mechanisms of the psychedelic treatment model are not well-understood (although see Carhart-Harris, 2019) and demand further investigation.
Acutely, serotonergic psychedelics interact with multiple receptor types (reviewed in Nichols, 2016), but have high affinity for the serotonin (5-HT) 2A receptor, which mediates their hallucinogenic effects via full or partial agonism (Preller and Vollenweider, 2018; Madsen et al., 2019). This receptor’s role in increased excitability, plasticity and adaptability has recently been put forward by Carhart-Harris and Nutt (2017) as potentially underlying their therapeutic effects. Psychedelic-induced plasticity and changes to network level functioning (e.g. increased cortical entropy, disintegration of networks and an enhanced communication between normally segregated brain regions) have been identified as underlying the acute psychedelic state (Carhart-Harris et al., 2012, 2016b; Lebedev et al., 2015; Majic et al., 2015; Muthukumaraswamy et al., 2013; Roseman et al., 2014). However, considering the sustained antidepressant effects and improvements in well-being observed after psychedelic experiences (e.g. Carhart-Harris et al., 2016a, 2018a; Griffiths et al., 2016), it is crucial to investigate potential neural changes beyond the acute state.
Previous work has identified some post-acute neurofunctional changes after psychedelics: A recent study in a group of healthy ayahuasca-experienced individuals showed increased resting-state functional connectivity (RSFC) between the posterior and anterior cingulate cortex, and between the anterior cingulate and the right medial temporal lobe, 24 h after one ayahuasca dose, a N,N-dimethyltryptamine (DMT)-containing plant brew (Sampedro et al., 2017). Similarly, RSFC analysis on data acquired at pre-treatment baseline and one day after a high dose treatment session with psilocybin for TRD revealed the following changes: (a) increased connectivity within the default mode network (DMN) and decreased parahippocampal-prefrontal RSFC; both predictive of treatment response at five weeks, and (b) decreased cerebral blood flow in the temporal cortex (including the amygdala) that correlated with reductions in depressed mood (Carhart-Harris et al., 2017).
In the same group of patients, which also constitutes the sample of the present study, a functional magnetic resonance imaging (fMRI) paradigm was used to assess amygdala responsiveness to emotional faces pre- and post-treatment with psilocybin (Roseman et al., 2018). The results of this study revealed an increased response in the right amygdala to fearful and happy faces one-day post-psilocybin, and right amygdala increases to fearful versus neutral faces which was predictive of depressive symptoms at one week and treatment response at one-day, one-week, and three-weeks (Roseman et al., 2018). Additionally, whole-brain analyses revealed increased activity in visual areas across all face conditions (happy, fearful, neutral) post-psilocybin, suggesting not just changes in amygdala reactivity, but also responsiveness in other brain regions. Additionally, Grimm et al. (2018) provided compelling evidence for decreased amygdala functional connectivity (FC) to the striatum and the frontal pole during angry and happy discrimination respectively under acute psilocybin as compared with placebo in healthy volunteers. However, post-acute FC alterations have yet to be explored. Moreover, there are reasons to believe that post-acute brain changes are quite different to the acute brain effects of psychedelics (Carhart-Harris et al., 2017) and are obviously vital to study given their potential relevance to therapeutic mechanisms.
The changes in post-acute amygdala reactivity in depressed patients (Roseman et al., 2018) and acute amygdala reactivity under psilocybin (Grimm et al., 2018) are particularly striking, as amygdala involvement, and particularly its hypersensitivity to negative stimuli, has long been thought to be a component of clinical depression, and thus a target for antidepressant medications (Drevets et al., 1992; Godlewska et al., 2012; Harmer et al., 2017; Ma, 2015). Dysfunctions in amygdala-prefrontal cortex (PFC) circuits and decreased amygdala-PFC FC are hypothesised to underlie disorders with disturbed emotional processing, such as depression and post-traumatic stress disorder (PTSD; Koenigs and Grafman, 2009; Kong et al., 2013; Ramasubbu et al., 2014; Stevens et al., 2013). Current neural models of depression and anxiety disorders emphasise abnormalities in both limbic responsiveness and metabolism and top-down emotional control mechanisms (Ramasubbu et al., 2014).
Accordingly, from a traditional antidepressant perspective, our previous finding of increased amygdala responsiveness to negative faces (that was predictive of better clinical outcome) may seem somewhat counterintuitive. In order to further investigate the therapeutic mechanisms of psilocybin, in the present study we aimed to examine the neural mechanisms responsible for this change in amygdala responsiveness post psilocybin. We predicted changes in amygdala FC during the same emotional face paradigm performed at pre-treatment baseline and one day after psilocybin for TRD. More specifically we predicted decreased FC between the amygdala and the ventromedial PFC (vmPFC) during face processing after psilocybin. The vmPFC is an important component of the emotional processing circuitry, shown to have a top-down inhibitory effects on limbic regions (Hariri et al., 2000) and is implicated in emotional control, inhibition and regulation (Rosenkranz et al., 2003). Changes in amygdala-PFC connectivity are also a common finding in depressed patients (e.g. Dannlowski et al., 2009; Moses-Kolko et al., 2010). We hypothesised that the reduction of FC between the vmPFC and amygdala might account for the increased amygdala responsivity after psilocybin, as shown previously by Roseman et al. (2018). Lastly, we predicted that altered amygdala and vmPFC FC might relate to post-treatment changes in depression severity and rumination tendencies. Rumination is a pattern of recursive self-directed and self-reflective thinking focused on one’s negative emotions (Cooney et al., 2010) and defined as the compulsively focused attention on the symptoms of one’s distress, and on its possible causes, consequences and implications, as opposed to its solutions (Nolen-Hoeksema, 1991). Alongside basic depression scores, rumination scores using the Ruminative Response Scale (RRS; Treynor et al., 2003) served as a secondary clinical outcome of interest. Rumination was chosen as a symptom of interest as: (a) it is a strong vulnerability factor in the development and maintenance of depressive episodes, and (b) it has been found to have different neural underpinnings in depressed patients as compared with healthy controls (Cooney et al., 2010).
Methods
Procedure
The full study procedure is reported in Carhart-Harris et al. (2016a, 2018a) and Roseman et al. (2018). For an overview of the trial schedule and study interventions, also see the Supplementary Material Figure 1. The present study focused on changes in brain FC after psilocybin in 19 patients with TRD who underwent two psilocybin-assisted therapy sessions a week apart, first 10 mg p.o. (test dose), then 25 mg, p.o. (therapeutic dose). Provided psychological support consisted of three components: one preparation session with the allocated trial psychiatrists (lasting 4 h) before the first dose, acute and peri-acute support (i.e. before, during and immediately after dosing) and one integration session with the same psychiatrists one day after the high dose. Seed-based FC analysis using psychophysiological interaction (PPI) methods was conducted on fMRI data collected during an emotional face paradigm pre- and post-psilocybin. Baseline fMRI scanning took place prior to any intervention, and the post treatment fMRI scan occurred the morning (10:00) after the high-dose psilocybin session and prior to any subsequent psychological integration. Clinical outcome measures were administered at baseline and at multiple time-points after the high-dose session (see the Method section ‘Relationship with clinical outcomes’ for details); the one week post-treatment time-point constitutes the primary clinical endpoint of the present analysis.
Inclusion criteria were (a) a diagnosis of moderate to severe major depression, as defined by a score of 16+ on the 21-item Hamilton Depression Rating Scale (HAM-D; Hamilton, 1960); and (b) treatment-resistance as defined by lack of improvement despite two adequate courses of pharmacologically distinct antidepressant medications over at least six weeks each, within the present depressive episode. Additionally, patients were asked to stop their antidepressant medication before study entry. Typically, washout occurred under close supervision and over a period of at least two weeks before study entry. The main exclusion criteria were a current or past diagnosis of a psychotic disorder or an immediate family member with a diagnosed psychotic disorder, a history of serious suicide attempts and/or mania, pregnancy and current drug or alcohol dependence.
Written informed consent was obtained from all participants during the screening visit. Ethical approval for the clinical trial was sought and granted by the National Research Ethics Service (NRES) London-West London, the UK Medicines and Healthcare Products Regulatory Agency (MHRA) and Imperial College London’s Joint Research and Complication Organisation (JRCO) – who also sponsored the trial. Psilocybin was obtained from THC-Pharm (Frankfurt, Germany) and formulated into the investigational medicinal product (5 mg psilocybin in size 0 capsules) by Guy’s and St Thomas’ Hospital’s Pharmacy Manufacturing Unit (London, UK).
Patient characteristics
In total, 20 patients underwent the two psilocybin-assisted therapy sessions. Nineteen patients (six females, Mage=44.7, standard deviation (SD)=10.9, age range: 27–64 years) completed both scanning sessions and constitute the current study sample (patient 17 did not complete the post-treatment fMRI scan and was therefore excluded from all analyses).
At baseline, 17 of the 19 patients met the criteria for severe or very severe depression (self-rated Quick-Inventory of Depressive Symptoms (QIDS-SR16)⩾16), with the other two classifying as moderately depressed (15⩾QIDS-SR16⩽11). On average, patients had been diagnosed with major depression for 17.7 years (SD=8.4, range=7–30 years) and had tried 4.6 (SD=2.6) antidepressant medications. Seventeen patients had tried at least one form of psychotherapy in the past. At the time of enrolment to the study, nine patients were taking antidepressant medication. Patients were asked to discontinue their antidepressant medication during the period of the trial, which all but two patients adhered to (patient 2 did not stop and continued venlafaxine during both dosing and magnetic resonance imaging (MRI) scans; patient 6 only stopped the opioid tramadol after the first psilocybin session, i.e. one week before the high dose session/post-treatment MRI scan). For a more detailed description of the study sample, see Table 1 in Carhart-Harris et al. (2018a).
Summary of results of the seed-to-voxel psychophysiological interaction analysis comparing functional connectivity (FC) during face processing (irrespective of facial expression) with rest (simple model) for both regions of interest (ROIs), the bilateral amygdala (AMG) and ventromedial prefrontal cortex (vmPFC).
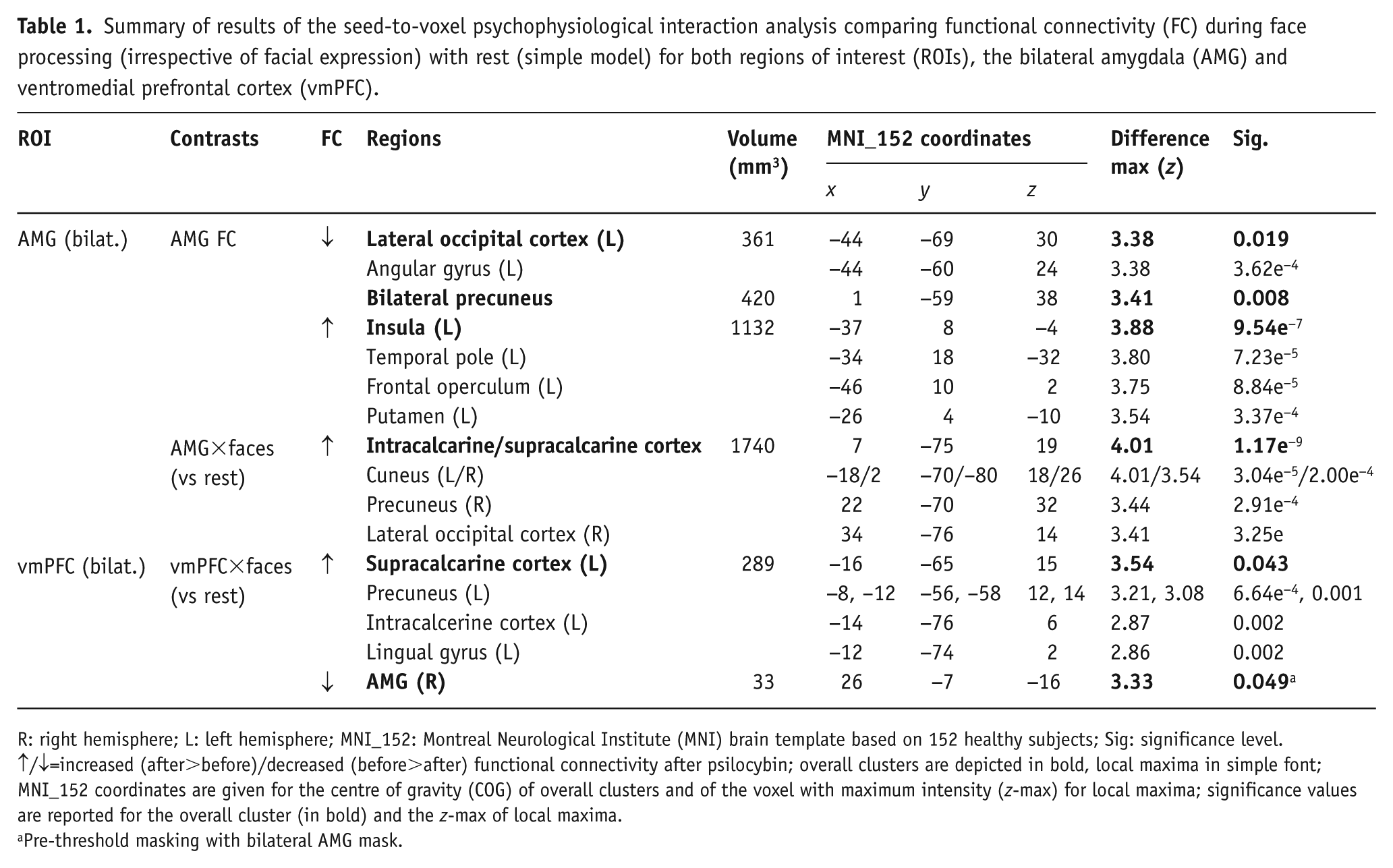
R: right hemisphere; L: left hemisphere; MNI_152: Montreal Neurological Institute (MNI) brain template based on 152 healthy subjects; Sig: significance level.
↑/↓=increased (after>before)/decreased (before>after) functional connectivity after psilocybin; overall clusters are depicted in bold, local maxima in simple font; MNI_152 coordinates are given for the centre of gravity (COG) of overall clusters and of the voxel with maximum intensity (z-max) for local maxima; significance values are reported for the overall cluster (in bold) and the z-max of local maxima.
Pre-threshold masking with bilateral AMG mask.
fMRI design
Imaging (anatomical and functional scans) was performed on a 3T Siemens Tim Trio using a 12-channel head coil at Invicro, London, UK. For a detailed description of the scanning procedure, see Roseman et al. (2018). Functional T2*-weighted echo-planar images (EPIs) were acquired for the functional task scans (3 mm isotropic voxels, repetition time (TR)=2000 ms, echo time (TE)=31 ms, 36 axial slices, 192 mm in-plane field-of-view, flip angle=80°, bandwidths=2298 Hz/pixel, in-plane acceleration=2, number of volumes=245).
Patients underwent counter-balanced versions of an emotional face paradigm on two occasions, at baseline and the morning after the high-dose session. Faces were presented in a blocked-design that lasted for eight minutes in total. Patients were shown blocks of fearful, neutral or happy faces of both genders selected from the Karolinska Directed Emotional Faces set (Goeleven et al., 2008 (see Supplementary Material Figure 2(a)). Patients passively viewed the faces, but were instructed to press a button with their thumb upon each face presentation, in order to check that they were still attentive. Faces were presented for three seconds, with each face block consisting of five faces of the same expression (i.e. 15 s per block). Each block was repeated eight times, inter-mixed with rest blocks (15 s, fixation cross) and presented in a pseudo-random sequence, resulting in 32 face blocks in total.
fMRI analysis
Preprocessing
Blood-oxygen-level-dependent (BOLD) pre-processing was conducted with four different software packages: FMRIB Software Library (FSL) 5.0.4 (Smith et al., 2004), AFNI (Cox, 1996), Freesurfer (Dale et al., 1999) and Advanced Normalization Tools (ANTS; Avants et al., 2008). More precisely, the following pre-processing steps were applied: (a) motion correction (3dvolreg, AFNI); (b) brain extraction (BET in FSL); (c) rigid body registration to anatomical scans (BBR in FSL); (d) non-linear registration to 2 mm MNI brain (Symmetric Normalization in ANTS); (e) scrubbing using a framewise displacement threshold of 0.90 mm as recommended for task fMRI (Siegel et al., 2014). Scrubbed volumes were replaced with the average of surrounding volumes; (f) spatial smoothing (FWHM) of 6 mm (3dBlurInMask, AFNI); (g) a high-pass filter of 0.01 Hz; and (h) regressing out of six motion-related parameters as nuisance regressors (three translations, three rotations, high-pass filtered with the same 0.01 Hz filter). A more comprehensive overview of the pre-processing pipeline is given in Roseman et al. (2018).
General linear models (GLMs)
The BOLD-weighted fMRI data were then analysed with a standard GLM approach using the FEAT module in FSL. Anatomical images were segmented into grey/white matter (WM) and cerebro-spinal fluid (CSF) masks using FMRIB’s Automated Segmentation Tool (FAST). These CSF and WM masks (thresholded at 0.75) were used to derive time-series from the functional data, which were entered as nuisance regressors in all GLMs.
In order to identify brain regions where psilocybin affected the FC of the amygdala and vmPFC, separate PPI analyses were conducted using seed regions of interest (ROIs) in the (a) amygdala and (b) vmPFC (Supplementary Material Figure 2(c)). The amygdala ROI was defined using a bilateral amygdala mask from the Subcortical Harvard-Oxford Atlas included in FSL (https://fsl.fmrib.ox.ac.uk/fsl/fslwiki/Atlases) while the vmPFC ROI was drawn on a standard brain and was the same as used in previous studies (Carhart-Harris et al., 2012, 2015). Mean time-series were extracted from the pre-processed functional data-series for each subject, for both ROIs and entered as physiological regressors in our PPI analyses. Two different PPI analyses were conducted (Supplementary Material Figure 2(b)):
A simple model including faces (irrespective of expression) compared to the rest-blocks and its interaction with the ROI time-series as the PPI regressor as the primary analysis.
A complex model including separate regressors for all facial expression conditions (happy, fearful, neutral) as compared to the rest-period, and their respective interactions with the ROI time-series (i.e. the PPI regressors) as a secondary analysis.
Face regressors derived from the onset times of the stimulus conditions were convolved with a gamma function, in order to simulate a canonical haemodynamic response function. Reasons for conducting both the simple and the complex PPI model were: (a) existing literature showing similar activation patterns for negative, neutral and positive emotional faces (e.g. Cunningham et al., 2008; Fusar-Poli et al., 2009; Somerville et al., 2004) and (b) power considerations due to the relatively small sample size. The simple model should provide more robust, though less specific, results, while the complex model enables a more comprehensive exploration of the data, and comparisons between different facial expressions.
Contrasts were defined that compared activity related to each regressor, i.e. (a) for faces, the ROI and their interaction in comparison to rest in the simple model; (b) for the different facial expressions (happy, fearful, neutral), the ROI, and their interactions relative to rest, as well as between the interaction terms for fearful vs neutral faces (ROI×fearful>ROI×neutral) in the complex model. Based on the results by Roseman et al. (2018) showing post-treatment effects for fearful and happy faces (vs rest) and fearful>neutral faces (see above), we expected differences in ROI FC for emotional (fearful, happy) faces in comparison to rest and neutral faces. Hence, the ROI×fearful and, ROI×happy versus rest, and ROI×fearful>ROI×neutral contrasts were of particular interest in this secondary analysis. We decided to also look at the ROI×neutral (vs rest) contrast, for completeness of results, as previous work has found similar activation patterns for different facial expressions (Roseman et al. (2018) and in order to disentangle which emotional expressions were driving the PPI effect.
Higher-level analyses across sessions, comparing subjects before any intervention and after the high dose of psilocybin, were performed with mixed-effects GLM (FMRIB’s Local Analysis of Mixed Effects [FLAME-1]) models. A statistical threshold of z>2.3 (cluster-corrected for multiple comparisons with p<0.05) was used for all whole-brain analyses. Additionally, for more direct testing of our hypothesis, we added the complementary ROIs as a pre-threshold mask (i.e. a bilateral amygdala mask for the vmPFC analyses, and a vmPFC mask for the analyses with amygdala as ROI).
Significant clusters were interpreted and labelled based on the Harvard-Oxford cortical and subcortical structural atlases provided in FSL (https://fsl.fmrib.ox.ac.uk/fsl/fslwiki/Atlases).
Relationship with amygdala BOLD response
Subsequently (post-hoc), response amplitudes (beta-weights) from the fearful PPI (vmPFC×fearful) contrast showing a significant decrease in vmPFC-amygdala FC (see Results) were correlated with results from the respective task analysis contrast (fearful vs rest), that had shown increased right amygdala reactivity post-treatment (see Roseman et al., 2018). This allowed for a more specific test of the hypothesised decreased prefrontal inhibitory input to the amygdala as being involved in its increased reactivity after treatment. In line with our primary hypothesis, we expected to find correlations between responses from the PPI contrast (representing the degree of FC between the ROI and identified cluster during fearful face processing) and basic amygdala BOLD responses to fearful faces (i.e. results from the task analysis) as well as their difference scores respectively.
Relationship with clinical outcomes
In order to analyse the relationship of the FC results (PPI analyses) with clinical outcomes, the average parameter estimates (i.e. beta weights from the GLM) of significant clusters identified in the two simple PPI models (representing the degree of FC of the ROIs with the respective clusters during face processing) were extracted for the baseline and post-treatment scan respectively. We focused our analysis on the results from the simple PPI models, as it constitutes the more robust and powerful model, significant clusters were very similar across the different interaction terms of the complex PPI model and in order to reduce the number of comparisons. The analyses in relation to clinical outcomes were carried out using the statistical software program IBM SPSS Statistics for Windows (Version 24.0, Armonk, New York, USA).
Depression
In order to reduce the number of tests and avoid excessive correction of alpha levels, we limited the primary analyses to one clinical outcome of interest: the self-rated Beck Depression Inventory (BDI; Beck et al., 1988) completed at baseline and one week after therapy. The QIDS-SR16 (Rush et al., 2003) was administered at more time-points (at baseline, one day, one week, two weeks, three weeks, five weeks, three months and six months after treatment). For better comparison with results by Roseman et al. (2018), relations between brain outcomes and the QIDS-SR16 were also investigated as supplementary analyses. An overview of correlations with BDI at three-month and all QIDS-SR16 time-points is provided in the Supplementary Material.
The BDI is a reliable and valid 21-item self-rating questionnaire for the assessment of depressive symptoms (Beck et al., 1988). Participants rate their mood and behaviour (e.g. sadness, feelings of guilt, disappointment, crying) according to four severity ratings per item (0–3). The total score determines the severity of depression, differentiating between absent (0–9), mild (10–18), moderate (19–29) and severe (⩾30) depression. Remission is defined as a score ⩽9 (Beck et al., 1988), and treatment response as a 50% drop in BDI score from baseline (Riedel et al., 2010).
The QIDS-SR16 is a 16-item self-report inventory assessing the severity of depressive symptoms in reference to the past seven days (Rush et al., 2003). Symptoms cover the nine Diagnostic and Statistical Manual of Mental Disorders (DSM)-IV symptom domains of major depression, namely: sad mood, concentration, self-criticism, suicidal ideation, interest, energy/fatigue, sleep disturbances, decrease/increase in weight and/or appetite, and psychomotor agitation or retardation. The total score determines the severity of depression, differentiating between absent (0–5), mild (6–10), moderate (11–15), severe (16–20) and very severe (21–27) forms. In line with the BDI classification, treatment response was defined as a 50% drop in score from baseline.
Rumination
As a secondary hypothesis, the relationship between brain outcomes and ruminative thinking and behaviour at one week was examined. During the trial, rumination was measured with the RRS at baseline, one week after and at three-months follow-up (Treynor et al., 2003). The RRS is a 22-item self-report scale assessing ruminative tendencies on a scale from almost never (=1) to almost always (=4), by asking to report what responders ‘generally do’ (e.g. ‘think about how alone you feel’ or ‘analyse your personality and try to understand why you are depressed’). The scale score is determined by summing all 22 items, with a higher score reflecting more ruminative thinking and behaviour. Relation to RRS was analysed at one week, although relation to RRS at three months was also examined for explorative purposes (see Supplementary Material).
Anxiety
While not being an explicit hypothesis of the current study and despite Roseman et al. (2018) not finding a significant relation between amygdala changes and anxiety, previous literature emphasises the 5-HT system (Gordon and Hen, 2004) as well as connectivity between the vmPFC and limbic regions, including the amygdala, in the aetiology of anxious states and anxiety disorders (Engel et al., 2009; Robinson et al., 2016). Levels of anxiety were measured through (a) in-scanner ‘state’ ratings of anxiety during both scanning sessions; and (b) the State-Trait Anxiety Inventory (STAI) (Spielberger et al., 1983) at baseline and one-week post-treatment. The STAI is a widely-used 40-item self-report inventory based on a four-point Likert scale (‘almost never’ to ‘almost always’), assessing anxiety as a state (e.g. ‘I am tense’) and trait (e.g. ‘I worry too much over something that really doesn’t matter’) characteristic. A higher score is reflective of higher anxiety levels. In terms of psychometric properties, internal consistency coefficients ranged from 0.86–0.95, test-retest reliability from 0.65–0.75 (Spielberger et al., 1983).
Statistical analysis
Changes in depression (BDI) and anxiety (STAI) levels post-treatment have already been analysed and reported by Carhart-Harris et al. (2018a; 2016a). Changes in rumination (RRS) from baseline, to one week and three months post-treatment were analysed with repeated measures analysis of variance (RM ANOVA). In order to analyse the relationship to the primary clinical outcome, BDI scores at one week and their change from baseline were correlated with the absolute average parameter estimates (beta-weights) of significant clusters and their difference scores (∆beta=after–before) respectively. The relationship to the secondary outcomes of interest (RRS at one week; STAI at one week; anxiety in-scanner rating) were investigated in the same way. Additionally, independent samples t-tests were conducted, analysing whether those patients who responded to treatment (50% reduction in BDI score) and/or met the criterion for remission (BDI⩽9) differed in terms of average beta-values post-treatment and their change scores (∆beta) per cluster.
We opted to correct for multiple comparisons for each cluster and construct separately, in order to reduce the risk of type two errors due to an overly conservative correction and because our tested clinical constructs were highly related (raw score, change score, remission and response). Accordingly, for the BDI at one week, six comparisons were made per identified cluster, leading to a Bonferroni-corrected α of 0.05/6=0.008. For the secondary outcome, RRS at one week, two comparisons were made per cluster, leading to a Bonferroni-corrected α of 0.025. Anxiety levels (in-scanner ratings and STAI at one-week) were tested at a Bonferroni-corrected α of 0.05/4=0.013. For transparency, an overview of all conducted correlation analyses and t-tests is provided in the tables in the Results section on ‘Relationship with clinical outcomes’.
Prior to all statistical tests, the clinical data were investigated in regard to statistical assumptions. Besides some deviation from normality in BDI scores, at one week in particular, all clinical data met the assumptions for parametric testing.
Results
PPI: amygdala
Results of the primary (simple) PPI model (with faces as the psychological, and amygdala time-series as physiological regressor; Table 1 and Figure 1(a)), revealed increased FC between the amygdala and visual areas, namely the intracalcarine and supracalcarine cortex, cuneus, precuneus and right lateral occipital cortex during face processing (as compared with rest) after psilocybin treatment (amygdala×faces).
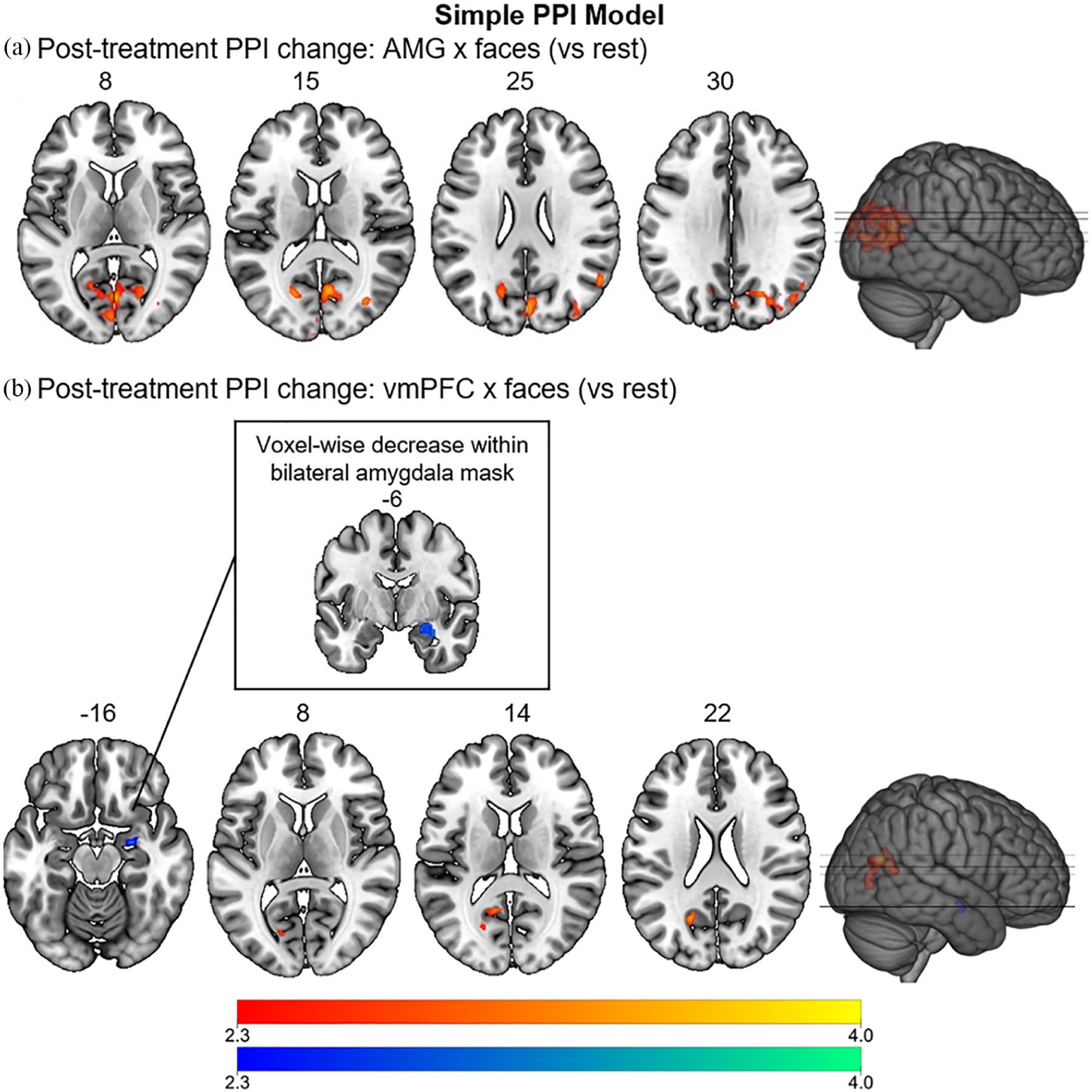
Results of the psychophysiological interaction (PPI) whole-brain and pre-thresholded analyses comparing functional connectivity (FC) of the amygdala (AMG) (a) and the ventromedial prefrontal cortex (vmPFC) (c) during face processing versus rest (simple model) before and after treatment with psilocybin. This analysis constituted the primary analysis of the study. Regions with increased connectivity (after>before) are depicted in red, decreased connectivity (before>after) in blue (cluster-corrected, p<0.05, z>2.3).
Results of the secondary analysis (complex) PPI model (Table 2; Supplementary Material Figure 3) showed stronger FC between the amygdala and similar visual areas during happy (amygdala×happy; zmax=3.96, p<0.001) and neutral (amygdala×neutral; zmax=3.31, p<0.001) faces, but not fearful faces when compared with rest after treatment with psilocybin. For both happy and neutral faces, the significant cluster was centred around the right intracalcarine and supracalcarine cortex. Contrasts between the different interaction terms, i.e. comparing amygdala connectivity between fearful and neutral faces (amygdala×fearful>amygdala×neutral), did not reveal any significant differences. Using the vmPFC as a pre-threshold mask did not produce any significant results for either of the models.
Summary of results of the seed-to-voxel psychophysiological interaction analysis comparing functional connectivity (FC) when processing different facial expressions (happy, fearful, neutral) versus rest (complex model) for both regions of interest (ROIs), the bilateral amygdala (AMG) and ventromedial prefrontal cortex (vmPFC).
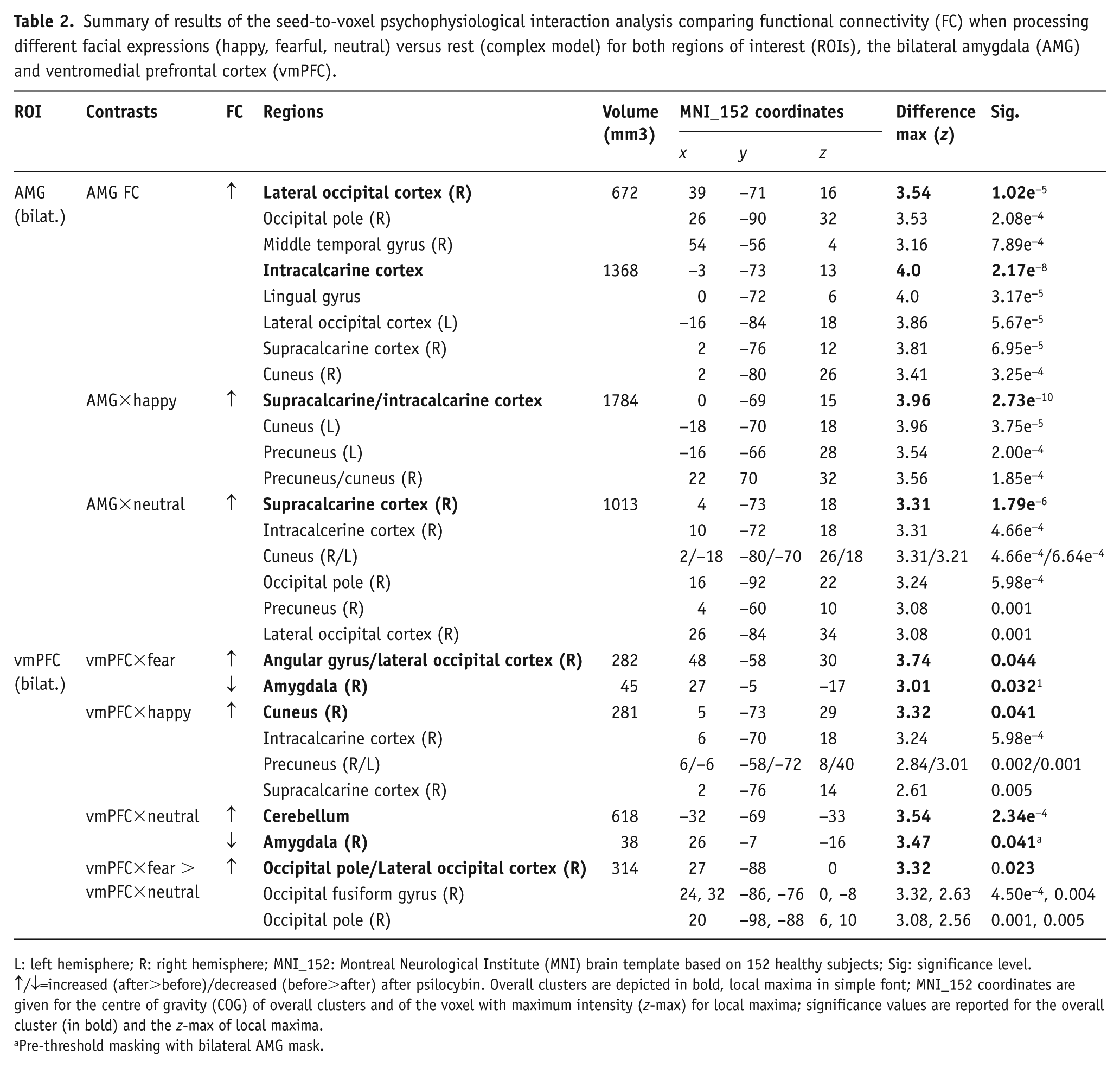
L: left hemisphere; R: right hemisphere; MNI_152: Montreal Neurological Institute (MNI) brain template based on 152 healthy subjects; Sig: significance level.
↑/↓=increased (after>before)/decreased (before>after) after psilocybin. Overall clusters are depicted in bold, local maxima in simple font; MNI_152 coordinates are given for the centre of gravity (COG) of overall clusters and of the voxel with maximum intensity (z-max) for local maxima; significance values are reported for the overall cluster (in bold) and the z-max of local maxima.
Pre-threshold masking with bilateral AMG mask.
PPI: vmPFC
Similar to the amygdala, whole-brain analysis of the primary (simple) PPI model (Table 1, Figure 1(b)), with the vmPFC time-series as physiological and faces as psychological regressor, showed significantly greater FC between the vmPFC and areas in the left occipital and parietal lobes (supracalcarine and intracalcarine cortex, precuneus and lingual gyrus) during face processing (vs rest) post-treatment versus pre-treatment (vmPFC×faces). When adding a bilateral amygdala mask for pre-threshold masking, the interaction term revealed significantly higher connectivity between the vmPFC and right amygdala during face processing before psilocybin treatment as compared to after; thus, supporting our primary hypothesis.
As secondary analysis, when taking the different facial expressions into account (complex PPI model, Table 2 and Figure 2), the vmPFC showed increased FC with the right angular gyrus and lateral occipital cortex during fearful faces (vmPFC×fearful; zmax=3.74, p=0.044), with occipito-parietal visual areas centred around the cuneal cortex during happy faces (vmPFC×happy; zmax=3.32, p=0.040) and with the left cerebellum during neutral faces (vmPFC×neutral; zmax=3.54, p<0.001) after psilocybin treatment as compared with before. Contrasting the interaction term for neutral with fearful faces (vmPFC×fearful>vmPFC×neutral) revealed significantly larger vmPFC FC with visual areas in the right hemisphere (lateral occipital cortex, occipital pole and occipital-fusiform gyrus) during processing of fearful faces (zmax=3.32, p=0.023) after treatment with psilocybin. In line with results from the primary analysis of the simple PPI model, when the bilateral amygdala was added as a pre-threshold mask, significantly higher FC between the vmPFC and right amygdala before treatment with psilocybin during processing of fearful (vmPFC×fearful; zmax=3.01, p=0.032) and neutral (vmPFC×neutral; zmax=3.47, p=0.041), but not happy (vmPFC×happy) faces was found.
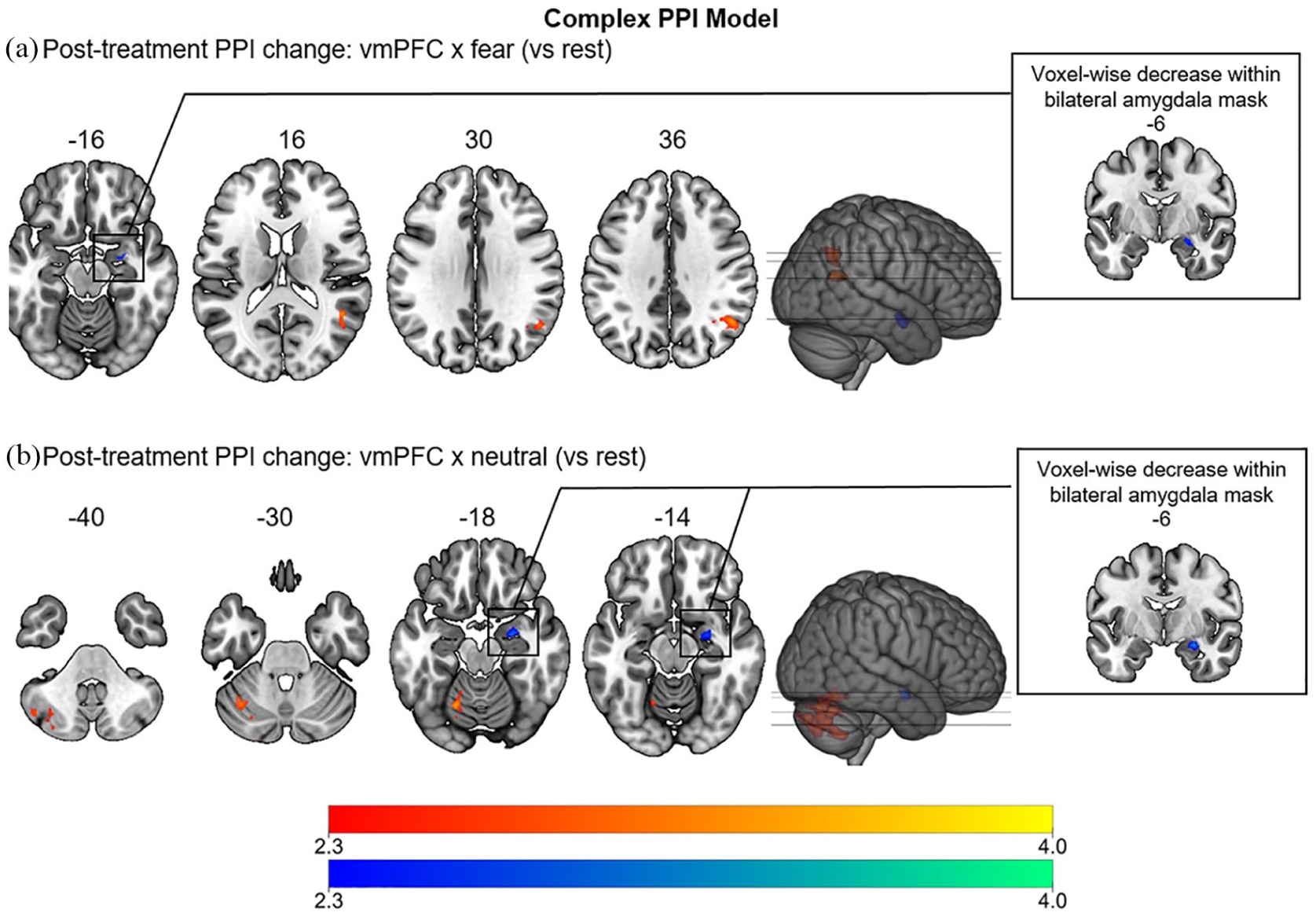
Results of the psychophysiological interaction (PPI) whole-brain and pre-thresholded analysis comparing ventromedial prefrontal cortex (vmPFC) functional connectivity when processing different facial expressions (a=fearful, b=neutral) with rest (complex model) before and after treatment with psilocybin. The contrast with happy faces (vmPFC×happy) did not reveal any significant results. This analysis was conducted as a secondary analysis to provide a more comprehensive exploration of the different conditions in the experiment. Regions with increased connectivity (after>before) are depicted in red, decreased connectivity (before>after) in blue (cluster-corrected, p<0.05, z>2.3).
Relation with amygdala response to fearful faces (task-analysis)
Response amplitudes (beta-weights) from the PPI fearful contrast (vmPFC×fearful) were correlated with amygdala responses to fearful faces in the task analysis (see Roseman et al., 2018). Contrary to expectations, the PPI contrast after treatment did not correlate with the task analysis (rfearful=0.19, p=0.425, R2=0.04); neither did the difference scores (rfearful=−0.12, p=0.629, R2=0.014). However, analyses did reveal a significant negative correlation between the PPI fearful contrast and the respective task contrast before treatment (rfearful=−0.48, p=0.040, R2=0.23), indicating that higher vmPFC-right amygdala FC was linked to a lower amygdala response to fearful faces before treatment with psilocybin.
Relationship with clinical outcomes
The primary clinical outcome of interest was the relationship between post-treatment FC changes and changes in BDI scores at the one-week endpoint. Important secondary outcomes of interest were the relation to rumination and anxiety scores at one week, plus in-scanner anxiety scores. The relevant results are shown in Tables 3 and 4.
Relation of amygdala and ventromedial prefrontal cortex (vmPFC) functional connectivity (FC) to clinical outcomes (Beck Depression Inventory (BDI), Ruminative Response Scale (RRS)) one-week post-treatment.
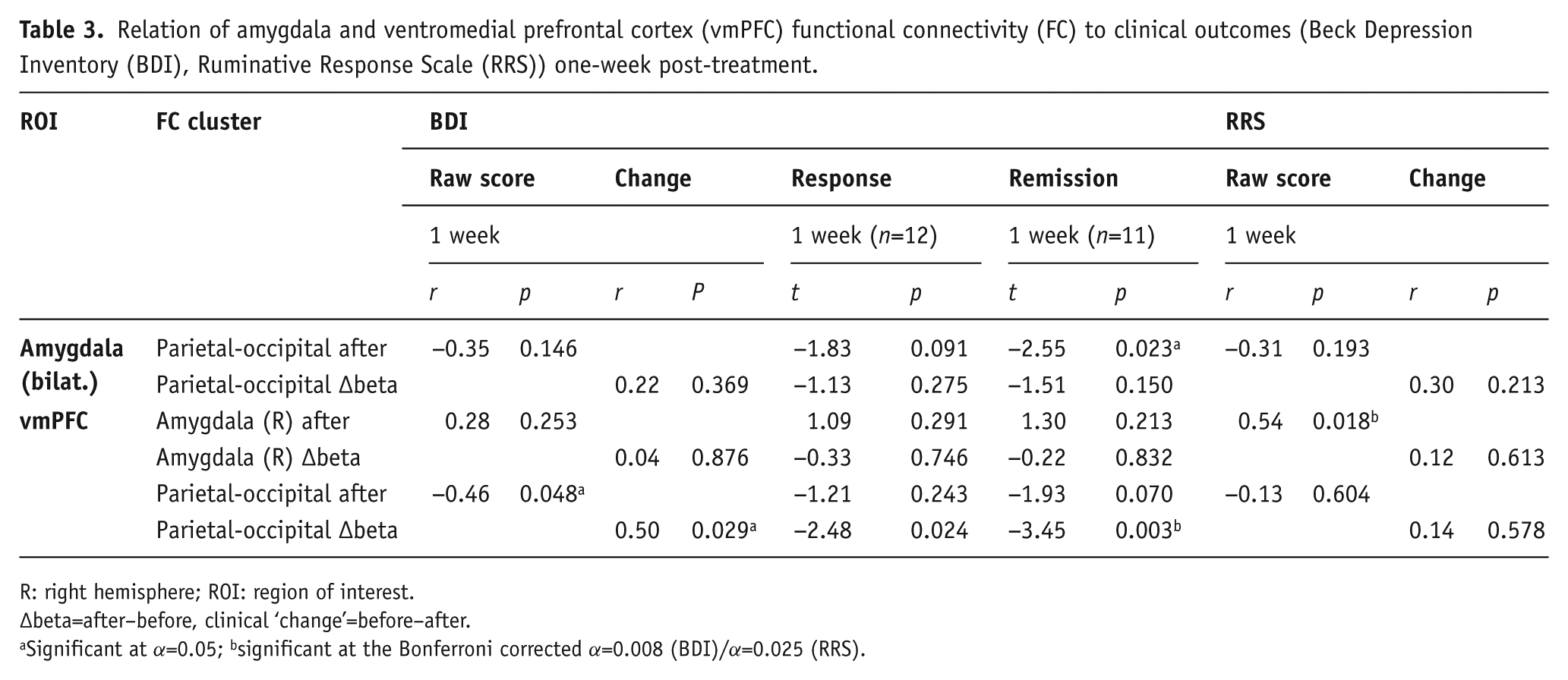
R: right hemisphere; ROI: region of interest.
∆beta=after–before, clinical ‘change’=before–after.
Significant at α=0.05; bsignificant at the Bonferroni corrected α=0.008 (BDI)/α=0.025 (RRS).
Relation of amygdala and ventromedial prefrontal cortex (vmPFC) functional connectivity (FC) to anxiety levels (in-scanner ratings one day, State-Trait Anxiety Inventory (STAI) one week) post-treatment.
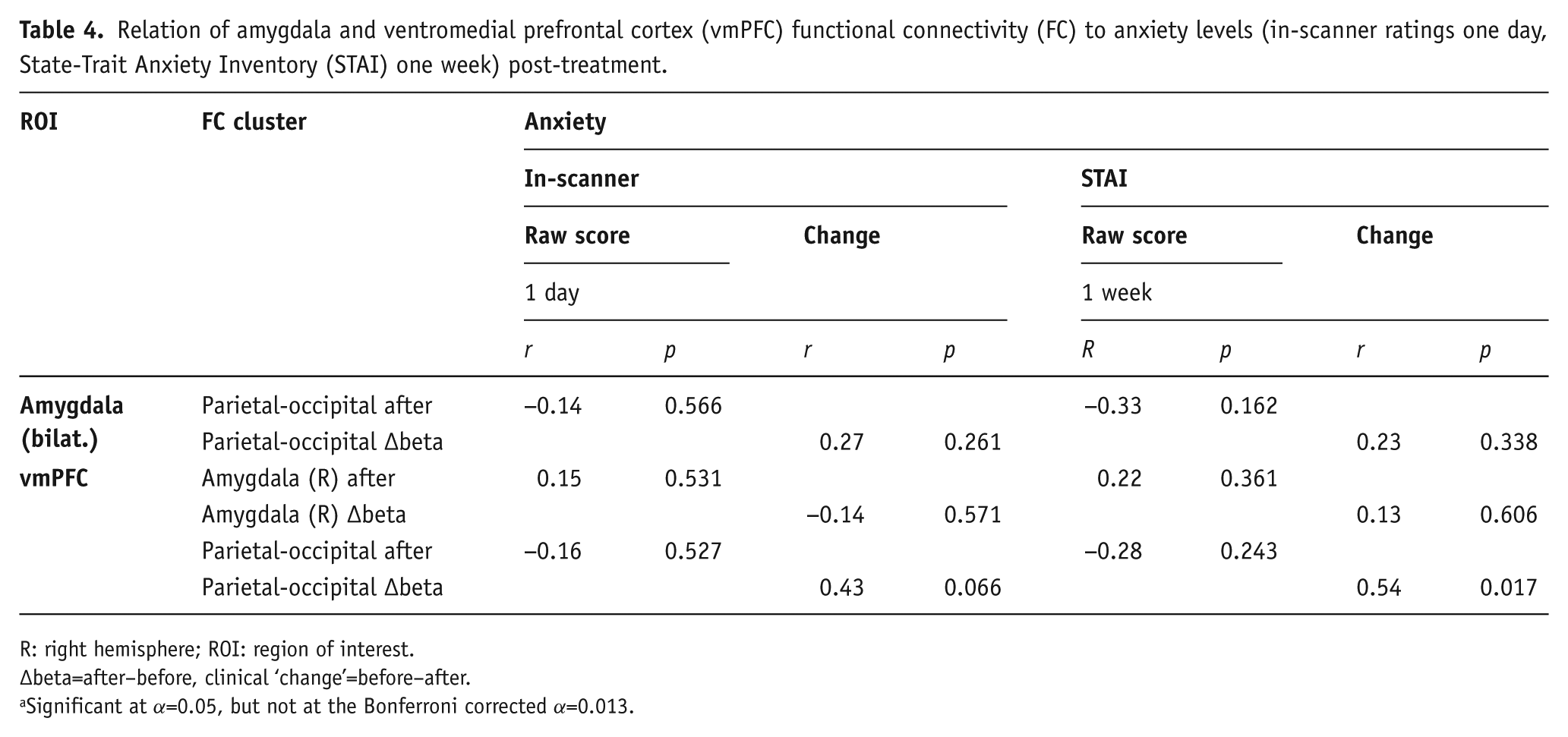
R: right hemisphere; ROI: region of interest.
∆beta=after–before, clinical ‘change’=before–after.
Significant at α=0.05, but not at the Bonferroni corrected α=0.013.
Clinical outcomes
As already reported previously (Carhart-Harris et al., 2018a; 2016a), BDI scores were significantly reduced at one week (mean reduction=22.26, SD=11.37, p<0.001); 63.2% of patients showed a treatment response (50% drop in BDI score) at this time-point, with 57.9% meeting criteria for remission (BDI⩽9). As evident from the RM ANOVA, rumination levels (RRS) differed significantly between time-points, F(2,36)=17.98, p<0.001, ηp2=0.50. Pairwise comparisons revealed a significant mean difference between baseline and one week (mean reduction=21.58, p<0.001, 95% confidence interval (CI) 14.3–28.83) as well as three months post-treatment (mean reduction=15.90, p<0.001, 95% CI 8.66–23.13), while there was no significant difference between the drop in scores at one week and three months (mean reduction=−5.68, p=0.197, 95% CI −14.6–3.23). Within-subjects contrasts further suggested a quadratic trend, F(1,18)=15.51, p=0.001, ηp2=0.463, indicating that rumination levels decreased from baseline to one week and then rose at three months, despite remaining significantly lower than at baseline.
Amygdala FC
With Bonferroni correction, none of the clinical outcome measures (BDI, RRS, STAI, in-scanner anxiety levels) was significantly correlated with amygdala FC after treatment or its change (∆beta) from baseline (Tables 3 and 4).
vmPFC FC
Right amygdala-vmPFC FC post-treatment was not significantly related to depression levels at one week (r=0.28, p=0.253), and neither did the change in right amygdala-vmPFC FC relate to change in BDI (r=0.04, p=0.876) or rumination from baseline (r=0.12, p=0.613). Absolute FC between the vmPFC and right amygdala after treatment did, however, significantly correlate with rumination scores one-week post-treatment (r=0.54, p=0.018), with lower connectivity being linked to lower rumination levels (see Figure 3c). This result remained significant with a Bonferroni corrected α of 0.025.
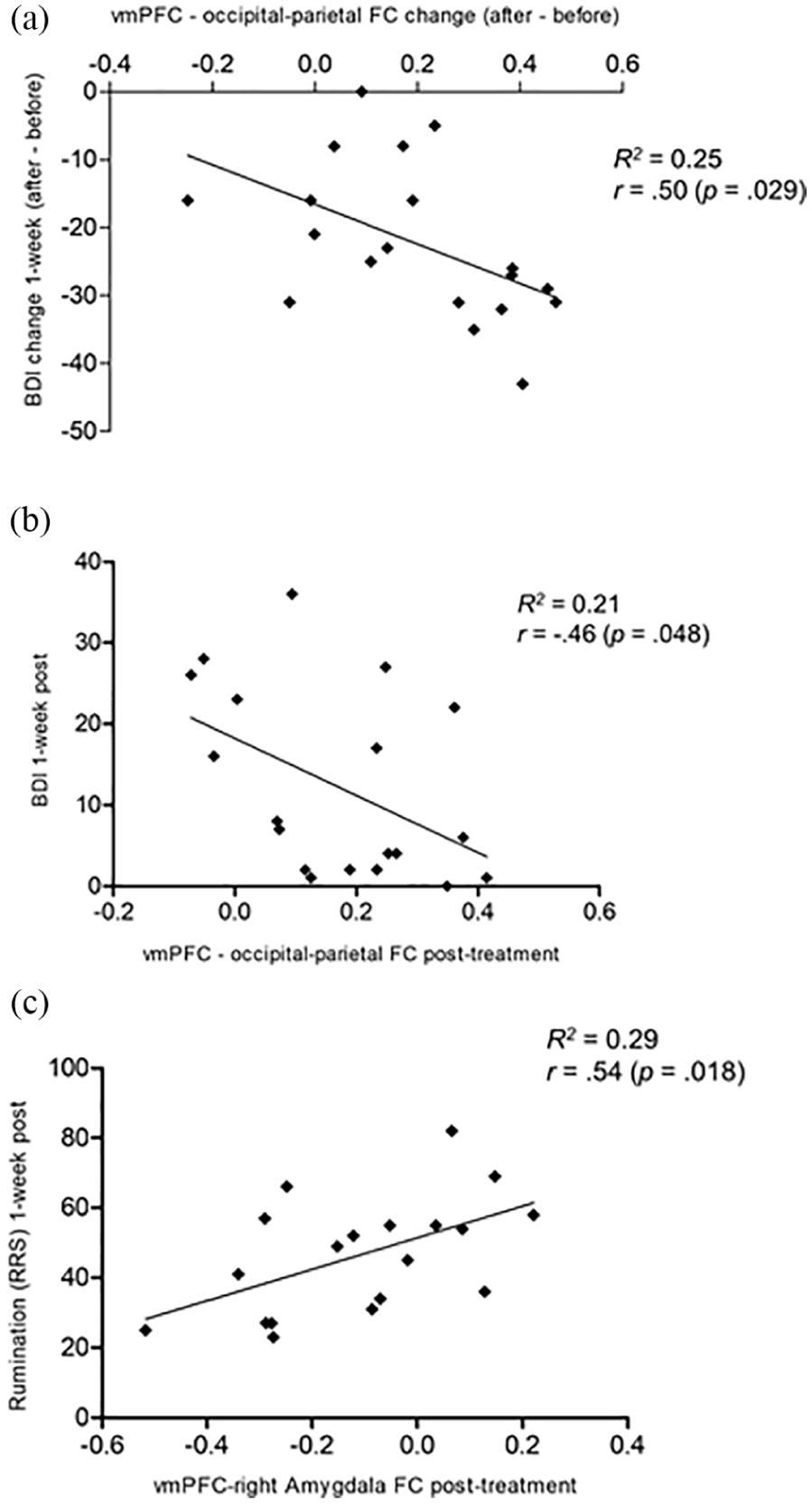
Ventromedial prefrontal cortex (vmPFC) functional connectivity (FC) during face processing (simple model) post-treatment plotted against clinical outcomes: (a) and (b) Beck Depression Inventory (BDI) and (c) Ruminative Response Scale (RRS) at one week.
FC between the vmPFC and the occipital-parietal cluster after treatment was significantly correlated with BDI scores at one week (r=−0.46, p=0.048; Figure 3b), and so were the respective change scores (r=0.50, p=0.029 ; Figure 3a). In line with this, responders (t(17)=−2.48, p=0.024) and remitters (t(17)=−3.45, p=0.003) at one week differed significantly in terms of vmPFC-occipital-parietal FC change, and the result for remitters remained significant after Bonferroni correction. Additionally, the vmPFC-occipital-parietal FC change was significantly correlated with change in STAI at one week (r=0.54, p=0.017), indicating that a larger increase in connectivity was associated with a greater decrease in anxiety from baseline. However, this result did not quite hold under a Bonferroni corrected α of 0.013. No significant correlations to anxiety levels rated during post-treatment scanning or its change from baseline were found.
Discussion
The current analyses constitute an extension to previous work that revealed increased amygdala responsiveness post treatment with psilocybin for TRD (Roseman et al., 2018). Here we aimed to investigate changes in FC of the amygdala and vmPFC during the same face-processing paradigm performed at baseline and one-day after treatment with psilocybin for TRD. Based on previous results of altered FC under and after psychedelics (Carhart-Harris et al., 2017; Grimm et al., 2018; Roseman et al., 2014) and the findings of Roseman et al. (2018) showing increased responsivity of the right amygdala to fearful and happy faces after treatment with psilocybin in patients with TRD, we hypothesised post-treatment changes in FC between the amygdala and the vmPFC, decreased amygdala-vmPFC FC in particular, during face processing. It was further hypothesised that the proposed FC changes of the amygdala and vmPFC would relate to improvements in depression (primary clinical outcome) at one-week, and anxiety and rumination scores one-week post-treatment, plus in-scanner anxiety (secondary clinical outcomes).
vmPFC-right amygdala FC
Our findings provide some support for the key hypothesis of altered amygdala-PFC FC after treatment with psilocybin. Consistent with the prior hypothesis, we observed a reduction in vmPFC FC to the right amygdala during face processing after treatment with psilocybin. Further analysis showed that this effect was driven by the viewing of fearful and neutral faces in particular. One plausible, although somewhat speculative, physiological mechanism which would be consistent with this effect is decreased inhibitory input from the vmPFC to the right amygdala during emotional face-viewing post-treatment with psilocybin resulting in a disinhibitory influence on amygdala responsiveness during the same paradigm.
We previously found that increased amygdala responsiveness to fearful (versus neutral) faces was related to improved clinical outcomes at one day, one week, and three weeks post-treatment (Roseman et al. 2018), but the reduction in vmPFC-right amygdala FC during face processing observed in the present analysis was not significantly associated with depression severity at one week. However, vmPFC-right amygdala FC post-treatment was significantly related to levels of rumination at one week, with lower connectivity being associated with less rumination. Additionally, rumination levels were significantly decreased at one week and three months post-treatment with psilocybin. Despite the non-significant relation to BDI scores, considering that rumination is a strong risk factor for the development and maintenance of depressive episodes (Cooney et al., 2010), decreased amygdala-prefrontal connectivity after psilocybin may still be an important underlying mechanism for the observed symptom improvement. This is consistent with work by Berman et al. (2011) who found that greater RSFC within the DMN, including the vmPFC, correlated with rumination scores in major depression. Failure to disengage DMN regions may be an important component of the mechanics of depression (Berman et al., 2011; Hamilton et al., 2015; Hayley and Litteljohn, 2013). At the same time – and in contrast to research on the aetiology of anxiety, fear conditioning and PTSD (Engel et al., 2009; Koenigs and Grafman, 2009; Robinson et al., 2016; Stevens et al., 2013) – decreased prefrontal-amygdala FC was not associated with levels of anxiety during scanning or one week after treatment. Similarly, Roseman et al. (2018) found no association between amygdala response and levels of anxiety.
Taken together, while certainly preliminary, these results challenge the general view of amygdala responsivity/hyperactivity and decreased amygdala-prefrontal connectivity as being indicative of psychopathology per se. While quite consistently described as a biomarker and risk factor in the literature on states of emotional dysregulation, such as depression, the current results, together with those of Roseman et al. (2018), suggest a more nuanced interpretation. One potential explanation is that conventional antidepressants and psilocybin have fundamentally different actions on emotional processing (Carhart-Harris and Nutt, 2017), with conventional antidepressants down-regulating emotional responsiveness (Goodwin et al., 2017) and psychedelics allowing patients to more fully engage with emotions (Carhart-Harris et al., 2018b; Roseman et al., 2018). One might connect these findings to patient reports of increased emotional acceptance and decreased emotional avoidance after psilocybin therapy, which contrasts with the emotionally-muting effects patients described with their past antidepressant use (Watts et al., 2017). Increased amygdala responsiveness, accompanied by decreased engagement of prefrontal control regions, as suggested by the current findings, might be an important underlying mechanism involved in the emotional reconnection reported with successful psychedelic therapy (Watts et al., 2017).
Caution is needed not to draw firm conclusions here, however. While, as predicted, we did observe decreased vmPFC-right amygdala FC during the processing of fearful and neutral faces, the degree of this connectivity change did not significantly correlate with the degree of increased amygdala responsiveness after treatment identified in the task analysis and neither do we have information about the direction of information flow from the present analyses.
Further changes in FC
Additionally, the amygdala and vmPFC showed increased FC with occipital and parietal cortices during face processing post-treatment with psilocybin. More precisely, the vmPFC showed significantly higher FC with areas in the visual cortex during face processing, and this was linked to change in depression and anxiety scores at one week as well as criteria for response and remission of depression. Increased visual cortex activity (reported by Roseman et al., 2018) and connectivity to the vmPFC and amygdala (found in the present analyses) could be the result of increased task engagement and changes in emotional stimulus recognition and processing post-treatment, e.g. perhaps patients were simply attending to the faces more after treatment (Harmer et al., 2009a; Harmer et al., 2009b; Jonassen et al., 2015) and thus engaging relevant circuitry. The results may also be reflective of increased engagement of amygdala-visual cortex circuitry post-treatment, a circuitry that is known to be involved in the normal detection and evaluation of emotionally salient stimuli (Morris et al., 1998; Vuilleumier and Pourtois, 2007). Increased connectivity to the precuneus is particularly intriguing as this region and the surrounding posterior cingulate is not just involved in visual-spatial navigation, but also a component of the DMN, thought to be involved in self-consciousness, reflective self-awareness, episodic memory and imagination (Cavanna and Trimble, 2006; Hassabis et al., 2007; Utevsky et al., 2014).
Importantly, considering our previous results (Roseman et al., 2018) and the often described hypersensitivity of the amygdala to negative stimuli in particular in depression (Ma, 2015), it is quite notable and worth mentioning that the secondary (complex) analysis did not reveal a significant difference when contrasting amygdala FC during fearful versus neutral faces (amygdala×fearful>amygdala×neutral). Accordingly, it must be emphasised that only some of the given results substantiate the made predictions for the analysis of the secondary (complex) PPI model.
Limitations
Focusing on design limitations, the given study was an open-label feasibility study; therefore, there was no blinding or placebo condition, and the sample size was relatively small. Small sample sizes generate low statistical power and increase the chances of inferential errors (Button et al., 2013). PPI models generally suffer from lack of power due to multicollinearity between regressors (O’Reilly et al., 2012: https://fsl.fmrib.ox.ac.uk/fsl/fslwiki/PPICaveats). This makes the use of larger samples particularly important. In this regard, it is also crucial to keep in mind that some of the analyses on relation to clinical outcome did not survive the applied Bonferroni correction for multiple testing, which weakens the respective results. In this respect, it must also be considered that in fact two separate PPI analyses – a primary (simple) model and a secondary (complex) model – were conducted with two ROIs, the amygdala and the vmPFC, adding to the problem of multiple comparisons. Reasons for analysing both PPI models are given above, but the number of comparisons have to be kept in mind as an important limitation. A potential solution and option for future studies to partly resolve this, would be the use of non-parametric testing.
Additionally, the sample comprised a very specific patient group: TRD. This limits the generalisability of the results and introduces bias in favour of alternative antidepressant treatments (e.g. psychedelic drugs), since multiple standard treatments had been unsuccessful in the patient sample. In order to rule out that these results are sample and/or TRD-specific, future studies should explore psilocybin as a treatment for non-treatment-resistant major depression in a larger sample, using a placebo-controlled design. However, it must be acknowledged that the inclusion of an effective placebo condition is very difficult when studying psychedelic drugs. This is due to the fact that the subjective effects of a high-dose psychedelic experience are so noticeable that they break the blinding. Alternative strategies include the use of a low dose of the psychedelic as a control condition or to compare the psychedelic against an active pharmacological control, such as stimulants (e.g. amphetamines or methylphenidate), a sedative (e.g. a benzodiazepine) or a selective serotonin reuptake inhibitor (SSRI). Furthermore, based on results from the present study, it is not possible to disentangle the drug-specific effects from those of the psychological support provided during the treatment. Adding a psilocybin without psychological support condition would resolve this issue, although it is questionable whether it is ethical to administer a high dose of psilocybin to a vulnerable population without psychological support, as it would imply risks for the patients and could certainly hinder clinical improvement, if not worsen outcomes (Carhart-Harris et al., 2018d; Studerus et al., 2012). In this respect, it is important to consider that the provided psychological support and in-session ‘check-ins’ in the present study were kept to a minimum (i.e. only one preparation and one integration session, plus acute and peri-acute support, but no specific psychotherapeutic interventions), in order to minimise their potential effect.
Additionally, while patients were requested to down-titrate their antidepressant medication over a minimum wash-out period of two weeks, it must be acknowledged that (a) patient 2 continued taking venlafaxine (a 5-HT-norepinephrine reuptake inhibitor) throughout both dosing and scanning sessions and was not excluded from the present analyses, and (b) in general a wash-out period of two weeks might be insufficient and could potentially bias the clinical and neurobiological results. Both factors constitute a limitation and should be ruled out in future studies.
FC analyses provide information on the temporal correlation between activity in spatially separate regions of the brain; however, they do not provide information on directionality and causal interactions between brain regions. Given our prior hypothesis, that reduced top-down control from the vmPFC to the amygdala might explain the increased responsiveness of the amygdala to emotional faces seen previously (Roseman et al., 2018), the next logical step would be to explore this question using a metric capable of providing information about causal information flow. Future work is now required to test this hypothesis more directly, not just by improving our imaging analyses, e.g. incorporating dynamic causal modelling (DCM; Frassle et al., 2017) and perhaps using other complementary imaging modalities (e.g. magnetoencephalography (MEG)) and supplementary emotional processing paradigms, but also by improving our indices of emotional re-engagement and acceptance/avoidance (although see Stroud et al., 2018). Using the Dynamic Emotional Recognition Task, Stroud et al. (2018) provided evidence for improved emotional processing to levels of healthy controls in TRD patients after treatment with psilocybin. Modulating techniques, such as repetitive transcranial magnetic stimulation (rTMS) to temporarily disrupt vmPFC function could also be applied. With the present paradigm, regressing out missed trials and/or including reaction times as a proxy of attention as a regressor in the GLM could be used to ensure that measured effects are not solely due to attentional changes or confounded by differences in engagement with, and more specifically, looking at, the relevant task stimuli (Harmer et al., 2004; Jonassen et al., 2015). Alternatively, for the same purpose, eye-tracking could be employed in future studies.
Conclusion
Psilocybin constitutes a novel and promising treatment strategy for TRD and perhaps other psychiatric disorders. The results of the current PPI analyses revealed greater FC between the vmPFC and, independently, the bilateral amygdala, and occipital-parietal cortices during face processing after treatment with psilocybin in TRD, with the vmPFC FC increases correlating with improvements in depression symptomatology. Consistent with our prior hypothesis, we also observed a decrease in FC between the vmPFC and right amygdala during face processing post-treatment with psilocybin, which was related to rumination levels at one week. These results provide further information on the mechanisms underlying the treatment effect of psilocybin and may be relevant to patients’ reports of an increased ability and willingness to engage with their emotions post-psilocybin therapy. Thus, we predict that the previously observed increase in amygdala responsiveness (Roseman et al., 2018), combined with a decrease in FC to prefrontal control regions, as cautiously implied here, may relate to an increase in emotional sensitivity and acceptance post-treatment with psilocybin (Watts et al., 2017). The proposed therapeutic mechanisms of the psilocybin treatment model are quite different to those hypothesised to underlie the action of classical antidepressants. However, there are also some similarities, most notably in relation to increased plasticity and the importance of context to the effectiveness of each treatment (Carhart-Harris and Nutt, 2017; Carhart-Harris et al., 2018c). Due to the methodological limitations of the design and analysis, the drawn conclusions must be regarded as preliminary. Future studies and more powerful and robust analyses are now required to examine the reliability of these inferences.
Supplemental Material
Supplementary_Material_Final – Supplemental material for Therapeutic mechanisms of psilocybin: Changes in amygdala and prefrontal functional connectivity during emotional processing after psilocybin for treatment-resistant depression
Supplemental material, Supplementary_Material_Final for Therapeutic mechanisms of psilocybin: Changes in amygdala and prefrontal functional connectivity during emotional processing after psilocybin for treatment-resistant depression by Lea J Mertens, Matthew B Wall, Leor Roseman, Lysia Demetriou, David J Nutt and Robin L Carhart-Harris in Journal of Psychopharmacology
Footnotes
Acknowledgements
The infrastructure support for this trial was provided by the NIHR Imperial Clinical Research Facility and NIHR Imperial Biomedical Research Centre.
Authors’ Note
Lea J Mertens is now affiliated to Department of Molecular Neuroimaging, Central Institute of Mental Health (CIMH), Heidelberg University, Mannheim, Germany.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This trial was funded by a UK Medical Research Council Grant and the Alex Mosley Charitable Trust. Trial Registration: ISRCTN number: ISRCTN14426797. EudraCT number: 2013-003196-35.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.