
Abstract
Background:
Antidepressant drugs in adolescent depression are sometimes mired by efficacy issues and paradoxical effects. Transcranial direct current stimulation (tDCS) could represent an alternative.
Aims/methods:
We tested the antidepressant action of prefrontal tDCS and paroxetine (20 mg/kg, intraperitoneal) in olfactory bulbectomised (OBX) adolescent rats. Using enzyme-linked immunosorbent assays and in situ hybridisation, we examined treatment-induced changes in plasma brain-derived neurotrophic factor (BDNF) and brain serotonin transporter (SERT) and 5-HT-1A mRNA.
Results:
OBX-induced anhedonia-like reductions in sucrose preference (SP) correlated with open field (OF) hyperactivity. These were accompanied by decreased zif268 mRNA in the piriform/amygdalopiriform transition area, and increased zif268 mRNA in the hypothalamus. Acute paroxetine (2 days) led to a profound SP reduction, an effect blocked by combined tDCS–paroxetine administration. Chronic (14 days) tDCS attenuated hyperlocomotion and its combination with paroxetine blocked OBX-induced SP reduction. Correlations among BDNF, SP and hyperlocomotion scores were altered by OBX but were normalised by tDCS–paroxetine co-treatment. In the brain, paroxetine increased zif268 mRNA in the hippocampal CA1 subregion and decreased it in the claustrum. This effect was blocked by tDCS co-administration, which also increased zif268 in CA2. tDCS-paroxetine co-treatment had variable effects on 5-HT1A receptors and SERT mRNA. 5-HT1A receptor changes were found exclusively within depression-related parahippocampal/hippocampal subregions, and SERT changes within fear/defensive response-modulating brainstem circuits.
Conclusion:
These findings point towards potential synergistic efficacies of tDCS and paroxetine in the OBX model of adolescent depression via mechanisms associated with altered expression of BDNF, 5-HT1A, SERT and zif268 in discrete corticolimbic areas.
Keywords
Introduction
Major depressive disorder is debilitating and underlies many other psychiatric disorders. It represents a leading cause of burden and disability (Ferrari et al., 2013). With a 12-month prevalence rate of 11.3% in adolescents and 9.6% in young adults it has been suggested that the rate of depression is increasing, especially among young people. These trends suggest that depression commonly develops in adolescence and can be persistent (Mojtabai et al., 2016). Nonetheless, it is likely that pathophysiological manifestations vary across age groups, mirroring divergences in symptom profiles (Rice et al., 2019).
Treatment response to conventional antidepressants, e.g., serotonin (5-hydroxytryptamine, 5-HT) reuptake inhibitors, is also dissimilar. Their efficacy relies on a number of synaptic and neuroplastic mechanisms, e.g., enhancement of transmitter availability via inhibition of transporter proteins for 5-HT (SERT) and other monoamines, desensitisation of somatodendritic auto-inhibitory 5-HT1A receptors and sensitisation of postsynaptic ones, and upregulation of growth factors, e.g., the brain-derived neurotrophic factor (BDNF) in key limbic compartments needed for repair and restabilisation (Bambico and Belzung, 2013). Among adolescents, these medications show little therapeutic effect, and may even worsen depression-related symptoms, e.g., anxiety and suicidality risk (Goldsmith and Moncrieff, 2011; Vitiello and Ordóñez, 2016). Recommended non-pharmacological therapies, e.g., cognitive-behavioural therapy (CBT), also show mixed results and often produce little benefit over active controls (for review, see Oar et al., 2017). The Treatment for Adolescents with Depression Study (TADS), the most extensive placebo-controlled study to date, showed that 5-HT reuptake inhibitors in combination with CBT yielded significantly greater amelioration of symptoms compared to CBT or placebo when given to severely depressed adolescents (Keller et al., 2001). On the other hand, other studies (e.g., the recent revaluation study of paroxetine) have shown that 5-HT reuptake inhibitors (paroxetine) are ineffective in adolescent depression and linked to suicidal ideation (Le Noury et al., 2015). Paradoxical behavioural effects are also apparent in rodent models, oftentimes marked by opposite patterns of neuroplastic molecular changes (Gomez et al., 2015; Homberg et al., 2011).
The precise biological underpinnings of unfavourable response outcomes in paediatric patients are highly debated but likely pertain to age-dependent developmental changes in key limbic areas. First, adolescents exhibit increased frontal activity (Mayberg, 1997) that may be attributed to incomplete neural maturation processes (Arain et al., 2013; Tao et al., 2012). Second, there is profound disparity in the prefrontal regulation of subcortical areas, which predisposes for increased emotional volatility and vulnerability in adolescents (Kerestes et al., 2014). Third, an underdeveloped expression of monoaminergic system constituents, e.g., transporters and receptors, can contribute to poor therapeutic response to 5-HT-modulating and noradrenergic antidepressants (Bowman and Daws, 2019; Bylund and Reed, 2007; Mulder et al., 2003).
Transcranial direct current stimulation (tDCS) is a novel, non-invasive neuromodulation strategy that sends weak electrical current through the scalp to induce synaptic and excitability changes (Nitsche et al., 2012; Pelletier and Cicchetti, 2015). Neuroplasticity-related changes in molecular and epigenetic correlates of the antidepressant response have also been consistently reported with an anodal-type configuration directed over the dorsomedial prefrontal cortex (Fritsch et al., 2010; Kim et al., 2017; Podda et al., 2016). In the rodent, this contains the cingulate and the prelimbic areas (Bambico and Belzung, 2013; Paxinos and Watson, 2007). Similar to prefrontal deep brain stimulation (DBS) (Bambico and Belzung, 2013; Mayberg, 1997; Price and Drevets, 2010), tDCS shows some benefits for cognitive performance and psychiatric symptoms, including depression (Alonzo et al., 2013; Boggio et al., 2008; Ferrucci et al., 2009; Mondino et al., 2014; but Palm et al., 2012). tDCS appears to meet safety, feasibility and tolerability standards in children and adolescents (Boggio et al., 2008; Gillick et al., 2015), and shows positive therapeutic outcomes in those with neurodevelopmental disorders (Amatachaya et al., 2014; Lee et al., 2019; Palm et al., 2016; Soff et al., 2017).
There is hitherto a paucity of experimental studies investigating the antidepressant properties of tDCS. And it has yet to be tested on paediatric and adolescent depression. Because previous studies with anodal tDCS, mostly applied to the left dorsolateral prefrontal cortex, showed positive outcomes on hypofrontality-related clinical conditions (Amatachaya et al., 2014; Koenigs and Grafman, 2009; Soff et al., 2017), and prefrontal DBS has antidepressant properties (Bambico et al., 2015; Hamani et al., 2010; Mayberg, et al., 2005), we elected to test the effect of prefrontal anodal tDCS. Brunoni et al. (2013) found that a combination of tDCS and the 5-HT reuptake inhibitor, sertraline, produced more potent antidepressant effects than either treatment alone. Therefore, we also examined whether concurrent tDCS and 5-HT reuptake inhibitor treatment results in potential synergistic effects. Next, we tested whether plasma BDNF that has been previously used as a peripheral marker for therapeutic response to some, but not all, antidepressant classes including somatic electrical interventions will accordingly be modified by the treatments. To induce the depressive-like phenotype, we employed the olfactory bulbectomy (OBX) procedure, which is well-validated for examining the progression of depression-related phenotypes and the time course of the antidepressant response to treatment. The two main outcomes we measured were sucrose preference (SP) attenuation – which mimics anhedonia, a core symptom of depression – and OF hyperactivity (Cryan and Mombereau, 2004; Ramaker and Dulawa, 2017; Song and Leonard, 2005). To achieve these goals, we developed a medial prefrontal tDCS model to appraise treatment effects across behavioural, peripheral and central/brain indicators.
Materials and methods
Animals
Adolescent male Sprague-Dawley rats (Charles-River Saint-Constant, Quebec, Canada), at postnatal (PND)20-21, were placed under standard conditions upon arrival. Animals remained pair-housed until electrode placement surgery at PND35-37. All animals were given ad libitum access to food and water and were kept on a 12-h light–dark cycle under standard laboratory conditions (lights on at 07:00).
All procedures and protocols for experiments and animal housing followed the guidelines of the Canadian Council on Animal Care and Memorial University of Newfoundland’s Animal Care Committee. For all surgical procedures, animals were anaesthetised with isoflurane (induction at 2.5%, maintenance at 2%) and mounted on a standard stereotaxic apparatus (David Kopf Instruments, CA, USA) in the skull-flat position. All brain coordinates were from Paxinos and Watson (2007).
Treatment groups
The study timeline is shown in Figure 1(a). After a week of acclimatisation, each animal was assigned to one of five groups. All animals, except for the control group (CTR), underwent OBX (PND 28-29). After 4-5 days (PND33-34), OF locomotor activity and SP scores were assessed before tDCS electrode implantation (PND35-37). Animals were then given acute treatments on PND44 and PND45. The 5-HT reuptake inhibitor paroxetine (a kind gift from Dr Gabriella Gobbi, McGill University; Sigma-Aldrich) was dissolved in physiological saline (0.9% NaCl) to obtain an injection volume of 1 mL/kg and was intraperitoneally administered at 20 mg/kg (after Amodeo et al., 2015; Erdemir et al., 2014). Each animal received one of the following combinations: paroxetine and tDCS (OBX+Pax+tDCS), vehicle and tDCS (OBX+Veh+tDCS), paroxetine and sham stimulation (OBX+Pax+Sham) or vehicle and sham stimulation (OBX+Veh+Sham). The CTR group received the vehicle and sham stimulation (CTR+Veh+Sham). An OF test (OFT) was conducted on PND45 and SP test (SPT) on PND46, after the second acute treatments and before any additional treatments. Then, the same animals then continued to receive chronic once-daily treatments before behavioural assessment on PND57 (OFT) and PND58 (SPT). Euthanasia and brain extraction were done on PND59.
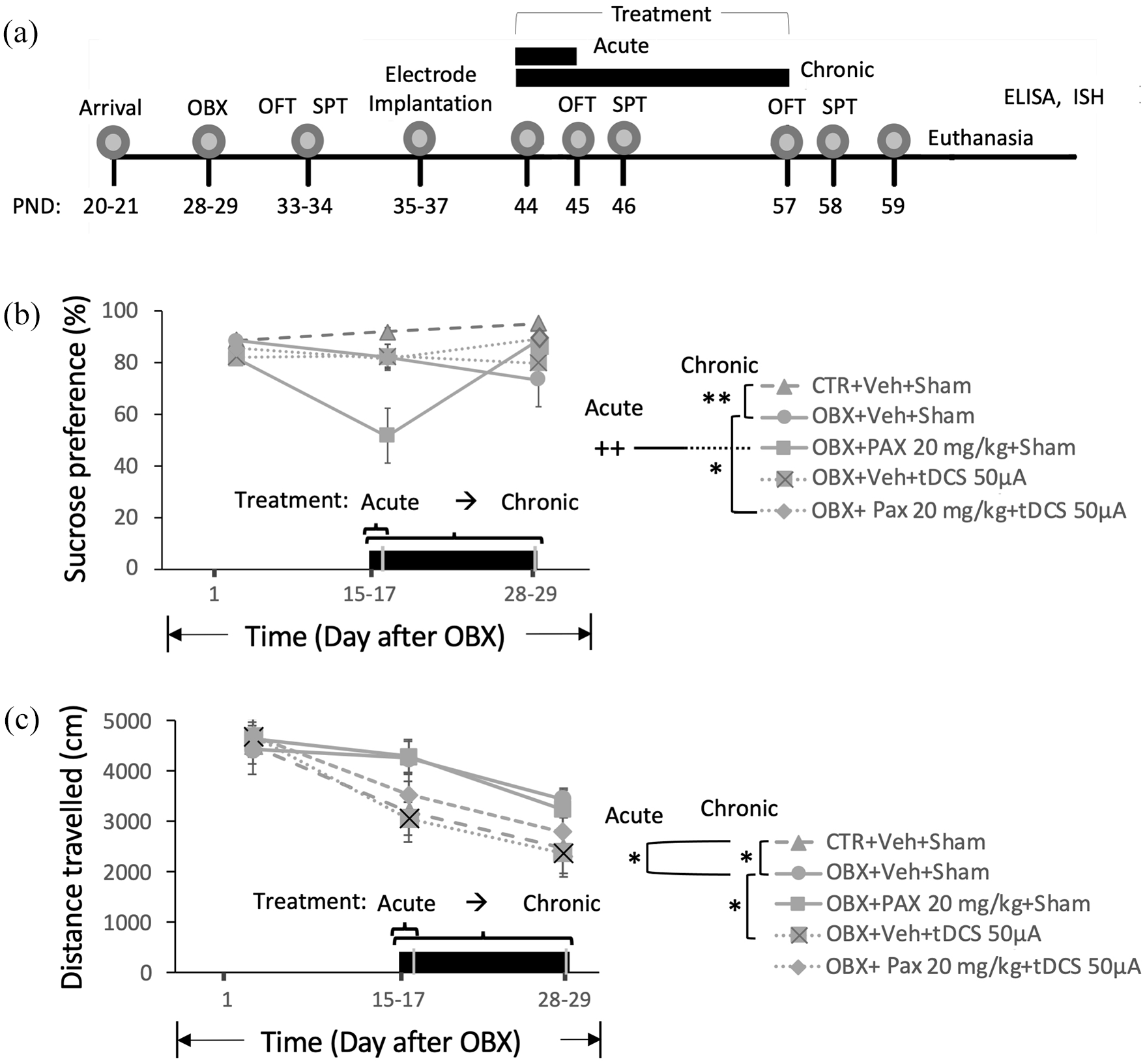
(a) Timeline of procedures. (b–c) SPT and OFT: progression of sucrose preference (SP) scores (b), and locomotor activity (LA) scores (c) following OBX and in response to acute (2 days) treatment tested 16–17 days post-OBX, and to chronic (14 days) treatment tested 28–29 days post-OBX. Black bars at the bottom of graphs denote duration of treatment. Data points are expressed as mean values±standard error of mean (SEM); n = 9−19. Rightmost legend panel, comparison of treatment effects as indicated (acute and chronic): *p < 0.05, **p < 0.01, ++p < 0.01 vs. rest of groups.
OBX surgery
We used a modified version of the OBX procedure described elsewhere (Kelly et al., 1997). A cranial window was drilled over the anterodorsal cranial cavities containing both olfactory bulbs (AP: +6.0 to +9.0 mm, ML: ±2.0 mm), stretching for 4.5 mm from the cribriform plate. The plate border is located 3.5 mm from the nasofrontal suture and 9 mm anterior to bregma. A sterilised 23G needle/syringe connected to a suction pump was then used to bilaterally aspirate the visible olfactory bulbs. Aspiration was carried out under a stereomicroscope. To prevent regrowth, sterile gel foam beads were placed within the vacated cavity against the cribriform plate. CTR animals underwent a similar procedure, with the dura being punctured but bulbs left intact. Following surgery, the animals were allowed to recover for 2 weeks before the commencement of treatment in order to allow for the development of OBX-induced neuroplastic changes and circuit rewiring within the extended olfactory–limbic network (for review see Song and Leonard, 2005).
Electrode placement
Small holes were drilled on both sides of the skull (AP: +2.0 mm, ML: ±2.0 mm) and far behind the cerebellum (AP: −15.0 mm, ML: +1.0 mm) and jeweller screws were tightly twisted into place to act as an anchor for the dental acrylic resin. A male JST connector was placed on the skull such that a conducting metal plate (2.5 ± 0.25 × 1.5 ± 0.25 mm) attached to the anodal end laid over the medial prefrontal cortex (mPFC) (AP: +2.2 mm to +4.7 mm). The cathodal end was attached to the post-cerebellar screw to act as the reference/ground electrode. Dental acrylic resin (Jet Set-4™ Denture Repair Powder and Jet™ Liquid, Lang Dental, USA) was applied to keep the assembly in place on the skull during treatment. Animals were left to recover for 7 days before the beginning of treatment.
Behavioural test
Behavioural tests were conducted beginning 5 hours following acute treatment and chronic treatment. Multiple correlational and covariability analyses were later performed for all behavioural data as well as for plasma BDNF (see section Blood sampling and BDNF analysis).
Sucrose Preference Test (SPT)
The SPT was conducted as an indicator of anhedonia (Papp et al., 1991). A two-bottle, overnight SPT was given to all animals, with one bottle containing a 1% sucrose solution and one bottle containing water. The animals had free access to both bottles for 16 hours. They were neither food-deprived nor water-deprived. Sucrose and water levels were measured before and after the test, and SP was determined by dividing the amount of sucrose consumed by the total amount of liquid consumed.
Open Field Test (OFT)
The OFT for LA was conducted using hyperlocomotion as a putative measure of hyperactivity and psychomotor agitation as proposed by others (Song and Leonard, 2005; Zueger et al., 2005). Rats were placed facing the corner in an OFT chamber (60 cm × 60 cm × 60 cm) with aluminium foil covering the walls and two 13-watt lights placed overhead for illumination, as described in previous experiments (Kelly et al., 1997). The animals were allowed to freely explore the chamber during their 10-min trial. Trials were counterbalanced based on group, and the chamber was cleaned with 70% ethanol in between trials. Each trial was videotaped and the distance travelled was analysed offline using Ethovision XT14 software (Noldus Information Technology, USA).
Effects of acute treatment
ON PND44, an injection of paroxetine was immediately followed by tDCS administration. A second treatment session occurred on PND45, 12–16 hours following the first session. CTR animals were injected with saline at 1 mL/kg. Rats were then connected to a cable assembly via a JST connector and subjected to 15 min of 50 μA anodal tDCS (13.33 A/m2), delivered from an external current generator (custom-built by Ed Yee according to the specifications by Drs José Nobrega and Francis Bambico, Centre for Addiction and Mental Health, Toronto). These parameters were based on preliminary intensity-response data and on behaviourally active properties used in rodent models. The generated charge density is also below the threshold (20 A/m2) shown to produce lesions (Jackson et al., 2016; Lee et al., 2019; Mishima et al., 2019). Each animal was mildly restrained while being connected to the generator but was promptly placed back in its home cage for the duration of the stimulation. Sessions were monitored using a multimeter attached to the generator to ensure that each animal received the full extent of the stimulation. Animals that lost electrode contact (<50 μA multimeter reading) were excluded from further tests. Rats receiving sham stimulation were handled in a similar manner, but no current was delivered. The OFT was conducted beginning 5 hours following treatment on PND45. The SPT was conducted for 16 hours on PND46.
Effects of chronic treatment
Procedures were similar to those under section Effects of acute treatment. Following the initial two sessions on PND44-45, treatment was administered to the same rats every 24 hours for 12 more days, resulting in a total of 14 consecutive daily sessions. The same behavioural tests were conducted upon completion of treatment on PND57-58.
Blood sampling and BDNF analysis
Following euthanasia by CO2 on PND60, trunk blood was collected in Microvette CB 300 K2E tubes (Sarstedt, Germany). Samples were immediately centrifuged for 10 min at 3000 rpm to obtain plasma and stored at −20 ℃ until analysis. BDNF plasma levels were measured in 96-well plates using a commercially available ELISA kit (Promega, WI, USA) according to the manufacturer’s instructions. Plates were coated with an anti-BDNF mAB + carbonate coating buffer overnight at 4°C. Nonspecific binding was blocked, followed by a wash with TBST buffer. A standard curve was prepared by performing six serial dilutions of the BDNF standard. The remaining plates were filled with plasma samples assayed in duplicates. Plates underwent a series of incubations and washes for the anti-human BDNF pAb, anti-IgY horseradish peroxidase conjugate and TMB One solution. The reaction was stopped with 1N hydrochloric acid. To determine the concentration of BDNF (pg/mL, a microplate reader (Molecular Devices, CA, USA) was used within 30 min of stopping the reaction, with absorbance for each plate set at 450 nm. Peripheral BDNF measures can reliably be used to make inferences on central BDNF activity as a principal substrate for neuroplasticity. Indeed, previous studies have found positive correlations between serum and cortical levels of BDNF in both rats and pigs (Karege et al., 2002; Klein et al., 2011).
Messenger RNA measurements for zinc finger transcription factor (zif268), 5-HT1A receptor and 5-HT transporter (SERT)
As a central indicator of brain regional activation, we performed in situ hybridisation (ISH) with [35S]UTP-labelled riboprobes complementary to sequences for zif268, 5-HT1A receptor and SERT mRNA as previously described (Bambico et al., 2019; Volle et al., 2018). Zif268 or early growth response protein 1 is an immediate early gene product that functions as a transcriptional regulator coupled to cell electrical activity and neuroplasticity. The 5-HT1A receptor is located either presynaptically or postsynaptically. Its desensitisation in somatodendritic domains of raphe 5-HT neurons and its sensitisation and tonic activation in the hippocampal CA3 subfield and in other postsynaptic sites have been linked with the mechanism of therapeutic action of antidepressants (Bambico and Belzung, 2013; Blier and de Montigny, 1994). The SERT protein is implicated in depression as some antidepressants, e.g., 5-HT reuptake inhibitors, block the activity of SERT or diminish its function (Bambico and Belzung, 2013; Blier and de Montigny, 1994). After euthanasia by CO2, the brains were extracted and unfixed coronal sections, 20 µm in thickness, were prepared on a Leica cryostat, thaw-mounted on Superfrost-plus slides (Fisher) then stored at −80°C until ISH processing. First, slide-mounted sections were fixed in 4% paraformaldehyde for 5 min at room temperature, and rinsed in 10 mM phosphate buffer (pH 7.4) containing 150 mM NaCl (2 min). The slides were then treated with 100 mM triethanolamine-HCl (pH 8.0) for 5 min, acetylated with 0.25% acetic anhydride for 10 min in the same buffer, and rinsed twice for 5 min in 0.1 M sodium citrate (pH 7.0) containing 300 mM NaCl (SSC). Slides were then dehydrated in graded ethanol, defatted in 100% chloroform for 5 min, rehydrated through ethanol and then stored frozen until hybridisation. Using the NCBI BLAST Tool, the designed primer sequences were checked for homology with the rat genome and found to be specific for their respective transcripts, and to include the T7 phage polymerase promoter sequence with the antisense primer. Primers were synthesised by ACGT (Toronto, ON, Canada). The following primers were used: zif260: GenBank # NM_012551, (bases 660–679) 5′-tcacctatactggccgcttc-3’ and (bases 1062–1043) 5′-aggtctccctgttgttgtgg-3′; 5-HT1A receptor: Genbank # NM_012585.1, (bases 331-350) 5’-ctgtttatcgccctggatgt-3’ and (bases 713-694) 5’- gctcccttcttttccacctt; and SERT: Genbank # NM_013034, (bases 94-113) 5’-tactgaccagcagcatggag-3’ and (bases 524-505) 5’-cagtgcgagctccatgtaaa-3’. T7 phage polymerase promoter: taatacgactcactataggg. [35S]UTP-labelled antisense RNA probes complementary to the sequences of interest were generated by in vitro transcription using the Maxiscript (T7) kit (Thermo Fisher Scientific, Waltham, MA, USA). Slides were incubated overnight at 60°C and labelled probes were diluted to 2500 cpm/mL in hybridisation solution containing 50% formamide, 35% Denhardts, 10% dextran sulfate, 50 mM sodium citrate (pH 7.0) containing 300 mM NaCl, fish DNA (300 mg/mL), yeast tRNA (100 mg/mL) and DTT (40 mM). Sections were rinsed in 6 mg/mL RNAse A (Sigma Aldrich, St. Louis, USA) at 45°C for 40 min in decreasing concentrations of SSC followed by 10 s rinses in deionised water and in 70% ethanol, and air dried. Then, the slides were exposed to Kodak BioMax film at 4°C for 2 weeks.
Densitometric analyses were performed with MCID 7.0 software (InterFocus). Standard curves obtained from calibrated radioactivity standards were used to convert raw optical density values to radioactivity levels in microcuries per gram of tissue (μCi/gT). Readings were first averaged across all sampling windows in a section and then across all sections to produce a final density value for each region for each animal.
Statistical analysis
We submitted the data to a one-way or to a two-way mixed design analyses of variance (ANOVA) with type of stimulation (tDCS or sham), type of drug (paroxetine or saline), treatment regimen (baseline, acute or chronic) or brain regions of interest (ROIs) as factors. Duncan’s multiple range test was used for post hoc, pairwise comparisons. Inter-domain Pearson correlation analyses (r) were conducted for behavioural and peripheral BDNF scores. The slope and y-intercepts of the regression lines were subjected to ANOVA. In view of the possibility that during adolescence peripheral BDNF changes were subtle, contingent upon symptom severity or subject to covariation with antidepressant-related behavioural measures in the SPT and OFT (Park et al., 2014), we performed multiple correlational analyses and generated a correlation matrix heatmap. Non-parametric analyses were used when assumptions of normality and homogeneity of variance were not met. A value of p≦0.05 was considered to be significant. Excel 2016 (Microsoft Corporation, WA, USA), Sigma Plot and Jamovi version 0.9 (the Jamovi project, 2019; retrieved from https://www.jamovi.org) were used for data analyses. All data are expressed as mean (M)±SEM.
Results
Combined paroxetine and tDCS treatment attenuated OBX-induced reduction of SP
For the SPT (Figure 1(b)), a two-way ANOVA on SP revealed significant main effects of treatment (F4,58=5.423, p<0.01) and time (F2,94=4.671, p=0.012), and a significant treatment × time interaction (F8,94=4.104, p<0.01) were obtained. Post hoc comparisons showed that at 16–17 days post-OBX, the mean SP score of OBX+Vehicle(Veh)+Sham animals (n=13) only achieved a nonsignificant 8.93% decline from the level of CTR+Veh+Sham animals (n=11). None of the acute treatments (OBX+Veh+tDCS, n=10; OBX+Paroxetine(Pax)+Sham, n=9; and OBX+Pax+tDCS, n=19) significantly raised mean scores from those of OBX+Veh+Sham and CTR+Veh+Sham. Rather, paroxetine (OBX+Pax+Sham) dramatically led to significant SP score reduction (≈ −32%, p<0.01 vs. all other groups); this did not occur when paroxetine was combined with tDCS (OBX+Pax+tDCS, n=19). At 28–29 days post-OBX, the mean score of OBX+Veh+Sham was markedly lower (−19.48%) than that of CTR+Veh+Sham (p<0.01). Neither chronic treatment with paroxetine (OBX+Pax+Sham) or tDCS (OBX+Veh+tDCS) was effective in preventing OBX-induced SP reduction. However, their combination (OBX+Pax+tDCS) restored mean score very close to the CTR+Veh+Sham value.
tDCS attenuated OBX-induced OFT hyperlocomotion
Two-way ANOVA on distance travelled in the OFT (Figure 1(c)) showed a significant main effect of time (F2,102=30.666, p<01) with nonsignificant effects of treatment (F4,62=2.110, p=0.089) and of treatment × time interaction (F8,102=1.375, p=0.217). We proceeded with post hoc tests within each time point, as these were planned comparison that sought to identify which specific means differed (Wei et al., 2012). We found that compared to CTR+Veh+Sham levels (n=16), OBX+Veh+Sham (n=13) displayed significantly greater LA at 16–17 days (+34.86%) and 28–29 days post-OBX (+45.58%) (p≦0.05), as previously shown (Song and Leonard, 2005). OBX+Veh+tDCS (n=10) but not OBX+Pax+Sham (n=9) or OBX+Pax+tDCS (n=19) blocked OBX-induced (OBX+Veh+Sham) hyperlocomotion/hyperactivity following chronic treatment (−36.02%, p=0.05) but also marginally following acute treatment (−24.87%, p=0.06).
Paroxetine and tDCS differently modify relationship among depressive-type behavioural and peripheral indicators
Plasma BDNF concentration (pg/mL) was assayed in OBX (n=7) and CTR (n=10) rats. A between-group difference was initially assessed in order to ascertain whether peripheral BDNF can serve as a valid physiological correlate of depressive-like behaviour in this model. An independent samples t-test revealed a marked reduction of plasma BDNF in OBX animals when compared to CTRs (t14= −2.07, p⩽0.05; Figure 2(a)). SP-BDNF correlational analyses (Figure 2(b)) revealed a significant difference in the regression lines for CTR and OBX, F1,12=23.9, p⩽0.01 for slope and F1,12=46.95, p⩽0.01 for the y-intercept. SP scores and plasma BDNF levels were weakly correlated in CTR, r = −0.22. OBX reversed directionality and enhanced the strength of this correlation >six-fold (r= −0.6), suggesting that variabilities in anhedonia-like behaviour and plasma BDNF are dependently influenced by a common pathophysiological mechanism. This provided the impetus to further probe into treatment effects on BDNF as well as into possible central correlate of these behavioural and peripheral indicators (zif268 mRNA analysis for regional cell activation, and 5-HT-1A and SERT mRNA analyses for functional changes in the 5-HT system-implicated in depression pathophysiology and in the mechanisms of action of antidepressants).
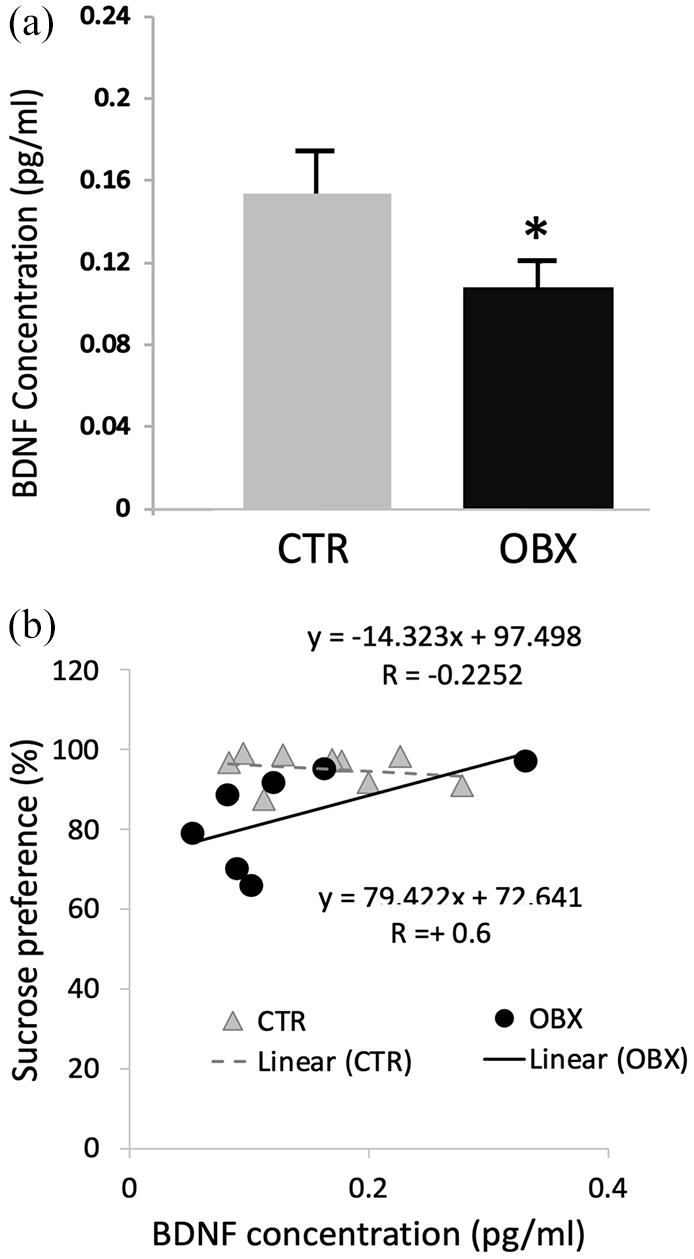
(a) Plasma BDNF concentration in CTR and OBX animals. (b) Regression lines for correlations between BDNF concentration and sucrose preference (SP) scores. Bars in graph represent mean values ± SEM; n = 7−9. *p < 0.05.
Subsequent one-way ANOVA on plasma BDNF for chronic treatment effects failed to detect any significant differences.
We performed multiple correlational analyses and found that in plotting SP against LA (Figure 3(a), top-right), a weak positive correlation was uncovered for CTR+Veh+Sham. This was diametrically divergent from the strong negative correlational profile of the regression line for OBX+Veh+Sham, which was roughly close to OBX+Pax+Sham. Those for OBX+Veh+tDCS and OBX+Pax+tDCS were positive-going, with the highest correlational coefficient garnered by OBX+Pax+tDCS. A similar trend was found when SP was plotted against BDNF (Figure 3(a), bottom-left). The negative correlational/regression profile for OBX+Veh+Sham conformed with OBX+Pax+Sham, and the positive profile of OBX+Veh+tDCS and OBX+Pax+tDCS with CTR+Veh+Sham. In plotting LA with BDNF (Figure 3(a), bottom-right), we uncovered a strong negative correlation for CTR+Veh+Sham that was considerably attenuated in OBX+Veh+Sham and OBX+Pax+Sham. OBX+Veh+tDCS and OBX+Pax+tDCS showed moderate negative and positive correlations, respectively.
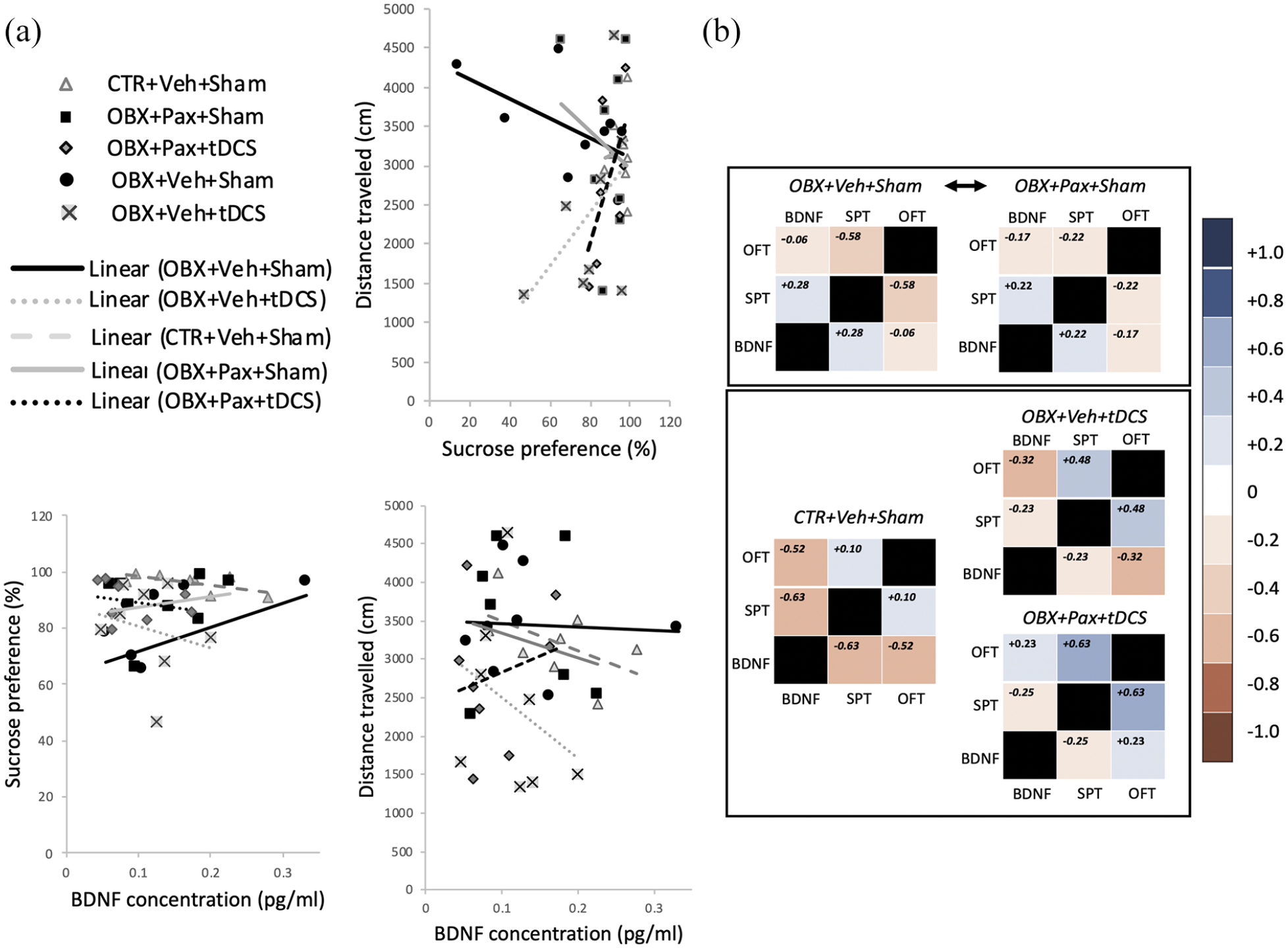
(a) Regression lines for correlations between OFT locomotor activity (LA) scores and sucrose preference (SP) scores (top right), SP and plasma BDNF (bottom left), and LA and plasma BDNF (bottom right). (b) Correlational matrix heat map. Correlational coefficients from Figure 3(a) were matrix-mapped under each treatment group. The spectrum of correlational strengths and directionality are represented as graded colours (right vertical bar).
Generating a matrix representation of all r coefficients (Figure 3(b); n=6–10/group), we found that the heatmap for OBX+Pax+Sham very closely resembled the one for OBX+Veh+Sham, and the OBX+Pax+tDCS and OBX+Veh+tDCS heatmaps were similar to CTR+Veh+Sham. These data indicate that tDCS treatment, compared to paroxetine mono-treatment, more effectively normalised the pattern of relationships among the peripheral and behavioural indicators, vectoring towards characteristics predictive of an antidepressant response.
tDCS and paroxetine modified corticolimbic zif268 mRNA levels
We analysed regional zif268 mRNA levels in 19 brain ROIs (Figures 4 and 5). Figure 4(a) shows a diagrammatic representation of their locations in four brain sections. There was a significant main effect of ROI (F4,652=147.932, p<0.01; n=8–9/group) and treatment × ROI interaction (F72,652=1.693, p<0.01). Post-hoc comparisons revealed that OBX resulted in a drastic attenuation of zi268 in the piriform cortex/amygdalopiriform transition area (CTR+Veh+Sham, n=9 vs. OBX+Veh+Sham, n=8; OBX+Pax+Sham, n=8; OBX+Veh+tDCS, n=8 or OBX+Pax+tDCS, n=8; p<0.01; Figure 4(b)) and an increase in the ventromedial and anterior hypothalamus (CTR+Veh+Sham vs. all others, p<0.01; Figure 5), and these effects were not reversed by any of the treatments. In the hippocampal formation, OBX did not alter zif268 in all ROIs (CTR+Veh+Sham vs. OBX+Veh+Sham). In CA1, paroxetine (OBX+Pax+Sham) led to increased zif268 (vs. CTR+Veh+Sham, p<0.01). This effect was completely blocked by tDCS co-treatment (vs. OBX+Pax+tDCS, p=0.01). tDCS alone (OBX+Veh+tDCS) also showed a moderate increase (vs. CTR+Veh+Sham, p<0.05). In CA2, tDCS mono-treatment (OBX+Veh+tDCS) or tDCS-paroxetine co-treatment (OBX+Pax+tDCS) increased zif268 (OBX+Veh+tDCS vs. all others except OBX+Pax+tDCS, p≦0.01; OBX+Pax+tDCS vs. all others except OBX+Veh+tDCS, p≦0.05). In the claustrum, paroxetine (OBX+Pax+Sham) showed a lower-than-normal zif268 (vs. CTR+Veh+Sham, p<0.05). No significant effects were obtained from the CTR groups (see Supplementary Figures S3–S4).
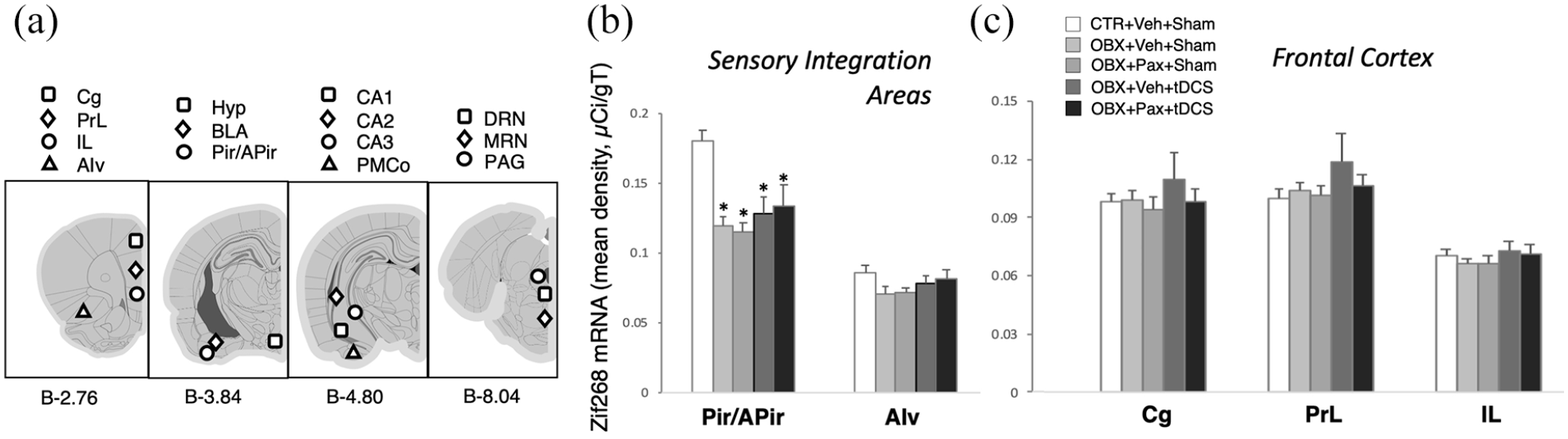
(a) Coronal brain section plates from Paxinos and Watson (2007) are presented with approximate locations of regions of interest (ROI) (millimetre posterior to bregma [b] as shown at the bottom) for zif268 in situ hybridisation densitometric analyses. Cg: cingulate; PrL: prelimbic; IL: infralimbic; AIv: ventral anterior insula; Hyp: ventromedial and anterior hypothalamus; BLA: basolateral amygdala; Pir/APir: piriform cortex and amygdalopiriform transition area; CA1–3: cornu ammonis subfields 1 to 3 of the hippocampus; PMCo: posteromedial cortical amygdala; DRN/MRN: dorsal and median raphe nuclei; PAG: periaqueductal grey. (b-c) Mean optical density (in micro-Curie per gram of tissue) across different ROIs in sensory integration areas (b) and frontocortical targets (c) of tDCS. Bars in graphs represent mean values ± SEM; n = 8−9. *p < 0.05.
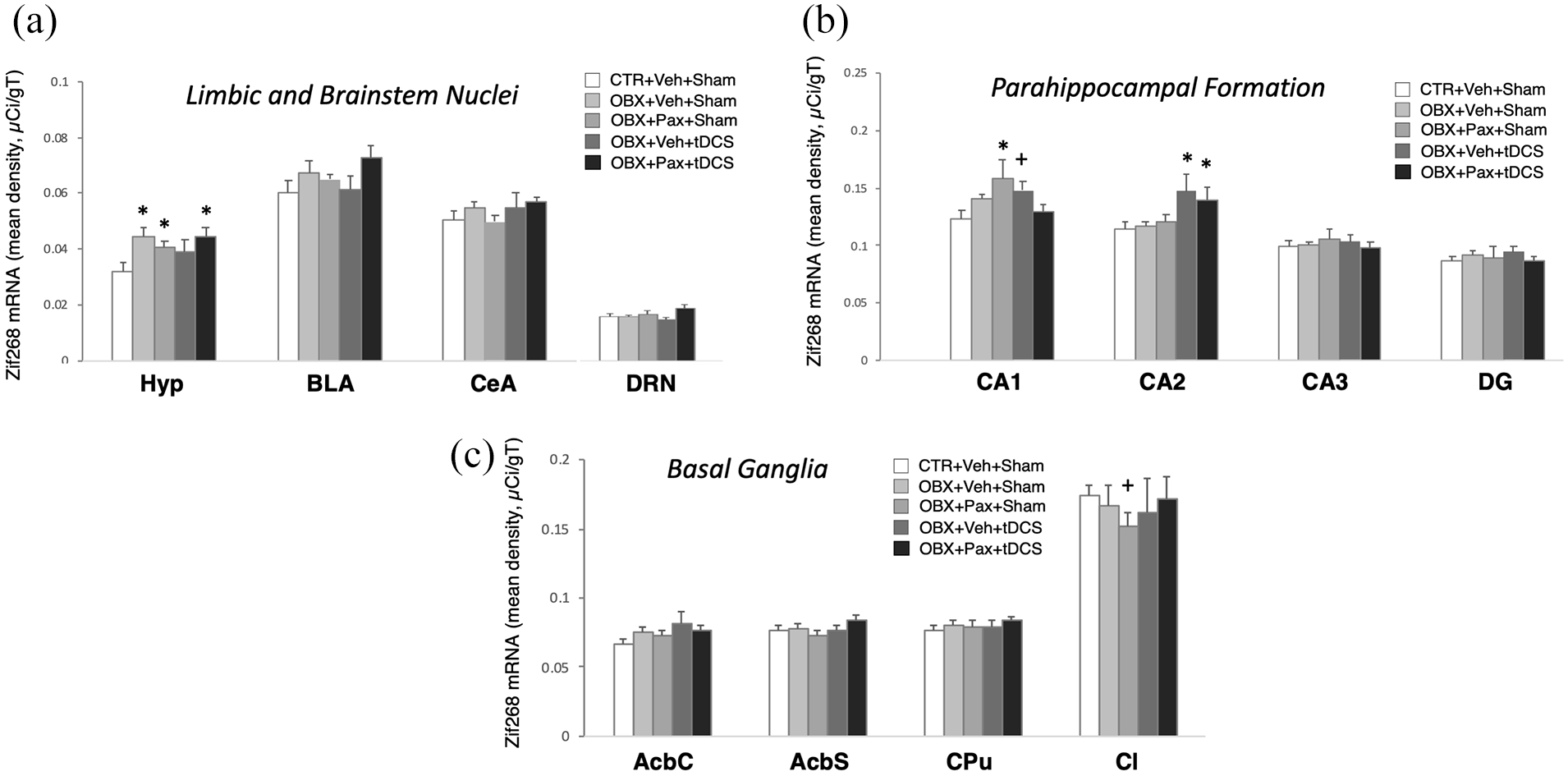
In situ hybridisation for zif268. Mean optical density (in micro-Curie per gram of tissue) across ROIs in limbic and brainstem areas (a), hippocampal formation (b) and basal ganglia (c). CeA: central amygdala; DG: dentate gyrus; AcbC: nucleus accumbens core; AcbS: nucleus accumbens shell; CPu: caudate-putamen; Cl: claustrum. Bars in graphs represent mean values ± SEM; n = 8−9. +p < 0.05, *p < 0.01.
tDCS and paroxetine modified 5-HT1A receptor mRNA levels in parahippocampal/hippocampal subregions
Two-way ANOVA on 5-HT1A receptor mRNA (Figure 6) uncovered a significant main effect of treatment (F4,750=5.753, p<0.001; n=7–10/group) and of ROI (F22,750=206.874, p<0.001; n=23), and a significant interaction (F88,750=1.279, p=0.05). Post hoc comparisons revealed pairwise differences exclusively in parahippocampal and hippocampal subregions. In CA3, OBX+Pax+Sham (n=7) and OBX+Pax+tDCS (n=10), but not OBX+Veh+tDCS (n=7), showed significantly greater 5-HT1A receptor mRNA levels than CTR+Veh+Sham (n=10) or OBX+Veh+Sham (n=10) (p≦0.01). In the entorhinal cortex, OBX+Veh+tDCS had significantly greater mRNA level compared to the rest of the groups (p
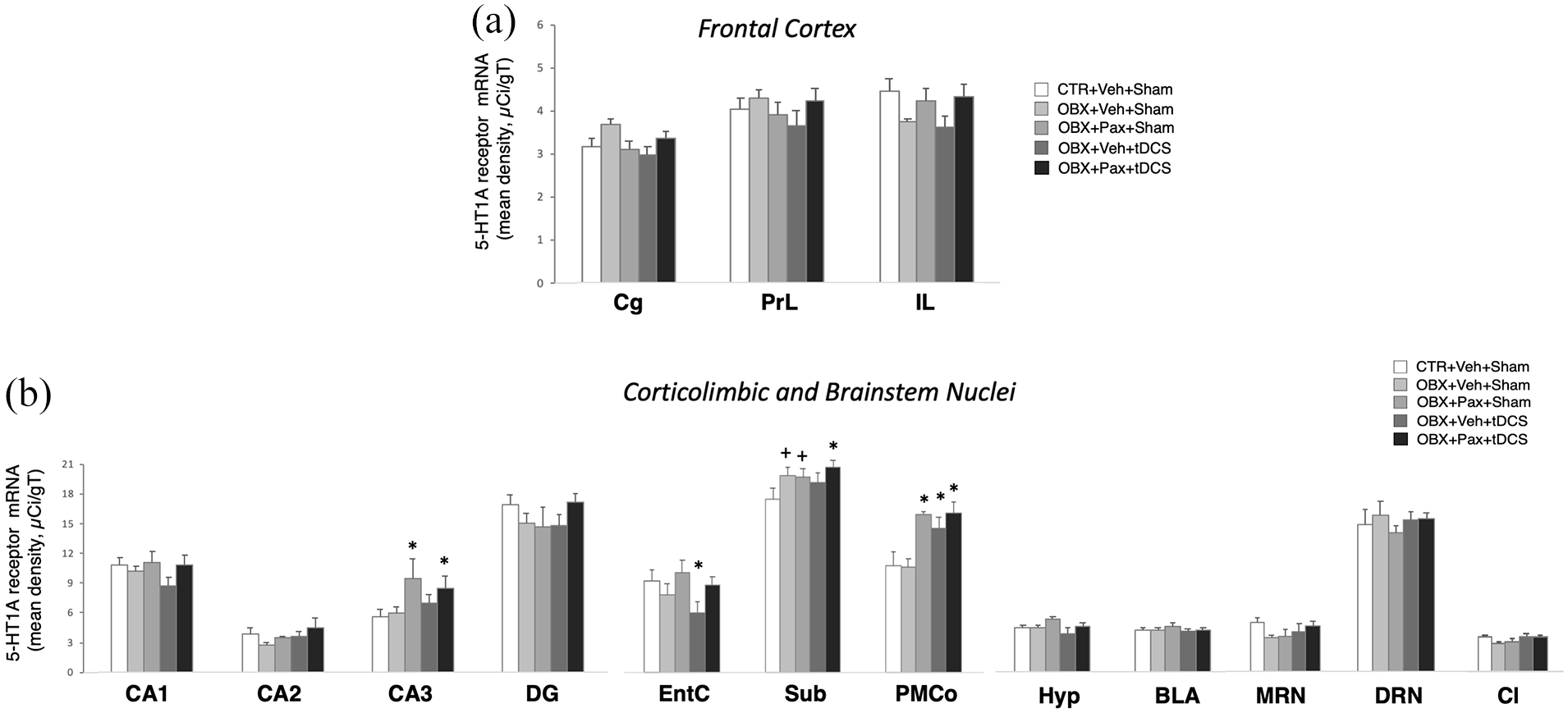
In situ hybridisation for the 5-HT1A receptor. Mean optical density (in micro-Curie per gram of tissue) across ROIs in frontocortical targets of tDCS (a), and corticolimbic and brainstem areas (b). EntC: entorhinal cortex; Sub: subiculum. Bars in graphs represent mean values ± SEM; n = 7−10. +p < 0.05, *p < 0.01.
tDCS and paroxetine modified corticolimbic and brainstem SERT mRNA levels
Two-way ANOVA on SERT mRNA (Figure 7) uncovered a significant main effect of treatment (F4,644=4.029, p=0.003; n=6–8/group) and of ROI (F19,644=184.462, p<0.001; n=20), and a significant interaction (F76,644=1.351, p=0.031). Post hoc comparisons revealed pairwise differences in subcortical and brainstem anxiety/fear circuits. SERT mRNA level for OBX+Veh+tDCS (n=8) was significantly greater than CTR+Veh+Sham (n=10) in the basolateral amygdala (p<0.05), and significantly lower than CTR+Veh+Sham, OBX+Veh+Sham (n=6), OBX+Pax+Sham (n=8) or OBX+Pax+tDCS (n=8) in the median raphe (p<0.01). SERT mRNA for OBX+Pax+Sham was significantly greater than CTR+Veh+Sham or OBX+Veh+Sham in the periaqueductal grey (p<0.05). In the interpeduncular nucleus, OBX+Pax+Sham or OBX+Pax+tDCS was significantly greater than CTR+Veh+Sham or OBX+Veh+Sham (p<0.05). In the zona incerta, OBX+Veh+tDCS was significantly lower than all others. OBX+Pax+Sham or of OBX+Pax+tDCS was significantly greater than CTR+Veh+Sham. No significant effects were obtained from the CTR groups (see Supplementary Figure S6).
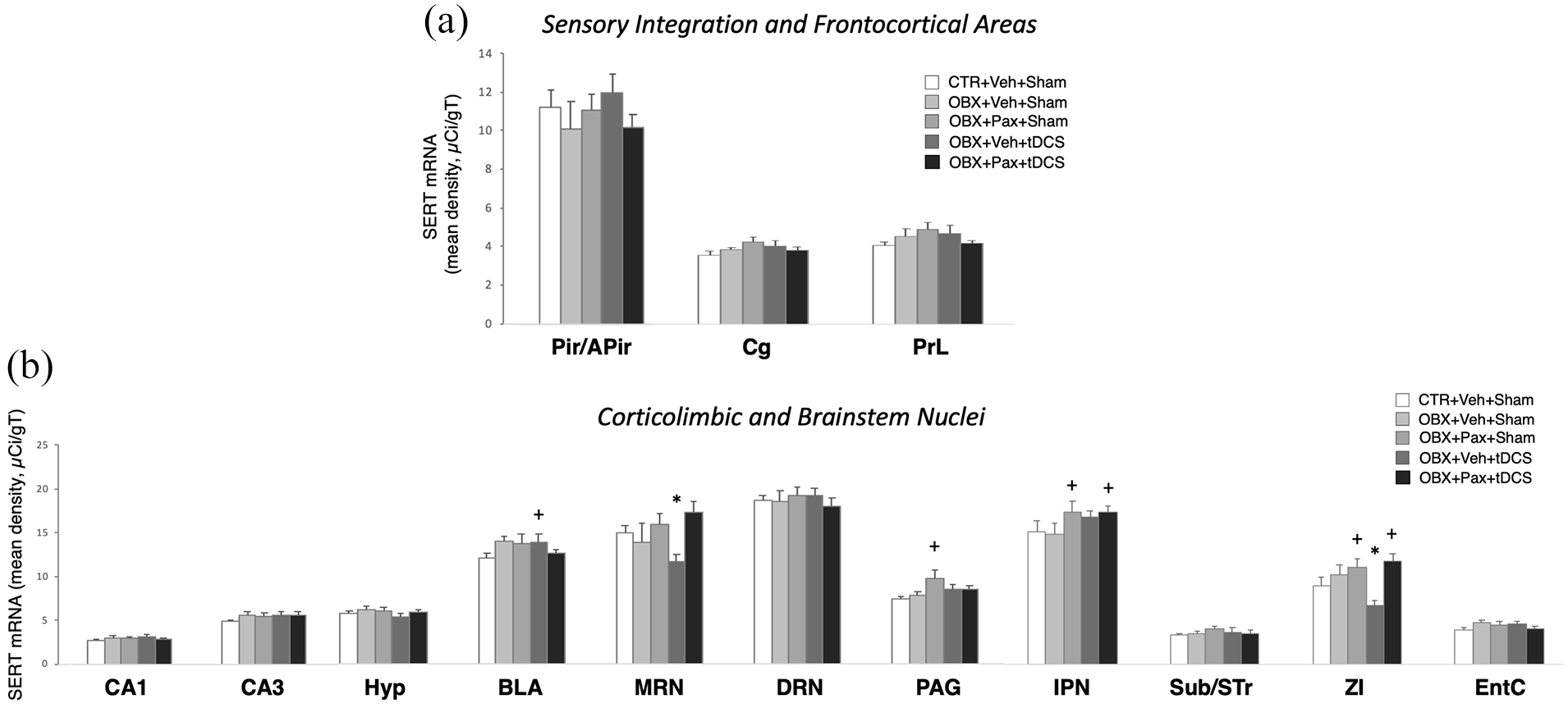
In situ hybridisation for the 5-HT transporter (SERT). Mean optical density (in micro-Curie per gram of tissue) across ROIs in sensory integration and frontocortical areas (a) and in corticolimbic and brainstem areas (b). IPN: interpeduncular nucleus; Sub/STr: subiculum and transition area; ZI: zona incerta. Bars in graphs represent mean values±SEM; n = 6−8. +p < 0.05, *p < 0.01.
Discussion
Here, we examined the effects of paroxetine and tDCS treatment in OBX adolescents in the SPT and OFT. We found that OBX adolescent rats displayed a gradually progressing reduction of SP, as reported with OBX adult rats (Padilla et al., 2018). We chose this as our primary behavioural outcome for a number of reasons. First, both rats lever press to self-administer sucrose, a response that is greatly impacted by stress and in turn mimics the cardinal anhedonia-related symptom of depression (Counotte et al., 2014). Second, it is known that OBX-induced depression-related deficits are not due to olfactory-gustatory and contextual associations or any learning-mediated bias, but rather to the direct behavioural consequences of the insidious cellular changes and rewiring of the limbic forebrain (Morales-Medina et al., 2017; Song and Leonard, 2005). This is likely true with SPT deficits that also rely on the integrity of the hippocampal pathway (Morales-Medina et al., 2017; Padilla et al., 2018; Shin et al., 2017). As such, top-down influences on stress adaptation such as perceived controllability could be somewhat controlled across animals. Lastly, among adolescents post-OBX SP scores most highly correlated with OFT hyperactivity that is classically used to evaluate the OBX depressive-like deficits and hypothesised to reproduce symptoms related to psychomotor agitation, hyperactivity or novelty-sensitivity (Song and Leonard, 2005; Zueger et al., 2005). SP also correlated with plasma BDNF, a peripheral bio-signature for depression pathophysiology (Karege et al., 2002; Klein et al., 2011; Kucera et al., 2019; Molendijk et al., 2011; Pudell et al., 2014). Indeed, in a pilot study (see Supplementary Information), we demonstrated the time course of OBX-induced anhedonia-like reduction of SP to be consistent with the temporal progression of OFT hyperactivity that is the classical pathology-related phenotype evoked by OBX (Song and Leonard, 2005). SP correlated strongly with OFT hyperactivity and weakly with forced swim immobility and novelty-induced suppression of feeding.
In the OFT, paroxetine had no effect on hyperactivity despite the fact that chronic 5-HT reuptake inhibitor treatment reduces OBX-induced hyperlocomotion in adults (Song and Leonard, 2005). This suggests that paroxetine has reduced efficacy in adolescents, consistent with other preclinical studies (Karanges et al., 2011; Vorhees et al., 2011; but see Peanlikhit et al., 2017). Moreover, there is some clinical evidence cautioning for the unwanted occurrences of a set of symptoms termed “activational syndrome”, which can emerge during the course of 5-HT reuptake inhibitor treatment in adolescents (Amitai et al., 2015). This can include agitation, akathisia, nervous hyperactivity, anxiety and apathy, and likely stems from the pharmacological action – possibly off-target – of paroxetine as it interacts with the vulnerable processes of the adolescent brain (Arain et al., 2013; Bowman and Daws, 2019; Bylund and Reed, 2007; Kerestes et al., 2014; Mulder et al., 2003; Tao et al., 2012).
On the other hand, chronic tDCS mono-treatment effectively decreased OBX-induced hyperactivity. Conversely, Filho et al. (2016) found no such effect in healthy adult controls. Among paediatric patients, dorsolateral prefrontal tDCS show improvements of psychomotor and impulsivity symptoms associated with attention-deficit hyperactivity disorder (Allenby et al., 2018). It is intriguing that although most antidepressant drugs require chronic treatment to reverse OBX-induced hyperlocomotion (Song and Leonard, 2005), we found beneficial albeit marginally significant effects after just two tDCS sessions. This putative rapid action has been to some degree observed with DBS (Mayberg et al., 2005) and electroconvulsive therapy (ECT) (Kerner and Prudic, 2014), as well as experimentally with prefrontal optogenetic stimulation (Fuchikami et al., 2015). Follow-up investigations are warranted to confirm whether underlying mechanisms are similar to those of the putative rapid-acting glutamatergic antidepressants, e.g., ketamine (esketamine or Spravato) (Gerhard and Duman, 2018).
In line with our hypothesis, acute paroxetine resulted in a sharp decrease in SP in OBX animals. Worsening of anhedonia and dysphoria has already been suggested by other clinical studies (Goldsmith and Moncrieff, 2011; Le Noury et al., 2015; Vitiello and Ordóñez, 2016). Along with the slow onset of therapeutic action observed in all conventional antidepressants, this may be related to the unfavourable recruitment of inhibitory mechanisms and related neuroplastic processes (Bambico and Belzung, 2013; Bambico and Gobbi, 2008; Bambico et al., 2009a; Bambico et al., 2009b; Gomez et al., 2015; Homberg et al., 2011). This includes, but not exclusively, the activation of somatodendritic 5-HT1A auto-inhibitory receptors arising from the initial indiscriminate effects of drug-induced 5-HT release. This event is an inadvertent early part of these medications’ time course of action that eventually culminates in the desensitisation of inhibitory receptors and/or recovery from inhibitory sequelae that ensue after chronic treatment. Indeed, neuro-physiologically, this time course coincides with the progression of changes in 5-HT neuron activity, with 5-HT reuptake inhibitors acutely decreasing firing rate and chronically recovering it to control levels (Bambico and Gobbi, 2008; Bambico et al., 2009). It is conceivable that among other yet to be identified mechanisms, the acute pro-anhedonic effect of paroxetine observed here was a direct consequence of 5-HT suppression. As to whether this paradoxical effect is shared by other 5-HT reuptake inhibitors or is a unique property of paroxetine requires further investigation. Iñiguez et al. (2010), for instance, found that acute fluoxetine treatment was inert in the SPT when tested in young rats. These however were manipulation-naive animals that were not subjected to any depression paradigm.
Another important finding speaks to the antidepressant potency of tDCS when combined with paroxetine. Remarkably, the decreased SP after acute paroxetine treatment was completely blocked by concurrent tDCS treatment. One possible explanation involves a normalisation of subcortical reward, mood-regulating and motivational systems since acute dorsolateral prefrontal tDCS is able to increase dopamine release in the ventral striatum (Fonteneau et al., 2018). Similarly, prefrontal DBS increases 5-HT activity in rodent studies (Bambico et al., 2018; Hamani et al., 2010). In chronic stress-based models, we have demonstrated decreased dopaminergic and 5-HT neuronal activity (Bambico et al., 2009a, 2009b, 2018), which corroborates some evidence for decreased monoamine activity in depressed patients (Blier and de Montigny, 1994; Bambico et al., 2009a; Bambico and Belzung, 2013). Because acutely administered 5-HT reuptake inhibitors decrease ventral tegmental area dopaminergic or raphe 5-HT activity (Prisco and Esposito, 1995), tDCS potentially normalised SP in OBX animals through an opposite action.
Chronic paroxetine treatment failed to show any antidepressant or psychomotor-stabilising property, which is at odds with the observed reversal of SPT and OFT depressive-like behaviour in adult rats (Casarotto and Andreatini, 2007). As raised by others (Karanges et al., 2011; Klomp et al., 2014), this differential efficacy between adults and adolescents could point to the protracted recruitment of secondary nonspecific mechanisms that nullify therapeutically relevant effects or to a heightened sensitivity of neurodevelopmental growth processes in corticolimbic circuitries. Conversely, tDCS mono-treatment was potent in attenuating OF hyperactivity. And when combined with paroxetine, achieved significant normalisation of SP. Nonetheless, while tDCS-paroxetine co-treatment markedly blocked OBX-induced SP reductions, we did not find the SP scores of chronic paroxetine-treated animals to be significantly lower than the tDCS-paroxetine co-treated animals. This could indicate that chronic paroxetine could still provide some degree of antidepressant protection. Moreover, this could indicate the possibility of further titrating tDCS currents to achieve the complete reversal of anhedonia. To address these questions, future investigations on mechanisms, incorporating isobolographic analyses with broader dose/current ranges, are of paramount importance. These findings support the efficacy of multi-modal treatment strategies, e.g., combined 5-HT reuptake inhibitor and cognitive-behavioural treatment, which have long been established to provide greater benefits for depression. To our knowledge, however, this is the first time we have shown possible evidence for potential synergistic or additive tDCS–paroxetine therapeutic action that can potentially be developed as a safer alternative for adolescent or early-life depression. Similar clinical indications have been demonstrated by Brunoni et al. (2013) but in adult patients.
A possible explanation for this interaction is that long-term potentiation-like plasticity associated with anodal tDCS can be effectively enhanced by concurrent 5-HT reuptake inhibitor treatment (Kuo et al., 2016). In support of this, we have demonstrated treatment-induced alterations in neuroplasticity-related protein and gene markers. OBX animals showed a substantial reduction of plasma BDNF, in agreement with other studies (Kucera et al., 2019; Pudell et al., 2014). Our chronic treatments did not produce any BDNF changes, which could be explained by measurement timing issues. Antidepressant treatments, including ECT, have varying detectability time for BDNF (Sorri et al., 2018; Zhou et al., 2017). Despite this, in our correlational analyses we found that the strength and directionality of the covariability of BDNF with SP and LA were largely reversed by tDCS mono-treatment or by combined tDCS–paroxetine treatment in a manner predictive of antidepressant activity. The heat map correlational matrix patterns for these treatments were closer to normal than either non-treated or paroxetine-treated OBX animals. This suggests that variabilities in depressive-like behaviour and plasma BDNF are dependently influenced by common pathophysiological mechanisms, and that the treatments acted upon these mechanisms.
Because BDNF is linked with activity-dependent plasticity, we measured treatment effects on mRNA indicators of cell activity. Likely due to anterograde neurodegeneration, OBX dramatically attenuated zif268 in the piriform and amygdalopiriform transition areas, an effect that was not reversed by any of the treatments. This was accompanied by increased zif268 in the ventral and anterior hypothalamic areas, known to directly stimulate the hypothalamic–pituitary–adrenal (HPA) stress axis (Bambico and Belzung, 2013). These transneuronal events may well alter the sensitivity and excitability of key limbic projection fields of the extended olfactory–limbic network. Indeed, we found that paroxetine led to increased zif268 in hippocampal CA1 that could be related to its pro-anhedonic effect and recapitulated by chronic stress exposure (Bambico et al., 2019). An abnormal hippocampal activity correlates with increased neuronal atrophy and diminished synaptic plasticity in CA1 (Morales-Medina et al., 2017). Paroxetine also decreased zif268 in the claustrum, which forms a parallel claustral/CA1-prefrontal pathway that is overactivated in patients and animal models (Airan et al., 2007; Bambico et al., 2013; Price and Drevets, 2010). Remarkably, this was blocked by tDCS-paroxetine co-treatment, likely due to electrophysiological modifications in the prefrontal cortex.
Unlike in CA1, the CA2 subfield was activated by tDCS and by tDCS–paroxetine co-treatment. Evidence is accumulating for the functional differences between CA1 and CA2 (Benes, 2010; Dudek et al., 2016; Gisabella et al., 2009). Unlike CA1, CA2 is endowed with dense afferences from the basolateral amygdala, with dense projections to the entorhinal cortex and septal nuclei, harbours neurotrophic activity without displaying long-term potentiation, and is more resistant to neurodegeneration. Increased CA2 activity may therefore be akin to an antidepressant activity.
We detected treatment-induced alterations in 5-HT1A receptor and SERT mRNA, which are the classical molecular targets of monoamine-modulating antidepressants/anxiolytics (Bambico and Belzung, 2013; Blier and de Montigny, 1994; Kraus et al., 2017). Treatment-induced changes in 5-HT1A receptor were almost exclusively observed in parahippocampal/hippocampal subregions, including the entorhinal cortex, subiculum and CA3. CA3 changes are in line with its function as the major hippocampal input component, and thus endowed with dense 5-HT afferences and postsynaptic receptors (Blier and de Montigny, 1994). The effects of paroxetine and tDCS–paroxetine co-treatment may therefore indicate a sensitised response or tonicity of postsynaptic 5-HT receptors, a feature shared by all tested antidepressant treatments (Blier and de Montigny, 1994; Haddjeri et al., 1998). On the other hand, SERT changes were detected mainly in brainstem fear/anxiety-relevant regions, composed of the amygdala, periaqueductal grey, zona incerta, interpeduncular nucleus and median raphe nucleus. The profound effect of tDCS on SERT in key structures is proposed to mainly contribute to the observed effects on OF psychomotor agitation and hyperactivity, similar to that reported by others (Mirza et al., 2007).
Prefrontal tDCS did not affect zif268, serotoinin-1A receptor and SERT in any of the tDCS target subregions, i.e., the cingulate, prelimbic and infralimbic. This seems to indicate a lack of persistent local in situ neuroplastic activity, corroborating reports of subtle electrophysiological changes within the tDCS current arch (Stagg et al., 2018). Rather, transsynaptic consequences of cumulative membrane potential augmentation, e.g., 5-HT release, could be occurring in the corticolimbic projections sites, as we have previously shown with prefrontal or cerebellar DBS (Bambico et al., 2018; Hamani et al., 2012; Volle et al., 2018).
Finally, in opposition to OBX animals, no treatment effects were noted in non-OBX controls. Hence, tDCS exerts therapeutically relevant changes under pathophysiological conditions, and hones in to correct or compensate for corticolimbic mechanisms to re-establish behavioural homeostasis and molecular adaptation.
There is a pressing need for novel treatments that can effectively relieve depression without causing adverse side effects, especially within the younger population. The results of the current study support the use of tDCS in combination with antidepressant drug therapy. As it stands, tDCS adjunctive treatment can be particularly useful in preventing the early adverse and paradoxical effect of paroxetine and improve its long-term therapeutic prognostic. Moreover, tDCS by itself shows some promise as a treatment for psychomotor disturbances with fast and sustained efficacy. Similar to DBS, the therapeutic mechanism of prefrontal tDCS seems to include restricted activational changes and widespread 5-HT effects within downstream corticolimbic regions. Future investigations could potentially guide its use as a non-invasive treatment option for early-life and adolescent depression.
Supplemental Material
sj-docx-1-jop-10.1177_02698811211000765 – Supplemental material for Antidepressant action of transcranial direct current stimulation in olfactory bulbectomised adolescent rats
Supplemental material, sj-docx-1-jop-10.1177_02698811211000765 for Antidepressant action of transcranial direct current stimulation in olfactory bulbectomised adolescent rats by Shannon C Waye, O Chandani Dinesh, SM Nageeb Hasan, Joshua D Conway, Roger Raymond, José N Nobrega, Jacqueline Blundell and Francis Rodriguez Bambico in Journal of Psychopharmacology
Footnotes
Author contributions
Shannon Waye: background research, study execution, data collection and curation, analyses, writing/original draft preparation. Francis Bambico: conceptualisation, methodology, supervision, writing, reviewing and editing. Jacqueline Blundell: methodology, supervision, reviewing and editing. José Nobrega: methodology, supervision, reviewing and editing. O. Chandani Dinesh: data analysis, reviewing and editing. Nageeb Hasan: study assistance and data analysis. Joshua Dean Conway: study assistance. Roger Raymond: study assistance and methodology.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by grants from the Janeway Foundation and Brain Canada/Azrieli Foundation.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.