
Abstract
Background:
Cycloprolylglycine (CPG) is an endogenous dipeptide with a wide range of psychotropic activity and putative therapeutic potential for depression. A small but growing body of data suggests that antidepressant-like effect of CPG is associated with neuroplastic changes in the brain or 5-HT system modulation. However, the mechanisms of the dipeptide action remain elusive.
Aims:
Here, we characterize the effects of chronic CPG administration on behavior and genes expression of antidepressants sensitive catalepsy (ASC) mice strain, characterized by depressive-like behavior.
Methods:
ASC mice were injected with saline, fluoxetine (10 mg/kg/day), or CPG (1 and 2 mg/kg/day) during 2 weeks. Behavior was studied using the open field test, novel object test, elevated plus maze test, forced swim test, and tail suspension test (TST). The expressions of genes coding BDNF, CREB, 5-HT1A and 5-HT2A receptors, TPH2, and SERT in the brain were measured with quantitative real-time reverse transcription polymerase chain reaction (RT-PCR).
Results:
Chronic intraperitoneal administration of 1 and 2 mg/kg of CPG revealed the significant antidepressant-like effect by decreasing immobility time in the TST. At the same time, CPG did not negatively affect locomotor activity, cognition, or anxiety. In the real-time quantitative polymerase chain reaction (PCR) assay, chronic CPG treatment (2 mg/kg for 14 days) increased Bdnf mRNA level in the frontal cortex.
Conclusions:
Our findings extend the evidence for the effectiveness of CPG to reduce depressive-like behaviors. The antidepressant-like effect of CPG is mediated, as least in part, by BDNF-dependent mechanism. The exact mechanism remains to be elucidated, and further studies are warranted.
Introduction
Major depression (MD) is a highly prevalent and debilitating mental illness. The World Health Organization (WHO) reported that MD was becoming the leading cause of disability worldwide. The global burden of depression is constantly growing due to high risk of relapse and treatment resistance (Lépine and Briley, 2011). Despite decades of extensive research and high incidence of depression, its pathogenesis up to the present remains unclear. Unfortunately, approximately half of all patients with MD fail to respond to first-line therapy, and more than 65% do not achieve remission (Liu et al., 2017). Furthermore, all classes of antidepressants have a wide range of undesirable effects (Jureidini et al., 2004; Kollhorst et al., 2019; Mazhar et al., 2016; Yekehtaz et al., 2013). These facts accentuate the need for developing a new effective medicine for depression treatment with a more tolerable side effect profile.
Natural compounds and their synthetic analogues are of great interest for the development of new psychotropic therapies. One such substance is a dipeptide cycloprolylglycine (CPG, pyrrolidino[1,2-a]-2,5-diazacyclohexane-1,4-dione). Since CPG synthesis in 1991 and its discovery as an endogenous compound in rat brain in 1996 (Gudasheva et al., 1996a), in vitro and animal studies have revealed a wide range of CPG psychotropic activities. It was shown that the dipeptide possesses antiamnesic (Gudasheva et al., 1999), anxiolytic (Seredenin et al., 2002), antihypoxic, and neuroprotective effects (Povarnina et al., 2016). Previously it was shown that CPG chronic administration significantly decreased immobility time in inbred BALB/c mice in the forced swim test (FST) (1 and 2 mg/kg, 2 weeks, i.p.) and C57BL/6 mice in the tail suspension test (TST) (1 mg/kg, 4 weeks, i.p.) (Vasileva et al., 2019). Radioligand binding studies in BALB/c mice brains after 2-week CPG administration (1 and 2 mg/kg, i.p.) revealed a decreased number of NMDAR (2 mg/kg) and 5-HT2AR (1 and 2 mg/kg) binding sites in hippocampus and striatum, respectively, and increased GABAAR (1 mg/kg) binding density in the frontal cortex (Abdullina et al., 2019). Previous in vitro studies have shown that CPG enhances AMPA receptor-mediated currents (Gudasheva et al., 2016a) and increases BDNF level under normal and neurotoxic conditions in neural cell culture (Gudasheva et al., 2016b).
It is well known that BDNF is involved in the mechanisms of depression and antidepressant drugs. Upregulation of the BDNF mRNA expression in hippocampus and cortical regions parallels the antidepressant-like effect of clinically used antidepressants (Nibuya et al., 1995, 1996). Subsequent studies showed that a single infusion of BDNF into the rodent midbrain area or hippocampus produces sustained antidepressant-like effects in behavioral tests (FST and TST) and in the learned helplessness paradigm (Naumenko et al., 2012; Shirayama et al., 2002; Siuciak et al., 1997). BDNF expression is regulated by the transcription factor CREB (cyclic AMP response element binding protein), which is upregulated by chronic antidepressants administration in a similar manner as for BDNF (Nibuya et al., 1996).
Research findings provide a strong evidence that links serotonergic system to the etiology of depression and the mechanisms underlying antidepressant response. A great majority of all antidepressants aim to increase extracellular 5-HT levels by targeting serotonin transporter or monoamine oxidase enzyme responsible for serotonin catabolism (Celada et al., 2004). But equally important therapeutic targets in depression treatment are serotonin receptors, in particular 5-HT1A and 5-HT2A, both of which were found to be highly expressed in the frontal cortex. Dysfunction of these receptors may contribute to depressive disorder, as evidenced by studies in suicide victims (Arango et al., 1997). Moreover, all findings seem to indicate that 5-HT1AR agonists as well as 5-HT2AR antagonists may augment the clinical effects of conventional antidepressants when they are given in combination (Celada et al., 2004). Taking into account the close link between BDNF, 5-HT system, and antidepressant activity as well as available data on CPG impact on the components of these systems, a more detailed study in this direction seems to be required.
To examine antidepressant-like effect of CPG and its possible mechanism, we used ASC (antidepressants sensitive catalepsy) mice with genetically determined depressive-like state. Mice of ASC strain were selected for high predisposition to catalepsy from a backcross population of catalepsy-prone CBA/Lac and catalepsy-resistant AKR/J strains (Kondaurova et al., 2006; Kulikov et al., 2008a). It was shown that the expression of catalepsy in mice can be enhanced by stressful stimuli suggesting a close association between the mechanisms of pinch-induced catalepsy and response to emotional stress. The catalepsy-prone genotypes demonstrated significantly higher susceptibility to stress compared with catalepsy-resistant mice (Tikhonova et al., 2013). Besides, the genetically determined catalepsy in these animals is sensitive to chronic administration of selective serotonin reuptake inhibitors (SSRIs) (Tikhonova et al., 2010) and tricyclic antidepressants (Tikhonova et al., 2006). ASC mice exhibit increased immobility in the FST and TST, which is considered as evidence for the predisposition to “depression,” low locomotor and exploratory activity (Bazovkina et al., 2005), disturbances in sexual behavior and motivation (Tikhonova and Amstislavskaya, 2016), and impaired extinction of a conditioned passive avoidance response (Dubrovina et al., 2009). Depressive-like phenotype of ASC mice is associated with significantly decreased immune response (Alperina et al., 2007) and altered brain serotonin system (Tikhonova et al., 2013), in particular, functional disturbances of the 5-HT1A and 5-HT2A receptors in the frontal cortex which may contribute to depression in humans (Slifirski et al., 2021). These findings indicate that ASC mouse strain can be a convenient model of stress-based neuropathology to study antidepressant effects on behavior.
Thus, the aim of this study was to assay the antidepressant-like effect of chronic CPG treatment on genetic mouse model of depression and, using model mice, identify the impact of the dipeptide on the expression of key components of BDNF and 5-HT systems. The behavioral response to CPG administration of ASC mice was assessed by two widely used preclinical screening tests, the TST and the FST in comparison with classic antidepressant fluoxetine. To exclude false positive results in the TST and the FST and examine possible side effects on anxiety and memory, the open field, the elevated plus maze (EPM), and the novel object recognition tests were conducted. Furthermore, to evaluate whether the observed antidepressant-like effect is associated with alterations in serotonergic system or BDNF, the expressions of Htr1a, Htr2a, Tph2, Slc6a4, Bdnf, and Creb1 genes in the brain structures of ASC mice chronically treated with CPG were measured using real-time quantitative polymerase chain reaction (PCR).
Materials and methods
Animals
Adult male ASC mice (2–3 months old, weighing 26 ± 2 g) were used. ASC strain was created in the Laboratory of Behavioral Neurogenomics, Institute of Cytology and Genetics, as the result of prolonged selective breeding for high predisposition to catalepsy from the (CBAx(CBAxAKR)) backcross population between CBA/Lac and AKR/J strains. CBA/Lac is an inbred mouse strain with genetical predisposition to catalepsy reaction in response to pinching the scuff of their neck, while mice of inbred AKR strain are catalepsy-resistant and never display cataleptic freezing under normal conditions (Kondaurova et al., 2006; Kulikov et al., 2008a). Since 2006, this strain has been maintained by brother–sister crossing. The breeding was conducted at the Center for Genetic Resources of Laboratory Animals at ICG SB RAS, supported by the Ministry of Education and Science of Russia (Unique identifier of the project RFMEFI62119X0023) in accordance with the Directive 2010/63/EU of the European Parliament and the Council as of 22 September 2010.
All experimental procedures were in compliance with the Guide for the Care and Use of Laboratory Animals, Eighth Edition, Committee for the Update of the Guide for the Care and Use of Laboratory Animals, National Research Council© 2020 National Academy of Sciences (USA) for animal experiments, and the trial was approved by the ICG SB RAS ethics committee and registered in ICG SB RAS (Protocol No. 34, 15.06.2016).
Drugs
CPG was synthesized at the Department of Chemistry, Institute of Pharmacology, as described in the article Gudasheva et al., 1996b. The synthesized CPG had the following characteristics. m.p., 213–214°C (decomp.); 1H NMR spectrum (DMSO-d6 + CF3COOD) (δ, ppm): 1.7–2.2 (m, 4H, CγH2(Pro), CβH2(Pro)), 3.39 (m, 2H, CδH2(Pro)), 3.53 and 4.0 (2d, 2H, J 16.65 Hz, unequiv. CαH2 (Gly)), 4.13 (t, 1H, CαH (Pro)), 8.11 (bs, 1H, NH); Anal. found: C, 54.41; H, 6.54; N, 18.20%. calcd. for C7H10N2O2.: C, 54.54; H, 6.54; N, 18.17%. CPG was dissolved in isotonic saline and injected intraperitoneally in doses of 1.0 or 2.0 mg/kg.
Fluoxetine hydrochloride ((±)-N-Methyl-γ-[4-(trifluoromethyl) phenoxy]benzenepropanamine hydrochloride) (Sigma-Aldrich) was dissolved in saline and administered intraperitoneally in dose of 10 mg/kg.
Experimental procedures
Thirty-two ASC mice were separated on four groups (eight animals each) and were daily treated for 16 days intraperitoneal with saline, fluoxetine (10 mg/kg), or CPG (1 and 2 mg/kg) (100 μL/10 g body weight). The dose choice was based on earlier studies (Vasileva et al., 2019), and literature data showing that serum fluoxetine levels for the 10 mg/kg/day dose correspond with the bottom of the range of plasma levels found in patients taking 20–80 mg/day Prozac (Dulawa et al., 2004). The mice were housed under standard laboratory conditions in a natural light–dark cycle (12 h light and 12 h dark) with free access to water and food. The animals were isolated to eliminate the group effect from the first day of drug administration. On the 11th day of treatment, the behavior battery of tests started, and all animals were subjected to the tests in the following order: open field, novel object recognition test, FST, TST, and EPM test. On test per day, object recognition test duration was 2 days. The assignment of treatments to the animals or measurements was counterbalanced. All behavioral procedures were performed between 15:00 and 19:00 in the dark time, 3 h after the drug administration. On the 16th day of injection after the EPM test, the mice were sacrificed and decapitated. The frontal cortex, hippocampus, and midbrain areas were dissected, frozen in liquid nitrogen, and stored at −80°C until RNA isolation.
Open field test
The open field test was performed in a circle arena (60 cm in diameter) bordered with a white plastic wall (25 cm high). The mouse was placed near the wall, and its behavior was recorded for 5 min. The distance traveled (cm), the time spent in the center of arena (%), and the duration of rearing and grooming (s) were automatically tracked with EthoStudio (Kulikov et al., 2008b). The arena was cleaned using napkins after each animal.
The novel object recognition test
The object recognition test was conducted in an open field arena the day next after the open field test. After 24 h, the mouse was allowed to explore the familiar arena (familiarization phase) with two identical objects (Fo + Fo) placed at an equal distance. The duration of this phase was 10 min. On the third day (test phase), the mouse was returned in the arena with two objects: one was familiar and other was novel (Fo + No). The test lasted for 10 min (Antunes and Biala, 2012). The index (iNo) of time spent near the novel object (%) was calculated as follows:
FST
Mice were placed in a transparent cylindrical tank (30 cm height, 18 cm diameter) filled with water at 25 ± 2°C for 6 min including 2 min of adaptation and 4 min of immobility time recording by an experienced rater blind to the experiment. Water was changed after each test.
TST
Mice were suspended by the tail using an adhesive tape on the horizontal bar positioned at 50 cm height. The behavior of mice was registered during 6 min with EthoStudio software. The complete time of immobility was measured by an experienced rater blind to the experiment.
EPM test
The EPM test was carried out in a plus-shaped apparatus that consisted of two closed and two open arms (35 × 7 cm) placed 50 cm above the ground. The closed arms bordered the walls 20 cm in height. Mouse was placed in the center of the apparatus facing the closed arm and was allowed to explore the maze for 5 min. Distance traveled (cm), time spent (%) in the center, open arms, and closed arms were analyzed automatically with EthoStudio software. The number of peeks and head dips were counted by an experienced rater blind to the experiment. The apparatus was cleaned after each session.
Real-time quantitative PCR
The extraction of total RNA was performed with Trizol reagent according to the manufacturer’s protocol. The cDNA was synthesized as discussed in previous articles (Kulikov et al., 2005; Kulikova et al., 2016; Naumenko et al., 2008; Naumenko and Kulikov, 2006). The number of cDNA copies of genes of interest were obtained using selective primers (Table 1) and SYBR Green I fluorescence detection kit (R-402, Syntol, Russia) for real-time PCR. As standards we used mouse genomic DNA extracted from the liver with known concentrations. The gene expressions were evaluated as the numbers of cDNA copies with respect to 100 copies of housekeeping gene Polr2a cDNA. Real-time PCR was carried out using the LightCycler 480 apparatus (Roche Diagnostics, Switzerland). The reaction protocol consists of 95°C, 4 min; then 40 cycles 95°C, 15 s; specific temperature for primers (Table 1), 30 s; and 72°C, 20 s. To monitor amplification specificity, we performed melting curve analysis at the end of each run for each primer pair (details in Supplemental material).
Primer sequences, annealing temperatures, and PCR-product lengths.
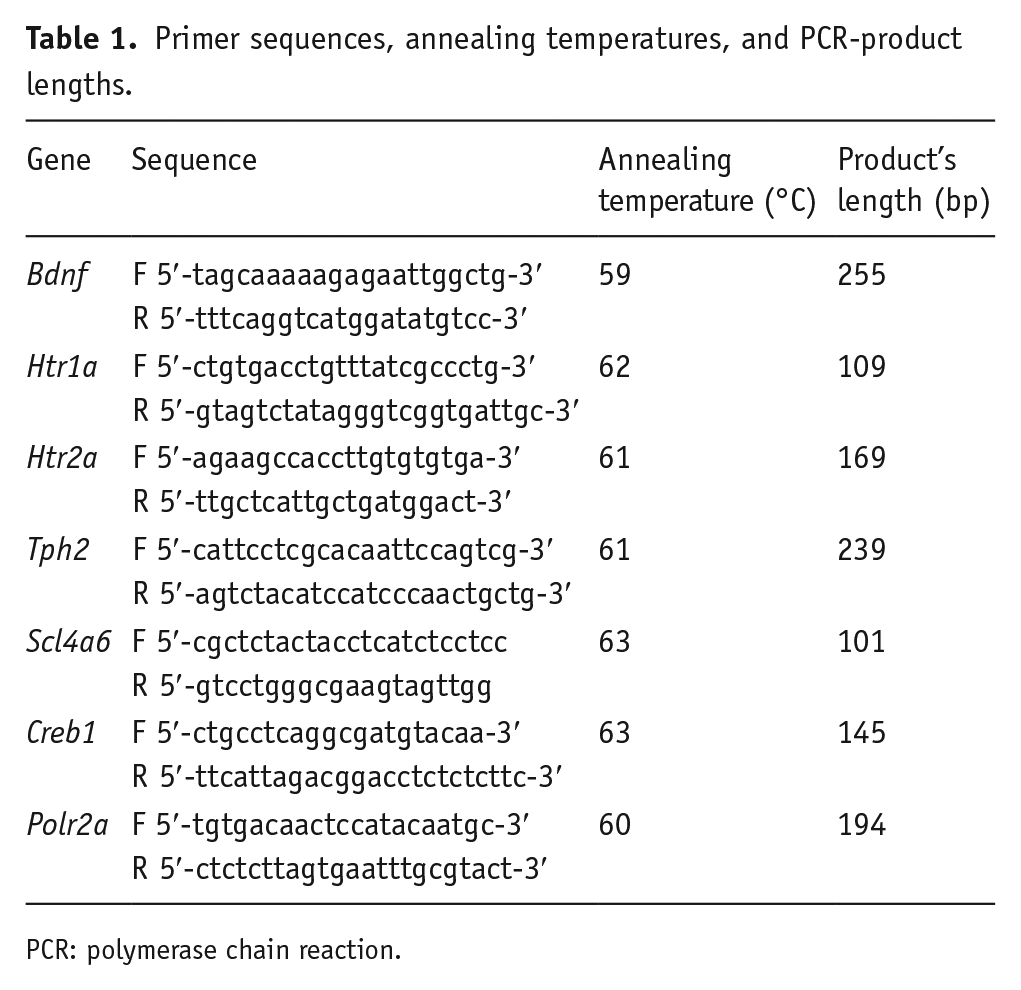
PCR: polymerase chain reaction.
Statistics
The values of other behavioral traits and mRNA levels were presented as means ± SEM (standard error of the mean) and compared with one-way analysis of variance (ANOVA) followed by Fisher’s least significant difference (LSD) test for multiple comparisons. The normality and equality of dispersion were checked using Lilliefor’s and Barlett’s test, respectively, and data conformed to normal distribution patterns.
Results
Effect of chronic CPG treatment on immobility in the TST and FST
Figure 1 shows that CPG markedly and dose dependently reduced immobility time in the TST in comparison with the vehicle (F(3,27) = 6.42, p < 0.01). These effects were comparable with those observed for the clinically used SSRI antidepressant fluoxetine. At the same time, in the FST, neither CPG in two doses nor fluoxetine produced any statistically significant effects on the immobility (F(3,25) = 1.77, p > 0.05).
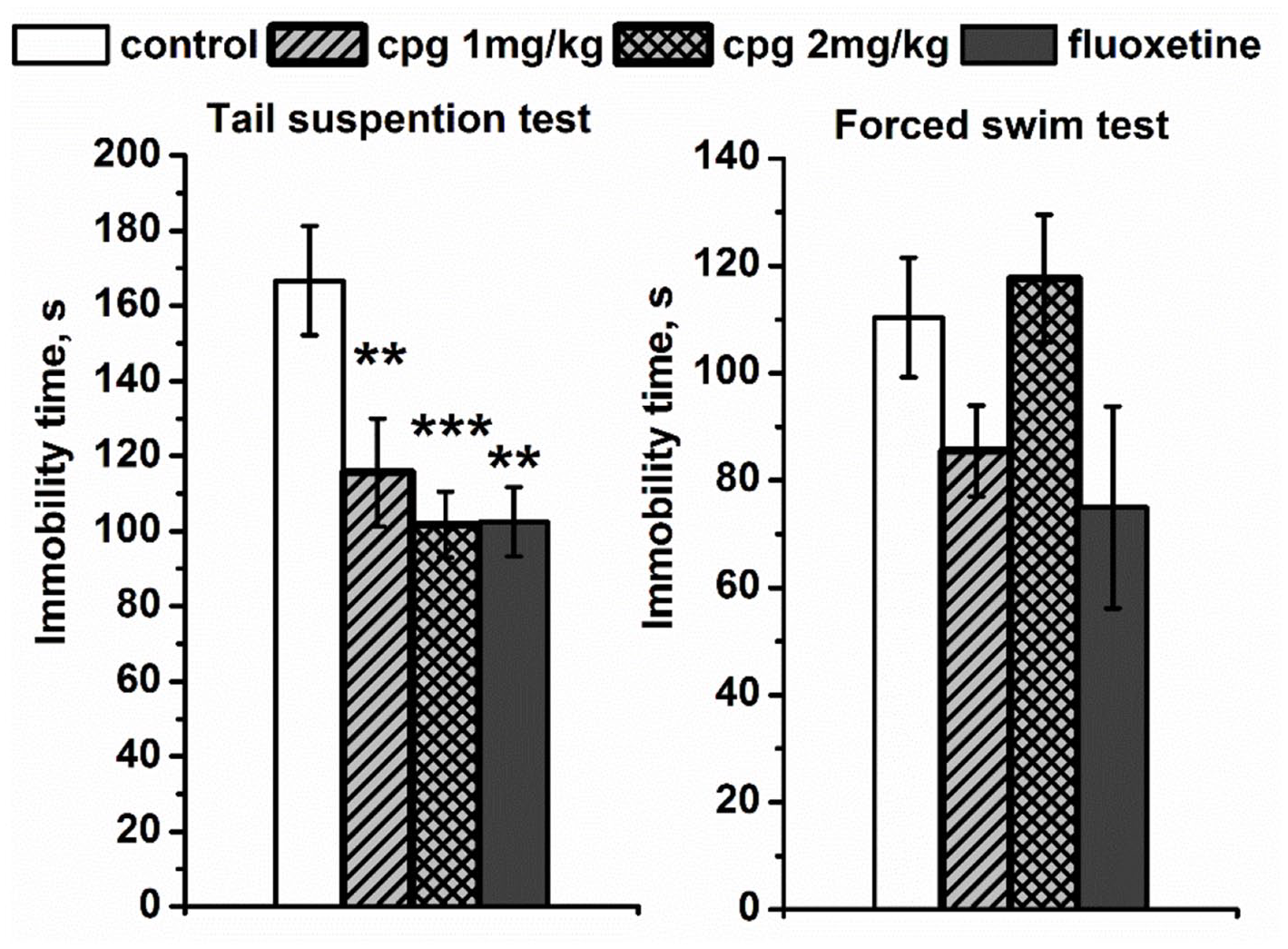
The effects of the chronic CPG administration (1 and 2 mg/kg, i.p.) on the immobility time in the tail suspension and forced swim tests. All values are presented as mean ± SEM. N = 8 for all groups.
Effect of CPG and fluoxetine on the behavior in the open field, novel object recognition, and EPM tests
To evaluate possible adverse effects of the dipeptide, mice chronically treated with CPG were subjected to behavioral tests for locomotor activity, anxiety, and cognition. In the open field test (Table 2), the distance run, time spent in the center, and time spent in grooming did not differ between all experimental groups. So, neither CPG nor fluoxetine induced a change in locomotor activity, excluding the possibility of a false positive in the TST. The anxiety component was also unaffected. At the same time, mice that received CPG at the dose 1 mg/kg showed increased duration of rearing compared with vehicle that may indicate increased exploratory activity.
The effect of chronic CPG (1 and 2 mg/kg) and fluoxetine (10 mg/kg) administration on the behavior in the open field test and the novel object recognition test.
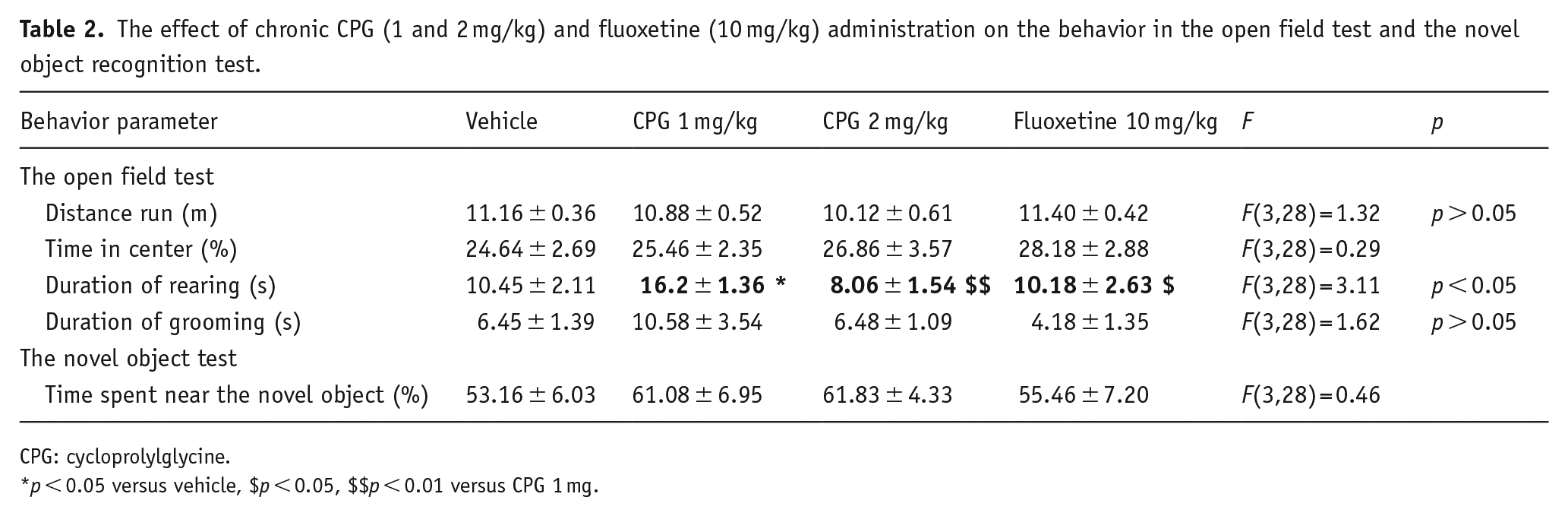
CPG: cycloprolylglycine.
p < 0.05 versus vehicle, $p < 0.05, $$p < 0.01 versus CPG 1 mg.
In the novel object recognition test (Table 2), no changes in total time of contact with the novel object were observed in any of the groups, suggesting that neither CPG nor fluoxetine influences mice cognitive functions, particularly memory processes.
In the plus maze (Table 3) test, no differences in total distance and time spent in the center, open arms, and closed arms were observed in CPG groups compared with vehicle. But fluoxetine treatment resulted in significant increase of time spent in the open arms and decrease of time spent in the closed arms relative to control-vehicle group.
The effect of chronic CPG (1 and 2 mg/kg) and fluoxetine (10 mg/kg) administration on the behavior in the elevated plus maze test.
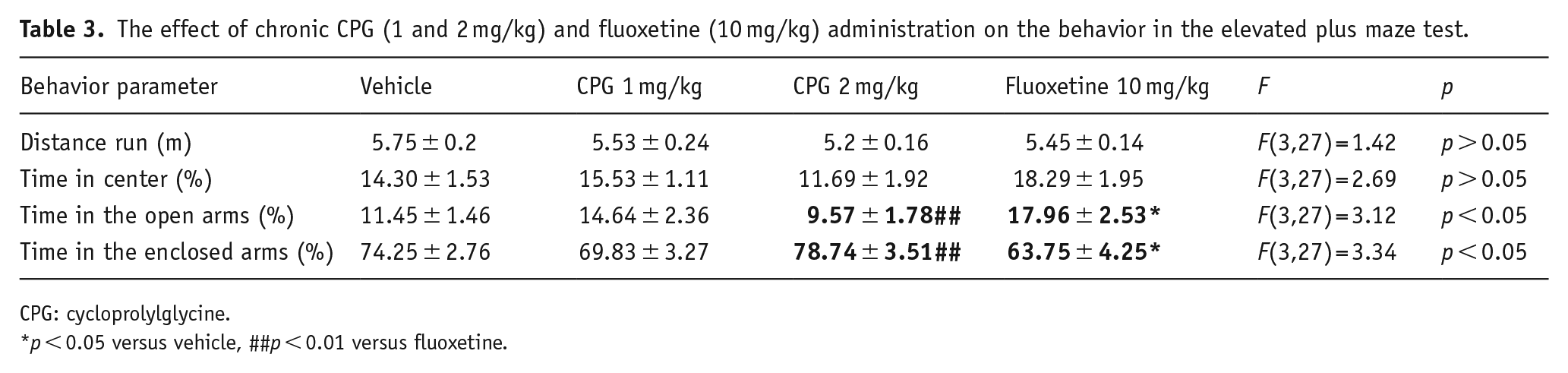
CPG: cycloprolylglycine.
p < 0.05 versus vehicle, ##p < 0.01 versus fluoxetine.
Effect of CPG and fluoxetine on the genes expression in the brain
In the frontal cortex, we found a slight effect of drugs on the Bdnf mRNA level (F(3,26 )= 2.86, p = 0.056); thus, CPG in the dose of 2 mg/kg increased the expression of this gene (p < 0.01) in comparison with control, whereas the Bdnf expression in the hippocampus was significantly elevated after fluoxetine administration (F(3,27) = 24.65, p < 0.001) in comparison with all other groups. At the same time, there was no effect of chronic drug administration on the Bdnf (F(3,27) = 0.16) (Figure 2), Tph2 (F(3,27) = 0.45), and Scl4a6 (F(3,24) = 2.98, p = 0.052) (Figure 3) mRNA levels in the midbrain as well as the mRNA level of Htr1a, Htr2a, and Creb genes in all studied structures (Figure 2, Table 4). The Slc6a4 and Tph2 mRNA expression is specific for the serotoninergic neurons, which are localized in the raphe nuclei of the midbrain (Bach-Mizrachi et al., 2006; Bengel et al., 1997; Fujita et al., 1993; Lin et al., 2007; Patel et al., 2004). Therefore, in this study, we did not investigate the mRNA level of Slc6a4 and Tph2 genes in frontal cortex and hippocampus.
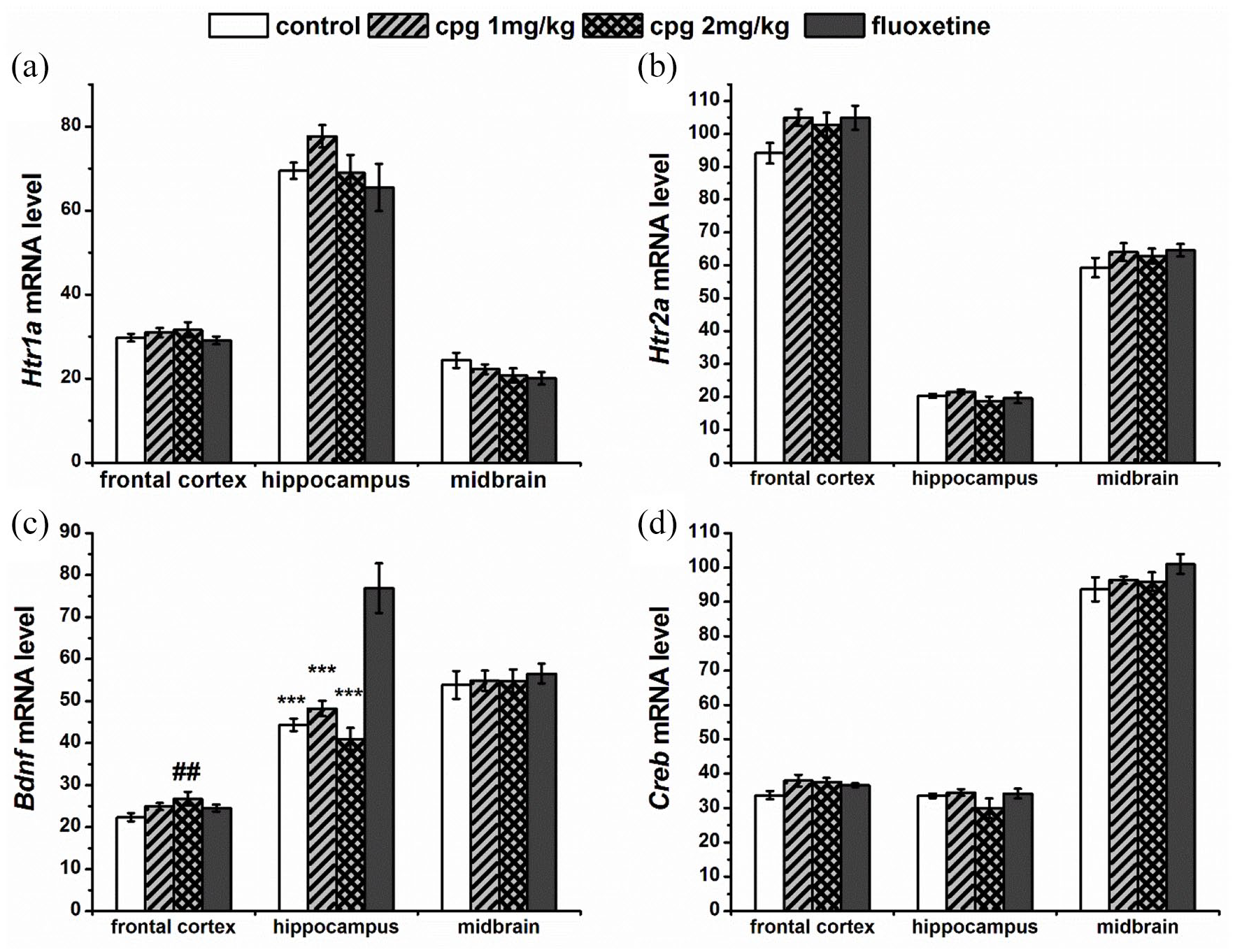
Effect of chronic drugs administration on Htr1a (a), Htr2a (b), Bdnf (c), and Creb1 (d) mRNA level in the frontal cortex, hippocampus, and Creb1 (d) mRNA level in the frontal cortex, hippocampus, and midbrain. Gene expression is presented as the number of cDNA copies with respect to 100 cDNA copies of Polr2a, which were used as a calibrator. All values are presented as mean ± SEM. N = 8 for all groups.
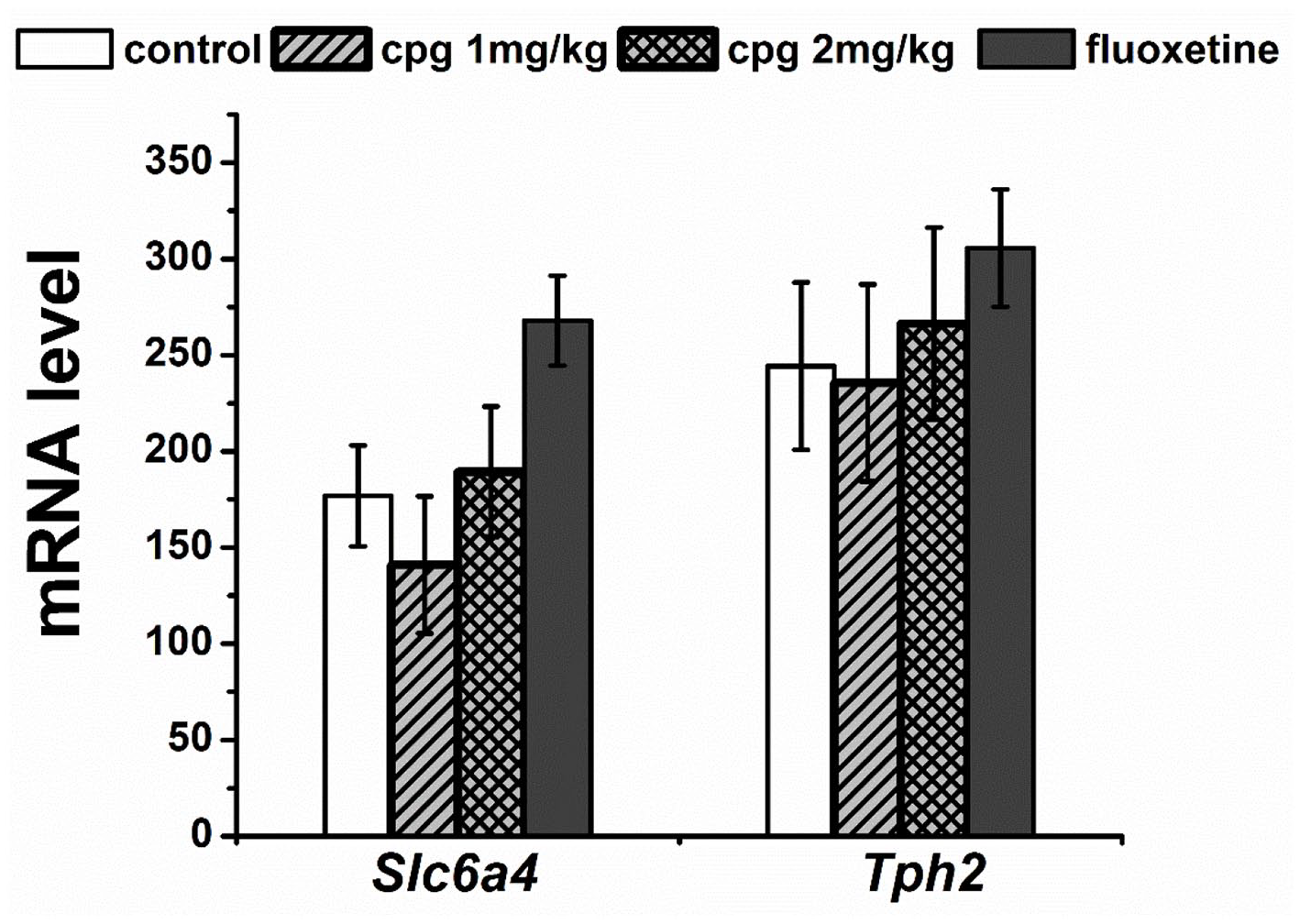
Effect of chronic drugs administration on Scl4a6 and Tph2 mRNA level in the midbrain. Gene expression is presented as the number of cDNA copies with respect to 100 cDNA copies of Polr2a, which were used as a calibrator. All values are presented as mean ± SEM. N = 8 for all groups.
Effect of chronic CPG (1 and 2 mg/kg) and fluoxetine (10 mg/kg) administration on the genes expression.
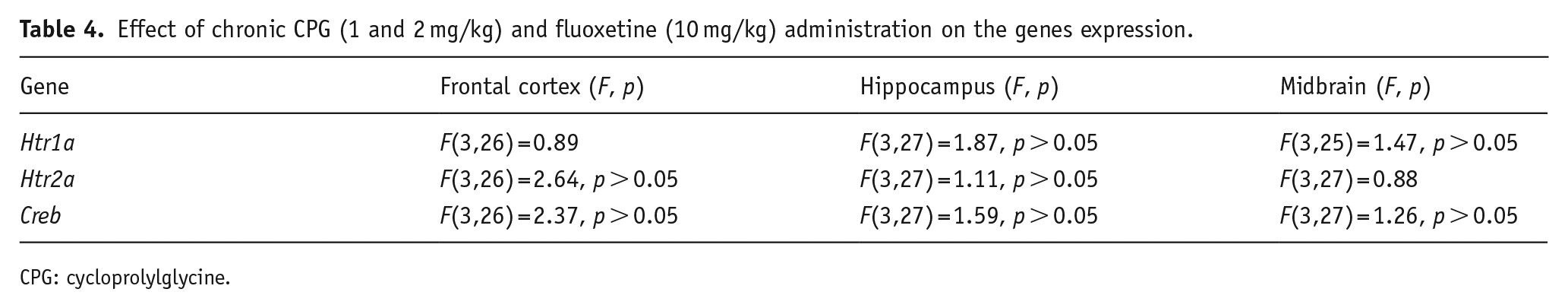
CPG: cycloprolylglycine.
Discussion
Previously we studied antidepressant-like effect of CPG in the FST and the TST using BALB/c and C57BL/6 mice that were not subjected to any intervention to induce a depression-like state. It was found that chronic CPG administration produces antidepressant-like effect in the FST in BALB/c mice only, but the opposite was found in the TST, in which CPG decreased immobility time in C57BL/6 but not in BALB/c strain (Vasileva et al., 2019).
In order to further study the antidepressant-like activity of CPG, we draw our attention to ASC mice, used as genetic model of depression (Bazovkina et al., 2005; Kondaurova et al., 2006). In particular, this study was designed to investigate behavioral effect of synthetic analogue of endogenous peptide CPG using animal model of depression and its related mechanisms focusing on the serotoninergic system and neuroplasticity markers.
It was shown that 2-week administration of 1 and 2 mg/kg of CPG produced strong antidepressant-like effect in the TST. The observed effect was comparable with that of fluoxetine at the dose 10 mg/kg. However, in the FST, chronic CPG administration at the both doses failed to reduce immobility time of ASC mice, as well as fluoxetine. So, CPG was effective in all tested mice strains but in different behavioral paradigms, affirming that these tests produce behavioral response to acute stress via different underlying mechanisms, which may vary depending on the strain characteristics of its biological substrates (Chatterjee et al., 2012; Jin et al., 2017).
An additional behavioral study on ASC mice revealed that CPG treatment did not affect distance run, grooming, and time spent in the center in the open field test. At the same time, the CPG induced increases in the duration of rearing that may be attributed to enhanced exploratory activity. Also CPG treatment did not affect mice behavior in the novelty object test as well as in the EPM, while fluoxetine produced anxiolytic effect as expected according to the literature (Kurt et al., 2000). Thus, antidepressant-like effect of CPG was not accompanied by alterations in locomotor activity or anxiety-related behavior as well as there was no negative influence on exploratory behavior or memory function. In contrast, mice treated with CPG at the dose 1 mg/kg showed increased exploratory activity compared with vehicle.
Behavioral studies on BALB/c, C57BL/6, and ASC mice demonstrated that strain-dependent response to CPG and fluoxetine in the TST and FST are identical, indicating that these agents may have a shared mechanism.
It was established that chronic antidepressant treatment increases the expression or function of BDNF in limbic structures, most notably the hippocampus (Nibuya et al., 1995, 1996). This is in line with present results that showed almost two-fold upregulation of BDNF mRNA in the hippocampus and an absence of any influence on BDNF expression in the cortex of fluoxetine-treated group. While the main effect of CPG treatment was found in the frontal cortex, we did not observe any effect in the hippocampus. We could suggest that this specificity may be due to the action of CPG on receptors, such as GABAA, NMDA, and 5-HT2A, which are abundantly expressed in this region. It should be noted that CPG does not bind to these receptors but alters their density (Abdullina et al., 2019) and possibly functional activity of these neurotransmitter systems. Interestingly, the both doses of CPG have been demonstrated to be effective in the TST, but only the higher dose increased significantly BDNF mRNA level. Although we have not yet determined what caused this paradox, there might be a myriad of explanations.
BDNF level could be a “biomarker” for effective antidepressant treatment. The activation of BDNF may be one of the final steps in a multistage process of intracellular events initiated by antidepressant treatment. The ability of antidepressants to increase the concentration of neurotransmitters in synapses, and the subsequent alterations in receptors density and function, can be accomplished by a variety of mechanisms, which result in the changes of a wide range of postsynaptic second-messenger systems (Reid and Stewart, 2001) and subsequent adaptive structural and functional changes at the synapse. In particular, this may require changes in the expression, secretion, or downstream functioning of BDNF (Castren, 2004; Duman et al., 1997). This is indicated by the fact that statistically significant increase in BDNF expression after CPG treatment at the dose of 2 mg/kg parallels the most pronounced antidepressant-like response in the TST, while CPG at the dose of 1 mg/kg, that produced less pronounced behavioral effect, also caused slight increase in BDNF mRNA level, but it has not yet reached statistical significance.
It may also be that CPG produces transient increase in BDNF expression/level which triggers changes on intracellular signaling that mediate synaptic plasticity effects, required for the antidepressant effect, suggesting that maintenance of increased BDNF level is not essential, and we observed this effect in a period of its decline. And besides, delayed onset of CPG antidepressant-like effect suggests that neuronal plasticity plays an important role in the mechanism of CPG action. In that context, it might be useful to draw attention to other elements involved in BDNF signaling, including TrkB receptors.
CREB is a transcription factor that regulates BDNF expression and participates in neuronal plasticity and long-term memory formation. The observed increased BDNF gene expression in both CPG and fluoxetine-treated mice did not correlate with any alteration in Creb1 gene expression. As the majority of studies examining alterations in CREB after chronic administration with some antidepressants including fluoxetine report increased CREB mRNA levels in the hippocampus and prefrontal cortex (PFC) (Blendy, 2006), this phenomenon may seem strange at first sight. However, not all antidepressants increase CREB levels and/or activity, and more importantly, the phosphorylation state of CREB rather than its total amount or its mRNA level is more essential for regulating CREB-directed gene transcription (Qi et al., 2008; Sairanen et al., 2007).
It should be noted that one major limitation of this study is using only mRNA expression data, which do not necessarily represent the exact gene profile. The integration of mRNA and protein expression profiles would give more insight into CPG effects.
As for the role of 5-HT, in this study, our hypothesis that antidepressant-like effect of CPG is mediated by serotonergic system was not substantiated. However, we do not exclude such possibility as earlier studies have found that 2-week CPG treatment resulted in a decreased number of 5-HT2AR binding sites in BALB/c mice brain (Abdullina et al., 2019). This discrepancy may be explained either by genotypic differences between BALB/c and ASC mice or by incomplete correlation between mRNA and protein expressions.
It has also been reported that CPG is a metabolite of insulin-like growth factor-1 (IGF-1) (Guan and Gluckman, 2009), which, in addition to the other functions, involved in long-lasting changes in neuronal activity (Malberg and Blendy, 2005; Trejo et al., 2001). It has been established that CPG is able to modulate the activity of IGF-1 by changing the binding of IGF-1 to its binding proteins and thereby increasing the availability of free IGF-1 (Singh-Mallah et al., 2016). Some studies link neurotrophic effects of IGF-1 with BDNF-induced mechanisms since IGF-I was found to increase BDNF in the brain (Carro et al., 2000; Kazanis et al., 2004), enhance expression of TrkB receptors, and increase BDNF responsiveness (McCusker et al., 2006). Furthermore, central administration of IGF-I produced long-lasting antidepressant-like effects in the modified rat-forced swimming test (Hoshaw et al., 2005). However, it was also shown that some neuronal effects of IGF-1 and BDNF are different and mediated by distinct signaling pathways (Yamada et al., 2001). Accordingly, antidepressant-like activity of CPG may be mediated by IGF-1-induced effects, which certainly require further clarification.
Accordingly, molecular targets of CPG are still not clear, and further research is needed to examine the role of BDNF signaling, IGF-1, serotonin, as well as other neurotransmitter systems in the antidepressant-like effects of CPG.
In conclusion, the results of these studies in genetic mouse model of depression indicate that chronic treatment with the dipeptide CPG produced potent antidepressant-like effect comparable with that of SSRI fluoxetine. The revealed activity was accompanied with increase in BDNF gene expression in the cortex, suggesting that CPG’s antidepressant-like effect can be mediated by BDNF signaling. The low effective dose, non-toxicity, and the absence of negative effects on locomotor activity, anxiety-like behavior, or cognitive function make CPG a promising drug for depression treatment.
Supplemental Material
sj-jpg-1-jop-10.1177_02698811211069101 – Supplemental material for The neuropeptide cycloprolylglycine produces antidepressant-like effect and enhances BDNF gene expression in the mice cortex
Supplemental material, sj-jpg-1-jop-10.1177_02698811211069101 for The neuropeptide cycloprolylglycine produces antidepressant-like effect and enhances BDNF gene expression in the mice cortex by Aliya A Abdullina, Ekaterina V Vasileva, Elizabeth A Kulikova, Vladimir S Naumenko, Alexandra V Plyusnina, Tatyana A Gudasheva, Georgy I Kovalev and Sergei B Seredenin in Journal of Psychopharmacology
Footnotes
Acknowledgements
This study was performed within the framework of the State Assignment No. 0521-2019-0009: Analysis of Receptor Mechanisms and Search for Means of Pharmaceutical Protection of the Central Nervous System upon Impaired Brain Circulation and Cognitive Deficit.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.