
Abstract
Background:
Dopamine is implicated in the effort-based control of motivational processes; however, whether tonic dopamine regulates the effort-cost impact on motivation, is still debated.
Aims:
The rats lacking the dopamine transporter (DAT), which have dramatically increased levels of the synaptic dopamine, were used in the present study to elucidate the role of the synaptic dopamine in motivational processes.
Methods:
To study the reward-related processes, the progressive ratio 3 (PR3) operant schedule of food reinforcement (the ratio increases by 3 after each earned reinforcer) was performed in adult male rats (DAT knockouts (DAT-KO), heterozygotes (DAT-HT) and wild-types (DAT-WT)).
Results:
During the PR3 session, the response rate of DAT-KO rats was gradually increased following the augmented required number of responses. In contrast, the local response rate of DAT-WT and DAT-HT decreased. d-Amphetamine sulfate salt (3 mg/kg, i.p.) altered the local response rate dynamics in DAT-WT, which became similar to that of DAT-KO. Interestingly, the reduction in response rate at low effort demands was associated with decreased rate of entries into the magazine tray in DAT-WT rats treated with amphetamine (3 mg/kg) but not in DAT-KO rats.
Conclusions:
Our results suggest that the elevated tonic synaptic dopamine can strongly affect motivation/effort-cost relation in rodents.
Keywords
Introduction
Positively reinforced goal-directed behaviour can be consider as a barter system in which the organism trades its work for the reward (Salamone et al., 2009; Vuchinich and Heather, 2003). Therefore, the instrumental response requirement is the price (cost) that must be paid to obtain the reinforcer. The majority of studies focus on ‘reward’, ‘reward value’ or ‘outcome valuation’. However, there is a growing body of evidence that studying the other side of the equation, the work or cost side, is equally important (Salamone and Correa, 2012; Salamone et al., 2016a, 2016b; Winstanley and Floresco, 2016).
Several papers have evaluated goal-directed behaviour with respect to some effort (Salamone and Correa, 2012; Salamone et al., 2016a, 2016b, 2018; Wassum and Izquierdo, 2015; Winstanley and Floresco, 2016). However, most of such studies are focused on effort-based decision making task, where high-effort experimental option is coupled with a higher reward value/amount (Bardgett et al., 2009; Cousins and Salamone, 1994; Cousins et al., 1996). Therefore, effort cannot be studied in isolation from reward, as the increase in difficulty can be compensated by the increase in reward. The progressive ratio (PR) schedule of operant reinforcement, which implies a gradual increase in the effort required to obtain the reward, is one of the experimental approaches that provides the possibility to exclude this confounding factor (Hodos, 1961). Although the methods of some of the studies mentioned above and others (Randall et al., 2012, 2014, 2015; Rotolo et al., 2019, 2020, 2021; Sommer et al., 2014; Yohn et al., 2016) included this schedule as a high-effort behavioural option, the PR schedule can be a highly informative method to assess motivational processes when used solo. Analysis of the local response rate (LRR) during the PR session provides the opportunity to obtain valuable information about motivation (Killeen et al., 2009; Oakeshott et al., 2012; Sizemore et al., 2003).
Dopamine (DA) is crucial for reward-related processes. DA neurons projections arise predominantly from the midbrain and form mesolimbic and nigrostriatal pathways that are essential for reward-seeking behaviour (Collins and Saunders, 2020). These neurons have two well-studied firing patterns associated with DA release – tonic (‘slow’) and phasic (‘fast’). Although phasic release appears to be more (but not exclusively) responsible for conveying information about potential reward rather than effort, the role of ‘tonic’ DA levels is much less clear (Walton and Bouret, 2019). It has been shown that under the influence of substances that reduce DA neurotransmission, such as systemic administration of DA receptor antagonists, rats show reduced performance in the PR schedule (Aberman et al., 1998). Similarly, administration of the vesicular monoamine transporter-2 inhibitor tetrabenazine, which reduces DA levels, resulted in a reduced response in the effortful PR behavioural option in rats (Randall et al., 2014). Conversely, pretreatment with DA reuptake blockers, such as amphetamine (AMP), cocaine, methylphenidate and other dopamine transporter (DAT) inhibitors, was associated with increased response in the PR schedule (Poncelet et al., 1983) and enhanced selection of high-effort option in effort-based choice tasks (Rotolo et al., 2019, 2020, 2021; Sommer et al., 2014; Yohn et al., 2016). Furthermore, Cagniard et al. (2006) showed that knockdown of DAT in mice resulted in increased choice of lever pressing in a cost-benefit procedure. However, there are some inconsistencies in the work evaluating the effects of DA enhancers. For example, other works reported an opposite or null effect of AMP in the PR schedule (Mobini et al., 2000; Sommer et al., 2014). Indeed, these results are still controversial.
Recently, Bradshaw and Killeen (2012) presented a new approach to the analysis of LRR in the PR schedule based on the mathematical principles of reinforcement (MPR). Accordingly, dynamics of LRR during the PR session are described by the model. This model has four fixed parameters: α (specific activation), δ (response time), T0 (minimum post-reinforcement pause (PRP)) and k (slope of the linear waiting function). The first one estimates the incentive value of the reinforcer. δ reflects the minimum time for a response and therefore the motor ability of the animal. The last two parameters (T0 and k) describe the PRP. So, applying the MPR for the behaviour analysis in PR schedule allows to unravel complex behavioural pattern and collect information about motivational state.
To clarify the role of ‘tonic’ DA levels in effort-cost dependent behaviour, we tested rats with genetically or pharmacologically induced increase of synaptic DA concentration in PR schedule.
Methods
Animals
All experiments were performed on the previously described DAT knockout (DAT-KO) rats (Leo et al., 2018). Drug- and experimentally naive male DAT-KO rats (n = 26) as well as their heterozygous (DAT-HT, n = 22), and wild-type (DAT-WT, n = 26) littermates were obtained by heterozygous crossing (2–3 months old and weighing 200–300 g at the beginning of the experiment) from the local colony of the Department of Psychopharmacology (Valdman Institute of Pharmacology, St. Petersburg, Russia). Animals were individually housed in TIIIH cages (Tecniplast, Buguggiate, Italy) with wood-based bedding (Lignocel, BK 8-15, JRS GMBH+Co., Rosenberg, Germany) under a 12 h/12 h light/dark cycle (lights on at 08:00 h) at 21°C ± 2°C and 50 ± 20% humidity. The animals had free access to filtered tap water (Filter AQUAPHOR®, St. Petersburg, Russia). Cages, bedding and water bottles were changed once a week. We restricted the animals’ food consumption (standard laboratory chow, formula ПК 120-1, Laboratorkorm, Moscow, Russia) before the start of the experiments for about 1 week so that their body weight decreased to approximately 85% of the initial one. During the experiments, the amount of food per day (15–16 g for WT and HT rats and 20 g for DAT-KO rats) was adjusted to limit body weight gain by 2–3 g/week. Pharmacological challenge with AMP was performed in a group of DAT-WT rats (n = 12).
Apparatus
The experiments were performed in six standard modular operant conditioning chambers for rats (interior dimensions: 30.5 cm × 24.1 cm × 29.2 cm; ENV-007, MED Associates Inc., East Fairfeld, VT, USA) placed in ventilated, light-proof and sound-attenuated boxes with an electric fan providing air circulation and a background white noise. A pellet tray (ENV-200R2MA) was set up at 2 cm from the floor on the middle panel of the right (five chambers) or the left (one chamber) wall. The tray was equipped with a pair of nose-poke photobeam infrared sensors (ENV254-CB). A pellet dispenser (ENV-203-45) was located outside of the chambers and delivered 45-mg food pellets (F0165, P.J. Noyes Inc., Lancaster, NH, USA) into the tray. A speaker (ENV-223AM) was situated under the tray, and a white house light (ENV-215M) was placed at the top of the middle panel. Retractable levers (model ENV-112BM) were placed on the sides of the food tray.
Behavioural procedure
All experiments were performed during the light period of the light-dark cycle. The experimental protocol was approved by the local animal care and use committee of the Pavlov First St. Petersburg State Medical University.
Habituation to food pellets
Approximately 48 and 24 h before the operant chamber test, each rat received 15 food pellets in the home cage in addition to lab chow to habituate them to pellet consumption. When the animal ate all the pellets during the night (from 8 pm to 8 am), we started the training in the operant boxes.
Magazine training
Rats were trained to consume food pellets from the magazine tray after a sound signal. During each of the three daily sessions, 28 pellets were individually delivered to the food tray in a variable-time 70-s schedule. Each food pellet delivery was paired with a sound stimulus.
Lever press training
First, the rats were subjected to a Pavlovian/instrumental autoshaping procedure to facilitate lever-press training. During the three daily sessions, rats received forty 8-second lever presentations (left or right levers were counterbalanced between subjects) followed by pellet delivery accompanied by the sound stimulus. However, if a rat pressed the lever, it was withdrawn, and the pellet was delivered immediately.
In the following sessions (maximum of 120 min), the animals had continuous access to the lever. Each lever press was reinforced by a food pellet delivery (fixed ratio 1, FR1). The ratio was then increased to 3 (FR3) and 5 (FR5). Rats progressed to the next FR level and on to the PR schedule when they met the criterion of receiving 50 reinforcers within 60 min during 2 consecutive sessions.
PR schedule
To ensure a linear increase in effort, we used the PR3 schedule, that is, the ratio increased by 3 after each reinforcer obtained (3, 6, 9 and so on). PR3 sessions lasted 120 min. The following parameters were measured: (1) number of reinforcers earned; (2) inter-reinforcement time (IRT); (3) number of magazine tray entries (ME); (4) PRP. LRR and local ME rate were calculated for each inter-reinforcement interval as ratio/IRT and ME/IRT respectively.
Drugs
d-Amphetamine (Sigma-Aldrich, St. Louis, MO, USA; 0, 0.3, 1 and 3 mg/kg) was dissolved in saline (0.9% NaCl) and administered intraperitoneally (i.p.) 30 min before the beginning of the PR3 session. A Latin square design was used for the pharmacological challenge, with a minimum 72 h washout period between tests.
Data analysis
Statistical data analysis and visualization was performed in the R software environment (R Core Team, 2023).
LRR and local ME rate under different effort demands (response/reinforcement ratio) were estimated by mixed model analysis of variance (mixed ANOVA) with the Satterthwaite method for degrees of freedom using the ‘lmerTest’ package (Kuznetsova et al., 2017) (version 3.1.3). As rats earned different numbers of reinforcers, we set a maximum ratio for which at least two values of the dependent variable remained in each experimental group. Data for ratio values exceeding this threshold were excluded from the analysis. Then, all ratio values were divided into blocks of 5 units (e.g. block 1 contains ratios 3, 6, 9, 12 and 15; block 2 contains ratios 18, 21, 24, 27 and 30; etc.). For each rat, the means of the variables analysed within each block were calculated and subjected to the following analysis. For further statistical analysis, all dependent variables were ranked (with average rank used for identical values). Follow-up tests for significant main effects or interactions were performed using the emmeans package (version 1.7.4.1). Dunnett’s and Tukey’s methods for adjusting p-values and Kenward-Roger method for degrees of freedom were used for multiple comparisons.
In addition, the PR performance mathematical model (Bradshaw and Killeen, 2012) was fitted to the data of each rat using MS Excel Solver add-in program. Then, the values of fitted parameters were subjected to statistical analysis.
Non-parametric statistical methods (Kruskal-Wallis and Friedman tests) were used to compare number of earned reinforcers and the model parameters. The Dunn’s test (‘dunn.test’ package, version 1.3.5) or the exact non-parametric many-to-one comparison test (EMC test) (Eisinga et al., 2017) were performed when the above statistical methods indicated significant results. Alpha was set at 0.05.
Results
DAT-KO rats
The number of earned reinforcers did not differ between the three genotypes: DAT-WT/HT and DAT-KO rats performed similarly (Kruskal-Wallis test: H = 1.456, df = 2, p = 0.48; Figure 1). In contrast, LRR was significantly influenced by genotype (factor ‘genotype’: F(2, 74) = 4.890, p < 0.05; factor ‘block’: F(9, 498) = 14.907, p < 0.001; interaction: F(18, 498) = 30.478, p < 0.001). While the LRR of DAT-WT and DAT-HT rats decreased with increasing work demands, DAT-KO rats showed the opposite pattern with lower LRR at low ratio demands and increased responding at high ratio demands (Figure 2).
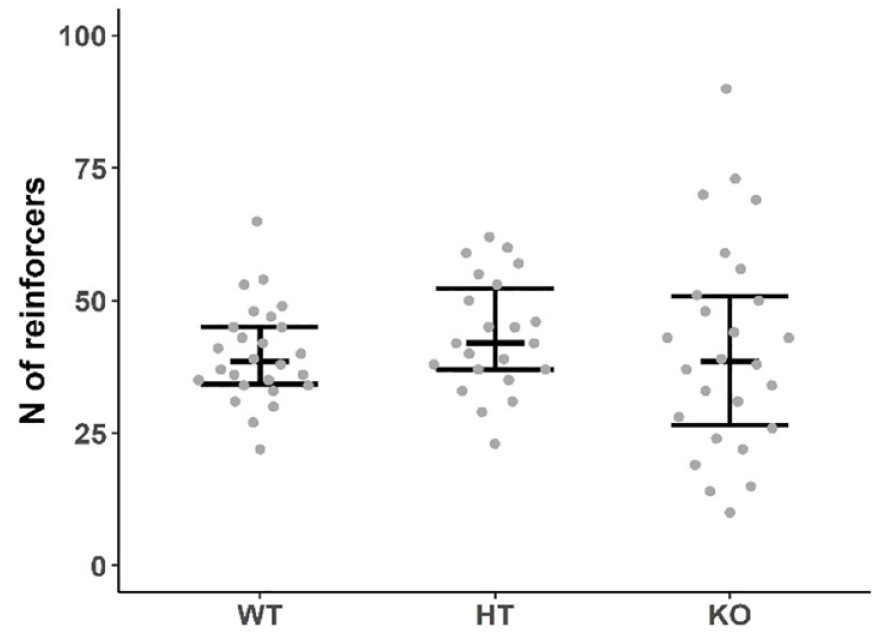
The number of reinforcers earned by DAT-WT, DAT-HT and DAT-KO rats under PR3 schedule. n = 26 (WT), 22 (HT), 26 (KO). Data are presented as median, first and third quartiles and individual values.
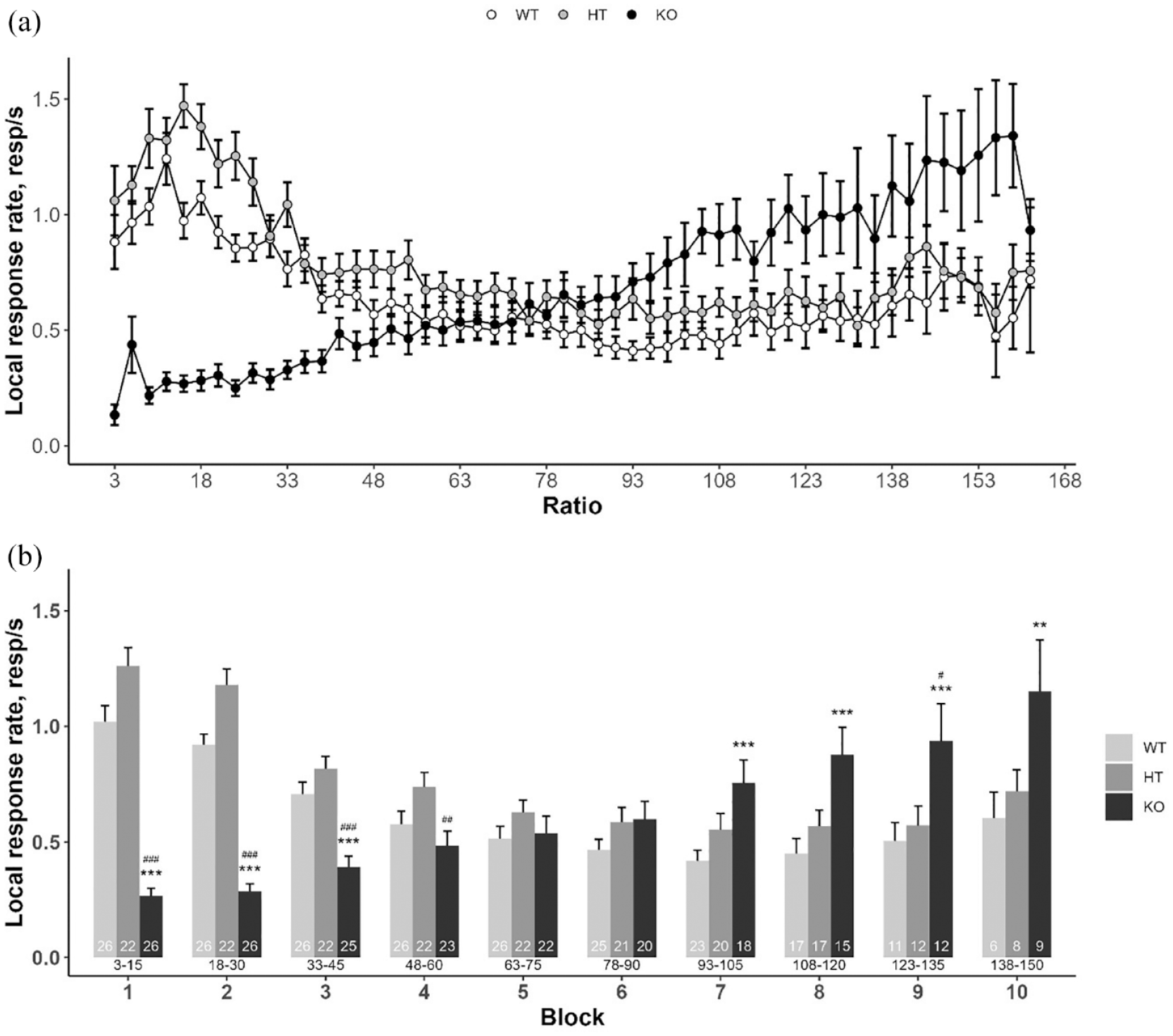
Effect of DAT gene knockout on local rate of responding under PR3 schedule. (a) LRR of DAT-WT, DAT-HT and DAT-KO rats; (b) results of LRR analysis by blocks of five reinforcers. Data are presented as mean ± SEM. **p < 0.01, ***p < 0.001, DAT-KO versus DAT-WT, Tukey’s test; #p < 0.05, ##p < 0.01, ###p < 0.001, DAT-KO versus DAT-HT, Tukey’s test. The number of observations is indicated into the bars. Current response/reinforcement ratio values (R) are under the bars.
PR model was successfully fitted to individual values of all rats except two DAT-KO animals. Statistically significant differences were found between the experimental groups in all parameters of the PR model (Kruskal-Wallis test – α: H = 21.9, df = 2, p < 0.001; δ: H = 22.8, df = 2, p < 0.001; k: H = 20.3, df = 2, p < 0.001; T0: H = 28.0, df = 2, p < 0.001). Post hoc comparisons revealed higher values of the α, δ and Т0 and a lower value of the parameter k in KO rats compared to WT and HT ones (Figure 3). There was also a trend toward a lower value of the δ in heterozygous rats compared to wild-type animals (p < 0.06).
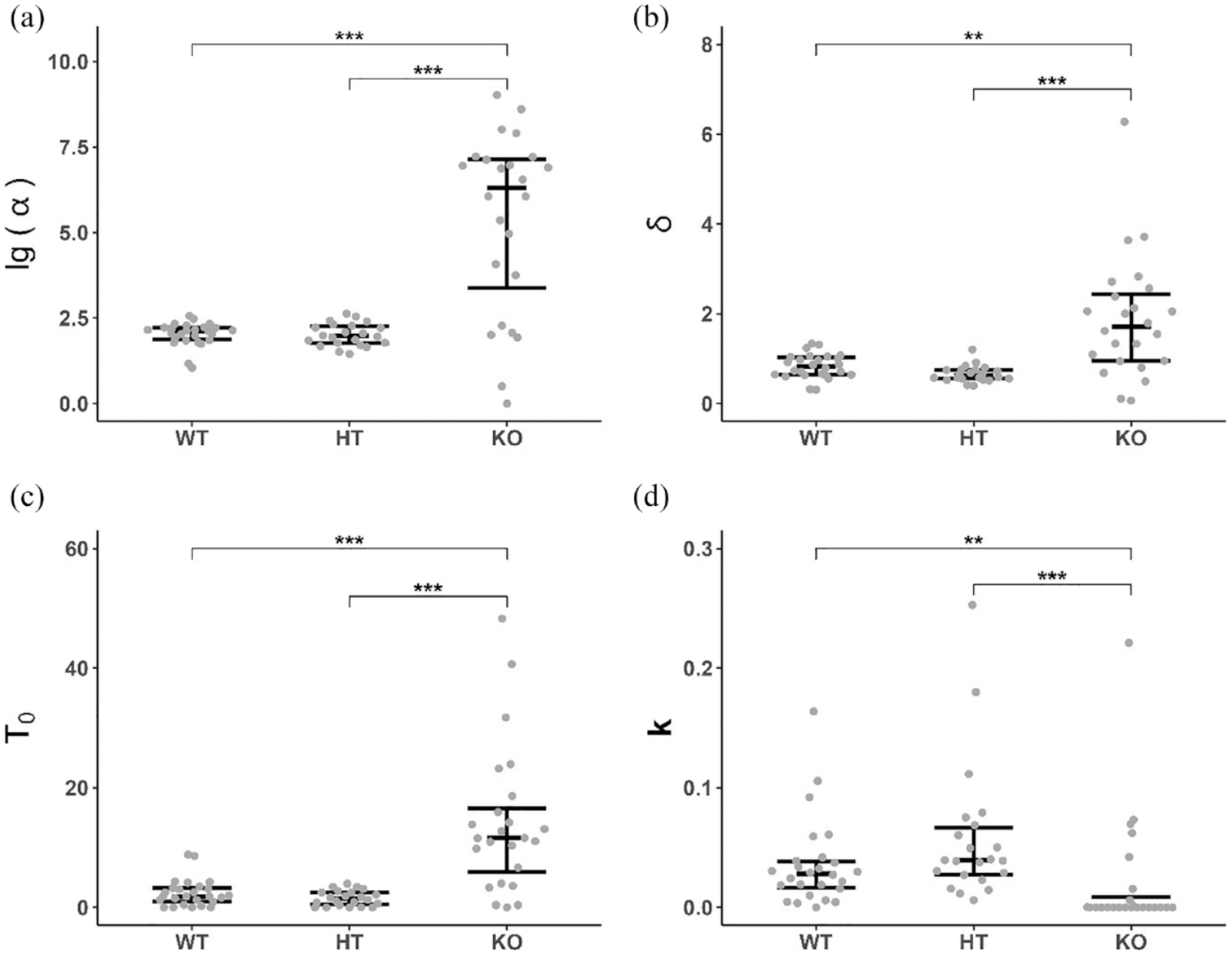
Effect of DAT gene knockout on PR model parameters. (a) α (specific activation, decimal logarithm of the initial value); (b) δ (response time); (c) Т0 (minimum PRP); (d) k (slope of the linear waiting function). n = 26 (WT), 22 (HT), 24 (KO). Data are presented as median, first and third quartiles and individual values. **p < 0.01, ***p < 0.001, Dunn’s test.
ANOVA detected significant effect of factors ‘block’ (F(2, 31) = 8.399, p < 0.01) and ‘genotype’ (F(7, 200) = 14.862, p < 0.001) as well as their interaction (F(14, 200) = 4.428, p < 0.001) on local ME rate. Post hoc comparisons revealed higher local ME rate in DAT-KO rats compared to WTs and HTs in blocks 3–8 (Figure 4).
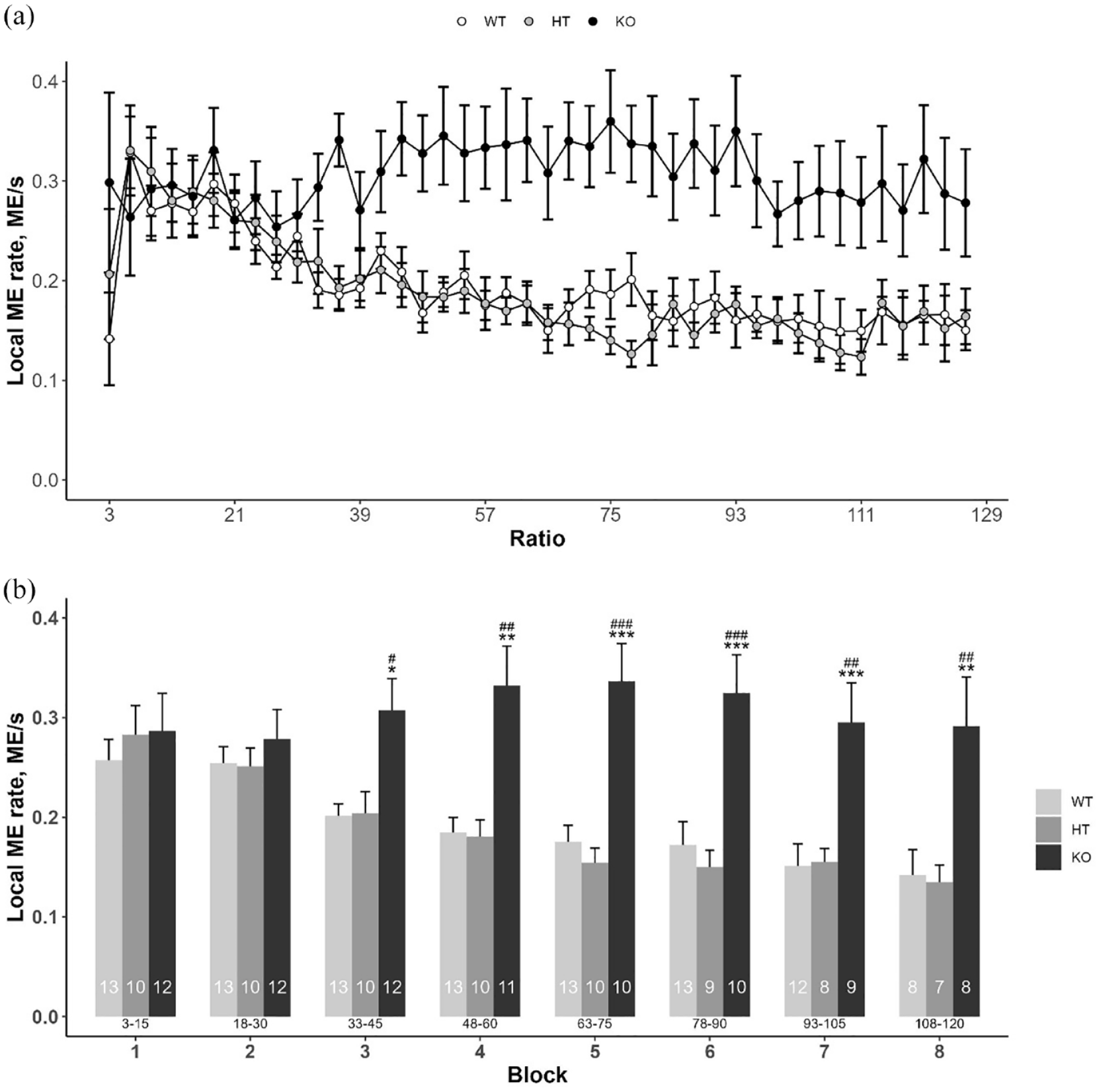
Effect of DAT gene knockout on rate of magazine entries under PR3 schedule. (a) Local ME rate of DAT-WT, DAT-HT and DAT-KO rats; (b) results of local ME rate analysis by blocks of five reinforcers. Data are presented as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001, DAT-KO versus DAT-WT, Tukey’s test; #p < 0.05, ##p < 0.01, ###p < 0.001, DAT-KO versus DAT-HT, Tukey’s test. The number of observations is indicated into the bars. Current response/reinforcement ratio values (R) are under the bars.
AMP treatment
AMP administration significantly affected the number of reinforcers earned by WT rats (Friedman’s test: F = 13.7, df = 3, p < 0.01), contrary to the effect reported for DAT-KO rats. However, after pairwise comparisons, we found that treatment with only a moderate AMP dose (1 mg/kg) increased the number of earned reinforcers (1 mg/kg vs vehicle p < 0.05, Figure 5). Further statistical analysis of the LRR revealed significant effects of both factors (‘dose’ and ‘block’) as well as their interaction (factor ‘dose’: F(3, 33) = 7.254, p < 0.001; factor ‘block’: F(5, 193) = 22.214, p < 0.001; interaction: F(15, 193) = 9.727, p < 0.001). Pairwise comparisons (EMC test) showed that after administration of the lowest dose of AMP (0.3 mg/kg), the LRR was higher at comparatively low ratio requirements (18–45) within blocks 2 and 3 (Figure 6). Conversely, pretreatment with higher AMP doses (1 and 3 mg/kg) induced an increased LRR at high-effort demands (fifth and sixth block). Interestingly, administration of the highest AMP dose (3 mg/kg) also decreased LRR at the beginning of the PR session when the ratio is minimal (3–30, Figure 6).
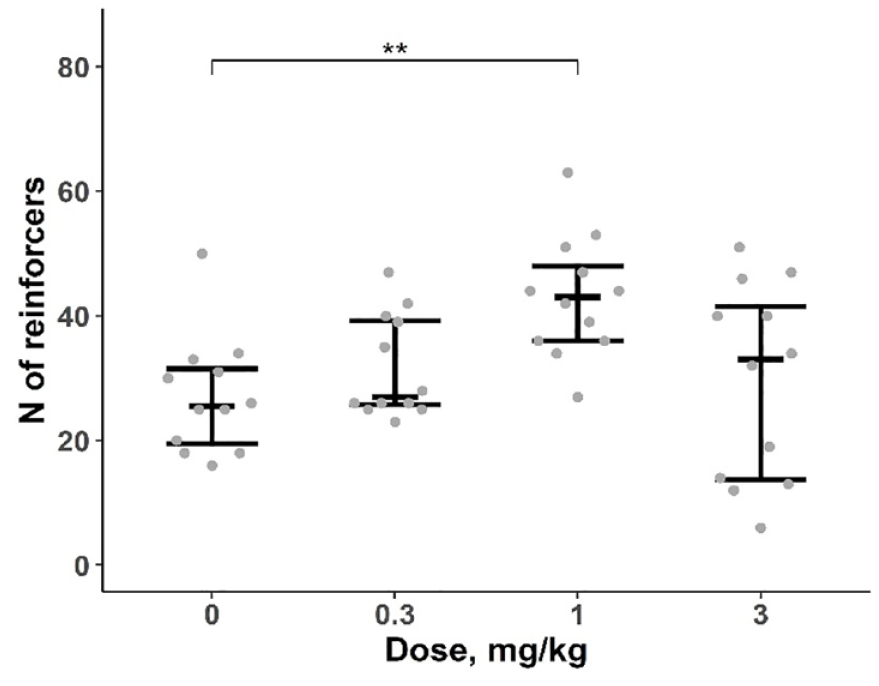
Number of reinforcers earned by DAT-WT rats received AMP injection under PR3 schedule. n = 12. Data are presented as median, first and third quartiles and individual values. **p < 0.01, AMP versus saline, EMC test.
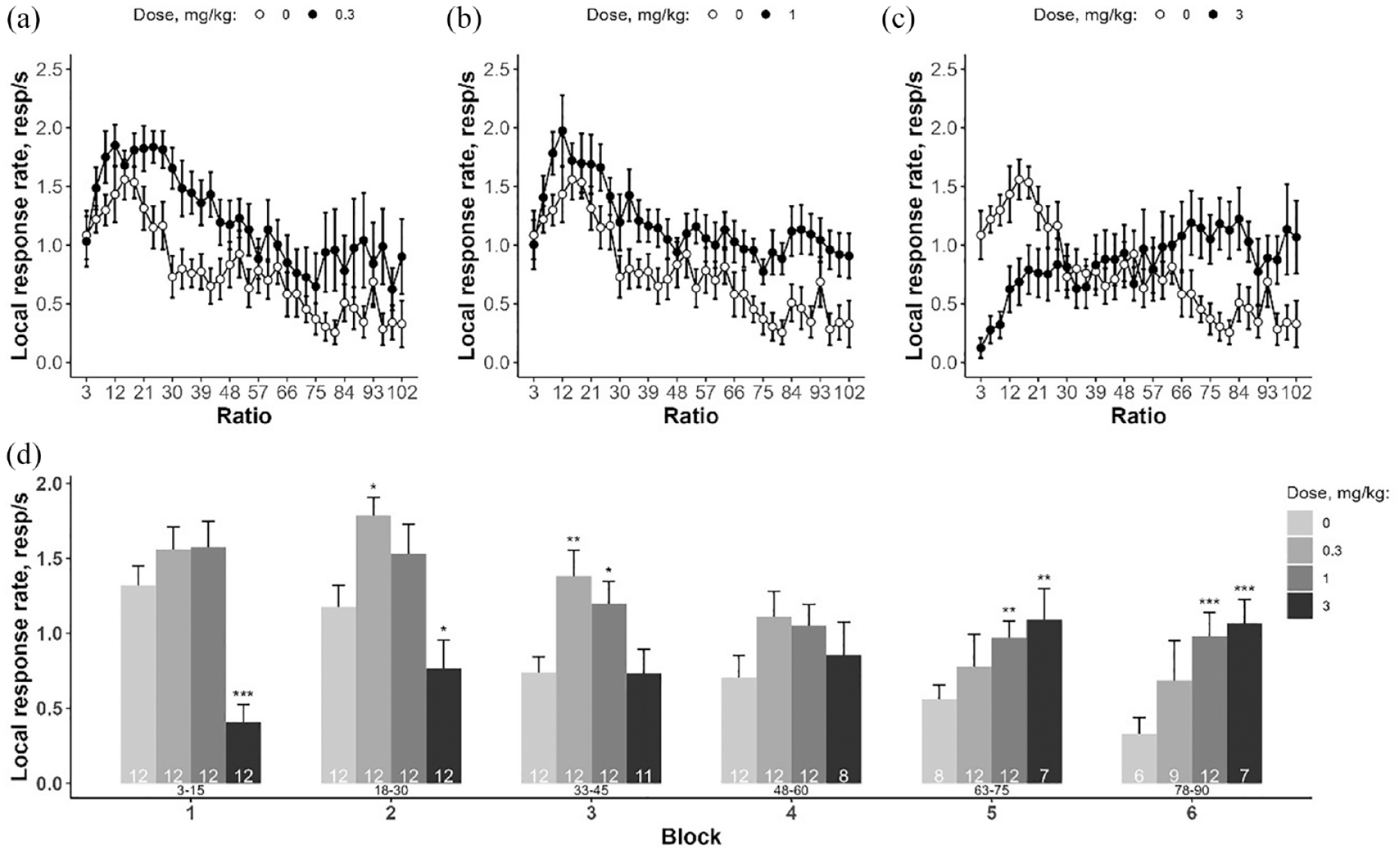
LRR of DAT-WT rats received AMP injection under PR3 schedule. (a–c) Effect of different doses of AMP treatment; (d) results of LRR analysis by blocks of five reinforcers. Data are presented as mean ± SEM. Number of observations is pointed within bars. Current response/reinforcement ratio values (R) are located under the bars. *p < 0.05, **p < 0.01, ***p < 0.001, Dunnett’s test (AMP vs saline).
LRR changes were reflected in the results of mathematical model of PR performance analysis (Friedman test – α: χ2 = 21.4, df = 3, p < 0.001; δ: χ2 = 19.9, df = 3, p < 0.001; T0: χ2 = 19.7, df = 3, p < 0.001; k: χ2 = 20.7, df = 3, p < 0.001). Post hoc (EMC test) comparisons detected higher values of the α and Т0 after treatment with AMP (3 mg/kg) and a lower value of k following AMP injection (1 and 3 mg/kg; Figure 7) compared to vehicle control. There was also a tendency to decrease of δ after pretreatment with 0.3 mg/kg (p < 0.1) and increase after pretreatment with 3 mg/kg of AMP (p < 0.07).
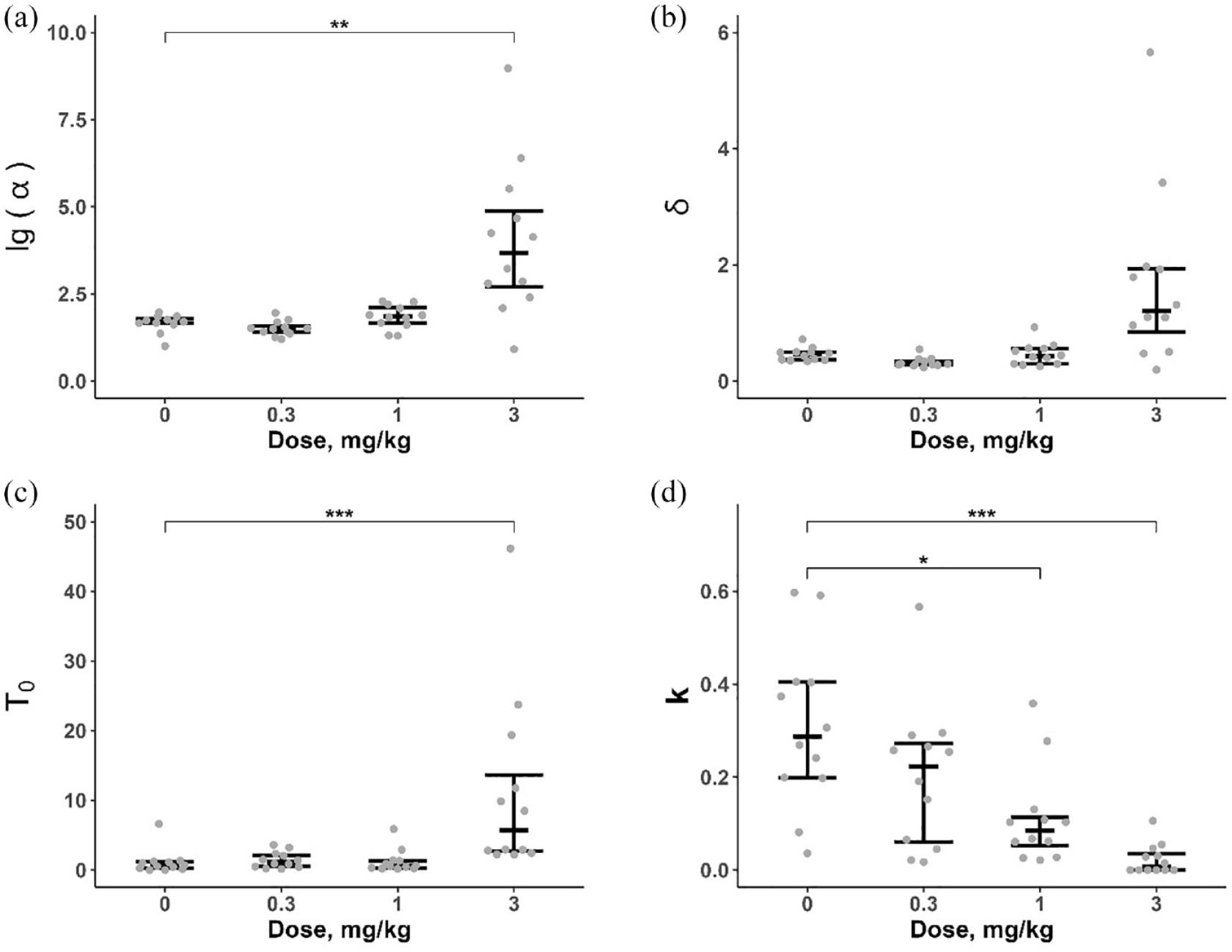
Effect of AMP on PR model parameters. (a) α (specific activation, decimal logarithm of the initial value); (b) δ (response time); (c) Т0 (minimum PRP); (d) k (slope of the linear waiting function). n = 12. Data are presented as median, first and third quartiles and individual values. *p < 0.05, **p < 0.01, ***p < 0.001, AMP versus saline, EMC test.
AMP administration also significantly affected local ME rate (factor ‘dose’: F(3, 33) = 6.265, p < 0.01; factor ‘block’: F(5, 193) = 86.700, p < 0.001; interaction: F(15, 193) = 8.194, p < 0.001). However, in contrast to DAT-KO rats AMP administration at the highest dose (3 mg/kg) was also associated with decreased local ME rate in block 1 (Figure 8).
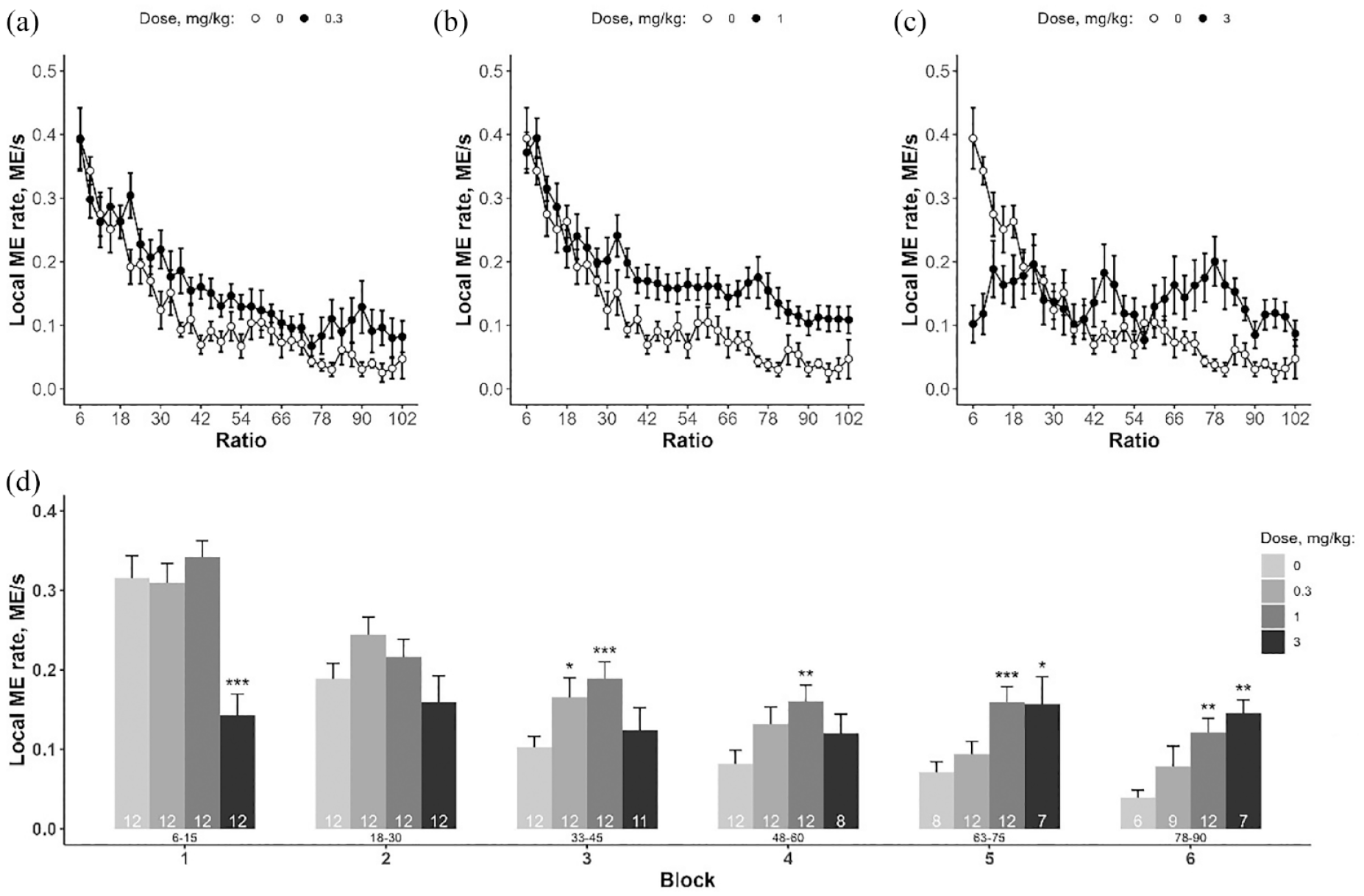
Effect of AMP on local rate of magazine entries under PR3 schedule. (a–c) Effect of different doses of AMP treatment; (d) results of local ME rate analysis by blocks of five reinforcers. Data are presented as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001, AMP versus saline, Dunnett’s test. Number of observations is pointed within bars. Current response/reinforcement ratio values (R) are located under the bars.
Discussion
We found a strong relationship between effort demands and operant performance in the PR3 schedule: the response rate is maximal at low values of ratio demand, but as the number of lever presses required increased, the animals’ performance decreased. However, we observed that under conditions of dramatically increased DA transmission (such as genetic DAT disruption or under the effect of the DA releaser AMP at a dose of 3 mg/kg), this pattern is reversed. Motivational changes were confirmed by the mathematical model of PR schedule performance. Thus, we can conclude that ‘tonic’ DA is associated with a relative preference for goal-directed activity as a function of effort-cost.
DAT functioning disruption is accompanied by reverse PR performance
DAT-KO rats showed reversed performance on the PR3 schedule. This result is consistent with the previously reported effect of cocaine, a potent DAT blocker, under the PR schedule (Sizemore et al., 2003). However, the amplitude of cocaine effect appears to be less considerable than the genetic DAT inactivation. This effect may be due to the more pronounced hyperdopaminergia in DAT-KO rats. Cagniard and colleagues also reported an increase in the breakpoint in the PR3 schedule in DAT knockdown (DAT-KD) mice, which have less enhanced DA tone (Cagniard et al., 2006), but LRR was not measured in this study. In contrast, in our study DAT-KO and WT rats earned a comparable number of reinforcers, which could be related to different criteria of termination of the PR session as well as neurochemical differences between DAT-KD mice and DAT-KO rats.
Unexpectedly, DAT-HT animals did not differ from WT littermates in PR behavior, although these animals have a twofold increase in synaptic DA concentration (Leo et al., 2018). One possible reason is a compensatory mechanisms in DA neurotransmission, such as DA receptor internalization (Dumartin et al., 2000), which could counteract the moderate DA increase. However, the observed tendency to δ lower value in DAT-HT rats can be interpreted as some motor facilitation of operant responding. Several papers have shown differences in locomotion of DAT-HT versus -WT littermates (Cristiana et al., 2020; Manoni et al., 2022; Pardo et al., 2022), so further studies are needed to better investigate the behaviour of heterozygous rats.
AMP effects in PR schedule
AMP effects were qualitatively different depending on the dose. AMP administration at doses of 0.3 and 1 mg/kg did not alter the overall relationship between work demands and behavioural performance but increased responding under different ratios (blocks 2, 3 – 0.3 mg/kg; blocks 3, 5, 6 – 1 mg/kg). Pretreatment with 3 mg/kg AMP resulted in an inverted performance similar to that observed in DAT-KO rats. Administration of 1 mg/kg AMP was also associated with an increase of the number of earned reinforcers, which contradicts some other works showing an opposite or absent effect of AMP (Mobini et al., 2000; Schulze and Paule, 1990; Sommer et al., 2014). These discrepancies could be dependent on differences in methods (different PR schedules and criteria for termination of PR sessions), animals (different rat strains and stocks as well as differences between local colonies), drugs (different sources and quality), etc. Our results highlight the advantage of using the LRR analysis, which can be more informative than the simple evaluation of the breakpoint. Indeed, in our study, only one of the three doses of AMP resulted in an increase of the number of earned reinforcers, whereas changes in LRR were found for all the doses.
The analysis of the PR model revealed a significant decrease in the parameter k after AMP 1 mg/kg administration. This result suggests a slowing of the typical PRP increment with the progression of ratio requirement, which is a typical pattern and probably reflects the rats’ aversion of responding with an increasing IRT (Killeen et al., 2009). Pretreatment with a smallest dose of AMP (0.3 mg/kg) resulted in a trend of δ decrease. Finally, administration of the highest AMP dose (3 mg/kg) was associated with a robust change of all the parameters evaluated (increased α, δ and T0 and decreased k). Thus, the underlying basis of AMP-induced changes of PR performance is also highly dependent on its dose: at low dose (0.3 mg/kg), the increase in LRR is probably due to motor effects of the drug, at medium dose (1 mg/kg), it is due to a change in the sensitivity to the increased IRT. Eventually the highest AMP dose (3 mg/kg) induced a reverse performance in consequence of a combined effect on the incentive value of reinforcer, motor capabilities and PRP.
In rats’ plasma, the half-life of AMP is approximately 60 min (Honecker and Coper, 1975; Hutchaleelaha et al., 1994), which can potentially introduce a confounding factor. For instance, when rats treated with 3 mg/kg of AMP are at the start of the session, they may take longer to complete the required ratio. As the ratio increases, the concentration of the drug may decrease more significantly compared to lower doses. Thus, the observed increase in LRR under high ratio requirements, similar to that observed after administering 1 mg/kg of AMP, may be attributed to a comparable amount of active drug in the organism. However, the results obtained in DAT-KO rats suggest that high DA level is related to similar LRR changes, so it is unlikely that higher blood AMP concentration would lead to different results. Besides, the AMP elimination from brain tissues can be a slower process compared to the one in plasma. Between 0.5 and 4 h post i.p. injection, AMP concentrations in different brain areas are about 7–10 times higher compared to plasma (Honecker and Coper, 1975).
Possible processes underlying changes in PR performance
The AMP effects here reported, provide further evidence that AMP affects instrumental responding in a rate-dependent manner (Heffner et al., 1974): low response rates are increased and high response rates are decreased under different operant schedules. However, the possible underlying behavioural and neurochemical processes remain poorly understood. Our results from DAT-KO rats in the PR schedule, suggest that DA excess in the synaptic cleft may be responsible for the rate-dependent phenomenon. It is also worth noting that, DAT-KO rats do not have a baseline response rate, so the amount of effort required most probably contributes to the level of responding in the hyperdopaminergic state.
While the increase in rate response is obviously due to increased responding activity, the decrease in lever pressing rate may reflect quite different behavioural effects of AMP. In particular, AMP and other substances that enhance DA signalling are known to increase locomotor activity and induce stereotyped behaviours such as licking, biting, gnawing, rapid head movements and so on. Thus, such effects may be responsible for the decrease in operant responding. In addition, Heffner and colleagues reported stereotyped behaviour following AMP treatment at 3 mg/kg, which was the highest dose in our study. Results of mathematical model analysis suggest that the decrease of LRR at the low ratios are related with both motor disturbances and increased PRP. Interestingly, decreased performance under low effort demands was linked to a decrease in food tray checking activity only in WT rats treated with AMP. This finding indicates that the well-known anorexigenic effect of AMP can be one of the reasons for the decreased PR performance. Remarkably, it was demonstrated that AMP, in contrast to serotoninergic drug fenfluramine, produces delay in the onset of eating in food-deprived rats (Blundell et al., 1976). AMP is also known to affect norepinephrine reuptake, so this fact can be responsible for the different effect of AMP compared to genetic DAT silencing.
The motivational nature of increased LRR under high-effort demands is substantiated by the heightened magazine checking activity. This finding contradicts the potential explanation that rats with elevated DA levels exhibit higher LRR solely due to the inability to cease lever pressing, caused by impaired inhibitory processes. DAT-KO animals as well as AMP-treated rats responded with higher frequency at the food tray, so they were clearly able to break a train of responses to move to the food tray.
PR schedules are widely used to study motivational processes as they allow the assessment of animal behaviour under conditions of increased effort. However, factors other than the amount of effort required may be responsible for the behavioural outcome. One such confounder could be temporal changes. It has been shown that the amplitude of DA release increases during both the PR session and the control session, where the reward is delivered after time intervals similar to those in the PR schedule and independent of the animal’s behaviour (Wanat et al., 2010). Thus, the behavioural differences associated with increased DA signalling may reflect different temporal behavioural strategies during reward anticipation at low and high time intervals. However, several studies have shown that DA-related changes in effortful choice cannot be explained simply by an increase in time to reward or a change in reward rate (Floresco et al., 2008; Mingote et al., 2005). Previously, Ostlund and colleagues reported that the extracellular concentration of DA is inversely proportional to the amount of effort required to obtain reinforcer; in this study, the order of high- and low-cost trials was indeed randomized (Ostlund et al., 2011). Analysis of PR model allows to separate time- (parameter k – slope of linear waiting function) and reward-dependent (parameter α – specific activation by reinforcer) changes in PR schedule. Nevertheless, the intermittent nature of reinforcement delivery may be an important factor influencing behaviour in PR schedules. Further studies are needed to disentangle effort-dependent and time-dependent processes.
Despite the clear difference between DAT-KO/AMP-treated rats and control animals it is worth to note that response rate of control rats was stabilized under high ratio values, especially in not treated WT and HT rats (blocks 7–10). This stabilization could be related with DA-independent factors. Since we used arithmetic progression, it can be the case when the difference between high ratios becomes less significant for an animal, according to the Weber–Fechner Law, which states that the ability of a sensory system to discriminate between two physical quantities decreases in inverse proportion to the magnitude of the quantities being compared (Perna et al., 2019). Another possible factor here is steadily decreasing number of animals reaching high ratio values so mean value can be biased.
Conclusion
Taken together, our findings demonstrate that enhancing ‘tonic’ DA transmission dramatically affects motivation/effort-cost relation in rodents. According to analysis of LRR, more effortful conditions seem to become more preferable than less effortful ones. However, our knowledge of these phenomena is limited by poor understanding of the underlying molecular mechanisms of the observed behavioural findings and further studies of effort-related processes are required.
Limitations
The data presented in this paper have been derived exclusively from research using male rats as the experimental animal model, and therefore our results must be interpreted without making biological generalisations.
Footnotes
Acknowledgements
The authors thank Dr Anton Bespalov for helpful suggestions; Marina Semina and Alexandra Gavrilova for maintaining the colony of DAT-KO rats; Dr Anna Ulitina for genotyping the animals.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.