
Abstract
Background:
Dysregulated ventral striatum function has been proposed as one important process occurring in individuals with substance use disorder. This study investigates the role of altered reward and loss anticipation, which is an important component of impaired decision-making, impulsivity, and vulnerability to relapse in individuals with amphetamine use disorder (AMP).
Aims:
To determine whether AMP is associated with blunted striatum, prefrontal cortex, and insula signals during win and loss anticipation.
Methods:
Participants with and without AMP (AMP+ n = 46, AMP− n = 90) from the Tulsa 1000 study completed a monetary incentive delay (MID) task during functional magnetic resonance imaging.
Results:
Group main effects indicated that: (1) AMP+ exhibited lower bilateral caudate/putamen and left nucleus accumbens signal than AMP− across anticipation of wins and losses; and (2) AMP+ showed slower reaction times than AMP− during loss anticipation. Group*condition interactions demonstrated that AMP+ exhibited greater right amygdala signal than AMP− while anticipating large wins, a pattern that reversed when anticipating small losses. Left caudate/putamen attenuations in AMP+ during small loss anticipation were also evident. Groups did not differ in prefrontal or insula signals.
Conclusions:
AMP+ individuals have altered neural processing and response patterns during reward and loss anticipation, potentially reflecting impairments in dopamine function, which may influence their decision-making and reactions to different win/loss scenarios. These findings help to explain why AMP+ have difficulty with decision-making and exhibit a heightened focus on immediate rewards or punishments.
Keywords
Introduction
Methamphetamine continues to be a public health concern in the United States, with an estimated 2.4 million adults aged 26 or older reporting past year use (Substance Abuse and Mental Health Services Administration, 2021). Of those, an estimated 1.4 million adults met criteria for methamphetamine use disorder in the past year, highlighting the association between stimulant use and adverse psychosocial, cognitive, and physical health effects. Although methamphetamine use in the U.S. persists, the effectiveness of evidence-based treatments offered for substance use disorders (SUD) is moderate at best, with estimates showing 50–60% of treatment-seeking individuals relapsing within the first year (Brecht and Herbeck, 2014). Therefore, investigation of the neural mechanisms that go awry in SUD is essential to inform the development and testing of effective treatments to reduce use and improve treatment outcomes.
Several theories have been proposed to explain the development and persistence of addictive behaviors, including SUD. One such theory is the reward deficiency hypothesis of addiction (Blum et al., 1996). In brief, this theory posits that a general deficit in reward responsivity exists among individuals with addictive behaviors, resulting in chronic under-recruitment of associated brain regions (i.e., striatum, prefrontal cortex [PFC]) in response to primary incentives (e.g., sex, food). This process leads individuals to seek out stronger rewards (e.g., illicit drugs) to compensate for this deficit and stimulate reward regions. Relatedly, the incentive sensitization theory (Robinson and Berridge, 2008) suggests that individuals with SUD demonstrate hypoactivation to primary incentives, compared to substance use-relevant, rewards due to the attribution of higher salience to substance-related cues. Taken together, these theories suggest that individuals with SUD should demonstrate hypoactivation in reward-relevant brain regions (e.g., striatum, PFC) in response to non-substance-related rewards (e.g., money).
Examination of the anticipation of reward or loss within SUD is essential, as alterations in the underlying neural mechanisms may play a key role in the perpetuation of SUD. More specifically, an exaggerated anticipatory response to substance cues may lead to heightened levels of motivation and emotion, but devaluation of nondrug rewards such as money; this valuation imbalance may result in maladaptive behavioral responses (e.g., substance seeking), further promoting the substance use cycle (Balodis and Potenza, 2015; Nestor and Ersche, 2022; Tanabe et al., 2019; Volkow et al., 2006). Given the potential role of reward processing in SUD development and maintenance, several studies have examined alterations in reward processing among individuals with SUD using the well-established monetary incentive delay (MID) task (Knutson et al., 2000). For example, individuals with an alcohol use disorder who recently underwent detoxification exhibited ventral striatum hypoactivation compared to healthy individuals when anticipating reward (Beck et al., 2009). Similarly, individuals with a cocaine use disorder have shown hypoactivation in the dorsolateral PFC and posterior cingulate cortex during reward anticipation (Yip et al., 2016). A recent meta-analysis of gain and loss processing in addiction revealed a pattern of hypoactivation within striatum, PFC, and insular cortex (a region involved in interoceptive awareness, salience, and emotional arousal; Khalsa et al., 2009; Singer et al., 2009; Uddin, 2014) during anticipation of nondrug rewards across SUD samples (e.g., alcohol, methamphetamine, cocaine, etc.) compared to individuals with no problematic substance use history (Qiu and Wang, 2021). Overall, previous findings generally demonstrate a pattern of hypoactivation in reward and frontal brain regions among individuals with various SUD types when anticipating reward and loss.
Despite the prevalence of studies investigating altered reward processing in abstinent individuals with alcohol, cocaine, and nicotine use disorders, a limited number of studies have focused on gain and loss anticipation among individuals with amphetamine use disorder (AMP+). In fact, the abovementioned meta-analysis of reward processing in SUD populations included 24 studies, only one of which included a sample of AMP+ individuals (Qiu and Wang, 2021). This study revealed a pattern of hypoactivation within ventral striatum and caudate during loss, but not reward, anticipation among 17 AMP+ males (compared to non-using males) using a modified MID task (Bischoff-Grethe et al., 2017). Importantly, within this sample, AMP+ reported higher depressive symptoms than the comparison group, leaving the potential influence of mood symptoms on the observed findings unclear. The acute administration of methamphetamine to healthy individuals has also recently been shown to increase activity within ventral striatum and anterior insula during loss anticipation (Crane et al., 2023; Molla et al., 2023). These findings suggest that amphetamine type stimulants and the associated use disorder alter neural reward responsivity and may have distinct effects on the processing of positive and negative stimuli. Further research examining the impact of AMP on reward-related brain regions is warranted to better understand how altered reward processing promotes continued maladaptive substance use despite aversive outcomes.
The present study compared reward and loss expectancy (i.e., anticipation) between individuals with and without amphetamine use disorder (AMP+, AMP−) using a MID task during functional magnetic resonance imaging (fMRI). This examination sought to replicate and expand on the existing literature regarding reward processing in individuals with SUD by: (1) focusing on reward process among AMP+ individuals given the limited number of studies utilizing the MID task within this population; (2) utilizing a larger sample of male and female AMP+ individuals (n = 46) than previous examinations (Bischoff-Grethe et al., 2017); and (3) reducing the impact of depression as a potential confound by including AMP− with major depressive disorders (MDD). The MID task is commonly used to investigate reward mechanisms as it allows for the examination of brain activity during both anticipation and receipt of monetary reward and loss (Knutson et al., 2001). In healthy individuals, the MID task has repeatedly been demonstrated to increasingly activate the ventral striatum (i.e., nucleus accumbens [NA]) and other reward regions with the anticipation of corresponding increases in monetary reward (Knutson et al., 2000, 2008). A whole-brain analysis was conducted to examine the role of NA and PFC as well as additional brain areas in anticipatory processing, given previous research indicating alterations in insular cortex and posterior cingulate cortex (May et al., 2013; Qiu and Wang, 2021; Stewart et al., 2014; Yip et al., 2016). We hypothesized that AMP+ would demonstrate hypoactivation in NA, PFC, and insula during reward and loss anticipation compared to AMP−. Support for these hypotheses would provide evidence for the reward deficiency theory of addiction and demonstrate the role of the striatum in reward expectancy.
Methods
Participants
Participants consisted of a subset of individuals enrolled in the Tulsa 1000 (T1000) project, a naturalistic study focusing on SUD as well as mood, anxiety, and eating disorders (Victor et al., 2018). The first 500 participants are available for hypothesis testing, whereas the second 500 are reserved for replication analyses. The present analysis focuses on two groups in the first 500 T1000 subjects: (1) AMP+ (n = 46): individuals meeting criteria for DSM-IV amphetamine dependence or DSM-5 AMP on the MINI who also reported that amphetamines (predominantly methamphetamine) were their current drug of choice on the Customary Drinking and Drug Use Record (CDDR); and (2) AMP− (n = 90): individuals who did not meet criteria for any diagnoses with the exception of past MDD. Individuals were not excluded from the AMP− group for MDD diagnoses or related symptoms given the high prevalence of mood disorders/symptoms among the AMP+ group (Brown et al., 1987, 1998). AMP+ were primarily recruited from two local recovery homes (12&12 and Women In Recovery) and AMP− were primarily recruited from the greater Tulsa community via radio, print, and internet advertisements. Interested individuals gave consent to participate in phone and in-person screening to assess T1000 inclusion/exclusion criteria (no psychotic disorders or obsessive–compulsive disorder, fMRI contraindications, moderate-to-severe traumatic brain injury, unstable medical conditions, or active suicidal ideation). Those who qualified were scheduled for an in-person visit where written informed consent was obtained (see Figure 1 for group selection flowchart). Participants then completed clinical interview, behavioral, questionnaire, neuroimaging, and blood draw sessions within 2 weeks of each other. T1000 study data were collected in alignment with the Declaration of Helsinki and participants were compensated for their time.
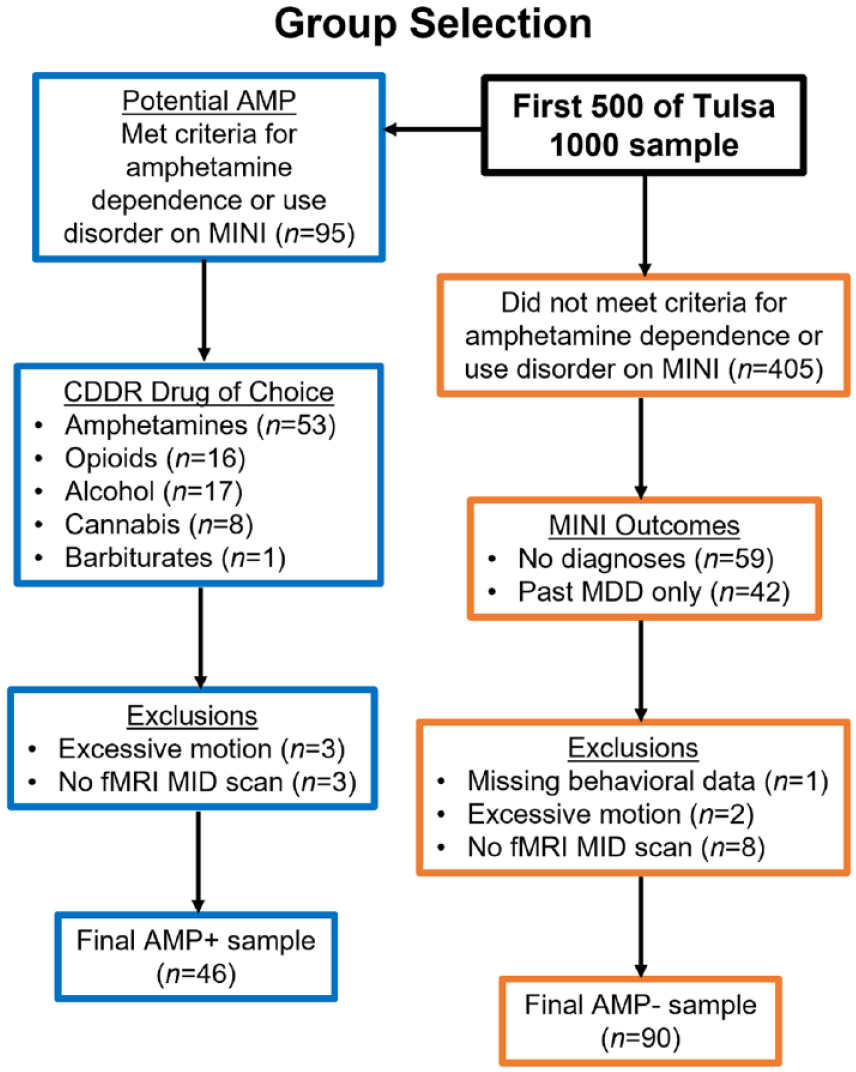
Selection of groups with and without amphetamine use disorder (AMP+ and AMP−) from the Tulsa 1000 sample for the present analysis. CDDR = Customary Drinking and Drug Record; MDD: major depressive disorder; MINI: MINI International Neuropsychiatric Interview.
Neuroimaging data collection and preprocessing
Participants completed two runs of the MID task (Knutson et al., 2001) with each run consisting of 45 trials and lasting 562 s. All participants completed a training session prior to the scan during which the task instructions were presented and individuals completed a practice run of the task to ensure comprehension of the instructions and the meaning of the cues. At the beginning of each trial of the MID task, a cue was presented to indicate a potential win or loss (circle or square) along with a magnitude of 0, 1, or 5 U.S. dollars. After a short delay, a target (white triangle) was presented, and participants were asked to press a button within a short time to win or avoid losing the amount indicated by the cue. Difficulty was calibrated by each participant’s reaction time (RT) during the practice session and updated during the scan such that participants would succeed on roughly two-thirds of the trials. The MID task was programmed in PsychoPy (Peirce, 2007), and all responses were recorded using a four-button response box (Current Designs, Philadelphia, PA, USA). Images were acquired on two identical GE MR750 3T scanners and scanning parameters were: TR/TE = 2000/27 ms, FOV/slice = 240/2.9 mm, 128 × 128 matrix, and 39 axial slices. High-resolution structural T1-weighted images were also acquired (TR/TE = 5/2.012 ms, FOV/slice = 240 × 192/0.9 mm, and 186 axial slices). All participants completed a urinary toxicology analysis prior to entering the scanner and participants with positive results did not complete the imaging session.
Neuroimaging data were preprocessed using the AFNI software package (Cox, 1996). The first three TRs were discarded, followed by despiking, slice timing correction, co-registration to anatomical volumes, motion correction, normalization to Montreal Neurological Institute space (with a final voxel size of 2 × 2 × 2 mm), and application of a 4 mm Gaussian full-width at half-max smoothing kernel. Four-second block regressors were convolved with a canonical hemodynamic response function and used to model the blood oxygen level dependent (BOLD) response to each of the six anticipatory task conditions: −5, −1, −0, +0, +1, +5. Additionally, nuisance regressors were included for the first four polynomial terms and six motion parameters. Censoring was done at the regression step by removing volumes with either a Euclidean norm of the derivatives of the six motion parameters greater than 0.3 or greater than 10% outlier voxels, determined by 3dToutcount. Percent signal change was defined as the estimated beta coefficient from single-subject analysis, which was relative to the implicit baseline during unmodeled fixation. Regressors in the model were the first four polynomial baseline terms, along with six motion parameters (roll/pitch/yaw/x/y/z translation), large loss (−5), small loss (−1), no loss (−0), no win (+0), small win (+1), and large win (+5). For visualization purposes, we averaged across no loss (+0) and no win (−0) conditions to create an overall neutral condition.
Statistical analyses
Demographic, behavior, and questionnaire analysis
Group differences on demographic variables (e.g., age Wide Range Achievement Test [WRAT] verbal IQ; Wilkinson and Robertson, 2006), Patient-Reported Outcomes Measurement Information System (PROMIS) depression and anxiety symptoms (Cella et al., 2010), BIS/BAS approach and avoidance motivation (Carver and White, 1994), Positive and Negative Affect Scale Extended (PANAS-X) positive/negative affect (Watson et al., 1988), and CDDR lifetime substance uses were examined using independent Welch t-tests and Wilcoxon rank sum (Mann–Whitney U-test) tests for variables that violated test assumptions. Group was the between subject variable and each demographic/self-report variable was the dependent measure. Fisher’s exact test was computed to compare groups on categorical variables of race and education.
Linear mixed-effects (LME) models were used to examine group (AMP+ and AMP−) differences in RT and hit rate for each condition of the MID task. A total of four LMEs were tested: (1) RT on win trials, (2) RT on loss trials, (3) hit rate on win trials, and (4) hit rate on loss trials. For each of the four LMEs, group and condition (no, small, and large wins or losses) were modeled as fixed effects, whereas subject was modeled as a random effect.
Neuroimaging analysis
AFNI’s group analysis program (Chen et al., 2014) with a multivariate modeling approach (3dMVM) was used for voxel-wise whole-brain analysis. Group (AMP+ and AMP−) was the between-subject variable, and condition (large-loss, small-loss, and no-loss) for loss anticipation was used as the within-subject variable. The resulting group main effect and group*condition interaction statistical maps were corrected for multiple comparisons at p < 0.05 using AFNI’s 3dClustsim -acf function. A similar LME model was also estimated for win anticipation (large-win, small-win, and no-win). For whole-brain corrected clusters showing significant group effects for win and/or loss anticipation, post-hoc tests were used to examine group differences. All reported findings are based on whole-brain analyses, with small volume corrections for specific a priori regions using the Brainnetome Atlas structural maps (Fan et al., 2016). A voxel level threshold of p < 0.05 was used for selected a priori regions and a threshold of p < 0.005 was used for all other regions.
To examine tasks effects, statistical maps for win versus neutral (average of small win and large win vs. no win), loss versus neutral (average of small-loss and large-loss vs. no-loss), main effect of condition during win anticipation (across large-win, small-win, and no-win), and main effect of condition during loss anticipation (average of large-loss, small-loss, and no-loss) were extracted for all subjects and corrected at p < 0.0001.
Effect sizes
For parametric tests, Hedges g is reported for group effects, whereas Cohen’s d is calculated for condition effects. For non-parametric Mann–Whitney U-tests, r values (the z statistic derived from U divided by the square root of the total sample size used in the U test) are reported.
Exploratory analyses
Additional linear regression analyses were conducted within AMP+ to explore the relationship between lifetime substance use and brain activation. Predictors were lifetime use of stimulants, nicotine, alcohol, and cannabis. Dependent variables consisted of behavioral and fMRI measures differing between groups (e.g., loss RT, left putamen to losses, etc.). All variables were log-transformed as needed and z-scored prior to analysis. Given the high co-occurrence of nicotine and methamphetamine use in prior studies (Kohut, 2017; Parker et al., 2019), Pearson correlations examined relationships between lifetime nicotine use and each dependence variable within the AMP+ group.
Results
Demographic and questionnaire results
Table 1 demonstrates group statistics for demographic and self-report data. Although groups were similar in age as well as sex and race/ethnicity distributions, AMP+ reported lower levels of education (p < 0.001) and demonstrated poorer WRAT reading scores (p < 0.0001, r = 0.51) than AMP−. There were no group differences regarding the overall number of participants taking daily prescription medications for psychiatric and physical health problems (see Supplemental Table S1 for additional details regarding medications). Groups however differed in the use of hormonal replacement/contraceptive medications (p = 0.026), with 23 AMP− versus 2 AMP+ reporting current use. With respect to questionnaire results, AMP+ reported greater BAS drive than AMP− (p < 0.001, r = 0.28), but groups did not differ in BAS reward responsiveness, BAS fun seeking, BIS inhibition, current PROMIS depression and anxiety symptoms, or PANAS positive and negative affect. Finally, AMP+ endorsed greater CDDR lifetime nicotine, cannabis, and other illicit substance use than AMP− (all p < 0.001 and r = 0.62–0.89), but groups did not differ in CDDR lifetime alcohol use.
Group demographics and self-report measures.
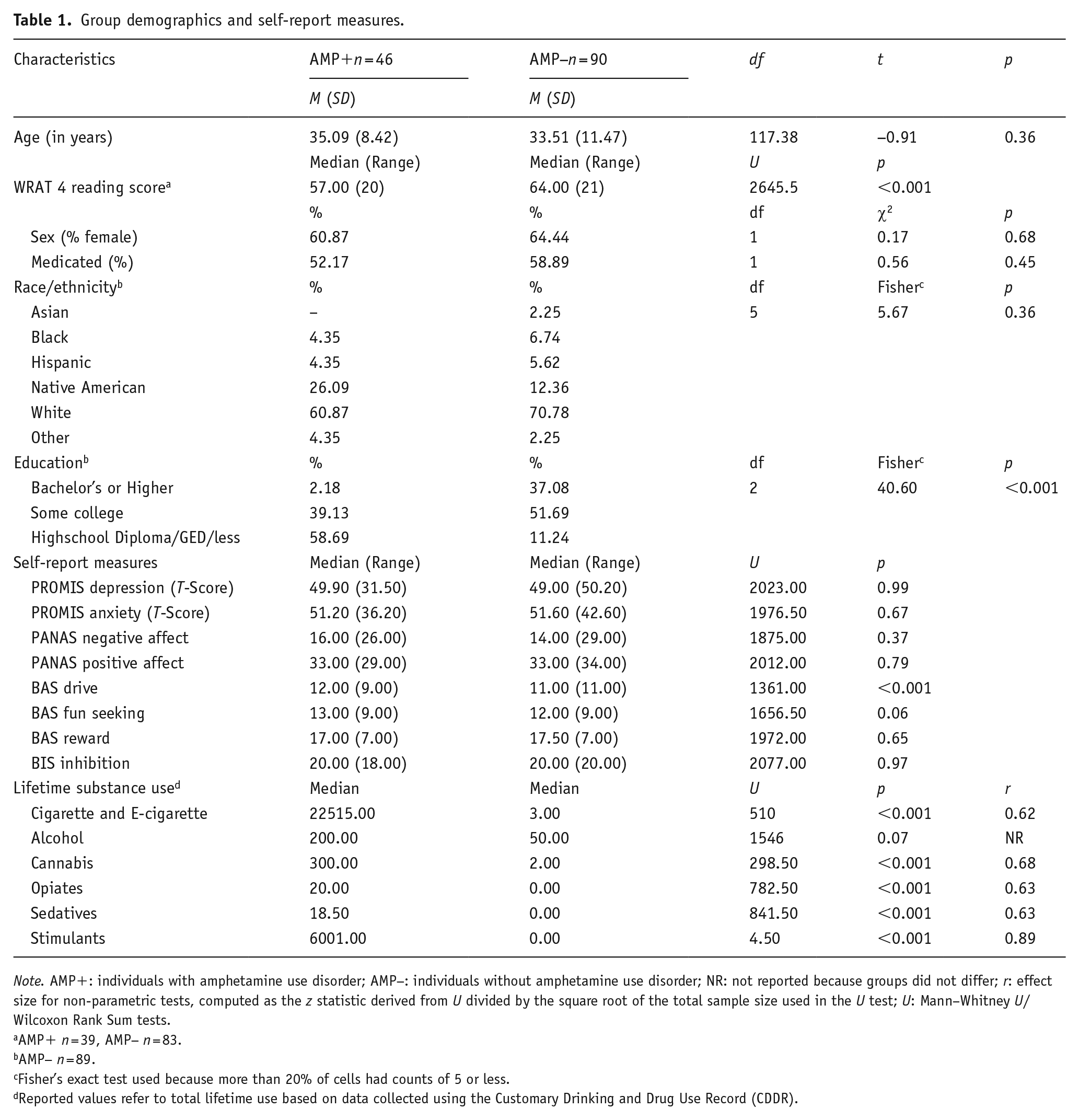
Note. AMP+: individuals with amphetamine use disorder; AMP−: individuals without amphetamine use disorder; NR: not reported because groups did not differ; r: effect size for non-parametric tests, computed as the z statistic derived from U divided by the square root of the total sample size used in the U test; U: Mann–Whitney U/Wilcoxon Rank Sum tests.
AMP+ n = 39, AMP− n = 83.
AMP− n = 89.
Fisher’s exact test used because more than 20% of cells had counts of 5 or less.
Reported values refer to total lifetime use based on data collected using the Customary Drinking and Drug Use Record (CDDR).
Behavioral results
Table 2 presents group statistics for behavioral data. Although no group main effect was evident for win anticipation RT, a significant group main effect emerged for loss anticipation RT (p = 0.02, g = 0.44), with AMP+ demonstrating slower RT than AMP− across all trials. Condition main effects showed that participants showed faster RT during small win trials compared to no win trials (d = 0.38) and during large win trials compared to no win trials (d = 0.64). Similarly, participants showed faster RT during small loss trials compared to no loss trials (d = 0.24) and during large loss trials compared to no loss trials (d = 0.40). In contrast, group*condition interactions for win and loss RT were not significant, and there were no significant findings for hits (all p-values > 0.05).
Group differences in behavioral performance.
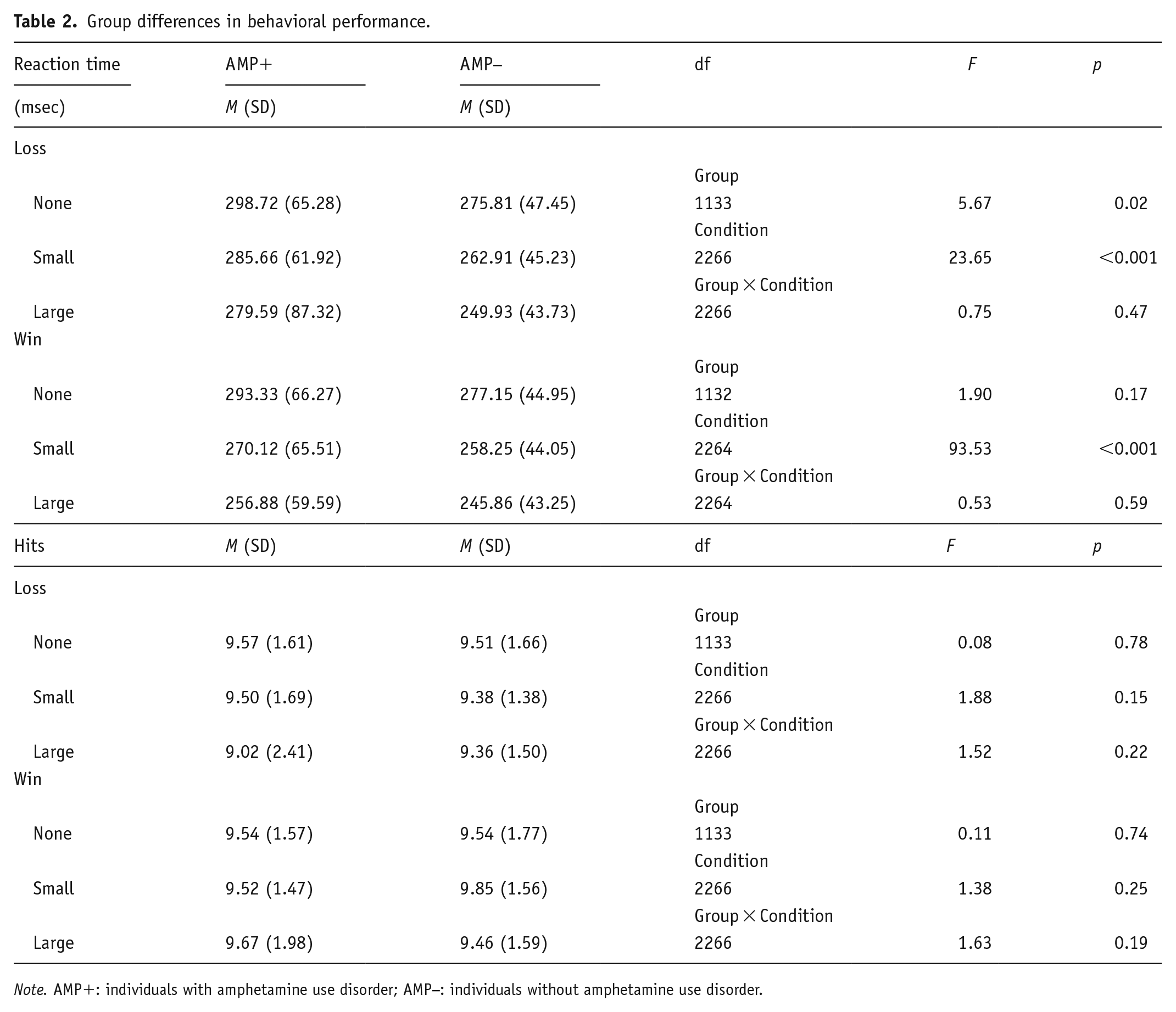
Note. AMP+: individuals with amphetamine use disorder; AMP−: individuals without amphetamine use disorder.
Neuroimaging results
Task effects
Task main effects are presented in Supplemental Table S2. The MID task elicited activation in large clusters across a wide range of brain regions including striatal, frontocingulate, insula, and temporoparietal areas. Results below center on primary hypotheses examining group and group*condition effects for win and loss anticipation.
Win anticipation
Group statistics are presented in Table 3. First, group main effects (depicted in Figure 2) demonstrated that AMP+ exhibited lower BOLD signal than AMP− in bilateral caudate (left: p = 0.004, g = 0.53; right: p = 0.002, g = 0.53), bilateral putamen (left: p = 0.007, g = 0.44; right: p = 0.02, g = 0.44), and left NA (p = 0.008, g = 0.48) across all trials. Second, post-hoc tests for group*condition interactions (shown in Figure 3) indicated that AMP+ exhibited greater BOLD signal than AMP− in two regions of right amygdala while anticipating large wins (p = 0.03; g = 0.41).
Whole-brain analysis: Group differences for win anticipation.
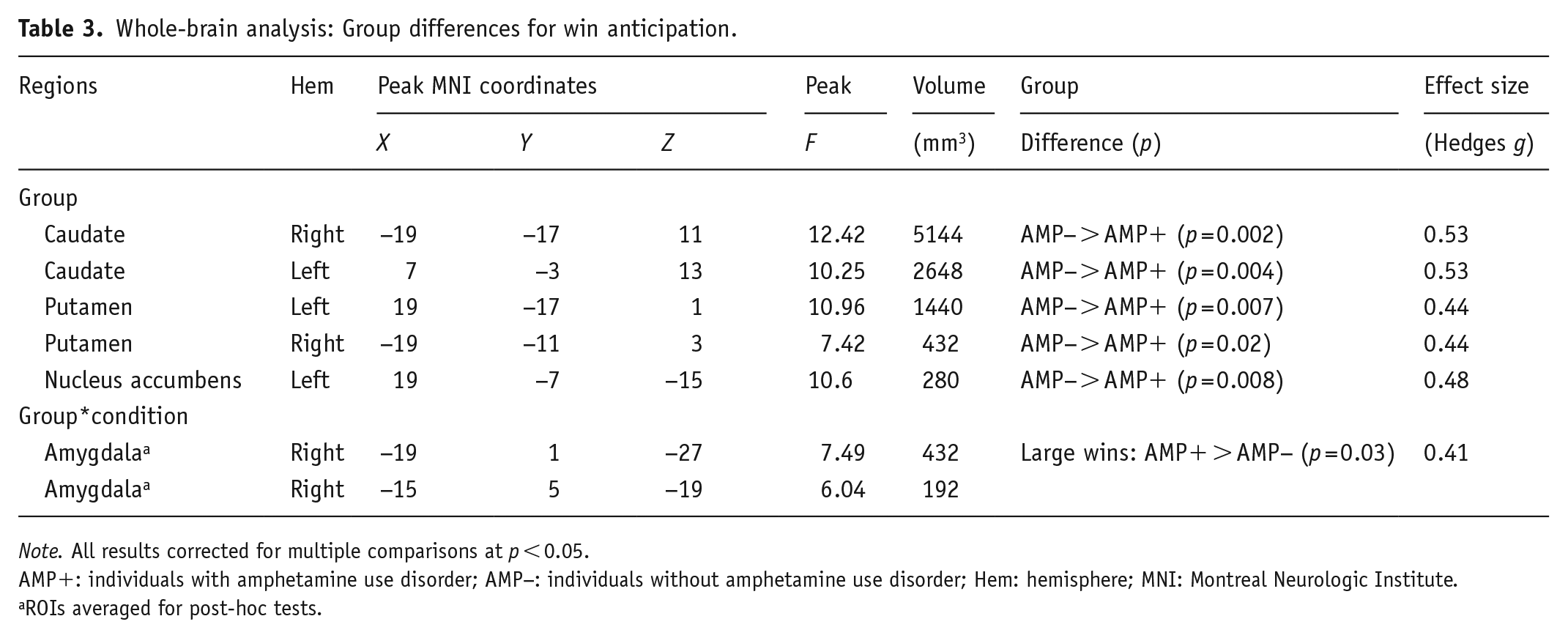
Note. All results corrected for multiple comparisons at p < 0.05.
AMP+: individuals with amphetamine use disorder; AMP−: individuals without amphetamine use disorder; Hem: hemisphere; MNI: Montreal Neurologic Institute.
ROIs averaged for post-hoc tests.
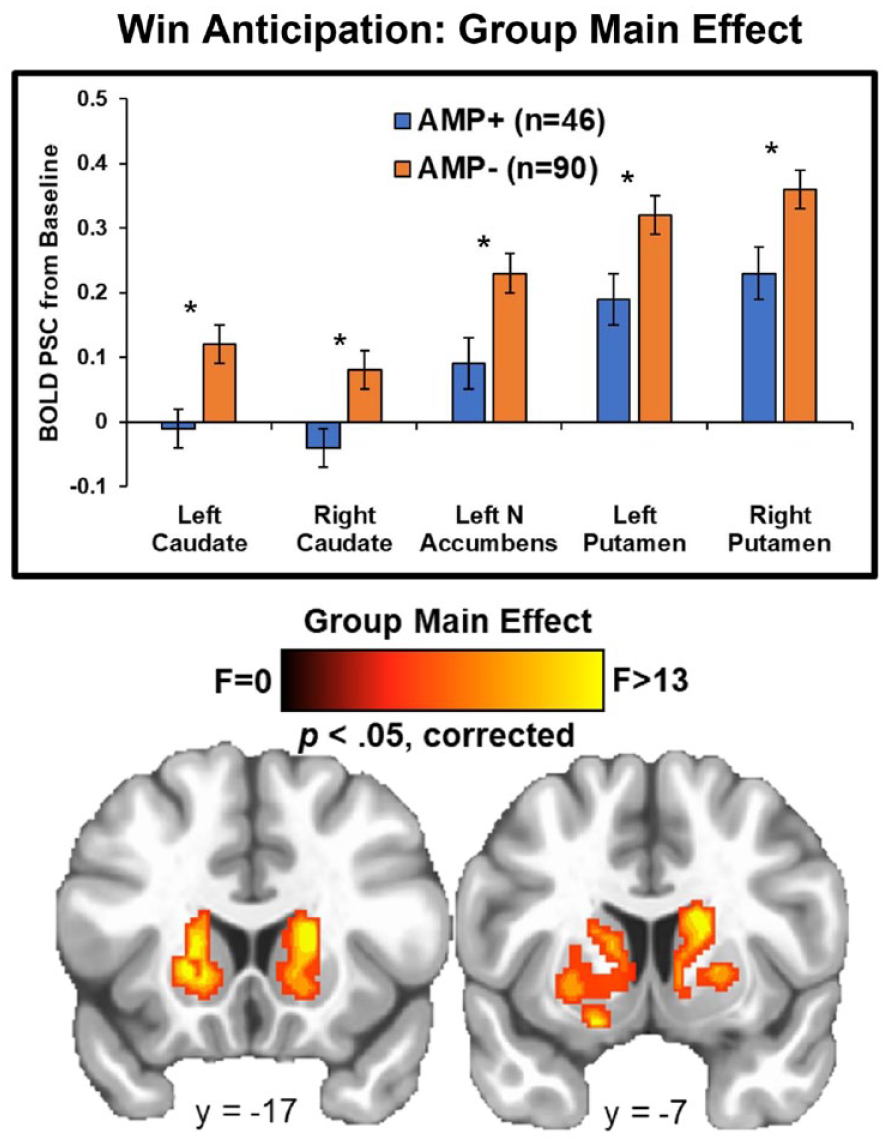
Whole-brain results for win anticipation depicting the group main effect. AMP+ exhibited lower activation than AMP− in multiple striatal regions across no win, small win, and large win trials. BOLD PSC = blood oxygen-level-dependent percent signal change. AMP+ = individuals with amphetamine use disorder. AMP− = individuals without amphetamine use disorder. NA = nucleus accumbens. Asterisks denote p < 0.05. Error bars reflect ±1 standard error. Brain image was resampled at 0.6 mm on a linear resample mode.
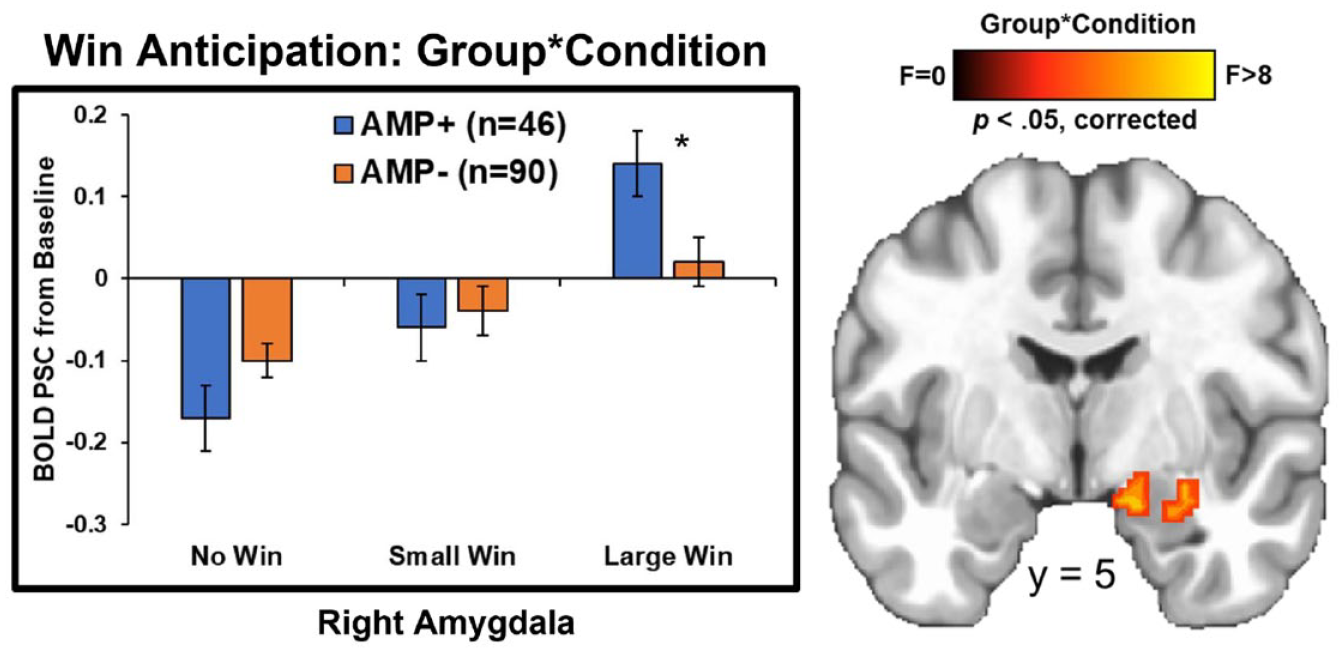
Whole-brain results for win anticipation depicting the group*condition interaction. AMP+ exhibited lower right amygdala activation than AMP− specifically during anticipation of large wins. BOLD PSC = blood oxygen-level-dependent percent signal change. AMP+ = individuals with amphetamine use disorder. AMP− = individuals without amphetamine use disorder. Asterisks denote p < 0.05. Error bars reflect ±1 standard error. Brain image was resampled at 0.6 mm on a Linear resample mode.
Loss anticipation
Group statistics are presented in Table 4. First, group main effects (depicted in Figure 4) demonstrated that AMP+ exhibited lower BOLD signal than AMP− across trials in bilateral caudate (left: p = 0.002, g = 0.50; right: p = 0.004, g = 0.55), bilateral putamen (left: p = 0.007, g = 0.47; right: p = 0.009, g = 0.44), bilateral NA (left: p = 0.004, g = 0.50; right: p = 0.005, g = 0.50), and left amygdala (p = 0.01, g = 0.48). Second, post-hoc tests for group*condition interactions (shown in Figure 5) indicated that: (1) AMP+ exhibited greater BOLD signal than AMP− in left fusiform gyrus when anticipating large losses (p = 0.02, g = 0.55); and (2) AMP+ displayed lower BOLD signal than AMP− in left caudate (p = 0.03, g = 0.41), left putamen (p = 0.002, g = 0.56), and right amygdala (p < 0.001, g = 0.62) when anticipating small losses.
Whole-brain analysis: Group differences for loss anticipation.
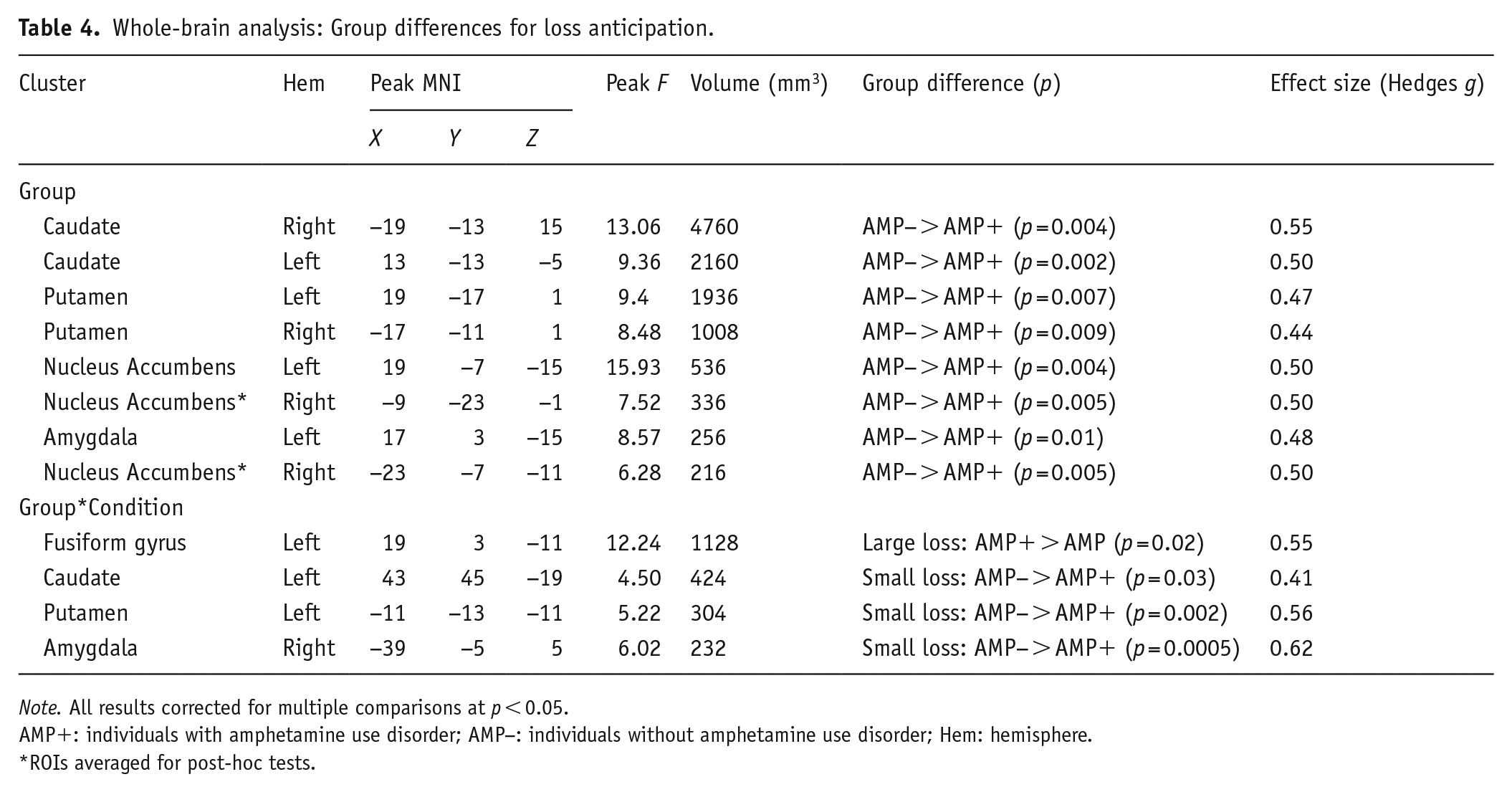
Note. All results corrected for multiple comparisons at p < 0.05.
AMP+: individuals with amphetamine use disorder; AMP−: individuals without amphetamine use disorder; Hem: hemisphere.
ROIs averaged for post-hoc tests.
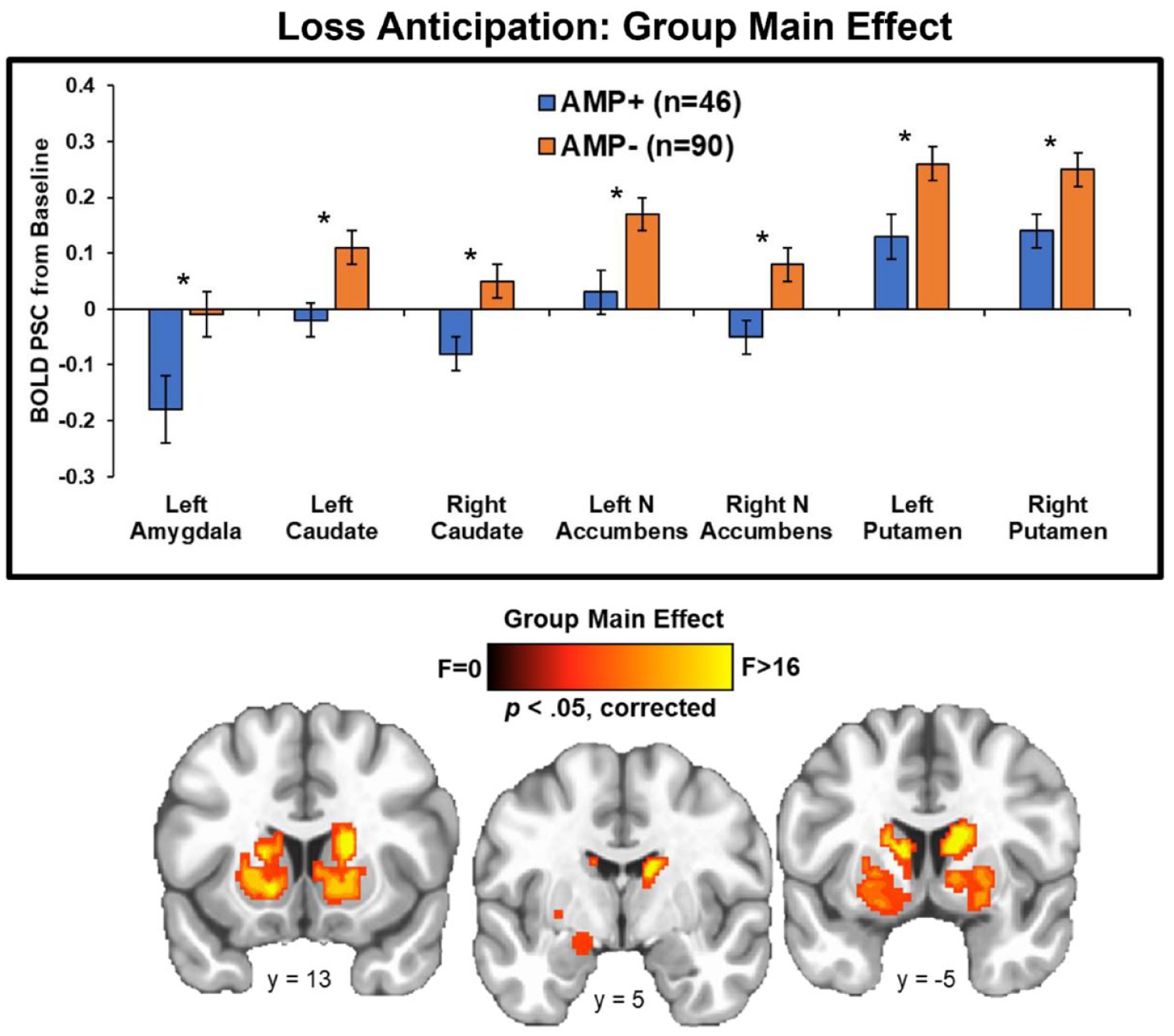
Whole-brain results for loss anticipation depicting the group main effect. AMP+ exhibited lower activation than AMP− in left amygdala and multiple striatal regions across no loss, small loss, and large loss trials. BOLD PSC = blood oxygen-level-dependent percent signal change. AMP+ = individuals with amphetamine use disorder. AMP− = individuals without amphetamine use disorder. N Accumbens = nucleus accumbens. Asterisks denote p < 0.05. Error bars reflect ±1 standard error. Brain image was resampled at 0.6 mm on a Linear resample mode.
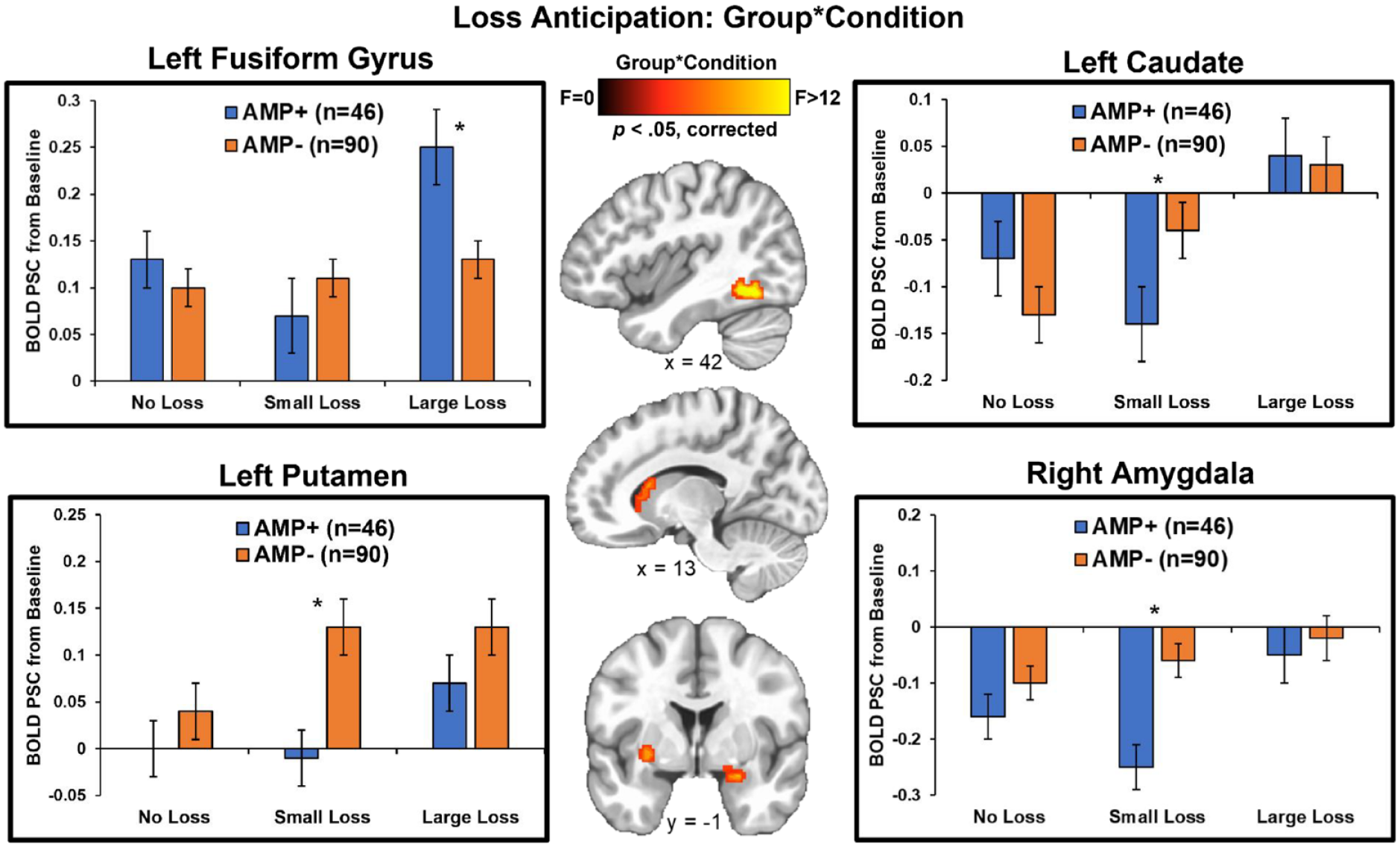
Whole-brain results for loss anticipation depicting the group*condition interaction. AMP+ exhibited greater left fusiform gyrus activation than AMP− when anticipating large losses, but showed lower left putamen, left caudate, and right amygdala activation than AMP− when anticipating small losses. BOLD PSC = blood oxygen-level-dependent percent signal change. AMP+ = individuals with amphetamine use disorder. AMP− = individuals without amphetamine use disorder. Asterisks denote p < 0.05. Error bars reflect ±1 standard error. Brain image was resampled at 0.6 mm on a Linear resample mode.
Exploratory analyses
No significant relationships were found between CDDR lifetime uses of stimulants, nicotine, alcohol, or cannabis and loss RT, nor activation within any of the significant brain regions identified by the neuroimaging analysis (all p-values > 0.05).
Discussion
This study utilized fMRI to examine differences in brain functioning during reward and loss expectancy among individuals with and without AMP. Consistent with previous research and theories regarding the persistence of addictive disorders, it was hypothesized that AMP+, compared to AMP−, would demonstrate reduced BOLD response within reward and frontal brain regions when anticipating win and loss outcomes. Overall, our findings present a general pattern of striatal hypoactivation when anticipating loss and gain among individuals with AMP compared to those without, potentially reflecting impairments in dopamine function, which may influence their decision-making and reactions to different win/loss scenarios.
Our first hypothesis predicted hypoactivation in the ventral striatum (i.e., NA) among individuals with AMP, compared to those without, during win anticipation. This hypothesis was supported; AMP+ exhibited hypoactivation within the left NA as well as the dorsal striatum (bilateral caudate and putamen) when anticipating reward, suggesting that AMP+ undervalue non-substance-related rewards compared to AMP−. As previously mentioned, alterations in reward processing have been documented in individuals with various SUD (e.g., alcohol, cocaine) and presented as a potential vulnerability factor for the development and maintenance of SUD (Qiu and Wang, 2021). Specifically, it has been theorized that reduced responsivity to non-substance-related rewards leads individuals to seek out stronger rewards, such as stimulant drugs, to activate relevant brain regions and compensate for this deficit. This would suggest that AMP+ overvaluate and pursue substance-related rewards at the expense of other rewards. Alternatively, recent work has demonstrated an overall reduction in functional connectivity within a network that includes the caudate among AMP compared to non-substance using individuals when anticipating monetary reward (but a lack of group differences when anticipating substance-related cues) (Nestor and Ersche, 2022). This finding suggests that altered reward processing among individuals with SUD applies only to non-substance-related rewards and does not involve an over-valuation of substance-related rewards; however, more evidence is needed (Nestor and Ersche, 2022). Indeed, research has demonstrated that individuals with AMP exhibit greater reward responsivity to substance-related cues than to other incentives, but less has been done to investigate whether reward processing of substance-related cues differs between AMP+ and AMP−. Our findings speak to the undervaluation of non-substance-related rewards present among AMP+ but does not examine whether an over-valuation of substance-related rewards occurs and contributes to drug-seeking behaviors. This highlights a need for additional work to examine whether the alterations in reward processing seen in AMP+ are universal and apply to all reward types.
Our study adds to the current literature by examining brain activity among individuals with AMP during loss anticipation. Consistent with our hypothesis, AMP+ demonstrated hypoactivation in bilateral striatum (ventral and dorsal) compared to AMP− when anticipating loss, and specifically within left striatum when anticipating small loss. To our knowledge, the only study to date to examine brain responsivity to loss anticipation among AMP+ reported findings of reduced ventral striatum BOLD signal among 17 individuals with methamphetamine use disorder who had elevated depression symptoms (Bischoff-Grethe et al., 2017). Our results expand on these previously reported findings by demonstrating a similar pattern of hypoactivation in bilateral striatum among a larger sample of AMP+ compared to AMP− who reported similar levels of depression, thus suggesting that the observed alterations in brain signaling are not simply due to current depression symptom severity. Instead, our findings lend support to the hypothesis that AMP persist due to poor decision-making despite negative consequences and that reduced sensitivity to the possibility of such consequences (i.e., loss anticipation) may perpetuate this behavior. Additionally, this implies that interventions focused on helping AMP+ develop skills to properly assess the potential negative consequences of their decisions and to build their motivation to use these skills may increase brain functioning when faced with these dilemmas and end the cycle of substance use.
Based on previous research, we predicted that AMP+ would exhibit hypoactivation in other regions implicated in reward processing and behavioral inhibition, specifically the insula and PFC, when anticipating reward. Our findings did not support this hypothesis; instead, alterations in amygdala signals were observed, with AMP+ showing heightened activity in right amygdala when anticipating large wins and reduced signals in left amygdala when anticipating loss, particularly when anticipating small losses. Although we did not explicitly predict these findings, they are consistent with the assertion that amygdala dysfunction is a core mechanism in the development of addiction (Koob, 2006) as well as recent work showing a positive association between amygdala activation and the subjective liking of D-amphetamine among stimulant naïve participants as a potential addiction risk factor (Langenecker et al., 2020). Additionally, the amygdala has been shown to play a role in decision-making by triggering autonomic responses to emotional stimuli (including monetary reward and loss) and providing cues to inform future decision-making (Gupta et al., 2011). These findings also converge with the theory of amphetamine sensitization, wherein repeated administration of the drug results in sensitivity to its effects and an increased release of dopamine to the NA, as well as associated alterations in BOLD response to reward outcomes (O’Daly et al., 2014). Therefore, our findings of altered BOLD signaling among AMP+ within the amygdala to the anticipation of reward and loss provide further evidence of underlying alterations in the dopaminergic network that is argued to be central to the development and persistence of SUD. Increased left fusiform gyrus signaling among AMP+ compared to AMP− when anticipating large losses was also an unpredicted finding. Although commonly implicated in facial processing, previous research has demonstrated that AMP+ have increased activity in this region in response to negative emotional faces (Kim et al., 2011; see May et al., 2020 for a review) as well as to non-facial drug-cues when compared to non-substance using individuals (Dakhili et al., 2022); taken together, the finding of altered fusiform gyrus signaling among individuals with AMP may be reflective of increased emotional processing when faced with the potential of large monetary loss.
As expounded on above, the current project replicates previous findings regarding deficits in the processing of reward anticipation among AMP+ and expands our knowledge about how these processes are also altered within the lesser researched area of loss anticipation. Despite these contributions, this study is not without limitations. First, this secondary data analysis focuses on cross-sectional data from the T1000 study, which was not designed to predict SUD or substance use (e.g., chronicity and recency) trajectories over time; additional longitudinal research is warranted to determine whether attenuated brain responses during reward anticipation predict the degree of future amphetamine use (Gowin et al., 2015). Second, AMP+ differed from AMP− in a number of characteristic ways including lower educational attainment and greater lifetime use of stimulants, cannabis, opioids, sedatives, and nicotine. Although it would be ideal to compare groups who only differed in their use of amphetamines, these variations are typical of AMP+ samples (Akindipe et al., 2014; Kohut, 2017; Liu et al., 2022; Parker et al., 2019), and our ability to rule out of the impact of these difference on brain responsivity is limited. The groups also differed in their reported use of hormonal replacement/contraceptives with a higher frequency of reported use among AMP− compared to AMP+. There is limited research to suggest that hormonal contraceptive use alters reward processing within the insular and prefrontal cortices (Bonenberger et al., 2013; Lewis et al., 2019); therefore, further research is warranted to clarify the influence of hormonal medication usage on reward processing among AMP+. Lastly, there may be other individual factors, such as genetic predispositions that could contribute to the observed alterations in brain functioning.
Conclusion
This project expands on prior work, which has primarily focused on brain functioning during reward anticipation, to include an examination of BOLD response to the anticipation of loss among individuals with AMP. Our findings suggest that alterations in reward processing contribute to the persistence of SUD by: (1) leading individuals to seek out stronger rewards (i.e., illicit substances) because they under-value primary incentives when anticipating the potential rewarding effects; and (2) perpetuating the cycle of poor decision-making due to an attenuated neural response to loss anticipation and underappreciation of the potential of negative consequences.
Supplemental Material
sj-docx-1-jop-10.1177_02698811231222355 – Supplemental material for Amphetamine use disorder is associated with striatum hypoactivation during anticipation of loss and reward
Supplemental material, sj-docx-1-jop-10.1177_02698811231222355 for Amphetamine use disorder is associated with striatum hypoactivation during anticipation of loss and reward by April C May, Kaiping Burrows, Rayus Kuplicki, Martin P Paulus and Jennifer L Stewart in Journal of Psychopharmacology
Footnotes
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by The Laureate Institute for Brain Research, the National Institute on Drug Abuse (R01DA050677; PI: Stewart), and the National Institute of General Medical Sciences (P20GM121312; PI: Paulus).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.