
Abstract
Background:
Maternal sleep deprivation (MSD) is a significant public health issue that adversely affects neurogenesis and synaptic plasticity in offspring, resulting in cognitive deficits in learning and memory. Resveratrol, an antioxidant with neuroprotective and anti-inflammatory properties, may help mitigate these effects. This study investigates resveratrol’s potential to counteract the negative impacts of MSD on neurodevelopment in male Wistar rat offspring.
Methods:
Ninety-six male Wistar rat offspring and 36 pregnant rats were used. Total MSD was induced using the water box device on gestational days 7, 11, and 17. Pregnant rats received resveratrol at doses of 25 or 50 mg/kg every 12 h during the sleep deprivation period. After parturition, offspring were divided into 12 groups for assessment at two months of age. Social interaction tests evaluated social memory, while the Morris water maze test assessed spatial learning and memory. Brain samples were prepared for Nissl staining, and brain-derived neurotrophic factor (BDNF) and tyrosine-protein kinase (TrkB) expression levels in the hippocampus were measured using western blotting.
Results:
Our findings indicate that the MSD group exhibited decreased BDNF/TrkB expression and increased neuronal damage in the hippocampus, which led to disrupted spatial and social memory compared to the control group. Subsequently, resveratrol administration, especially at a dose of 50 mg/kg during pregnancy, significantly reversed MSD’s detrimental effects on cognitive function in offspring.
Conclusion:
Our results provide novel evidence of resveratrol’s neuroprotective effects in rat pregnancy models of MSD, suggesting its potential for developing therapeutic interventions targeting prenatal neurodegenerative disorders.
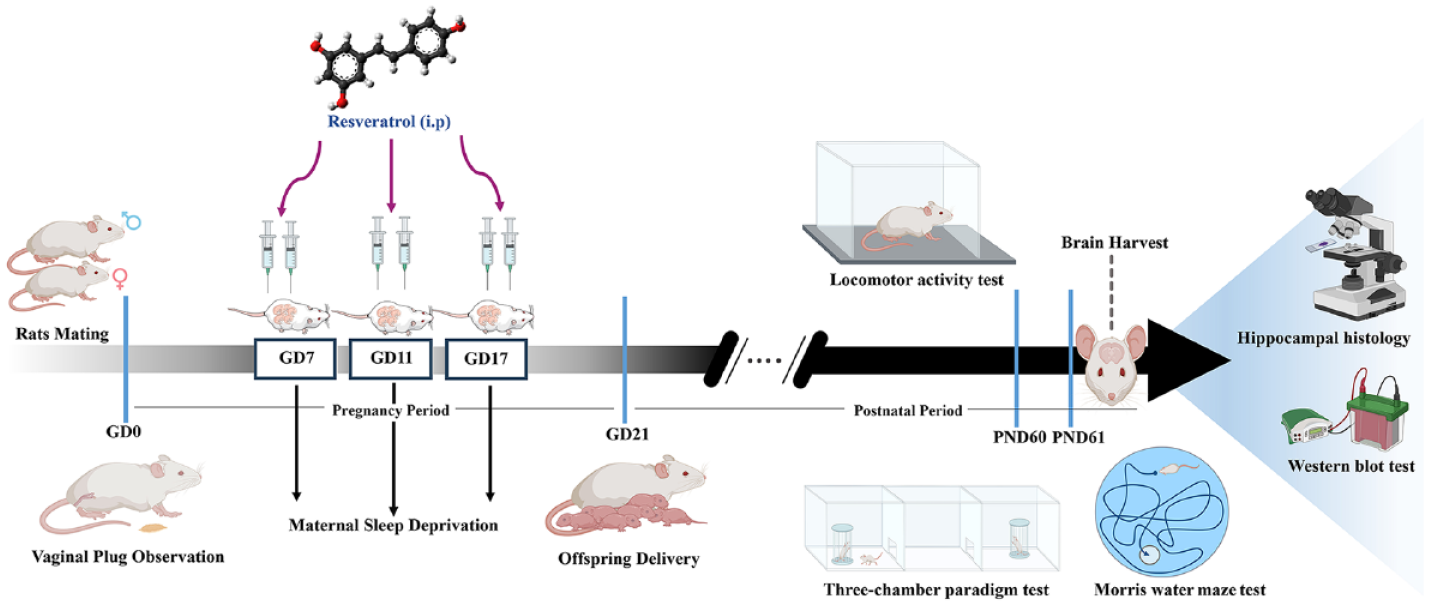
Introduction
Sleep is crucial for survival and health. It is a dynamic recovery process that partially disconnects the central nervous system from the external environment, reducing sensory, motor, and mental functions (Yao et al., 2022). This period plays a pivotal role in memory formation, learning, maintaining oxidant-antioxidant balance, endocrine regulation, emotional stability, and immune function enhancement (Rezaie et al., 2020, 2021).
Maternal sleep deprivation (MSD) is becoming increasingly prevalent during pregnancy due to lifestyle changes, hormonal fluctuations, and physical or behavioral factors (Sedov et al., 2018). Sleep disturbances during pregnancy not only impair maternal health but also disrupt the intrauterine environment, negatively affecting fetal growth and nervous system development. These disruptions can lead to long-term cognitive deficits, altered synaptic plasticity, and neurodevelopmental impairments in offspring that may persist into adulthood (Chang et al., 2010; Zhao et al., 2014; Zhang et al., 2023c).
Despite the protective role of the placenta, MSD has been shown to disrupt the hypothalamic-pituitary-adrenal (HPA) axis and placental function, elevating maternal and fetal pro-inflammatory cytokines, which increase the risk of long-term neurodevelopmental disorders in offspring (Howerton and Bale, 2012). The overall effects of maternal sleep disruptions on offspring physiology and behavior remain poorly understood, though rodent studies consistently show that MSD during pregnancy leads to spatial memory deficits, anxiety-like behaviors, and cognitive impairments in offspring (Radhakrishnan et al., 2015; Trzepizur et al., 2017).
The hippocampus, a crucial brain region for neurogenesis, is integral to mood regulation, learning, and memory. Disrupted maternal sleep significantly impairs hippocampal neurogenesis, alters circadian rhythms, and increases neuroinflammation, negatively impacting cognitive functions in offspring (Zhao et al., 2014, 2015). Brain-derived neurotrophic factor (BDNF) and its receptor, tyrosine-protein kinase (TrkB), are essential for hippocampal function, particularly learning and memory. BDNF expression peaks during critical windows of development, including the early embryonic period and postnatal days 10–14, and is crucial for dendritic sprouting, cell survival, synaptic plasticity, and stress resistance (Neeley et al., 2011). BDNF also regulates key processes such as learning, memory, and sleep by influencing neurotransmitter release (Hassanzadeh et al., 2021; Zhang et al., 2023a). TrkB, which mediates BDNF activity, plays an indispensable role in hippocampal neuron function and memory formation. The BDNF-TrkB signaling pathway is essential for maintaining optimal brain function, particularly in regions associated with learning and memory processes (Qiu et al., 2020).
Resveratrol (3, 4′, 5 trihydroxystilbene) is a natural polyphenol found in grapes, berries, and peanuts and exhibits neuroprotective properties. The trans-resveratrol isomer, with a low molecular weight (228 Da), allows it to cross the blood-brain barrier (BBB) effectively (Komorowska et al., 2024). This polyphenol reaches the brain within just four hours of administration, demonstrating its prompt neurological impact (Wang et al., 2002). Resveratrol has been shown to enhance cognitive functions such as learning, memory, and emotional stability while protecting neurons and glial cells from damage through its antioxidative, anti-inflammatory, and neuroprotective effects (Albani et al., 2010; Gocmez et al., 2016; Izquierdo et al., 2019).
This study aims to investigate the molecular aspects of resveratrol’s effects on the BDNF-TrkB signaling pathway during total sleep deprivation (TSD) in pregnant rats and its subsequent impact on the cognitive and hippocampal development of male offspring.
Materials and methods
Drugs
The Resveratrol (CAS No: 501-36-0) was procured from Chemenu Inc. (shanghai, China) and dissolved in Dimethyl Sulfoxide (CAS No:67-68-5). The BDNF antibody (Cat# PAB7956, RRID: AB_1673512) and TrkB antibody (Cat# PAB18110, RRID: AB_10716656) were provided by Abnova. Beta-actin (Cat# 4967, RRID: AB_330288), rabbit Immunoglobulin G, and horseradish peroxidase-related antibody (Cat# 7074, RRID: AB_2099233) were utilized in the western blot. Also, Ketamine hydrochloride and xylazine were purchased by Alfasan (Woerden, the Netherlands).
Animals and experimental protocol
The study began with Thirty-six adult female Wistar rats (ICSS Institute, Tehran, Iran), which were used for mating. These female rats were randomly assigned into 12 experimental groups, with three females per group. Virgin female rats were paired overnight with male rats, and vaginal plugs were checked daily to determine the gestational day (GD 0) (Mohammadi-Mahdiabadi-Hasani et al., 2020). To induce MSD, pregnant rats were completely deprived of sleep for 24 h on three specific gestational days (GD7, GD11, and GD17) using an automated water box (BorjSanatazma Co, Tehran, Iran). Based on previous studies, the most critical embryonic days (gestational days) for forming the central nervous system in rats are E7, E11, and E17 (Chen et al., 2017). According to the short half-life of resveratrol in rodents due to rapid metabolism (Lacerda et al., 2023), during this gestational period, pregnant rats received intraperitoneal (i.p.) injections of resveratrol at doses of 25 or 50 mg/kg, administered every 12 h (at 8 a.m. and 8 p.m.). Following birth, offspring remained with their mothers until postnatal days 21 (PND1–PND21). After this period, only the male offspring were randomly selected for further study and assigned to experimental subgroups based on their respective maternal groups. All animals were kept in laboratory conditions with a temperature of 22 ± 2℃ and under a light/dark cycle (12 h/12 h). Hygienic conditions were kept with four rats per cage, and they had unrestricted access to food and water.
Experimental groups
The experimental groups were categorized as follows:
Non-sleep-deprived groups:
Group 1 (Control): Pregnant rats in this group received no intervention.
Group 2 (Vehicle): Pregnant rats were administered DMSO injections (10%, i.p.) every 12 h.
Group 3 (Resveratrol 25 mg/kg): Pregnant rats were administered resveratrol injections (25 mg/kg, i.p.) every 12 h.
Group 4 (Resveratrol 50 mg/kg): Pregnant rats were administered resveratrol injections (50 mg/kg, i.p.) every 12 h.
Sham-sleep-deprived groups:
Group 5 (Control): Pregnant rats were placed in the turned-off TSD apparatus without any injections.
Group 6 (Vehicle): Pregnant rats were administered DMSO injections (10%, i.p.) every 12 h and then placed in the turned-off TSD apparatus.
Group 7 (Resveratrol 25 mg/kg): Pregnant rats were administered resveratrol injections (25 mg/kg, i.p.) every 12 h and then placed in the turned-off TSD apparatus.
Group 8 (Resveratrol 50 mg/kg): Pregnant rats were administered resveratrol injections (50 mg/kg, i.p.) every 12 h and then placed in the turned-off TSD apparatus.
Maternal sleep-deprived groups:
Group 9 (Control): Pregnant rats were placed in the turned-on TSD apparatus (under MSD condition) without any injection.
Group 10 (Vehicle): Pregnant rats were administered DMSO injections (10%, i.p.) every 12 h and then placed in the turned-on TSD apparatus.
Group 11 (Resveratrol 25 mg/kg): Pregnant rats were administered resveratrol injections (25 mg/kg, i.p.) every 12 h and then placed in the turned-on TSD apparatus.
Group 12 (Resveratrol 50 mg/kg): Pregnant rats were administered resveratrol injections (50 mg/kg, i.p.) every 12 h and then placed in the turned-on TSD apparatus.
Offspring and experimental procedures
The male offspring were maintained under standard conditions with unrestricted sleep access until PND60. Based on previous studies on calculating sample size in animal studies (Arifin and Zahiruddin, 2017; Charan and Kantharia, 2013; Forghani et al., 2023), eight male rats were considered for statistical analysis in each experimental group. Behavioral procedures were performed between 9:00 a.m. to 2:00 p.m. After completing the behavior assessments, five male offspring per experimental group were euthanized using CO2 inhalation. The brains were then properly extracted, and the hippocampus was quickly frozen in liquid nitrogen. After that, the samples were kept at −80°C for Western blotting. Subsequently, the remaining rats were anesthetized with ketamine and xylazine and perfused with phosphate-buffered saline, and their brains were fixed in 4% formaldehyde for histomorphological studies.
All experimental procedures followed the established international guidelines for laboratory animal usage and were approved by the local Ethics Committee of Tehran University of Medical Sciences (IR.TUMS.AEC.1400.052). The experimental protocol was summarized in a schematic diagram presented in Figure 1.
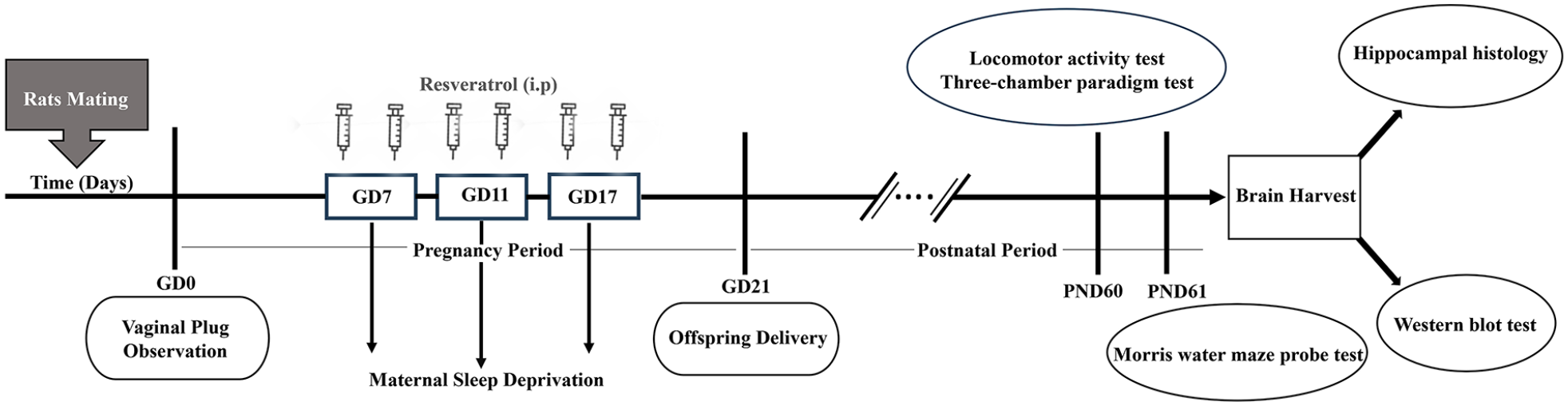
Schematic illustration of the experimental protocol. Male rats were mated with female rats, and vaginal plugs were checked daily to identify gestational day (GD 0). Pregnant rats were then deprived of sleep for 24 h on three key gestational days (GD7, GD11, and GD17) and received intraperitoneal injections of resveratrol at 25 or 50 mg/kg doses every 12 h. After birth, offspring remained with their mothers for 21 days during infancy. Only male offspring were randomly separated. On postnatal days 60 or 61, behavioral tests were assessed. Finally, the animals were sacrificed, and their brain tissues were harvested for western blot and histological studies.
MSD procedure
Female rats were sleep-deprived on GD7, 11, and 17 of pregnancy using an automated water box apparatus. The device consists of four boxes measuring 30 cm × 30 cm × 50 cm, each capable of completely depriving one pregnant rat of sleep. The apparatus is connected to a tank that measures 120 cm × 50 cm × 30 cm, filled with water at ambient temperature. Two circular pedals, each 15 cm in diameter and 3 mm edged, perforated surfaces, each powered by a 1-W DC motor, are located within each box and move in opposing directions at a 1.5 cm/s speed. The rats had to remain awake throughout the experiment. After reaching the highest platform, the pedals were held for 20 s, and the pregnant rats had access to water and food pellets. During the TSD protocol, the pregnant rats remained fully awake. After the procedure, they were returned to their home cages (Nasehi et al., 2021). The pregnant rats in the MSD groups were placed for 24 h in the device with it turned on, while those in the sham groups were exposed to the switched-off apparatus for the same duration.
Locomotor activity test
On postnatal day 60, the offspring rats were tested for locomotion activity in a dimly lit, quiet room without prior habituation. The locomotion apparatus consisted of a transparent Perspex container measuring 40 cm × 30 cm × 30 cm. The device floor was made of a 2.2 cm thick gray Perspex panel divided into 16 equal squares using 16 photocells. Each experiment lasted 300 s, during which locomotor activity was measured by counting movements between the squares (Khakpoor et al., 2021).
Three-chamber paradigm test
On the 60th day after birth, offspring were subjected to the three-chamber paradigm test. Crawley’s sociability and social novelty preference assess social interaction behaviors. This apparatus was a rectangular box divided into three equal-sized chambers measuring 76 × 30 × 35, separated by walls. The lateral chambers, each made of wire glass and with a removable top, held one rat, each placed vertically. The compartments contained familiar and stranger rats. The test was divided into three phases: habituation, sociability assessment, and social preference evaluation. Experiments were conducted in a room with 650 lx lighting. A camera recorded all parameters and the determined time spent in each compartment (Rezaie et al., 2020).
Habituation phase: The gates separating the apparatus were closed in this phase. Each rat was placed in the middle chamber for 5 min to acclimate.
Sociability phase: For 10 min, the walls separating the chambers were taken down, allowing the offspring rats to move freely between all three chambers. The time spent on the subject offspring rat’s direct interactions with a stranger 1 in one side chamber and an empty box in the other was measured.
Social preference phase: During this phase, rat stranger two was placed inside the wire cup that was previously empty, opposite the cup where stranger one was placed for 10 min. The time spent on the subject offspring rat’s direct interactions with stranger 1 (Familiar rat) in one side chamber and stranger 2 (novel rat) in the other was measured (equations (1) and (2)).
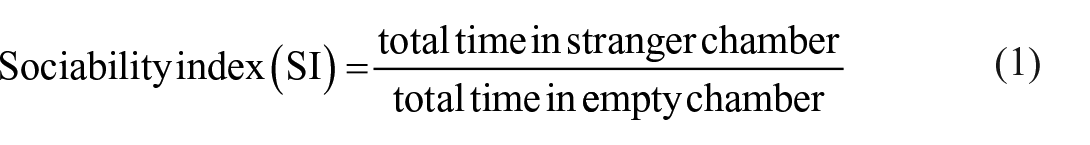
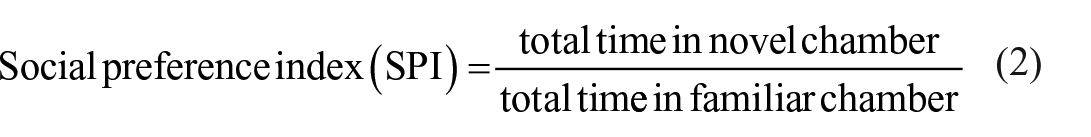
Morris water maze
Sixty days after birth, all male offspring of Wistar rats were trained in the Morris water maze, a valid method for assessing spatial memory performance in rodents. This apparatus is a round black pool that is 150 cm in diameter, 60 cm deep, and filled with 22 ± 24°C water to a depth of 30 cm. The test room has a spatial layout to facilitate navigation with clear and stable cues attached to the walls. The pool was divided into four equal zones, each with starting locations at equal distances on the rim: north (N), south (S), west (W), and east (E). An escape platform with a 10 cm diameter was placed 1 cm below the water level in the center of the target zone (the northwest quadrant). During the experiment, a camera above the pool recorded each rat swimming, sending the data to a computer. A video tracking system (BorjSanatazma Co, Tehran, Iran) measured three parameters during the trial session. These parameters include escape latency (time to reach the hidden platform), traveled distance (path length to find the hidden platform), and swimming speed (rat swimming velocity in each training session).
The escape latency and traveled distance in the target zone were measured during the probing session. On the first day, eight trials were performed with four different starting positions during training sessions. The starting position of each rat was evenly distributed around the circumference of the maze. At the onset of each trial, the animal was placed within one of the zones. The rats had 60 s to find the escape platform using distal spatial cues. Upon discovering the escape platform, the rats were given 20 s to memorize spatial cues and situations. Before the subsequent trial, they were placed in a cage for 20 s. If the rat fails to find the platform, the researcher guides it to the platform and gives it a 20-s rest. The probe trial (retrieval test session) was performed on the second day. Before starting, the escape platform was taken out of the pool. In the probe trial, rats freely swam for 60 s while recording their time spent in the target quadrant and distance traveled to determine their spatial memory retrieval, which is crucial in identifying memory impairment. The nonspatial visible test was conducted after the probe experiment. For the visibility test, the platform was raised above the water level and visually sealed with aluminum foil in the center of the northeast quadrant. This assessment evaluates nonspecific effects on motivation and motor and visual skills unrelated to memory or learning. Following the probing test, a visible platform test was performed to ensure that the rat’s motor skills, visuomotor abilities, and desire to escape water did not affect the results (Vaseghi et al., 2018).
Western blot
After the probe and nonspatial behavioral tests, the brains (right hippocampus) of five rats were randomly selected from each group. Western blot analysis assessed BDNF and TrkB expression levels in hippocampus tissue. The hippocampus tissues were homogenized in a lysis buffer with a cocktail of protease inhibitors using a homogenizer (Micro Smash MS-100). The homogenates were centrifuged for 30 min at 13,000 g at 4°C, and the resulting upper phase was competently collected as total protein extracts. By employing Bradford’s assay, the protein concentration in the samples was determined. Subsequently, 60 μg of protein samples were loaded on a gel composed of 12.5% sodium dodecyl sulfate-polyacrylamide (SDS = 8%, pH = 8.3). During electrophoresis, a voltage of 120 V was maintained for 120 min. Subsequently, the proteins were transferred to the polyvinylidene fluoride (PVDF) membrane. The PVDF membranes were incubated with a 2% blocking solution for 75 min at room temperature to block nonspecific protein binding. Then, the blots were incubated overnight at 4°C with specific primary antibodies against BDNF and TrkB (Abnova), diluted to 1:1000 in 2% BSA. The following day, after washing the membrane three times with TBST (Tris-buffered saline with Tween 20), it was incubated with a horse radish peroxidase-linked secondary antibody for 90 min at room temperature. Protein bands were detected by chemiluminescence using an ECL kit (Bio-Rad, CA, USA) and observed via radiography film—ImageJ software (RRID: SCR_003070, National Institute of Health, USA) quantified protein band density. Blots were stripped and re-probed with rabbit monoclonal β-actin (1:1000) antibodies to ensure equal loading (Mohammadi et al., 2023).
Cresyl violet (Nissl) staining
The Nissl staining method was utilized to identify and quantify neuronal loss in the hippocampus. Brain hemispheres were embedded in paraffin, sliced into 7 μm thickness, and placed on glass slides. After deparaffinization and rehydration, the tissue section was stained with a 0.1% solution of cresyl violet acetate. Three slides were observed with light microscopy and captured with a computer camera. We counted dark neurons in a 1 mm length of the CA1 region using ImageJ software. Finally, we averaged the results across different groups to obtain the final value (Karimi-Zandi et al., 2022).
Statistical analysis
All statistical analysis was performed using GraphPad Prism software (version 9.0, Inc., La Jolla, CA, USA). Data was divided into three categories: behavioral, molecular, and histological assessments. Repeated measures, two-way ANOVA, one-way ANOVA, and post hoc Tukey’s tests were used to compare the behavioral data. Similarly, two-way ANOVA and one-way ANOVA, with post hoc Tukey’s tests, were conducted to compare the molecular changes (western blot) and histology in the hippocampus (Nissl staining). All data was presented as mean ± SEM, and statistical significance was considered at p-value < 0.05.
Results
The effect of MSD and different doses of resveratrol on spatial learning
The results from the eight trials of the training phase showed that MSD significantly increased the escape latency time during the trials for the male offspring to find the hidden platform across the trials (as shown in Table 1, Figure 2(a)). Repeated measures of ANOVA and post hoc Tukey test results confirmed that MSD induced a deficit in spatial learning performance in the male offspring. However, maternal administration of resveratrol at 25 or 50 mg/kg during pregnancy significantly reduced the escape latency time, effectively restoring spatial learning performance in male offspring (Figure 2(a)).
Comparison of Morris water maze parameters between all the experimental groups of male rat offspring.

Panel left: one-way ANOVA analysis comparing the non-sleep-deprived groups; Panel middle versus left: two-way ANOVA analysis comparing the non-sleep-deprived groups and sham-sleep-deprived groups; Panel right versus left: two-way ANOVA analysis comparing the non-sleep-deprived groups and maternal sleep-deprived groups (see Figure 2).
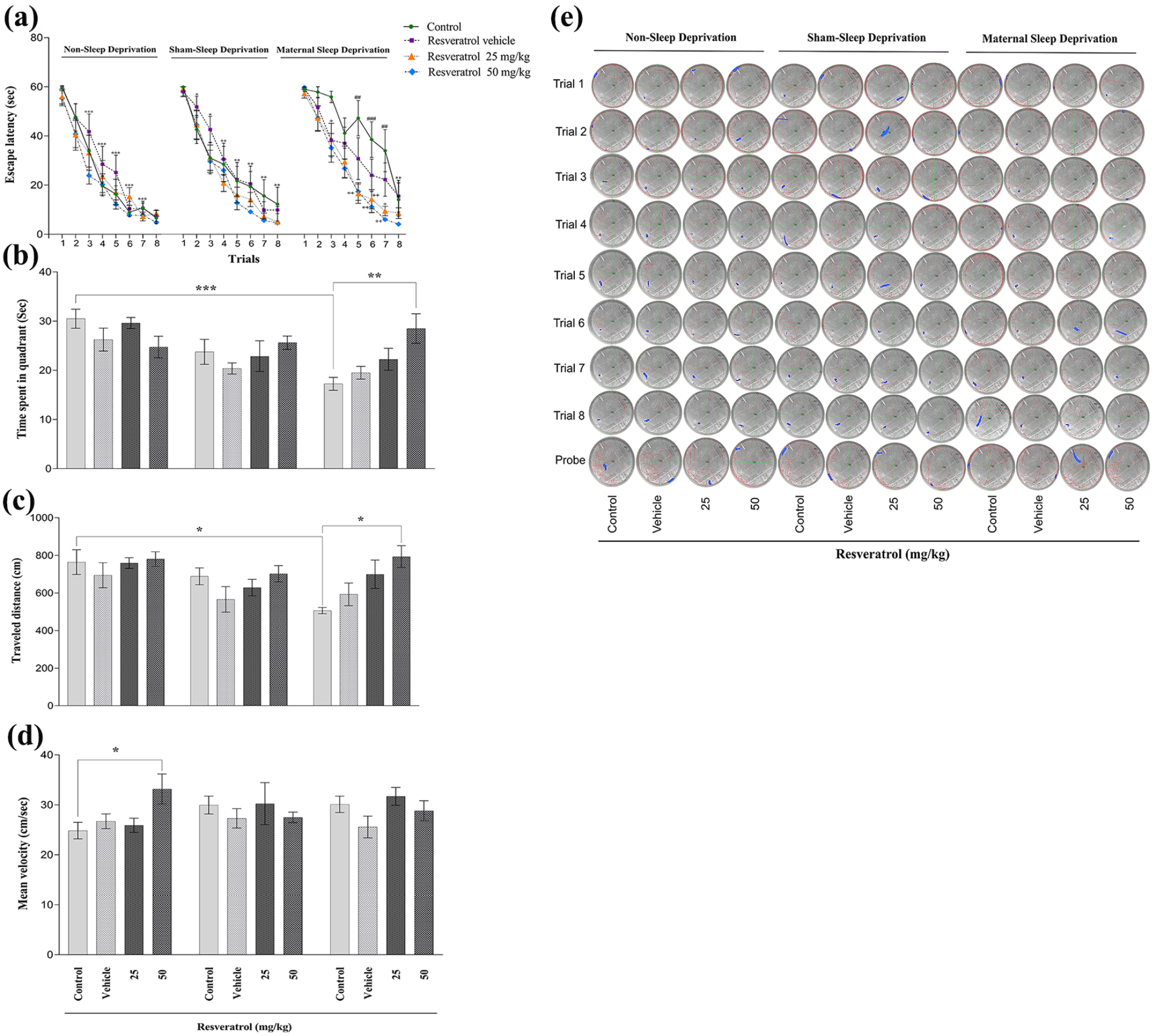
The effect of maternal sleep deprivation and different doses of resveratrol (25 or 50 mg/kg) on spatial memory performance in male Wistar rat offspring in the non-sleep-deprived, sham-sleep-deprived, and sleep-deprived groups. The escape latency (a), time spent in the target quadrant (b), traveled distance (c), mean velocity (d), and a sample trace of the traveled distance-heat map in each group were depicted in the (e) section. Values represented are mean ± SEM (n = 8). In (a) section: *p < 0.05, **p < 0.01, and ***p < 0.001 as compared to the first day of trials; +p < 0.05 and ++p < 0.01 as compared to respective controls in the maternal sleep-deprived group; ##p < 0.01 and ###p < 0.001 compared to the respective controls in the non-sleep-deprived group. In (b–d) section: *p < 0.05, **p < 0.01, and ***p < 0.001 represent significant differences between the studied groups (non-sleep deprivation (left panel), Sham-sleep deprivation (middle panel), and maternal sleep deprivation (right panel); see Table 1).
The effect of MSD and different doses of resveratrol on spatial memory
Time spent in the quadrant
During the probe test, the results of the two-way ANOVA and post hoc analysis revealed that MSD significantly reduced the time male offspring spent in the target quadrant, indicating impaired spatial memory (Table 1, Figure 2(b)). Furthermore, administering resveratrol at 50 mg/kg to the pregnant rats was able to restore the spatial memory deficit. In contrast, the time spent in the target quadrant was not altered in the sham-sleep-deprived groups compared to the control groups of the non-sleep-deprived group. A one-way ANOVA and post hoc analysis indicated that the time spent in the target quadrant was not significantly different among the non-sleep-deprived groups (Figure 2(b)).
Traveled distance
Further analysis using two-way ANOVA and post hoc tests demonstrated that prenatal sleep deprivation led to a reduction in the distance traveled by male offspring within the target quadrant (Table 1, Figure 2(c)). This impairment was successfully reversed after maternal administration with resveratrol at 50 mg/kg (Figure 2(c)).
Mean velocity
The one-way ANOVA and post hoc Tukey test revealed that the offspring of mothers who received 50 mg/kg of resveratrol dosage significantly increased mean velocity within the target quadrant compared to the non-sleep-deprived control group (Table 1, Figure 2(d)).
The effect of MSD and different doses of resveratrol on locomotion
There was no statistically significant difference in locomotor activity between the non-sleep-deprived groups (Figure 3). Similarly, two-way ANOVA analysis confirmed no statistically significant variation in locomotion across all experimental groups (Table 2, Figure 3).
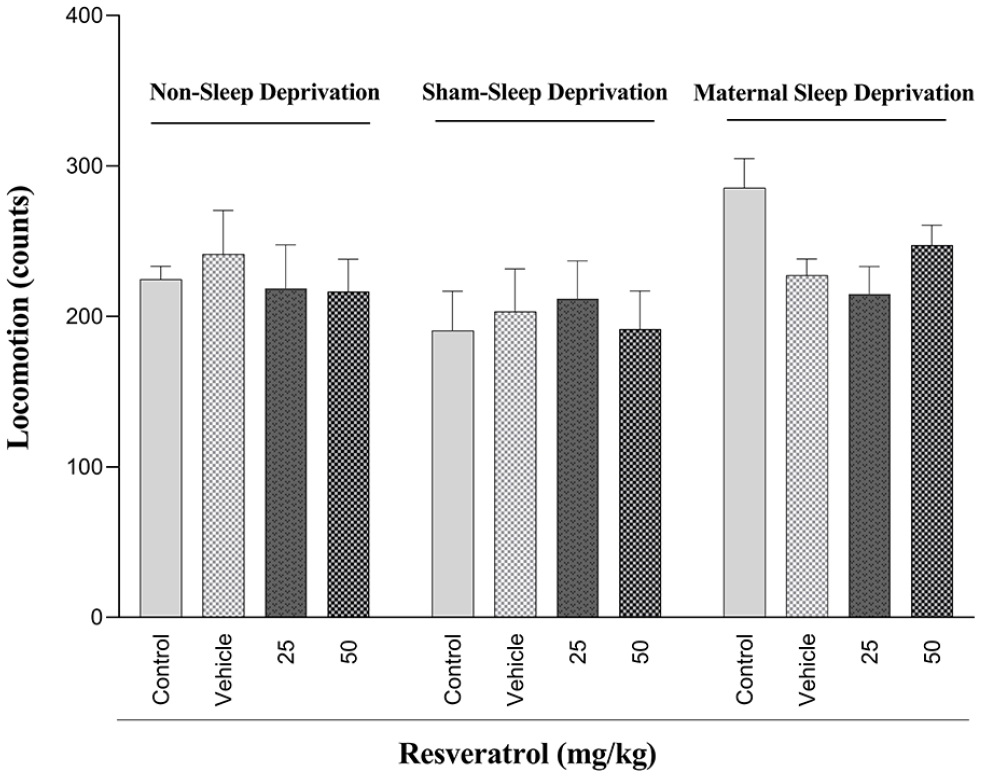
The effect of maternal sleep deprivation and different doses of resveratrol (25 or 50 mg/kg) on locomotion in male Wistar rat offspring in the non-sleep-deprived, sham-sleep-deprived, and sleep-deprived groups. Values represented are mean ± SEM (n = 8). (Non-sleep deprivation (left panel), sham-sleep deprivation (middle panel), and maternal sleep deprivation (right panel); see Table 2).
Comparison of social memory and locomotor activity parameters between all the experimental groups of male rat offspring.

Panel left: one-way ANOVA analysis comparing the non-sleep-deprived groups; Panel middle versus left: two-way ANOVA analysis comparing the non-sleep-deprived groups and sham-sleep-deprived groups; Panel right versus left: two-way ANOVA analysis comparing the non-sleep-deprived groups and maternal sleep-deprived groups (see Figures 3 and 4).
The effect of MSD and different doses of resveratrol on social memory
Sociability behavior
Two-way ANOVA and post hoc Tukey analysis confirmed that MSD significantly reduced sociability behavior in the male offspring compared to the non-sleep-deprived control group (Table 2, Figure 4(a)). Furthermore, maternal administration of resveratrol at a dose of 50 mg/kg effectively restored normal social interactions (Figure 4(a)).
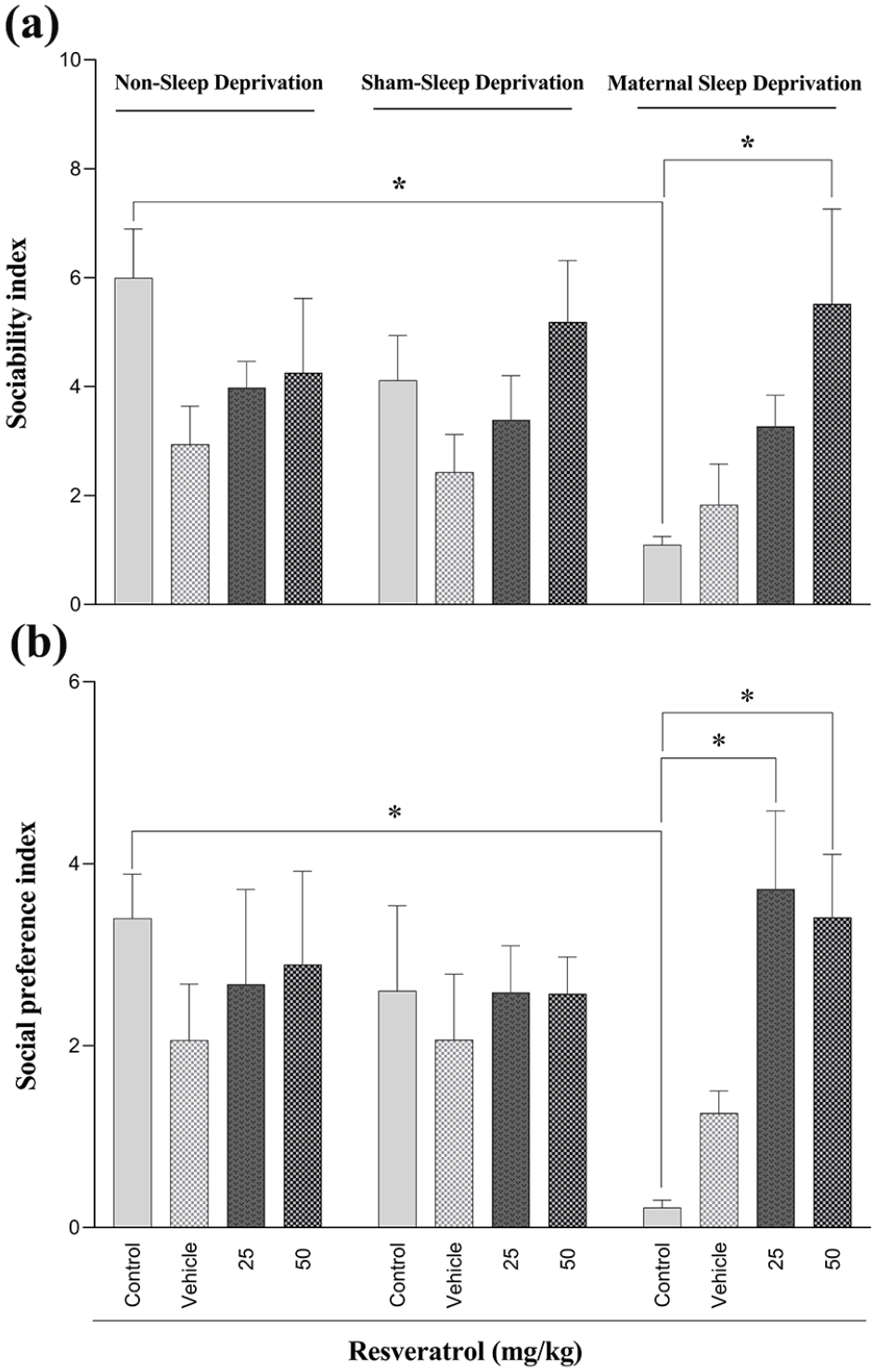
The effect of maternal sleep deprivation and different doses of resveratrol (25 or 50 mg/kg) on sociability index (a) and social preference index (b) in male Wistar rat offspring in the non-sleep-deprived, sham-sleep-deprived, and sleep-deprived groups. Values represented are mean ± SEM (n = 8). *p < 0.05 represents significant differences between the studied groups (non-sleep deprivation (left panel), sham-sleep deprivation (middle panel), and maternal sleep deprivation (right panel); see Table 2).
Social preference
Furthermore, analysis of the social preference test revealed that MSD significantly decreased the social preference index in male offspring (Table 2, Figure 4(b)). Notably, maternal resveratrol administration at both 25 and 50 mg/kg effectively mitigated this deficit, restoring social preference to control levels (Figure 4(b)).
The effect of MSD and different doses of resveratrol on hippocampal BDNF and TrkB expression
The current study investigated the effects of MSD and maternal consumption of resveratrol on the expression levels of BDNF and TrkB, which are crucial for synaptic plasticity modulation in rat offspring. The obtained results exhibit that MSD during pregnancy led to a significant reduction in BDNF expression levels in the hippocampus of male rats’ offspring, indicating an impairment in synaptic plasticity and cognitive function (Table 3, Figure 5(a)). However, administering resveratrol to the pregnant rat at a dosage of 50 mg/kg was able to effectively reverse this decline (Figure 5(a)). No statistically significant differences were observed between the sham-sleep-deprived and non-sleep-deprived groups.
Comparison of western blot and Nissl staining parameters between all the experimental groups of male rat offspring.

Panel left: one-way ANOVA analysis comparing the non-sleep-deprived groups; Panel middle versus left: two-way ANOVA analysis comparing the non-sleep-deprived groups and sham-sleep-deprived groups; Panel right versus left: two-way ANOVA analysis comparing the non-sleep-deprived groups and maternal sleep-deprived groups (see Figures 5 and 6).
BDNF: brain-derived neurotrophic factor; TrkB: tyrosine-protein kinase.
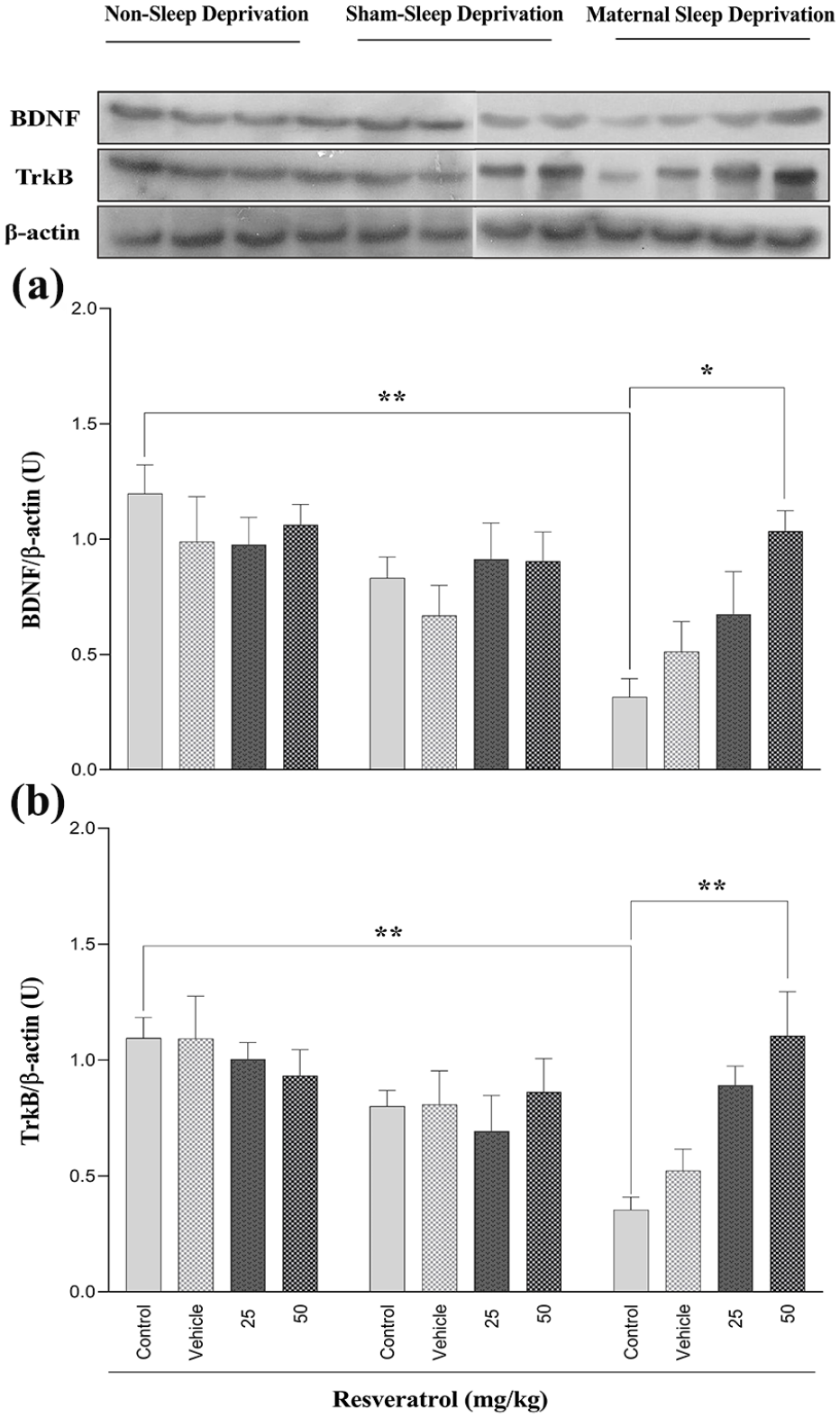
The effect of maternal sleep deprivation and different doses of resveratrol (25 or 50 mg/kg) on hippocampal BDNF (a) and TrkB (b) expression levels in male Wistar rat offspring in the non-sleep-deprived, sham-sleep-deprived, and sleep-deprived groups. Values represented are mean ± SEM (n = 8). *p < 0.05 and **p < 0.01 represent significant differences between the studied groups (non-sleep deprivation (left panel), sham-sleep deprivation (middle panel), and maternal sleep deprivation (right panel); see Table 3).
Similarly, TrkB expression was significantly downregulated in MSD-exposed offspring compared to a control group of the non-sleep-deprived groups (Table 3, Figure 5(b)). Administering resveratrol at a dosage of 50 mg/kg during pregnancy effectively prevents these detrimental effects in the signaling pathway in the hippocampal of male rats’ offspring (Figure 5(b)).
Hippocampal histology
Nissl staining was used to evaluate the impact of MSD on the hippocampal tissue of male rat offspring. As shown in Figure 6, MSD disrupted the integrity and reduced the thickness of pyramidal cells in the CA1 region of the hippocampus.
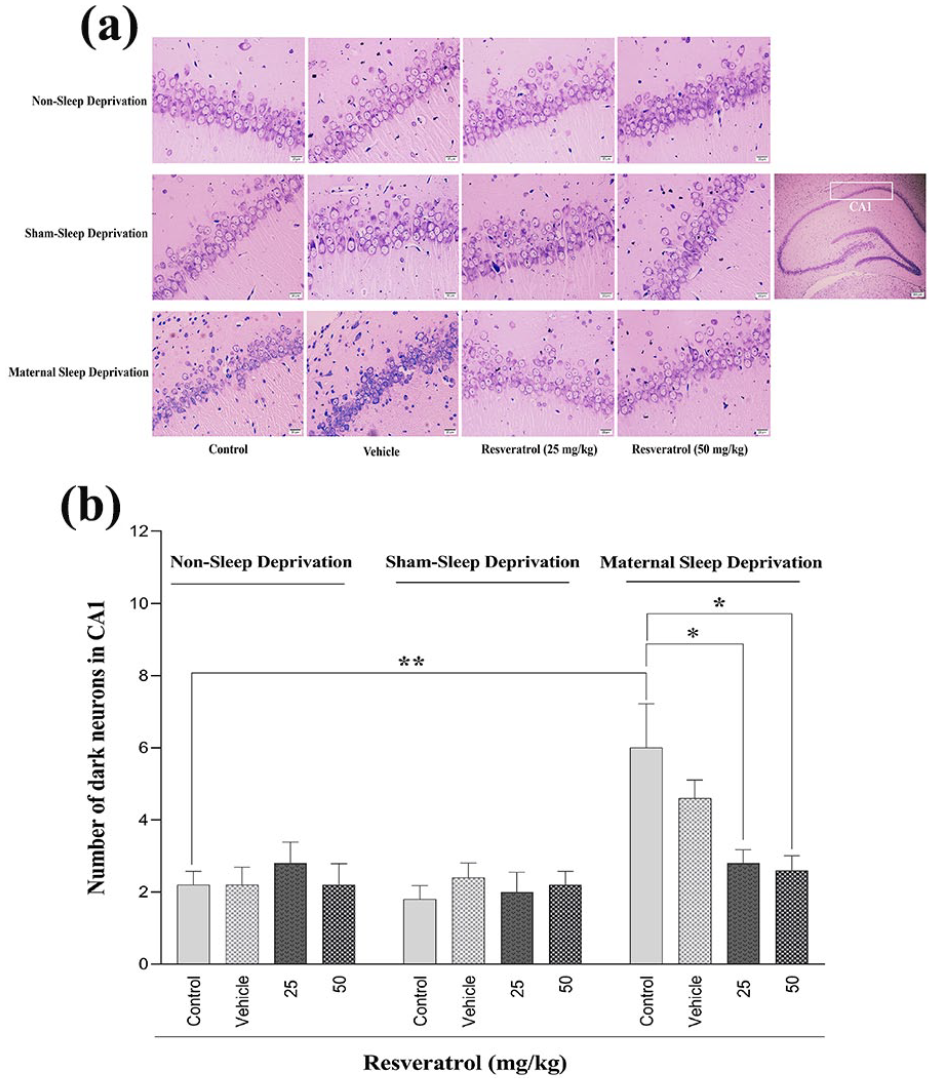
The effect of maternal sleep deprivation and different doses of resveratrol (25 or 50 mg/kg) on hippocampal morphology assessment using Nissl staining (a) and the number of dark neurons in the CA1 region (b) in male Wistar rat offspring in the non-sleep-deprived, sham-sleep-deprived, and sleep-deprived groups. Values represented are mean ± SEM (n = 8). *p < 0.05 and **p < 0.01 represent significant differences between the studied groups (non-sleep deprivation (left panel), sham-sleep deprivation (middle panel), and maternal sleep deprivation (right panel); see Table 3).
A quantitative analysis between non-sleep-deprived groups and MSD groups indicated that the number of dark neurons in CA1 was significantly increased in the control group of MSD (Table 3, Figure 6(b)). Administering resveratrol at doses of 25 or 50 mg/kg significantly reduced dark neuron density, effectively restoring hippocampal integrity to near-control levels. No significant differences were observed between the non-sleep-deprived and sham-sleep-deprived groups (Figure 6(b)).
Discussion
The present study showed that MSD during pregnancy leads to cognitive impairments in offspring, including deficits in spatial learning, memory, and social behavior, likely due to hippocampal dysfunction. In addition, the study revealed that resveratrol, a polyphenolic compound, could mitigate the detrimental effects of MSD on hippocampal function and related behaviors.
Animal studies have demonstrated that prenatal stress, including MSD, disrupts hippocampal oxidative phosphorylation, impairing spatial learning and memory in offspring (Li et al., 2020). Late pregnancy stress increases glucocorticoid levels, which cross the placenta and cause neuronal atrophy, reduced synaptic plasticity, and cognitive deficits in the fetus (Sahu et al., 2013). Since the prenatal period is a critical window for hippocampal development, disturbances such as MSD can profoundly and lastingly affect this brain region by disrupting the intrauterine environment and impairing neurogenesis (Peng et al., 2016; Wei et al., 2022; Zhao et al., 2014). One of the key mechanisms underlying these cognitive deficits is the reduction of BDNF protein expression in the hippocampus due to prenatal stress and inflammation (Guan et al., 2016).
Inflammation reduces BDNF protein expression in the hippocampus, a key contributor to cognitive deficits. Elevated cytokines such as IL-6, TNF-α, and IL-1β can cross the placenta, disrupting offspring’s brain structure and nervous system development (Shang et al., 2019; Wei et al., 2022). Acute sleep loss during pregnancy further exacerbates this state by disrupting maternal stress levels, placental immune function, and fetal brain development (Baratta et al., 2020; Zhao et al., 2014). MSD alters kynurenic acid levels, leading to neurodevelopmental changes, cognitive impairments, emotional behavior deficits, and altered hippocampal protein expression in offspring (Zhang et al., 2023b; Zhao et al., 2014).
Moreover, other research suggest that sleep deprivation exacerbates cognitive and synaptic impairments by disrupting BDNF-related miRNAs. Key miRNAs such as miR-124 and miR-132, which suppress BDNF, are upregulated during sleep loss, reducing BDNF levels in regions like the hippocampus. Simultaneously, miR-134, crucial for synaptic plasticity, becomes dysregulated, further impairing neuronal function (Davis et al., 2007; Lyons et al., 2023; Zhang et al., 2024).
Interestingly, the effects of sleep deprivation differ between rodents and humans due to variations in developmental context, biological mechanisms, and duration. In rodents, prolonged MSD increases oxidative stress and inflammation, disrupting neurogenesis and leading to long-term cognitive deficits in offspring (Wei et al., 2024). In contrast, short-term therapeutic sleep deprivation in adult humans enhances serotonin, norepinephrine, and BDNF levels, promoting neuroplasticity, suppressing hyperactive default mode network, and improving mood without negatively impacting brain development (Krause et al., 2017; Massa et al., 2011; Sochal et al., 2024; Tse et al., 2024). Thus, the differences in how sleep deprivation affects the brain in rodents and humans underscore the complexity of its impact across species and developmental stages.
Substantial evidence highlights that the hippocampal BDNF-TrkB signaling pathway plays a pivotal role in synaptic plasticity, learning, and memory. MSD disrupts this pathway, reducing BDNF expression (Zhang et al., 2023b). Insufficient sleep further reduces acetylation levels of histones H3 and H4 at BDNF promoter regions, downregulating BDNF expression (Duan et al., 2016). Several rodent studies have reported that dysregulation of the HPA axis and immune system due to stress and MSD can impair normal brain development, alter BDNF levels, and affect social interaction memory in offspring, with lasting effects into adulthood (Gur et al., 2019; Madhyastha et al., 2013; Menezes et al., 2020). Structural changes in hippocampal CA1 and CA3 regions, such as dendritic atrophy, have also been linked to prenatal stress (Soares-Cunha et al., 2018). Our findings align with previous research showing that MSD disrupts the BDNF-TrkB signaling pathway and increases neuronal loss in the hippocampus, leading to dramatic deficits in spatial and social memory performance in rat offspring. These effects were specifically attributed to MSD by including sham sleep-deprived groups, and the influence of potential confounders such as water box stress or animal handling was thus ruled out.
In light of the disruptions mentioned above caused by MSD on hippocampal development and cognitive function, resveratrol, a polyphenol known for its neuroprotective, antioxidant, and anti-inflammatory properties, has shown the potential to mitigate the adverse effects. Resveratrol’s ability to enhance synaptic plasticity, reduce neuronal apoptosis, and protect against neurodegenerative diseases such as Alzheimer’s and Parkinson’s diseases highlight its significance as a neuroprotective agent (Komorowska et al., 2024; Sun et al., 2010; Zhang et al., 2010). This polyphenol crosses the placenta and can influence fetal brain development, as evidenced by its detection in fetal circulation after maternal consumption or administration (Darby et al., 2023; Dumont et al., 2020; Lacerda et al., 2023; Madhyastha et al., 2013; Reyes-Corral et al., 2021). Such properties make it a promising candidate for counteracting the detrimental effects of prenatal stress and sleep deprivation on fetal brain development.
Resveratrol’s antioxidant properties have been linked to improved memory performance. It has been shown to modulate BDNF and TrkB expression in the cerebral cortex following ischemia-reperfusion (Ma et al., 2023; Serra et al., 2019) and to enhance cognitive function by activating the cAMP response element-binding protein (CREB-BDNF signaling pathway in the hippocampus under chronic unpredictable mild stress (Shen et al., 2018). In addition, animal studies indicated that resveratrol reduces neuronal apoptosis in the CA1 region of the hippocampus (Zhang et al., 2019) and protects against neuronal death induced by pentylenetetrazole (Meng et al., 2014). Further supporting its neuroprotective potential, a study by Zhao et al. (2020) reported that maternal resveratrol supplementation alleviated Nissl body loss in the hippocampal CA1 region of offspring exposed to fluoride, protecting them from developmental neurotoxicity and cognitive function. It also reduces oxidative stress by scavenging reactive oxygen species and enhancing antioxidant enzymes such as superoxide dismutase and glutathione peroxidase, promoting neuronal homeostasis and hippocampal neurogenesis (Navarro-Cruz et al., 2023; Azargoonjahromi et al., 2025).
Resveratrol’s anti-inflammatory effects—achieved through NF-κB inhibition and cytokine reduction help suppress microglial activation and mitigate neuroinflammation, further enhancing hippocampal health (Chung et al., 2020). Resveratrol’s anti-inflammatory and antioxidant properties also extend to mitigating BBB dysfunction in offspring, as research indicates that resveratrol can reduce BBB permeability, influence cell adhesion proteins, and enhance tight junction protein expression, which are vital for maintaining BBB integrity (Komorowska et al., 2024; Ma et al., 2023). In addition, resveratrol activates SIRT1, which is essential for CNS development and neuroprotection. It enhances stress resistance, promotes neuronal survival, supports synaptic plasticity, and contributes to cognitive function (Moraes et al., 2020; Romeo-Guitart et al., 2018). Notably, resveratrol pretreatment has been shown to prevent BDNF-TrkB downregulation by activating the Sirt1/miR-134 pathway, highlighting its neuroprotective potential (Zhang et al., 2024). In this regard, our findings showed that resveratrol restores the BDNF-TrkB signaling pathway, mitigating the adverse effects of MSD on offspring neurodevelopment and cognitive function.
Furthermore, our study highlights that a 50 mg/kg dose of resveratrol effectively reversed MSD-induced hippocampal impairments, including deficits in spatial learning, memory, neuronal loss, and reduced BDNF-TrkB levels. In contrast, the 25 mg/kg dose provided partial benefits but was less consistent in its outcomes, suggesting it may not be enough to activate essential neuroprotective pathways fully. This indicates a potential dose threshold for optimal neuroprotection, with higher doses of resveratrol necessary to reliably enhance synaptic plasticity and cognitive function.
In conclusion, the current study investigates an MSD model and highlights its long-lasting neurodevelopmental effects on offspring, which persist into adulthood and impair behavior. Administering resveratrol intraperitoneally during pregnancy significantly improves memory performance and reduces neuronal loss in rat offspring. This improvement is attributed to restoring hippocampal proteins related to synaptic plasticity and memory performance, such as BDNF and TrkB, which are negatively affected by MSD. By targeting these pathways, resveratrol substantially alleviated learning and memory deficits in offspring exposed to MSD during gestation. As a natural polyphenol compound, resveratrol offers several advantages that enhance its therapeutic potential, including a low risk of side effects, cost-effectiveness, wide availability in natural sources, and an excellent safety profile. These attributes position resveratrol as a promising candidate for the development of innovative therapeutic interventions, particularly in the context of prenatal neurodegenerative disorders and cognitive impairments. While these findings provide valuable insights, further research is needed to evaluate the safety and effectiveness of resveratrol in humans. Furthermore, future investigations should focus on understanding how resveratrol affects various brain regions influenced by MSD.
Limitations
The absence of phospho-TrkB expression measurements in our analysis is a limitation, as phospho-TrkB, the active form of the TrkB receptor, serves as a key marker of BDNF signaling activation. Its assessment would provide a more precise evaluation of TrkB receptor activity and the downstream signaling processes that regulate synaptic plasticity, neurogenesis, and cognitive function. Another limitation of the current study was the lack of evaluation of the action mechanism of resveratrol, such as its antioxidant and anti-inflammatory properties. Since these resveratrol effects have been proven in various studies and the association of BDNF/TrkB to these pathways has been identified, we have not investigated these mechanisms. However, incorporating these analyses would offer deeper insights into resveratrol’s neuroprotective mechanisms during MSD.
Future directions
This study provides compelling evidence for resveratrol’s neuroprotective effects in mitigating cognitive and hippocampal deficits caused by MSD. However, advancing these findings into clinical applications requires further investigation. Clinical trials are essential to evaluate resveratrol’s safety, bioavailability, and effectiveness during pregnancy. Identifying the optimal dosage and assessing the long-term impact on maternal and fetal health will be pivotal. Exploring the molecular mechanisms underlying resveratrol’s neuroprotective properties will also enhance our understanding of how it counters MSD-induced neurodevelopmental impairments. Combining resveratrol with other neuroprotective agents may also improve its therapeutic potential. Future research should also extend beyond the hippocampus to examine resveratrol’s impact on different brain regions affected by MSD.
Supplemental Material
sj-docx-1-jop-10.1177_02698811251334034 – Supplemental material for Therapeutic effects of resveratrol on memory deficits in offspring of sleep-deprived rats: Involvement of hippocampal BDNF-TrkB pathways
Supplemental material, sj-docx-1-jop-10.1177_02698811251334034 for Therapeutic effects of resveratrol on memory deficits in offspring of sleep-deprived rats: Involvement of hippocampal BDNF-TrkB pathways by Mohammad-Hossein Mohammadi-Mahdiabadi-Hasani, Maryam Farahmandfar, Mohammad-Reza Zarrindast, Mohammad Nasehi, Anahita Torkaman-Boutorabi and Gholamreza Hassanzadeh in Journal of Psychopharmacology
Supplemental Material
sj-docx-2-jop-10.1177_02698811251334034 – Supplemental material for Therapeutic effects of resveratrol on memory deficits in offspring of sleep-deprived rats: Involvement of hippocampal BDNF-TrkB pathways
Supplemental material, sj-docx-2-jop-10.1177_02698811251334034 for Therapeutic effects of resveratrol on memory deficits in offspring of sleep-deprived rats: Involvement of hippocampal BDNF-TrkB pathways by Mohammad-Hossein Mohammadi-Mahdiabadi-Hasani, Maryam Farahmandfar, Mohammad-Reza Zarrindast, Mohammad Nasehi, Anahita Torkaman-Boutorabi and Gholamreza Hassanzadeh in Journal of Psychopharmacology
Footnotes
Acknowledgements
We extend our gratitude to the Tehran University of Medical Sciences for providing financial support for this study. We also acknowledge the collaboration of the Institute for Cognitive Science Studies (ICSS) and the Cognitive Science and Technology Council of Tehran, Iran.
Author contributions
Mohammad-Hossein Mohammadi-Mahdiabadi-Hasani: Methodology, Project administration, Investigation, Software, and Writing – Original Draft. Maryam Farahmandfar: Funding acquisition, Supervision, Conceptualization, Resources, and Writing – Review & Editing. Mohammad-Reza Zarrindast: Conceptualization, Writing – Review & Editing, Supervision. Mohammad Nasehi: Supervision, Conceptualization, Software, Writing – Review & Editing. Gholamreza Hassanzadeh: Software and Writing – Review & Editing. Anahita Torkaman-Boutorabi: Writing – Review & Editing.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The study was funded by the Vice Chancellor of Research Affairs, Tehran University of Medical Sciences. This manuscript is extracted from the PhD thesis of Mohammad-Hossein Mohammadi-Mahdiabadi-Hasani (Academic Grant No. 54994).
ORCID iDs
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.