
Abstract
Background:
Recent evidence suggests that psychedelics can induce rapid and long-lasting antidepressant effects. The generally acknowledged explanation for these traits is the phenomenon of neuroplasticity, although the exact underlying molecular mechanisms remain unclear.
Aims:
This study investigates the effects of psilocin, lysergic acid diethylamide (LSD) and N,N-dimethyltryptamine (DMT) on synaptogenesis and immediate early genes (IEGs) expression in direct comparison with ketamine, fluoxetine and lithium after acute (1 h) and/or prolonged (24 h) treatment in vitro.
Methods:
Rat primary cortical cultures were treated with 10 µM psilocin, 1 µM LSD, 90 µM DMT, 1 µM ketamine, 10 µM fluoxetine and 5 mM lithium. Analysis of synaptic puncta was performed; puncta of presynaptic marker synapsin I/II, postsynaptic density protein 95 (PSD-95) and their co-localization (established synapse) were assessed 24 h after drug treatment. Next, expressions of IEGs encoding activity-regulated cytoskeleton-associated protein (Arc), early growth response 1 (Egr1), and neuronal PAS (Per-Arnt-Sim) domain protein 4 (Npas4) were analysed 1 and 24 h after drug treatments.
Results:
Psilocin increased synaptic puncta count and induced Arc expression. The effect to promote synaptogenesis was comparable to ketamine and lithium; ketamine additionally increased PSD-95 puncta count. LSD and DMT did not induce any significant effects. Interestingly, fluoxetine had no effect on synaptic puncta count, but upregulated Egr1 and Npas4.
Conclusions:
Psilocin demonstrated synaptogenic effects comparable to those of ketamine and lithium, and acutely upregulated IEG Arc expression, adding another piece of evidence to its profile as a promising therapeutic agent.
Introduction
Neuroplasticity, the brain’s ability to modify synaptic connections in response to experience, plays a fundamental role in neuropsychiatric disorders. A growing body of research suggests a correlation between maladaptive neuroplastic mechanisms, such as synaptic remodelling, dendritic atrophy and impaired neurogenesis with neuropsychiatric disorders, such as depression, anxiety, post-traumatic stress disorder, addiction, anorexia and chronic pain, often exhibited as atrophy of neurons in the cortical and limbic brain regions (Rădulescu et al., 2021; Serafini, 2012; Soeiro-de-Souza et al., 2012). Given the central role of neuroplasticity in these disorders, therapeutic strategies that restore the mechanisms involved have been a primary focus of psychiatric research.
The selective serotonin reuptake inhibitors, targeting and blocking serotonin transporter (SERT), represent the most widely used class of antidepressants. They have been suggested to modulate neuroplasticity in the adult cortex, potentially reopening a plasticity window similar to that observed in juvenile brains (Maya Vetencourt et al., 2008; Umemori et al., 2018). However, direct evidence linking SERT blocker-induced neuroplasticity to therapeutic outcomes in neuropsychiatric disorders remains limited. While some studies have shown SERT blocker-induced plasticity effects in the visual cortex, their role in reshaping synaptic connections in emotional regulation circuits remains an area of active investigation (Casarotto et al., 2021, 2022; Rosas-Sánchez et al., 2024; Winkel et al., 2021). Lithium, a drug interacting with enzymes, is primarily used as a mood stabilizer in bipolar disorder but also as an augmentation strategy option in the treatment of depression. Similarly to SERT blockers, it has been shown to effectively enhance neuroplasticity by promoting neuroprotection, glioprotection, elevated adult neurogenesis and synaptic plasticity (Diniz et al., 2013; Gray and McEwen, 2013; Hammonds and Shim, 2009; Puglisi-Allegra et al., 2021; Won and Kim, 2017).
Although these medications have been the mainstay of treatment for depression, and despite their neuroplastic effects, they possess several drawbacks; the antidepressant effects of SERT blockers take 6–8 weeks of adequate doses to become evident, various side effects can appear, and many patients are unresponsive to this first-line treatment. In line with the delayed onset of antidepressant actions, the neuroplastic changes take place only after chronic administration. While in the case of lithium, its therapeutic use is constrained by its narrow safety margin and adverse effects, including nephrotoxicity, thyroid and parathyroid dysfunction, weight gain, and thus high discontinuation rate (Antoniadou et al., 2018; Cameron et al., 2023; Gitlin, 2016). These drawbacks highlight the need for alternative treatments with more rapid and sustained effects on neuroplasticity.
In contrast to common antidepressants, psychedelics and ketamine exhibit rapid and robust therapeutic effects, even in treatment-resistant depression. Consequently, after years of marginalization, research on their pharmacology is gaining robust support regarding their potential to treat a plethora of neuropsychiatric conditions including depression, anxiety, post-traumatic stress disorder, addiction, anorexia or chronic pain (Cameron et al., 2023; Carhart-Harris and Goodwin, 2017).
While classic psychedelics and ketamine act on distinct receptor systems (serotonergic and glutamatergic, respectively), they share significant features beyond being psychoactive. They are thought to promote structural and functional plasticity in the prefrontal cortex (PFC), hippocampus and other brain regions involved in emotional processing, by rapidly increasing dendritogenesis, spinogenesis and synaptogenesis. Hence, they induce downstream network reconfiguration through complex neuroplastic mechanisms (Davoudian et al., 2023; de la Fuente Revenga et al., 2021; Krystal et al., 2019; Li et al., 2010; Lu et al., 2021; Ly et al., 2018, 2021; Moda-Sava et al., 2019; Moliner et al., 2023; Phoumthipphavong et al., 2016; Raval et al., 2021; Shao et al., 2021; Vargas et al., 2023).
Depending on the specific compound and dosage regimen, sustained antidepressant effects have been observed. For instance, repeated ketamine infusions have demonstrated prolonged therapeutic benefits in patients with treatment-resistant depression (Siegel et al., 2021; Zheng et al., 2018, 2019). Similarly, both single high doses and dose-escalation regimens have demonstrated long-lasting effects. For 5-MeO-DMT, individualized dosing with up to three doses of the compound (GH001) on a single day was superior to single-dose administration in providing potent and ultra-rapid antidepressant effects, while high-dose psilocybin resulted in persistent positive psychological effects (Nicholas et al., 2018; Reckweg et al., 2023). The duration of neuroplastic effects has also been linked to the duration of intoxication, as shown in psilocybin and ketamine models (Nardou et al., 2023). Furthermore, set (mindset) and setting (environment) significantly influence the therapeutic outcome of psychedelics, and as such, guidelines to support safe therapeutic use of these drugs have been published (Johnson et al., 2008).
At the molecular level, immediate early genes (IEGs) are a family of genes that are rapidly and transiently transcribed after a wide variety of signals. As opposed to the ‘late response’ genes, IEGs expression does not depend on any precedent protein synthesis. They encode many functionally distinct proteins, such as transcription and growth factors, cytoskeletal proteins or signalling molecules, and play an essential role in cellular responses that contribute to long-term neuronal plasticity.
Of these, activity-regulated cytoskeleton-associated protein (Arc), early growth response 1 (Egr1) and neuronal PAS (Per-Arnt-Sim) domain protein 4 (Npas4) are three IEGs strongly linked to synaptic plasticity processes and expression of all three genes has been previously shown to be induced by various psychedelics (Desouza et al., 2021; González-Maeso et al., 2003, 2007; Lerer et al., 2024; Levy et al., 2019; Nichols and Sanders-Bush, 2002; Pei et al., 2000; Villalba et al., 2024). Arc is a postsynaptic effector protein localized primarily in the dendrites of excitatory glutamatergic neurons of the cortex and hippocampus. Arc has a key role in long-term plasticity, as it is involved in an inactivity-dependent redistribution of synaptic weights on the synapse in the process of inverse synaptic tagging (Okuno et al., 2018; Zhang and Bramham, 2021). Arc modulates dendritic spine morphology and excitatory synaptic strength by coordinating the regulation of α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) trafficking. It also indirectly associates with F-actin and helps to stabilize the primary cytoskeleton of dendritic spines.
Egr1 is a synaptic activity-inducible zinc finger transcription factor linked to long-term synaptic plasticity. Following synaptic activation, it directly regulates Arc by binding to its promoter and activating its transcription (Li et al., 2005). Egr1 also acts as a transcription suppressor of postsynaptic density protein 95 (PSD-95) and modulates N-methyl-D-aspartate receptor-dependent AMPAR trafficking (Qin et al., 2015). Egr1 is specifically involved in synaptic organization at multiple levels, resulting in synaptic stabilization.
Finally, Npas4 is a plasticity-associated transcription factor with a homeostatic ability. After its rapid expression elicited by an excitatory activity, it regulates the genes such as brain-derived neurotrophic factor that trigger formation and maintenance of inhibitory synapses on excitatory neurons, toning down neuronal firing events and that is directly involved in neuroplasticity induced by psychedelics (Lin et al., 2008; Moliner et al., 2023; Sun and Lin, 2016). Npas4 expression is selectively activated after a membrane depolarization-induced increase in intracellular calcium concentration and occurs selectively in neurons (primarily in the excitatory type).
Despite the exact pathways activated by psychedelics have been under intense investigation in recent years, to the best of our knowledge, no study has directly compared these promising agents to regular psychiatric medication in terms of their neuroplastic effects. In this study, psilocin, DMT and LSD were examined and compared to two of the most prescribed psychiatric drugs, fluoxetine and lithium, as well as with ketamine, a generally acknowledged psychoplastogen, in the ability to promote synaptogenesis and to enhance the above specified gene expression of selected IEGs Arc, Egr1 and Npas4.
Methods
Primary cortical culture isolation and maintenance
All experiments were conducted in accordance with the guidelines of Directive 2010/63/EU of the European Parliament and approved by the Animal Care and Use Committee of the Ministry of Health (reference number MZDR 17637/2020-4/OVZ) in line with the Animal Protection Code of the Czech Republic 246/1992. NIMH is authorized for the use of experimental animals (accreditation ref. no. MZE-51088/2021-13114).
Primary cortical cultures were prepared as previously described by Jorratt et al. (2023) with minor modifications. Briefly, embryos were extracted from time-mated female Wistar rats on embryonic day 18. The cortices were dissociated under the stereomicroscope VisiScope® 250 (VWR, USA) and put in ice-cold Dulbecco’s phosphate-buffered saline (DPBS, Gibco, 14190144). Then, the cortices were washed thrice with ice-cold DPBS and mechanically dissociated with a 0.9-mm needle (B. Braun, 4657519), followed by a 0.45-mm needle (B. Braun, 4657683), each needle thrice. The dissociated supernatant was transferred into a seeding medium containing advanced Dulbecco’s modified Earle’s medium (Gibco, 12491015) supplemented with 10% foetal bovine serum (FBS, Biowest, S1810-500), 1% antibiotic-antimycotic solution (Gibco, 15240062) and 1% GlutaMAX supplement (Gibco, 35050061) and filtered twice through a 100-µm cell strainer (VWR, 732–2759). The singularized cells were counted using a haemocytometer and seeded on poly-L-lysine (Merck, P1274-25MG) pre-coated well plates. 24-well plates with inserted 13-mm, #1.5 coverslips (VWR, 631-0150) with a seeding density of 50,000 cells/well were used for immunocytochemistry (ICC), and 6-well plates with a density of 500,000 cells/well were used for real-time quantitative polymerase chain reaction (qPCR). The cells were cultured in a humidified incubator at 37°C and 5% CO2. On the next day, the seeding medium was replaced with neurobasal medium (Gibco, 21103049), enriched with 2% B27 supplement (Gibco, 17504044), 1% antibiotic-antimycotic, and 1% GlutaMAX. Half of the growth medium was changed twice a week.
Drugs and chemicals
On day in vitro 17, the cultures were treated for 24 h for the ICC analysis of colocalization, and 1 and 24 h for the qPCR analysis of changes in IEGs expression. The cells were treated with 10 µM psilocin (THC Pharm, Germany), 1 µM LSD fumarate (Alfarma, Czechia), 90 µM DMT fumarate (Forensic Laboratory of Biologically Active Substances, University of Chemistry and Technology, Czech Republic), 10 µM fluoxetine hydrochloride (Molekula Group, Czechia), 5 mM lithium chloride (Merck, Germany) and 1 µM ketamine (Merck, Germany). The doses were selected based on a previously published study by Ly et al. (2018) and our pilot data. All drugs were of ⩾99% purity and were diluted in sterile distilled water. Sterile distilled water was used as a negative control (Ctrl).
Immunocytochemistry
Cells were quickly washed with pre-warmed (37°C) phosphate-buffered saline (PBS) and fixed with pre-warmed 4% paraformaldehyde in PBS for 4 min, then rinsed 3 min × 5 min with PBS. Next, blocking and permeabilization were done in one step with 10% FBS, 0.1% Triton X-100 in PBS for 45 min at RT. The coverslips were subsequently incubated with primary antibodies chicken anti-MAP2 (1:10,000, Abcam, ab5392), mouse anti-PSD-95 (1:500, Abcam, ab192757) and guinea-pig anti-synapsin I/II (1:1000, Synaptic Systems, 106004) at 4°C overnight. The following day, coverslips were washed 3 min × 5 min with PBS and incubated with secondary antibodies AlexaFluor (AF) 647 conjugated donkey anti-chicken (703-605-155), AF488 donkey anti-guinea pig (706-545148) and AF594 donkey anti-mouse (715-545-151) for 1 h at room temperature in the dark. All secondary antibodies were purchased from Jackson ImmunoResearch and diluted 1:500 for working solutions. After the incubation, coverslips were washed 3 min × 5 min with PBS in the dark and rinsed with distilled water, dried from excess liquid, mounted onto microscope slides using ProLong™ Gold Antifade Mountant with DAPI (Invitrogen, P36941) and allowed to cure overnight. All the antibodies were diluted in the staining solution consisting of 0.1% Triton X-100 in PBS.
Image acquisition and analysis
Images were acquired with a confocal laser scanning microscope, Leica SP8 X, on an inverted microscope scope, Leica DMi8 (Leica Microsystems, Germany). Plan apochromat oil objective Leica 63×/1.40 CS2 was used. The confocal pinhole was set to Airy1, and bidirectional scanning with line averaging four times was applied for Z-stacks at a 0.3 μm Z-step size. The acquisition was done in 8-bit, 1024 × 1024 pixels, and 2.6× digital zoom. For every experiment, samples were imaged under identical software settings. Automatic threshold selection was performed, and binary pictures of proximal dendrites were analysed using the Synapse Counter plugin (Dzyubenko et al., 2016) for Fiji (version 1.52e, Schindelin et al., 2012), following a protocol from Li et al. (2016). The plugin identifies the pre- and post-synaptic puncta separately and detects their spatial overlap, termed colocalization. The region of interest for the plugin was set at 32 μm × 5 μm and placed 10 μm from the soma.
Changes in IEGs expression
Cells were lysed in RNA Blue reagent (Top-Bio, R013) and phenol-chloroform-based extraction of the homogenate was performed according to the manufacturer’s instructions. Complementary DNA (cDNA) was generated using the same amount of each total RNA sample within one batch. High-capacity cDNA Reverse Transcription Kit (Applied Biosystems, 4368814) was used according to the manufacturer’s instructions. qPCR was performed with CFX96 Touch Real-Time PCR Detection System (Bio-Rad Laboratories, USA) using Luna Universal qPCR Master Mix (New England Biolabs, M3003L) according to the manufacturer’s instructions. IEGs signal was normalized to the average of two housekeeping genes, glyceraldehyde-3-phosphate dehydrogenase (Gapdh) and peptidylprolyl isomerase A (Ppia), which were selected as an optimal reference via the NormFinder tool (version 0.953, Andersen et al., 2004) and according to the literature research (Lerer et al., 2024; Martin and Nichols, 2016; Nogueira et al., 2024). The comparative Ct (ΔΔCt) method was used to determine the changes in mRNA levels of Arc, Egr1 and Npas4 (Livak and Schmittgen, 2001).
The primers were designed in the NCBI Primer-BLAST tool (Ye et al., 2012) or selected based on literature (Adams et al., 2017; Heroux et al., 2018) and the sequences were chosen as follows: Arc (5′-GCAGAAAGCGCTTGAACTTG-3′, 5′-AGCGGGACCTGTACCAGAC-3′), Egr1 (5′-CATGCAGATTCGACACTGGAAG-3′, 5′-GTATGCTTGCCCTGTTGAGTCC-3′), Npas4 (5′-GCCACTATGTCTTCAAGCTCT-3′, 5′-CTGCATCTACACTCGCAAGG-3′), Gapdh (5′-GGCATGGACTGTGGTCATGA-3′, 5′-CAACTCCCTCAAGATTGTCAGC-3′), Ppia (5′-TCCTTTCTCCCCAGTGCTCAG-3′, 5′-TCAACCCCACCGTGTTCTTC-3′).
Data and statistical analysis
Statistical analyses were performed in GraphPad Prism (version 8.0.1, GraphPad Software, Inc., USA). Outliers were removed using the robust regression and outlier removal (ROUT) method with a 1% Q-value. Data violating the assumption of normality (Shapiro–Wilk test) were normalized by square root transformation. Homogeneity of variance was tested using the Brown–Forsythe test. One-way analysis of variance (ANOVA) was used to model the relationship, with each substance as an independent predictor. We used Dunnett’s post hoc test to perform the comparison of drug effects to Ctrl.
Results
Psilocin induces synaptogenesis
Synaptic puncta were identified by the co-localization of synapsin I/II and PSD-95 puncta. Analysis of changes in synapsin I/II density did not show any effect of the treatment (Figure 1(a)). Significant changes were observed in PSD-95 puncta density after the treatment (F(6, 330) = 5.55, p < 0.001); the post hoc analysis revealed the prominent effect of ketamine (p < 0.001, Figure 1(b)). Analysis of co-localized (i.e. synaptic) puncta also displayed a significant effect of the drug treatment (F(6, 333) = 8.60, p < 0.001), and further post hoc analyses revealed a strong effect of psilocin, lithium and ketamine (all p < 0.001, Figure 1(c)).
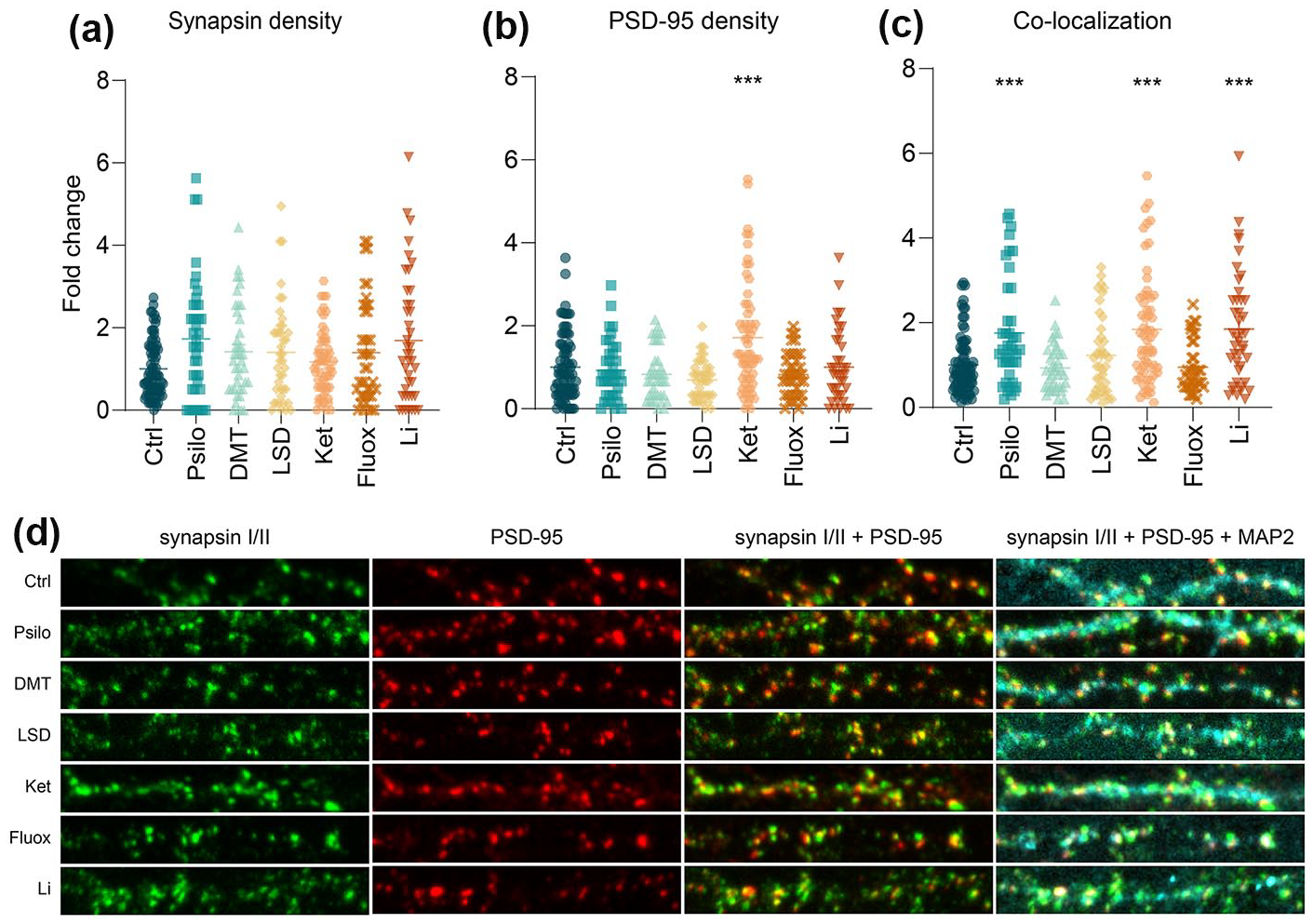
The effect of selected psychoplastogens on synaptogenesis. (a–c) Quantification of presynaptic density (a Synapsin puncta), postsynaptic density (b PSD-95 puncta) and synapse density (c Synapsin and PSD-95 co-localized puncta) after 24 h drug treatment of primary cortical neurons. n = 38–79 randomly chosen neurons per treatment from at least three biological replicates. One-way ANOVA followed by Dunnett’s post hoc test was performed, and data are presented as mean fold change ± SEM.
Psilocin induces gene expression of Arc after acute treatment
After 1 h (Figure 2(a)), Arc expression was significantly affected by the treatment (F(6, 24) = 4.73, p < 0.01), and post hoc analysis showed a significant effect of psilocin (p < 0.01). Expression of Npas4 was also significantly changed after 1 h (F(6, 26) = 2.55, p < 0.05), although post hoc test did not reveal any specific drug treatment effect.
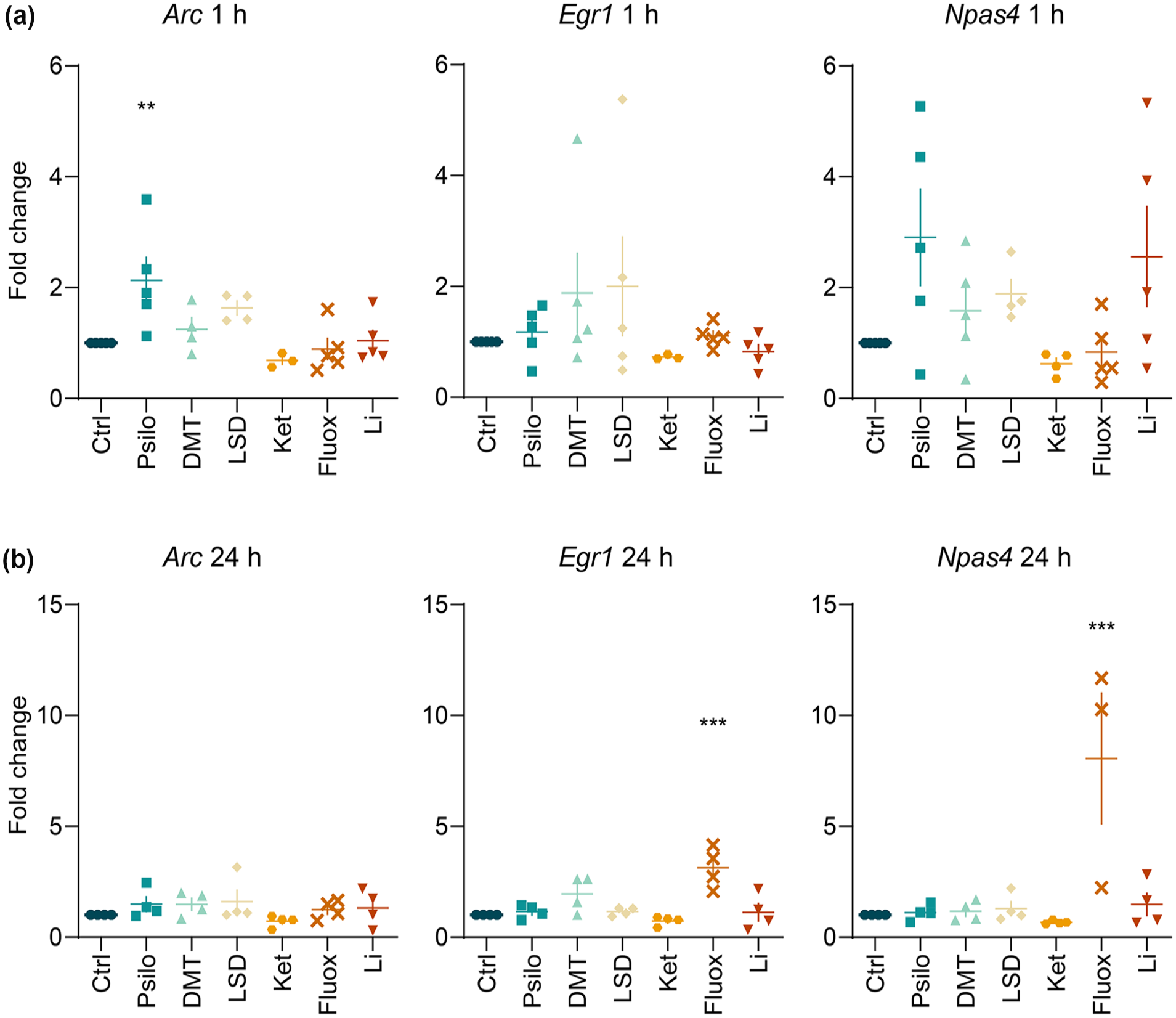
Effect of selected psychoplastogens on the expression of IEG mRNA. (a + b) Assessment of the changes in mRNA expression of IEG Arc, Egr1 and Npas4 after 1 h (a) and 24 h (b) drug treatment of primary cortical neurons. Data were collected from at least four biological replicates. One-way ANOVA followed by Dunnett’s post hoc test was performed. Data are presented as mean fold change ± SEM.
After 24 h (Figure 2(b)), Egr1 levels were significantly affected by the treatment (F(6, 21) = 8.34, p < 0.001), and the post hoc test showed a significant effect of fluoxetine (p < 0.001). Npas4 expression was also significantly altered by the treatment (F(6, 20) = 7.59, p < 0.001), and the post hoc test revealed prominent changes following fluoxetine treatment (p < 0.001).
Discussion
The main finding of our study is that psilocin significantly promotes synaptogenesis and expression of the IEG Arc, and the effects of psilocin were comparable to those of lithium and ketamine. Surprisingly, we did not observe such effects after LSD and DMT treatment. Concerning the positive control treatments, ketamine also increased PSD-95 density, and fluoxetine did not affect synaptic density, but upregulated Egr1 and Npas4 expression.
Our finding that psilocin promotes structural neuroplasticity is in line with other studies (Du et al., 2023; Moliner et al., 2023; Raval et al., 2021; Vargas et al., 2023). In accordance with Ly et al. (2018), we did not observe any effect on PSD-95 density after the treatment with DMT, but we discovered the increased PSD-95 and synapse densities associated with ketamine treatment, which was described also in other studies (Li et al., 2010; Ly et al., 2021). Interestingly, Ly et al. (2018) also observed the effect of LSD on presynaptic and synaptic puncta count, as opposed to our results. The fact that we did not find any significant effect of LSD and DMT on synaptogenesis was unexpected. The explanation could be that we did not add dimethyl sulfoxide (DMSO) into the drug solution for the elimination of any noxious effect on the primary cells. However, since DMSO is a permeation enhancer, its lack in the solution might cause the non-significant results for LSD and DMT in our study. Another explanation may lie in the fact that we used LSD and DMT in the form of fumarate salts to ensure better solubility and prolonged stability in the growth medium compared to a free base. However, in light of recent data suggesting that the ability of psychoplastogens to induce structural plasticity depends on crossing the cell membrane and activating the intracellular pool of serotonin 2A receptors (Vargas et al., 2023), it seems that LSD and DMT may have been inefficient in inducing neuroplasticity in our study because of the fumarate moiety. As for fluoxetine, a frequently used SERT blocker, its ineffectiveness in elevating synaptic density in our study could stem from the necessity to be applied chronically rather than in one acute treatment (Barone et al., 2018; Zahrai et al., 2020).
In the study of changes in IEGs mRNA expression after psychedelic treatment, we found a significant upregulation of Arc after the acute treatment with psilocin, and of Egr1 and Npas4 following the prolonged treatment with fluoxetine. In contrast to our study, 90-min psilocybin treatment did not exhibit any significant changes in Arc in the PFC, but decreased expression in the hippocampi of rats (Jefsen et al., 2021). Another animal study demonstrates upregulated Arc expression following 90-min LSD treatment in PFC, but not in the hippocampus (Nichols and Sanders-Bush, 2002). Davoudian et al. (2023) describe comparable elevation of IEG Cfos in many brain regions after psilocybin and ketamine administration, with selected brain regions manifesting drug-preferential differences. Concerning Egr1, it has been shown that an acute dose of psychedelics increases its expression (Desouza et al., 2021; Liu et al., 2023), while chronic microdosing does not (Cameron et al., 2019). A study using noribogaine observed upregulated Egr1 and Npas4 expression in male mice with serotonin 2A receptor (5-HT2AR) knockout, Npas4 expression in 5-HT2AR knockout females, and Egr1 expression in wild-type females, indicating sex and genotype differences in IEGs expression (Villalba et al., 2024). In this study, Npas4 was clearly upregulated in a 5-HT2AR activation-independent manner. On top of that, noribogaine has a polypharmacological profile and does not aim directly at 5-HT2AR, as opposed to psychedelics. Another reason for the divergent results with the aforementioned studies could lie in our in vitro model, which comprises the mixture of cortical cells from both male and female embryos. This aspect may be necessary to consider for the following studies focusing on IEGs, as it could be the cause of major variances.
Overall, our study provides additional support for the hypothesis that psilocin is a potent neuroplastic agent. The co-localization results also indicate similar mechanisms by which psilocin, ketamine and lithium promote their neuroplastic effect on the synapse. On the other hand, our findings also suggest that fluoxetine’s neuroplastic effects are produced through different mechanisms than those of serotonergic psychedelics or ketamine.
Footnotes
Author contributions
All authors made a significant contribution to the design, acquisition, analysis or interpretation of data for this study. AP and TP designed the project, YV and KS carried out the experiments, YV, KS and IK wrote the manuscript. VK performed the dissections and supervised the ICC assay, KŠ, VM and MN carried out the statistical analyses, JP trained the experimenters in confocal microscopy and supervised the setup, MK and RJ synthesized and provided the group with ketamine, TP and ZB supervised the project and manuscript writing. All authors gave final approval for the current version of this work to be published. All authors agree to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Data availability
All data supporting the described findings can be obtained from the corresponding author upon reasonable request.
Declaration of conflicting interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: TP declares having shares in ‘Psyon s.r.o.’ and in ‘Společnost pro podporu neurovědního výzkumu s.r.o’ and together with MK in AVI-X Aviation Experts s.r.o.. TP founded ‘PSYRES’ – Psychedelic Research Foundation, and he reports consulting fees from GH Research and CB21-Pharma outside the submitted work. TP is/was also involved in clinical trials of Compass Pathways with psilocybin, MAPS clinical trial with MDMA, Ketabon clinical trial with KET01 (slow-reelease oral formulation of ketamine) and GH-Research clinical trial with 5-MeO-DMT outside the submitted work.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This work was supported by the grants from Czech Ministry of Education, Youth and Sports (Inter-Excellence Inter-Action sub-programme LUAIZ24146), Czech Science Foundation (project 20-25349S), the Czech Health Research Council (project NU21-04-00307), Long-term conceptual development of research organization (RVO 00023752), Specific University Research, Czech Ministry of Education, Youth and Sports (project 260648/SVV/2024), Charles University research program Cooperatio-Neurosciences, Grant Agency of Charles University in Prague, Czech Republic (project GAUK 430122) and private funds obtained via PSYRES – Psychedelic Research Foundation (psyresfoundation.eu).