
Abstract
Background:
Impulsivity may be defined as a heterogeneous construct characterized by difficulties in inhibiting actions and premature decision-making. Although a high level of impulsivity is recognized as a risk factor for cannabis use disorder, the effects of cannabinoids on impulsive choices have been less explored.
Aims:
This study aimed to determine the acute and persistent effects of CB1/2 agonism on impulsive choice in rats using a delay-discounting task (DDT).
Methods:
Trained adult male Sprague-Dawley rats were injected with either vehicle or increasing doses (0.1, 1, and 5 mg/kg) of the CB1/2 agonist WIN 55212-2 and tested in the DDT.
Results:
Our results showed that the effect of WIN55212-2 correlated with baseline impulsivity and reduced impulsivity in rats classified as high impulsive, while no effect was observed in rats classified as low impulsivity. Two weeks after the last WIN55212-2 injection, the rats were injected with vehicle and re-exposed to DDT. Rats classified as high impulsive maintained a significantly high AUClog value, suggesting a long-lasting effect of WIN 55212-2. Finally, the CB1 receptor antagonist rimonabant (1 mg/kg) reversed the effect of repeated treatment with WIN55212-2 on impulsivity in the high-impulsive population, suggesting an active role of the CB1/2 receptor in the persistent effect of WIN55212-2.
Conclusions:
Our results suggest a potential benefit of CB1/2 agonism in vulnerable subpopulations with high levels of impulsivity. To maximize therapeutic benefits and minimize potential iatrogenic effects, assessing choice impulsivity and other variables is essential, aligning with personalized medicine principles to effectively tailor interventions.
Introduction
Impulsivity may be defined as a heterogeneous construct characterized by difficulties in inhibiting actions and premature decision-making (Pattij and Vanderschuren, 2008). Although impulsive behaviors have enabled individuals to adapt successfully to rapidly unexpected environmental changes, exacerbated impulsive behaviors in the long term are counterproductive (Fenneman and Frankenhuis, 2020). In this sense, maladaptive impulsivity is a characteristic of several neuropsychiatric disorders, such as attention-deficit/hyperactivity disorder (ADHD) and substance use disorder (Hamilton et al., 2015; Setlow et al., 2009; Vassileva and Conrod, 2019).
Impulsive behaviors may be assessed in preclinical models with two dimensions: impulsive action and impulsive choice. Impulsive action may be defined by a deficit in motor inhibition and failure in attentional processes, whereas impulsive choice is understood as an intolerance to the delay of rewards or delay aversion (Pattij and Vanderschuren, 2008). Typically, impulsive choice can be assessed in clinical and preclinical contexts using a delay-discounting task (DDT). The DDT evaluates the choice of a larger delayed reward versus a small immediate reward; as the delay of a large reward increases, its subjective value decreases. Impulsive individuals tend to exhibit higher delay aversion, resulting in a higher choice of a small immediate reward over a larger delayed reward (Jentsch et al., 2014; Marshall et al., 2014; Mazur, 1987). Considering the multidimensional nature of impulsivity, impulsive choice, assessed in the DDT, has also been classified as “waiting impulsivity,” because it requires waiting to make a favorable choice (Dalley and Robbins, 2017). Previous evidence has demonstrated that impulsive choice is a trait that can predict future maladaptive behavior. Preclinical data show that higher levels of impulsive choice predict a lack of inhibition of drug seeking and higher sensitivity to relapse induced by cues (Diergaarde et al., 2008). In addition, clinical longitudinal research has shown that higher levels of impulsive choices predict a higher consumption of cannabis among adolescents (Kim-Spoon et al., 2019; Rinehart and Spencer, 2021).
Cannabis is the third most popular substance of abuse, following nicotine and alcohol, with an estimated 192.2 million people having used cannabis at least once in the last year (United Nations, 2018). Furthermore, recent regulations on cannabis use in Canada and some states in the United States have led to a decrease in perceptions of the risks and negative consequences associated with its use among the general population. Delta-9-tetrahydrocannabinol (THC), the primary psychoactive compound in cannabis, activates the cannabinoid CB1 receptor in the brain and induces significant effects on higher cognitive functions, such as attention, behavioral flexibility, time perception, and working memory (Pertwee, 2014).
Previous evidence has shown the complex effects of CB1 agonists on impulsivity, which vary according to task, protocol treatment, and research model. In humans, acute exposure to THC increases impulsivity in tasks related to inhibitory control but does not alter delay aversion (McDonald et al., 2003). However, it has been observed that daily, but not occasional, cannabis use increases impulsive choice in DDT (Petker et al., 2021). In addition, Sofis et al. (2020) showed that a higher frequency of cannabis use is associated with higher levels of impulsive choice.
Preclinical studies have reported mixed findings regarding the effect of CB1 receptor activity on impulsive behavior. Pattij et al. (2007) showed that CB1 antagonists decrease impulsive action without modifying impulsive choices. In addition, the same study indicates that the CB1/2 receptor agonist WIN55212-2 did not modify the impulsive action and choice (Pattij et al., 2007). By contrast, infusion of CB1 agonists into the orbitofrontal cortex (OFC) increases impulsive choice (Fatahi et al., 2018). It has recently been proposed that the mixed effect observed after CB1 receptor activation could be dependent on individual differences in baseline levels of impulsivity (Rinehart and Spencer, 2021). In this sense, previous evidence has shown that stimulants may reduce impulsive decision-making in individuals with high impulsivity induced by an isolated environment (Perry et al., 2008). In fact, it has been proposed that “high impulsive individuals might reduce their impulsivity to a higher degree than low impulsive individuals, after marihuana use” (Ucha et al., 2019). To address this hypothesis, we tested the effects of WIN 55212-2 in DDT individually and determined whether cannabinoid effects on impulsive choice depended on the baseline impulsivity of each subject.
Experimental procedures
Animals
Fourteen male Sprague-Dawley rats, approximately postnatal day (PND) 25, were obtained from the Animal Care Faculty of Biological Science, Pontificia Universidad Católica de Chile (Charles River, Wilmington, MA, USA), under the strict supervision of a veterinarian. The rats were maintained in the Animal Care Faculty of the Department of Pharmacy, Pontificia Universidad Católica de Chile. All rats were housed in pairs per cage and kept at room temperature (22°C–24°C) on a 12 h light/dark cycle (lights on at 7:00 EST) with access to water ad libitum. Rats were handled for 1 week before starting the treatment. Six days after arrival (PND 31), food access was restricted. Rats were weighed daily to maintain their body weight at 90% of their free-feed body weight. We permitted an increase in the restricted body weight per week to allow age-associated growth. The rats started with an approximate weight of 75 g and weighed approximately 420 g at the end of the experiment. The target weights were adjusted according to the normal growth curves obtained from the animal supplier. All procedures were carried out in strict accordance with the guidelines published in the “National Institute of Health (NIH) Guide for the Care and Use of Laboratory Animals” (8th edition) and the principles presented in the “Guidelines for the Use of Animals in Neuroscience Research” by the Society for Neuroscience.
Reagents
The CB1/2 agonist, WIN 55212-2 mesylate (Sigma-Aldrich, St. Louis, MO, USA), and CB1 antagonist, Rimonabant (MedChemExpress, Monmouth Junction, NJ, USA), were emulsified in 2% Tween 80 (Sigma-Aldrich, St. Louis, MO, USA) in saline solution 0.9%.
Apparatus
Sessions were performed in ventilated, sound-attenuating enclosures containing an operant conditioning chamber with an interior space of 31 cm × 24 cm × 21 cm (Med Associates, Inc., St. Albans, VT, USA). The front door and the rear panel were made of clear polycarbonate, and both ends were made of aluminum panels. The right panel was equipped with two response levers, horizontally aligned 11.5 cm apart. Above each lever, there was a translucent circle transilluminated with white light, 2.5 cm in diameter. The pellet receptacle was centrally located between the two levers. Forty-five milligrams of food chocolate pellet (F0299 chocolate dustless precision pellets, Bio-serv, Flemington, NJ, USA) was delivered to this receptacle from an external food dispenser. The panel on the opposite side of the chamber was equipped with a house light located centrally near the top of the chamber. Data were collected using the MED-PC IV software and a PC-compatible interface (Med Associates, Inc., St. Albans, VT, USA).
Lever press training
The rats were tested using the behavioral procedures described below. All procedures were adapted from Moreno et al. (2021). After PND 35, a shaping program was used to train the rats to begin pressing the response lever (Figure 1(a)). Both levers were extended at session onset. The delivery was programmed according to a 3-min fixed-time schedule, in which a food pellet was dispensed every 3 min. Furthermore, when the lever was pressed, an additional pellet was delivered. Sessions were finished after either 60 min had elapsed or 100 pellets had been delivered. The criterion for advancement to the next phase of training was 100 lever presses performed for three consecutive sessions.
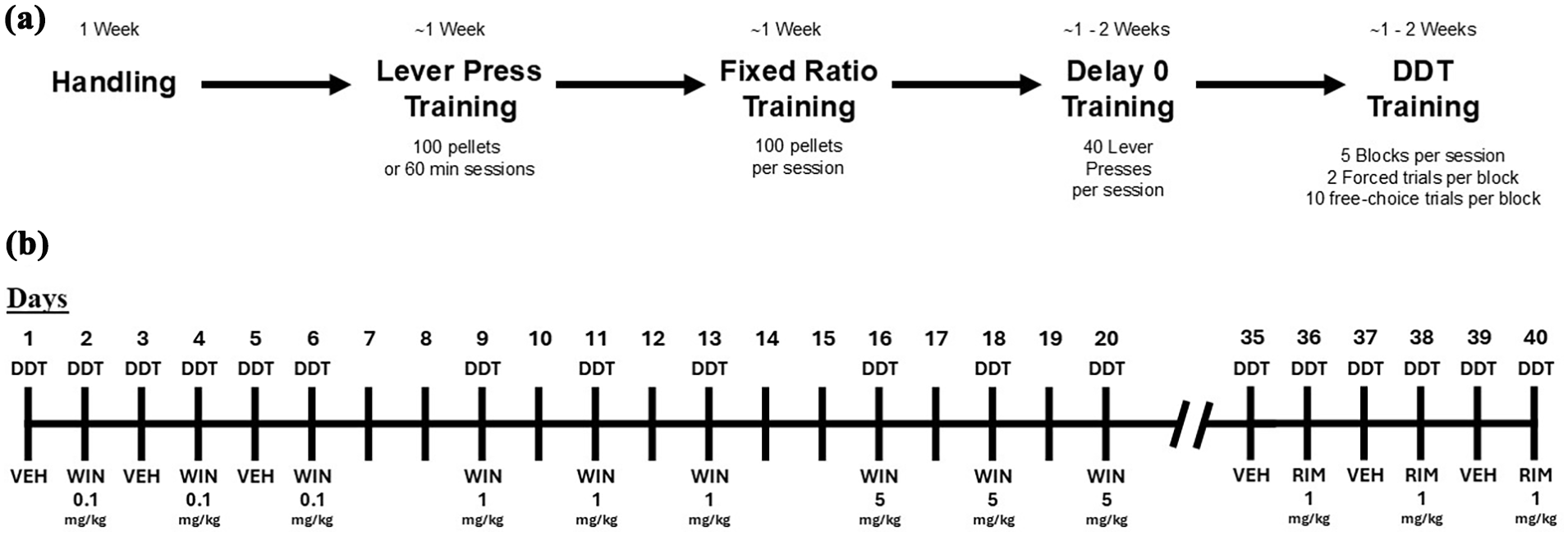
Training and experimental protocol. (a) Summary of training protocol before the delay-discounting task. (b) Experimental schedule and treatment protocol used before each delay-discounting task.
Fixed ratio training
The second phase of training started between PND 39 and 40 (Figure 1(a)). In this phase, only one lever was extended, and the associated cue light was activated. After five lever presses, the extended lever was retracted, and the opposite lever was extended. Each lever press resulted in the delivery of a food pellet following a fixed ratio 1 schedule. After receiving 100 pellets delivered over 3 consecutive sessions, the rats progressed to the next training phase.
Delay 0 training
This training phase began around PND 45 (Figure 1(a)). This phase consisted of 40 trials per session. Two levers with two different reinforcer amounts were presented and were accompanied by an illuminated cue light: the lever press of the larger reinforcer (LR) resulted in three pellets, and the small reinforcer (SR) lever resulted in one pellet. There was no delay in the delivery of either the small or large reinforcers in this training. The session ended after 40 lever presses between the large and SRs. The criterion for advancement to the next phase of training required a preference proportion for large rewards higher than 0.8.
DDT training
The DDT began in darkness, with the levers retracted (blackout). The session started 10 min after the blackout, and then the house light was turned on. Each session consisted of 5 blocks of 12 trials each. The duration of each trial was 60 s. The first two trials of each block were forced-choice trials, in which the LR or SR lever was alternately introduced. If the rat pressed the SR lever, one pellet was immediately delivered, and the lever was retracted. If the LR lever was activated, the stimulus light blinked during the delay, and three pellets were delivered following the delay. Then, 10 free-choice trials followed the force trials in each block. At the beginning of the free-choice trial, the lights over the levers were turned on, and the levers appeared. If the lever was pressed, both levers were retracted. If the rat pressed the SR lever, one pellet was immediately delivered, and both levers were retracted. If the LR lever was chosen, the stimulus light blinked during the delay, and three pellets were delivered following the delay. Delays to LR were increased from 0 s in Block 1, 4 s in Block 2, 8 s in Block 3, 16 s in Block 4, and 32 s in Block 5. Failure to choose a response within 20 s resulted in the levers retracting, and the trial was considered an omission. The rats were trained five times per week for approximately 3–4 weeks until stable performance was achieved before assessing the effect of vehicle or WIN55212-2 administration. The criterion for stability was three consecutive DDT tests with a variation of less than 10% on the AUClog.
Drug administration protocol
Rats were injected with vehicle and increasing doses of the CB1/2 agonist WIN 5212-2 (0.1, 1, and 5 mg/kg) interspersed, as illustrated in Figure 1(b). Five minutes after each injection, the rats were subjected to a DDT to determine their impulsive choice behavior. Two weeks after the last injection of WIN, the rats were injected with vehicle and rimonabant (1 mg/kg), as illustrated in Figure 1(b), and subjected to DDT. Each treatment test was performed three times, and the average value was used for the data analysis.
Data analysis
Delay-discounting analysis
Delay-discounting curves were obtained by plotting the proportion of LR choices related to the total choices of each block versus each delay(s) (Figure 2(a), 4(a), 4(b), 5(a), and 5(b)). The area under the delay-discounting curve (AUC) was used to measure impulsive choices. The AUC was calculated using a logarithmic transformation of delays (AUClog) to avoid disproportionate contributions at long delays in the delay-discounting curves (Borges et al., 2016). Specifically, each delay was transformed into the logarithm of the delay and normalized by dividing by the largest logged delay. To avoid the problem of calculating the logarithm of zero, a small amount (value = 1) was added to each delay before the transformation. GraphPad Prism was used to calculate the AUClog (GraphPad Software, Inc., La Jolla, CA, USA). Higher AUClog values indicate lower discounting by delay (i.e., preference choice for larger rewards, indicating lower impulsivity), whereas lower AUClog values correspond to higher discounting by delay (i.e., preference choice for small and immediate rewards, indicating higher impulsivity; Myerson et al., 2001).
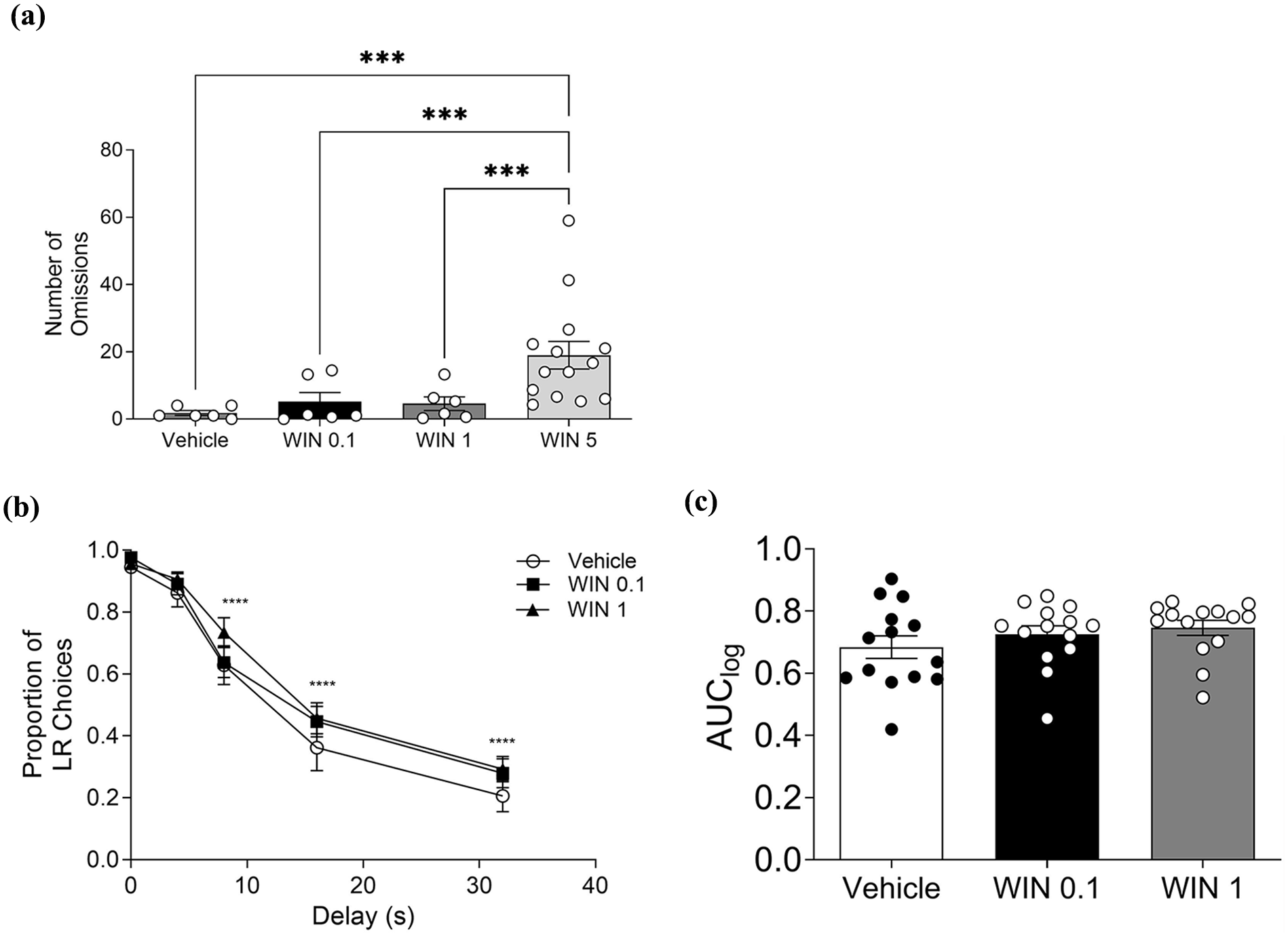
Acute effect of WIN55212-2 on the delay-discounting task. (a) Number of omissions during DDT. (b) Delay-discounting curve. (c) AUClog of each treatment group (Vehicle, WIN 55212-2, 0.1 and 1 mg/kg). Data bars represent mean ± SEM. One-way ANOVA, Tukey test post-hoc; ****p < 0.0001 compared to delay 0.
Statistical analyses
All statistical analyses were performed using the GraphPad Prism 9.4 software (GraphPad Software, Boston, MA, US). The resulting data were analyzed using one-way and two-way analysis of variance (ANOVA) with repeated measures when appropriate. Normality was assessed using the Shapiro–Wilk test, and sphericity was assumed. Post hoc analyses were performed using a paired Tukey test. Correlation analyses were performed using Spearman’s correlation coefficient. Rats were classified as low impulsive and high impulsive based on the median split of AUClog. All data are reported as mean ± standard error of the mean (SEM). The significance level was set at p < 0.05.
Results
Effects of WIN in DDT
One-way ANOVA showed a significant effect of WIN 55212-2 treatment on the number of omissions (Figure 2(a), F3, 15 = 10.76; p = 0.0005). Post hoc analyses revealed that the dose of 5 mg/kg of WIN 55212-2 increases the number of omissions compared to vehicle (VEH vs WIN 5: p = 0.0002), 0.1 mg/kg (WIN 0.1 vs WIN 5: p = 0.001), and 1 mg/kg of WIN 55212-2 (WIN 0.1 vs WIN 5: p = 0.0008). Given the high number of omissions (Figure 2(a)) and a preference proportion for large rewards lower than 0.8 at delay 0 s (Supplemental 1), a dose of 5 mg/kg was excluded from the impulsive choice analysis. Considering the delay-discounting curves (Figure 2(b)), two-way ANOVA showed a significant effect of delay (F4, 52 = 153.9; p < 0.0001) without a treatment effect (F2, 26 = 1.63; p = 0.21). According to AUClog obtained from the delay-discounting curve (Figure 2(b)), one-way ANOVA did not show a significant effect of WIN (Figure 2(c), F2, 26 = 1.734; p = 0.196).
Acute treatment of WIN 55212-2 induces differential effects on impulsivity behavior depending on basal impulsivity
To evaluate whether the effects of WIN 55212-2 on impulsive choice depend on baseline impulsivity, a correlation analysis was performed between the AUClog of rats when treated with a vehicle and the change in AUClog (ΔAUC = WIN AUClog – Vehicle AUClog) after the WIN 55212-2 injection. Interestingly, the ΔAUC for both doses of the CB1/2 agonist (0.1 and 1 mg/kg) showed a significant negative linear correlation with baseline impulsive choice (Figure 3(a) and (b), WIN 0.1: r = −0.685, p = 0.007; WIN 1: r = −0.799, p = 0.0006).
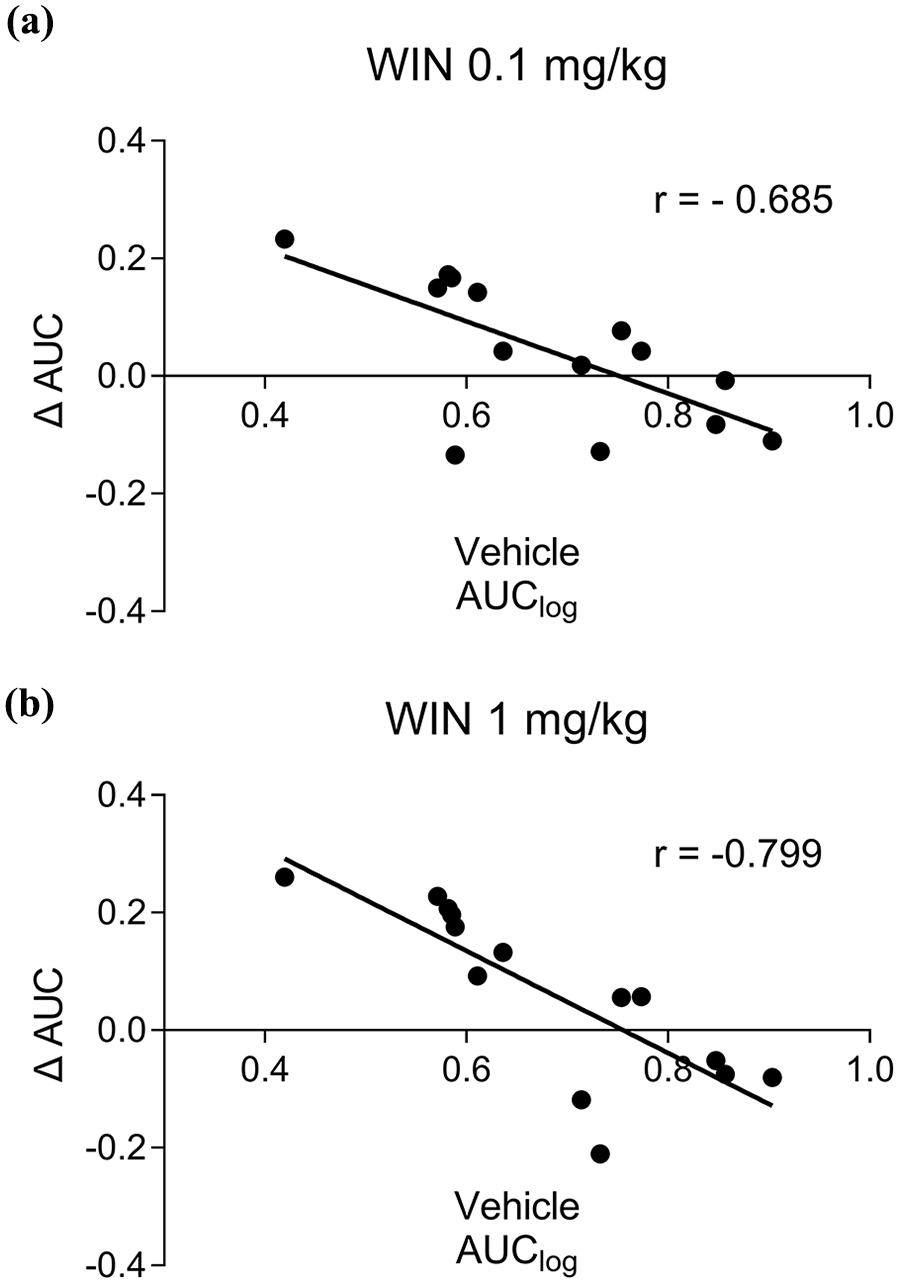
Individual differences in the effects of WIN 55212-2. (a) Correlation between ΔAUC with WIN 55212-2 treatment at 0.1 mg/kg and AUClog with vehicle treatment. A significant negative correlation was observed (r = −0.685; p = 0.007 according to Spearman correlation; n = 14). (b) Correlation between ΔAUC with WIN 55212-2 treatment at 1 mg/kg and AUClog with vehicle treatment. A significant negative correlation was observed (r = −0.799; p = 0.006 according to Spearman correlation; n = 14).
To further analyze the individual differences in the effect of WIN, rats were classified as low impulsive and high impulsive based on the median split of the AUClog. Considering the delay-discounting curves (Figure 4(a) and (b)), two-way ANOVA showed a significant effect of delay in both low-impulsive (F4, 24 = 68.52; p < 0.0001) and high-impulsive rats (F4, 24 = 109.8; p < 0.0001). Interestingly, WIN 55212-2 treatment showed a significant effect (F2, 12 = 9.03; p = 0.004) and interaction with delays (F8, 48 = 3.62; p = 0.002) only in high-impulsive rats (Figure 4(b)). Considering the AUClog, a two-way ANOVA showed a significant effect of baseline impulsivity (Figure 4(c), F1, 12 = 7.08; p = 0.021) and an interaction between the WIN 55212-2 treatment and classification (Figure 4(c), F2, 24 = 12.57; p = 0.0002). A post hoc analysis corroborated that vehicle AUClog was higher in the low-impulsive group than in the high-impulsive group (VEH/low impulsive: 0.797 ± 0.027 vs VEH/high impulsive: 0.57 ± 0.026; p < 0.0001). In addition, 1 mg/kg WIN 55212-2 increased AUClog values in high-impulsive rats (VEH/high impulsive: 0.57 ± 0.026 vs WIN 0.1/high impulsive: 0.68 ± 0.04; p = 0.146; vs WIN 1/high impulsive: 0.76 ± 0.017; p = 0.001), but did not modify the AUClog values in low-impulsive rats (VEH/low impulsive: 0.797 ± 0.027 vs WIN 0.1/low impulsive: 0.77 ± 0.031; p = 0.714; vs WIN 1/low impulsive: 0.737 ± 0.047; p = 0.21).
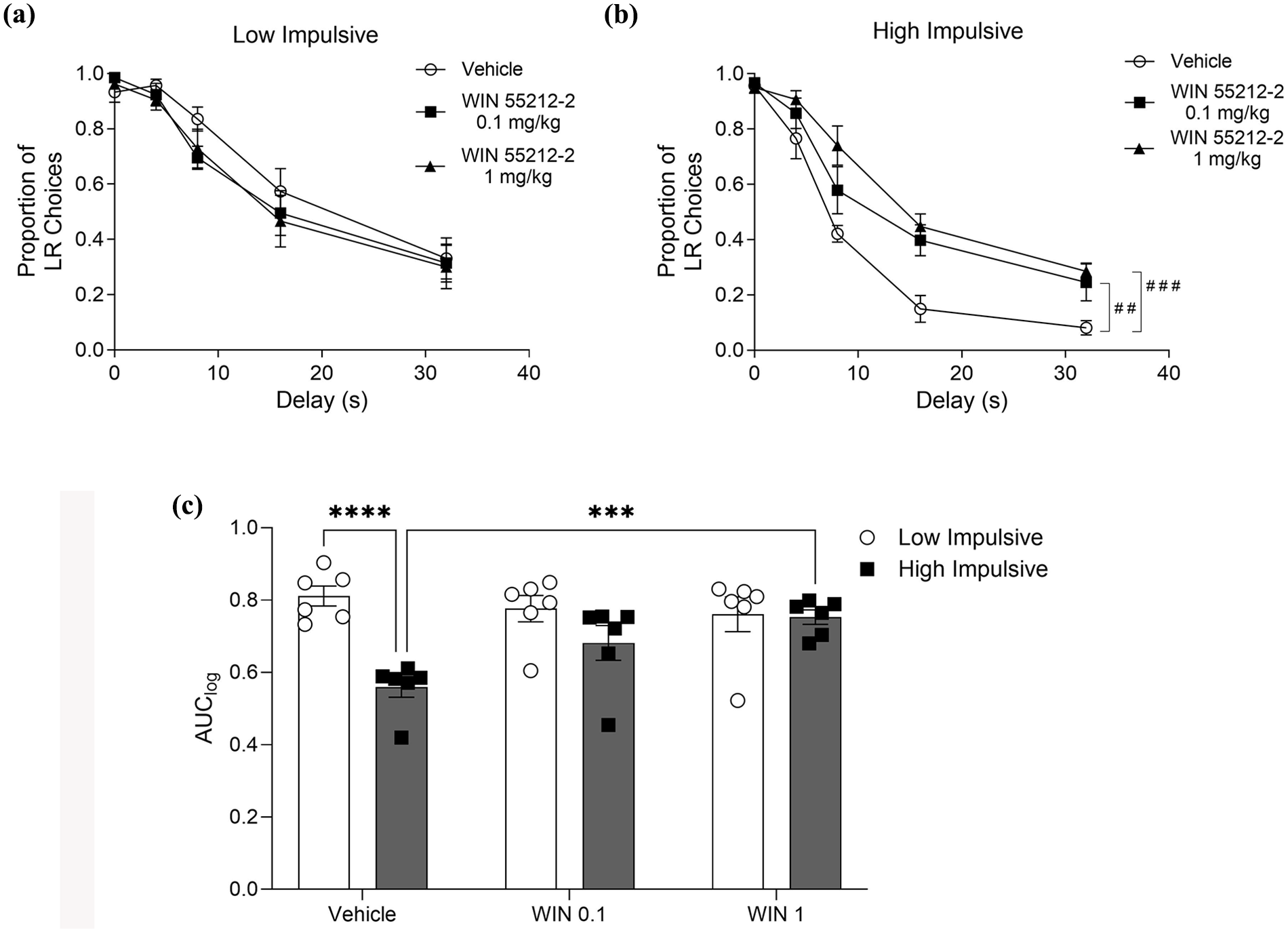
Baseline impulsive choice defines individual differences in WIN 55212-2 treatment. (a) Proportion of large reward choices in each delay trial during DDT of the low-impulsive subpopulation treated with Vehicle, WIN 55212-2 at 0.1 and 1 mg/kg. (b) Proportion of large reward choices in each delay trial during DDT of the high-impulsive subpopulation with treatment of Vehicle, WIN 55212-2 at 0.1 and 1 mg/kg. (c) AUClog of low- and high-impulsive subpopulations for each treatment group (Vehicle, WIN 55212-2, 0.1 and 1 mg/kg). Data bars correspond to mean ± SEM, two-way ANOVA, Tukey test post hoc; ***p < 0.001; ****p < 0.0001; ##p < 0.01; ###p < 0.001 compared to vehicle/delay 32 s.
Differential persistent effects of WIN 55212-2 depend on the CB1 receptor
Two weeks after the last injection of the CB1/2 agonist, DDT was conducted to assess the persistent effects of WIN55212-2 treatment on impulsive choice. Although no significant differences were found when comparing pre- and post-treatment impulsive behaviors in the whole sample (p = 0.0776; see Supplemental 2), we observed a differential persistent effect of WIN 55212-2 when analyzing the subpopulations (Figure 5(a) and (b)). Considering the delay-discounting curves (Figure 5(a) and (b)), a two-way ANOVA showed a significant effect of delay in both low-impulsive (F4, 24 = 76.74; p < 0.0001) and high-impulsive rats (F4, 24 = 85.09; p < 0.0001). In addition, a significant effect of the treatment in both low-impulsive (F2, 12 = 10.03; p = 0.003) and high-impulsive rats (F2, 12 = 11.28; p = 0.002) was observed. Two-way ANOVA showed a significant effect of treatment (Figure 5(c), F2, 24 = 18.71; p < 0.0001), baseline impulsivity (F1, 12 = 8.48; p = 0.013), and interaction (F2, 24 = 4.3; p = 0.025). Post hoc analysis revealed a significant difference in AUClog after WIN 55212-2 treatment, but only in the high-impulsive rats (VEH Pre-treatment/high impulsive: 0.57 ± 0.026 vs VEH Post-treatment high impulsive: 0.73 ± 0.04; p = 0.0058), suggesting a long-lasting effect of WIN 55212-2 treatment. By contrast, low-impulsive rats did not exhibit persistent effects of WIN 55212-2 treatment (VEH Pre-treatment/low impulsive: 0.79 ± 0.027 vs VEH Post-treatment/low impulsive: 0.78 ± 0.044; p = 0.944). Rimonabant was injected to assess the role of the CB1 receptor in the persistent effect of WIN 55212-2 on impulsive choice. First, one-way ANOVA analyzing the overall population of rats showed a significant effect of treatment (Supplemental 2, F2, 24 = 14.92; p < 0.0001). Post hoc analysis revealed a significant decrease in the AUClog after the injection of rimonabant compared to pre-treatment (VEH Pre-treatment: 0.68 ± 0.036 vs Rimonabant: 0.55 ± 0.032; p = 0.023) and post-treatment vehicle injection (VEH Post-treatment: 0.76 ± 0.032 vs Rimonabant: 0.56 ± 0.033; p < 0.0001). When the effect of CB1 receptor blockade was analyzed in the subpopulations, a significant decrease in the AUClog was observed after rimonabant injection in both low-impulsive rats (Figure 5(c), VEH Post-treatment/low impulsive: 0.78 ± 0.04 vs Rimonabant/low impulsive: 0.596 ± 0.05; p = 0.0012) and high-impulsive rats (VEH Post-treatment/high impulsive: 0.73 ± 0.04 vs Rimonabant/high impulsive: 0.52 ± 0.04, p = 0.0005).
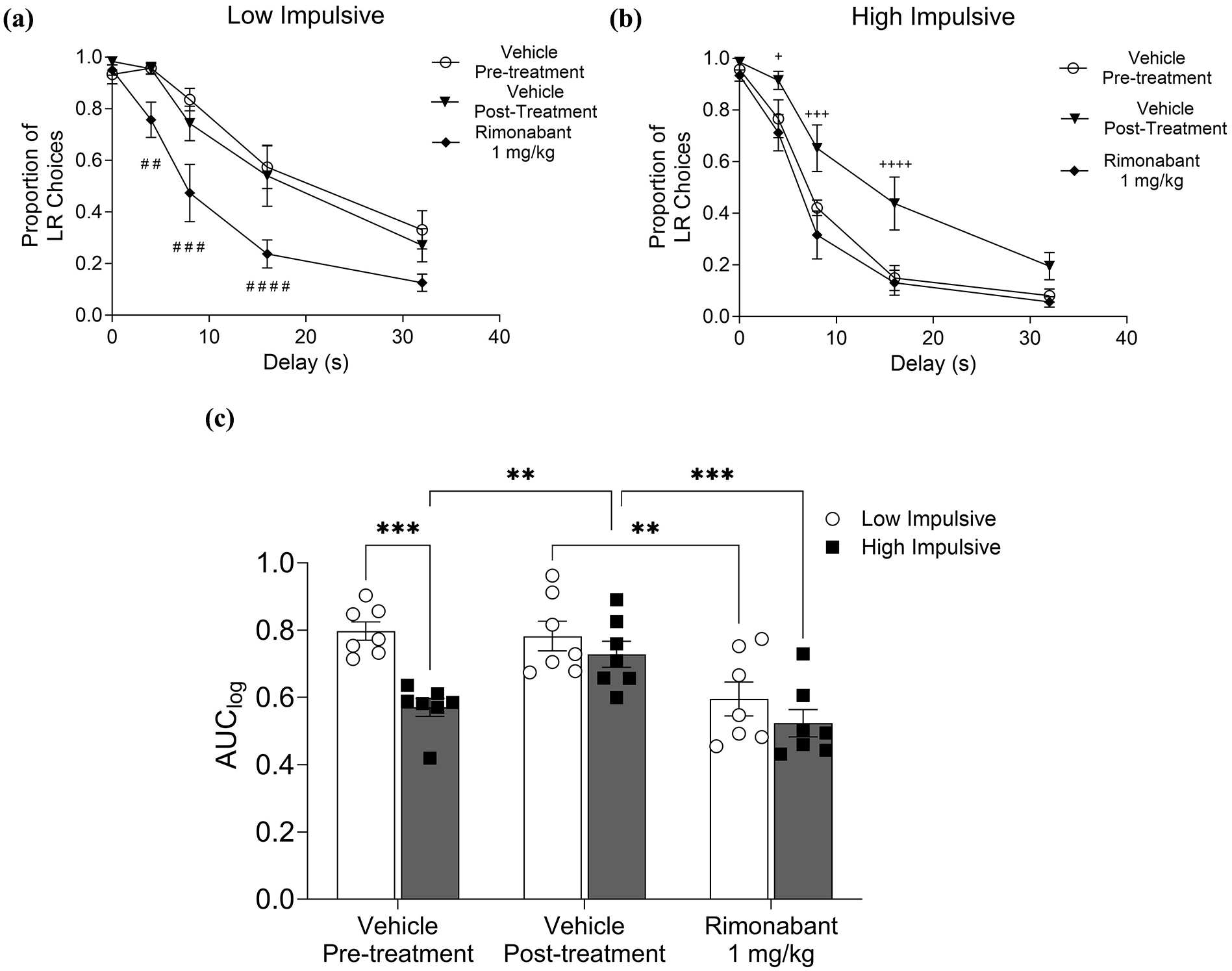
Persistent effect of WIN55212-2 on impulsive choice depends on the CB1 receptor. (a) Proportion of large reward choices in each delay trial during DDT of the low-impulsive subpopulation in vehicle pre-treatment, vehicle post-treatment condition, and with rimonabant 1 mg/kg. (b) Proportion of large reward choices in each delay trial during DDT of the high-impulsive subpopulation in vehicle pre-treatment, vehicle post-treatment condition, and with rimonabant 1 mg/kg. (a) AUClog of low- and high-impulsive subpopulations for each treatment condition (Vehicle pre-treatment, Vehicle post-treatment, and Rimonabant 1 mg/kg). Data bars correspond to mean ± SEM, two-way ANOVA, Tukey test post hoc; **p < 0.01; ***p < 0.001. ##p < 0.01; ###p < 0.001; ####p < 0.0001 compared to Vehicle pre-treatment and Vehicle post-treatment. +p < 0.05; +++p < 0.001; ++++p < 0.0001 compared to Vehicle Pre-treatment and Rimonabant 1 mg/kg.
Discussion
New regulations on cannabis use in Europe, Canada, Uruguay, and some states of the United States have boosted research into its therapeutic properties and targets related to the endocannabinoid system (Cox, 2018; Howard and Osborne, 2020; Hutchison et al., 2019; Schauer et al., 2023; Zarrabi et al., 2020). CB1 agonists, in particular, have shown therapeutic potential for several conditions, including epilepsy, anxiety, glaucoma, and cancer (Cairns et al., 2016; Chakravarti et al., 2014; Moreira et al., 2009; Pérez-Valenzuela et al., 2023; Rosenberg et al., 2017). However, the effects of CB1 agonists on impulsive choice are complex and depend on factors such as the frequency of exposure, dosage, and individual differences in basal levels of impulsivity. These differences may contribute to variations in responses to CB1 agonists (Rinehart and Spencer, 2021). Our results showed a dose-dependent reduction in impulsive choice induced by WIN 55212-2 among high-impulsive rats. By contrast, rats classified as low impulsive did not exhibit changes in impulsive choice following acute injection of WIN-55212-2. Since our results indicate a baseline impulsivity-dependent effect, we discuss both the potential benefits and deleterious consequences of CB1/2 agonist exposure. Our study provides new evidence highlighting both the acute and persistent effects of WIN 55212-2 on impulsive choice. Furthermore, the observed persistent effects on impulsive choice depend on the endogenous CB1 receptor activation.
First, we assessed the acute effects of different doses (0.1, 1, and 5 mg/kg) of the CB1/2 agonist WIN55212-2 on the impulsive choice in the current experimental population (n = 14). However, the highest dose significantly increased omissions during the DDT, leading to its exclusion from the impulsive behavior analysis. Changes in omissions during DDT may reflect cognitive processes distinct from impulsivity, such as attention, arousal, or motivation (Cardinal et al., 2000). Preclinical evidence indicates that WIN 55212-2 produces a dose-dependent decrease in time sensitivity, associated with impairments in attentional processes (Crystal et al., 2003). This effect may contribute to the increase in omissions observed after administration of the higher dose of WIN 55212-2 in our experimental setup. In addition, a decrease in locomotor activity may directly affect performance during DDT, thereby increasing the number of omissions (Cardinal et al., 2000). High doses of CB1 agonists have been shown to cause locomotor deficits, including catalepsy, akinesia, immobility, grooming suppression, and exploratory activity inhibition (Rodríguez De Fonseca et al., 1998). Thus, the increase in omissions observed only with a high dose of WIN 55212-2 may be attributed to the hypolocomotor effect and changes in the time sensitivity induced by CB1/2 agonism.
Previous evidence has shown inconsistent results regarding the effects of cannabinoid signaling on impulsive choice. For example, clinical evidence from an online self-report study on the voluntary use of cannabis in patients with ADHD reported beneficial effects, harmful effects, and no effect on ADHD symptomatology (Mitchell et al., 2016). Consistent with our findings in the overall population, Pattij et al. (2007) found that WIN55212-2 did not modify impulsive choice. Interestingly, while acute administration of THC dose-dependently decreases impulsive choice (Wiskerke et al., 2011), the specific activation of CB1 receptors in the OFC has been shown to increase impulsive choice (Fatahi et al., 2018). By contrast, blocking CB1 receptors has been associated with either no effect (Wiskerke et al., 2011) or an increase in impulsive choice (Boomhower and Rasmussen, 2014). This inconsistency has been proposed to arise from individual differences in the baseline levels of impulsive choice (Rinehart and Spencer, 2021). In this sense, rats displaying high baseline levels of impulsive choice show higher expression of CB1 receptors in the OFC (Ucha et al., 2019) and higher DA release in the DLS (Moreno et al., 2021) than rats classified as low impulsive.
A preliminary analysis of the overall population showed that the effect of acute injection of WIN55212-2 on impulsive choice correlated with baseline levels of impulsive choice. While CB1/2 agonism reduces impulsive choice in rats with lower AUClog value, it increases impulsive choice in rats displaying higher AUClog value. A further analysis, following a median split of AUClog values, showed an increase in the AUClog after injection of 1 mg/kg of WIN 55212-2, but only in high-impulsive rats, demonstrating a decrease in impulsive choice after CB1/2 receptor activation. In line with the proposal of Ucha et al. (2019), these results suggest that CB1 agonism could have positive effects on impulsive choice, particularly in vulnerable populations exhibiting high levels of impulsivity.
Considering the neurobiological basis of impulsivity, variations in dopaminergic and cannabinoid signaling have been associated with individual differences in baseline levels of impulsivity. Dopaminergic pathways are strongly linked to the modulation of impulsive choice, showing an inverted U-shaped relationship wherein both excessively high and low dopamine signaling may contribute to impulsive decision-making (Petzold et al., 2019). Accordingly, Moreno et al. (2021) demonstrated that dopamine release in the dorsal striatum correlates with AUClog in the DDT, with low dopamine release associated with low impulsivity and high dopamine release linked to high-impulsive choice (Moreno et al., 2021). In addition, it has been observed that participants with suboptimal dopamine signaling exhibit more impulsive behaviors, which can be attenuated by L-Dopa treatment (Petzold et al., 2019). Conversely, supra-optimal dopamine signaling induced by L-Dopa increases impulsive choice in healthy individuals. This suggests that dopamine-enhancing drugs would be therapeutically effective only for impulsivity choices associated with suboptimal dopamine signaling (Petzold et al., 2019). Considering that CB1 agonism may indirectly regulate dopamine signaling in the mesolimbic and nigrostriatal pathways (Pérez-Valenzuela et al., 2019; Perez-Valenzuela and Evans, 2023), it is possible that WIN increases dopamine release in highly impulsive subjects, normalizes dopaminergic neurotransmission, and reduces impulsive choice. Further experiments with dopamine antagonists are required to test this hypothesis.
In addition, we observed a differential and persistent effect of CB1/2 agonism on impulsivity depending on the baseline levels of impulsivity. In low-impulsive rats, we did not observe differences in the AUClog value between pre- and post-WIN 55212-2 treatment. However, an increase in the AUClog was observed 2 weeks after the last injection of WIN 55212-2 in high-impulsive rats, suggesting a protracted diminishing effect on impulsive choice after repeated exposure to cannabinoids. Despite limited evidence regarding chronic exposure to cannabinoid agonism during adulthood and its effect on impulsive choice, similar enduring changes in impulsivity have been observed using other paradigms. For instance, Miederer et al. (2024) showed that chronic treatment with THC for 21 days increases the correct no-go response associated with decreased rat brain activity in regions such as the hippocampus and amygdala (Miederer et al., 2024). In addition, Marco et al. (2007) showed that chronic enhancement of the endocannabinoid system during adolescence reverses increased impulsive choice induced by maternal separation (Marco et al., 2007). On the other hand, enduring deleterious effects on impulsivity have been observed after more intense exposure to THC. A regimen of nine cycles, each consisting of 5 days of twice-daily THC exposure per day and nine withdrawal days, increased motor impulsivity that persisted for 5 weeks (Irimia et al., 2015). In summary, the long-term consequences of cannabinoids on impulsivity are influenced by baseline impulsivity and intensity of exposure.
To determine the influence of endocannabinoid signaling on the differential, persistent effect of WIN55212-2 on impulsive choice, rats were injected with the CB1 antagonist rimonabant 2 weeks after the last injection of WIN 55212-2. Interestingly, rimonabant reversed the increase in AUClog after repeated treatment with WIN55212-2 in high-impulsive rats. On the other hand, although the repeated treatment with WIN 55212-2 did not result in persistent changes in impulsive behavior, rimonabant decreased the AUClog in the rats classified as low impulsive. These results suggest that the activation of CB1 receptors by endogenous cannabinoids contributes to basal levels of impulsive choice in low-impulsive rats. In line with these findings, Boomhower and Rasmussen (2014) also showed that rimonabant increases impulsive choice and suggested that blocking CB1 receptors induces an overestimation of time, thereby increasing delay aversion. In addition, in rats classified as high impulsive, endogenous cannabinoids appear to contribute to the persistent attenuation of impulsive choice observed after repeated exposure to WIN 55222-2, suggesting an enduring increase in tonic endocannabinoid neurotransmission following repeated exposure to CB1/2 agonists. In this context, chronic prenatal exposure to WIN 55212-2 has been found to induce long-term potentiation of the endocannabinoid system in the striatum of adult rat offspring. This effect is mediated by increased levels of the endogenous CB1 agonist anandamide, which is linked to a reduction in its degradation by fatty acid amide hydrolase (Castelli et al., 2007). Considering that high CB1 receptor expression in the OFC has been observed in rats with high levels of impulsive choice (Ucha et al., 2019), it is plausible to suggest that an increase in anandamide following repeated WIN 55212-2 exposure might lead to a decrease in CB1 receptor expression, thereby reducing delay aversion in highly impulsive rats.
However, there remains a gap in our understanding of the mechanism underlying the long-lasting effect of CB1 agonism on impulsive behavior. For example, cannabinoid agonists have been shown to alter time perception; specifically, clinical studies have reported an overestimation of time, suggesting a subjective slowing of temporal experience (Sewell et al., 2012). Since DDT measures delay aversion, it is plausible to hypothesize that an acute injection of WIN 55212-2 would increase impulsive choice. However, our results show the opposite effect: a decrease in impulsive choice was observed in animals classified as high impulsive. Thus, whether time perception influences impulsive decision-making remains an open question. Furthermore, it is important to note that this study was exclusively conducted in male rats. Given the significant sex differences in impulsivity (Kok et al., 2020) and cannabis consumption (Hinckley et al., 2024), future studies should include female rats. This would provide a more comprehensive understanding of the applicability of these findings to different populations.
In summary, this study demonstrated a differential effect of acute CB1/2 agonism on impulsive choice depending on baseline levels of impulsivity. The acute injection of WIN55212-2 increased the AUClog only in rats classified as high impulsive. These results suggest the potential benefits of CB1/2 agonism in a vulnerable subpopulation and support the possible medical use of cannabis or other CB1 receptor-affine compounds for treating disorders associated with dysfunctional impulsive behavior, such as ADHD or addiction. Such treatments should consider an assessment of choice impulsivity, among other variables, to ensure that the intervention is only delivered to individuals who are likely to benefit from it, thus minimizing potential iatrogenic effects in line with the principles of personalized medicine.
Supplemental Material
sj-jpg-2-jop-10.1177_02698811251355603 – Supplemental material for “Modulatory role of baseline impulsivity on the acute and persistent effects of CB1 agonism on impulsive choice”
Supplemental material, sj-jpg-2-jop-10.1177_02698811251355603 for “Modulatory role of baseline impulsivity on the acute and persistent effects of CB1 agonism on impulsive choice” by Enzo Pérez-Valenzuela, Victor Azocar, Andrea Gräber-Martinez, Alvaro Vergés and José Fuentealba Evans in Journal of Psychopharmacology
Supplemental Material
sj-jpg-3-jop-10.1177_02698811251355603 – Supplemental material for “Modulatory role of baseline impulsivity on the acute and persistent effects of CB1 agonism on impulsive choice”
Supplemental material, sj-jpg-3-jop-10.1177_02698811251355603 for “Modulatory role of baseline impulsivity on the acute and persistent effects of CB1 agonism on impulsive choice” by Enzo Pérez-Valenzuela, Victor Azocar, Andrea Gräber-Martinez, Alvaro Vergés and José Fuentealba Evans in Journal of Psychopharmacology
Supplemental Material
sj-xlsx-1-jop-10.1177_02698811251355603 – Supplemental material for “Modulatory role of baseline impulsivity on the acute and persistent effects of CB1 agonism on impulsive choice”
Supplemental material, sj-xlsx-1-jop-10.1177_02698811251355603 for “Modulatory role of baseline impulsivity on the acute and persistent effects of CB1 agonism on impulsive choice” by Enzo Pérez-Valenzuela, Victor Azocar, Andrea Gräber-Martinez, Alvaro Vergés and José Fuentealba Evans in Journal of Psychopharmacology
Footnotes
Author contributions
Enzo Pérez-Valenzuela: conceptualization, methodology, writing – original draft, data acquisition, formal analysis, and investigation. Victor Azocar: Conceptualization, Methodology, Data acquisition, Software, Data analysis, and investigation. Andrea Gräber-Martinez: Data acquisition and analysis. Alvaro Vergés: Methodology, Supervision, and Data Analysis. Jose Fuentealba Evans: Conceptualization, Methodology, Resources, Supervision, and Funding Acquisition.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by FONDECYT, Fondo Nacional de Desarrollo Científico y Tecnológico, Chile. ID (1141088; JAF).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.