
Abstract
Background:
Although psychedelics have regained attention as potential treatment tools for various mental disorders, little research has examined their impact on temporal perception.
Aims:
This double-blinded placebo-controlled study aimed to investigate changes in temporal perception under psilocybin, both through performance during the Temporal Bisection Task (TBT) and through subjective self-report scales.
Methods:
Twenty-four healthy volunteers were assessed by comparing their performance on two parameters of the TBT —the Bisection Point (BP) and the Just Noticeable Difference (JND) with subjectively reported changes measured using the Hallucinogen Rating Scale (HRS) and the Altered States of Consciousness (ASC) questionnaires.
Results:
We observed a rightward shift in BP under psilocybin compared to placebo (t(23) = 2.27, p = 0.033, g = −0.37). This shift corresponded to reports of subjective time slowing down under psilocybin as measured by HRS and ASC. Psilocybin also increased JND compared to placebo (t(23) = 2.48, p = 0.021, g = −0.47), indicating decreased temporal precision. Consistent with previous findings, these effects were significant for durations longer than 2 seconds.
Conclusions:
Based on Bayesian framework of timing, we emphasised that psilocybin alters time perception through disruptions in cognitive functions, particularly working memory and attention. We also outlined directions for future research, which would allow us to not only understand time perception under psychedelics better, but help elucidate the role of serotonergic system on timing.
Research ID:
The research was conducted as part of a clinical trial registered at EudraCT database under the number 2012-004579-37.
Introduction
Psilocybin is a tryptamine psychedelic present in various species of fungi, most notably the genus Psilocybe (Nichols, 2016). Upon consumption, it undergoes dephosphorylation into the active metabolite psilocin, which is responsible for its psychedelic effects (Tylš et al., 2014). Its primary mode of action is as a serotonin (5-hydroxytryptamine; 5-HT) receptor agonist, binding with high affinity to the 5-HT2A receptor (Vollenweider et al., 1998), as well as the 5-HT1A receptor (Halberstadt and Geyer, 2011) and other 5-HT receptor subtypes with lower affinities. It induces profound and wide-ranging changes in consciousness (Tylš et al., 2014), affecting cognitive, affective and sensory domains, as well as altering time perception (Wittmann et al., 2007).
Alterations in time perception have been reported since the early psychedelic studies in the 1960s (Preller and Vollenweider, 2016) and have also been reflected in key scales of psychedelic phenomenology, such as the Hallucinogen Rating Scale (HRS; Strassman et al., 1994) and the Altered States of Consciousness Scale (ASC; Dittrich, 1998). The ASC includes questions like ‘Time passed slowly in a tormenting way’ or ‘Everything happened so fast that I could not follow it all’, highlighting how temporal perception can vary widely, sometimes slowing down, sometimes speeding up or occasionally giving rise to a sense of timelessness as reflected in the question ‘I experienced past, present, and future as a oneness’. Although there are similarities in their effects, different psychedelics can influence time perception in unpredictable ways, with 5-methoxy-N,N-dimethyltryptamine (5-MeO-DMT) more frequently leading to mystical-type experiences characterised by a sense of timelessness (Davis et al., 2018). They also differ in duration of effects, ranging from the several-minutes-long but extremely intense experience of vaporised 5-MeO-DMT (Reckweg et al., 2022) to the hours-long experience after lysergic acid diethylamide (LSD) administration (Holze et al., 2024).
Temporal processing is a complex system governed by distinct cognitive and neurobiological mechanisms (Buhusi and Cordes, 2011) operating across multiple scales – from circadian rhythms to mechanisms responsible for timing in the millisecond to second range (Buonomano, 2007); this study specifically addresses time perception within the seconds range, commonly termed ‘interval timing’.
Scalar Expectancy Theory (SET; Gibbon, 1977) is a classical model of time perception proposing an internal clock mechanism in which a pacemaker emits pulses that are accumulated in working memory. A switch, modulated by attentional processes, regulates the transmission of pulses. The accumulated pulses are compared against a reference interval stored in memory, corresponding to subjective duration estimation. Building upon this conceptual foundation, the Striatal Beat Frequency (SBF) model offers a neurophysiological account of interval timing (Oprisan and Buhusi, 2011). The SBF model posits the existence of cortical oscillators operating at various frequencies located most likely in the prefrontal cortex. At the onset of to-be-timed interval, cortical oscillators are phase-reset, and at the offset of the interval, the state of the oscillators forms a unique activation pattern, which is detected by neurons located in the striatum (coincidence detectors) allowing the estimation of elapsed time.
Alternative contemporary models utilise a Bayesian framework, offering a distinct theoretical approach (Sadibolova and Terhune, 2022). Bayesian models conceptualise time perception as the integration of prior temporal knowledge with incoming sensory data via Bayesian inference. The resulting perceived interval (posterior distribution) reflects a weighted combination of prior temporal memory and sensory evidence, modulated by a cost function that optimises expected gains and minimises losses associated with timing decisions.
Temporal processing is modulated by various neurotransmitter systems, for example, dopaminergic releasers and reuptake inhibitors accelerate the subjective flow of time, whereas D2 receptor antagonists slow it down (Lake and Meck, 2013). The role of the serotonergic system (which is psilocybin’s primary target (Meck, 1996)), and thus the effects of psychedelics, remains less clear. Asgari et al. (2006) demonstrated that the 5HT2A receptor mediates the time-dilating effects of 2,5-dimethoxy-4-iodoamphetamine in rats, as these effects were completely reversed by administration of highly selective 5HT2A receptor antagonist volinanserin. The first modern study examining the effects of psilocybin (115 and 250 μg/kg) on time perception in humans found that participants under-reproduced temporal intervals longer than 3 seconds (Wittmann et al., 2007), had difficulty synchronising motor response (finger tapping) to intervals over 2 seconds and showed an increase in their preferred tapping rate from 692 to 949 ms. A similar study using a lower dose of psilocybin (12 μg/kg) yielded comparable results (Wackermann et al., 2008). Yanakieva et al. (2019) investigated the effects of LSD microdose on temporal reproduction of older adults with timing deficits, finding that participants under LSD and placebo both under-reproduced intervals longer than 2 seconds. However, this under-reproduction was less prominent in the LSD group, leading researchers to speculate that the LSD microdose may have corrected their timing deficits to some degree. These results are in contrast to those of Wittmann et al. (2007), who have observed less accurate time perception after psilocybin administration compared to placebo. However, a population with timing deficits being used in the LSD study and the fact that a microdose was used could explain why the results are different from the other studies on psychedelics and time perception.
As previous human studies testing time perception under psilocybin used only temporal reproduction and production tasks, our study was designed to examine whether time perception is also affected during temporal categorisation, specifically in a temporal bisection task (Penney and Cheng, 2018). Temporal bisection presents a different set of cognitive and motor demands compared to temporal reproduction. It requires a categorical decision to be made about a duration by comparing it to two other durations (Cantarella et al., 2023); therefore, three durations have to be remembered and compared. Contrastingly, during temporal reproduction, only one target duration has to be remembered and then reproduced. On the other hand, temporal reproduction requires planning and then executing motor action that is supposed to reflect the interval being reproduced (Mioni et al., 2014). By using temporal bisection, the influence of possible psilocybin-induced motor alterations should be minimised. Moreover, we presented stimuli during the task visually, as opposed to using auditory stimuli used in previous studies with psilocybin. Sensory modality has been shown to influence temporal perception, with visual stimuli being judged as shorter, compared to auditory stimuli of the same duration (Bratzke, 2025). Using visual stimuli will support broader generalisations regarding whether temporal distortion following psilocybin administration is a general phenomenon, independent of sensory modality. Furthermore, we used responses on the ASC and HRS scales regarding subjective experience of time and tested whether the subjectively reported time distortion correlated with the measures of time perception obtained from bisection task. We hypothesised that the speeding up or slowing down of subjective time reported on the ASC and HRS questionnaires would be reflected in a corresponding shift of bisection point (BP, measure of timing accuracy extracted from the results of bisection task). For more information about the indices obtained from the bisection task, see methods section.
To test this hypothesis, we have administered both psilocybin and placebo to participants and asked them to complete the temporal bisection task 260 minutes after administration. We then compared their performance on psilocybin relative to placebo by comparing the Just Noticeable Difference (JND) and BP indices of time perception.
Methods
Study design
The study received approval from the ethics committee of the National Institute of Mental Health (former Prague Psychiatric Centre) and the Czech State Institute for Drug Control. These experiments are part of a larger clinical trial registered at the European Union Drug Regulating Authorities Clinical Trials (EudraCT) database under the identifier 2012-004579-37. The study was not pre-registered, and all of the analyses in this article are exploratory.
The full trial consisted of multiple experiments; further details can be found in Klučková et al. (2024) and Viktorin et al. (2022). The study employed a double-blinded, placebo-controlled design, with participants receiving both placebo and psilocybin. Randomisation was performed using the block permutation method. We have not controlled for unblinding. Each session was conducted by a research team consisting of a clinician (psychiatrist), a second researcher and a nurse or technician. In every research team, there was at least one male and one female member. The research team assigned to each particular participant, as well as the intake procedures have stayed consistent across both sessions. We tried to ensure that all the factors in our control were identical in both conditions (room, duration of study etc.). The study design adhered to the Guidelines for Safety in Human Hallucinogen Research (Johnson et al., 2008). The minimum and maximum times between the two dosing sessions were 28 and 147 days respectively, with the mean time of 66.25 days (SD = 37.96). However, having three participants with over 130 days between sessions skews the mean upwards, with the majority of participants having shorter duration between sessions, reflected in the median time of 50 days. The sessions for female participants were scheduled outside of their menstrual phase, as previous research has shown altered responses to psilocin in female mice during their metestrus and diestrus periods (Tylš et al., 2016).
Participants
A total of 25 healthy participants (15 males/10 females) were recruited through the snowball sampling method. Participants ranged in age from 28 to 55 years, with a mean age of 38.2 years (SD = 7.91 years). One female participant was excluded from the subsequent analysis due to a corrupted dataset under psilocybin condition (where all responses were recorded as 6000 ms, something that should not have been possible, due to 1000 or 3000 ms being the only two possible responses, corresponding to the ‘short’ and ‘long’ durations). An a priori power analysis for sample size was not performed for the outcomes of the timing experiment, as the data were collected as part of a larger study involving functional magnetic resonance imaging (fMRI), for which the sample size was more than adequate (Szucs and Ioannidis, 2020).
The inclusion criteria consisted of being aged between 28 and 65 years old, having no psychiatric history (no mental illnesses included in ICD 10 F0.X–F99.X) and having no family history of psychotic disorders up to second-degree relatives. Exclusion criteria consisted of pregnancy, hypertension, stroke, congestive heart failure, having a pacemaker, metal clamp in the head and/or face, celiac disease, left-handedness, usage of any medication on a daily basis (except for contraception) and being a student of medical faculty. All participants underwent pre-screening to ensure that they fulfilled all inclusion criteria including assessment with the Mini-International Neuropsychiatric Interview v. 5.0 (Sheehan et al., 1998), conducted by a study clinician. They were additionally screened for clinically significant psychopathologies using Minnesota Multiphasic Personality Inventory-2 (Butcher et al., 1989). Eligible participants were informed about the study design and provided written informed consent.
To minimise potential confounding factors, participants were instructed to abstain from alcohol and other psychoactive substances for at least 1 week before the experiment and between sessions. On measurement day, participants arrived in a fasted state, refrained from consuming coffee, and smokers were instructed to refrain from smoking for at least 2 hours before the start of the experiment. Smoking was prohibited throughout the intoxication phase of the session.
Procedure
On the dosing days, psilocybin was administered, and the session lasted for 6–8 hours, most of which was spent in a comfortable, decorated room. The temporal bisection task was performed 260 minutes after drug/placebo administration, during the descending part of intoxication, when major perceptual and cognitive disturbances – potentially rendering the task inoperable – were expected to subside (Hasler et al., 2004). We believe that completing the task 260 minutes after administration offered a good compromise between ensuring some effects of psilocybin were still present, but they were not too strong to make the task inoperable, as the duration of psilocybin experience lasts from 240 to 360 minutes (MacCallum et al., 2022).
To account for the possibility that psilocybin-induced visual perceptual effects could influence performance on the temporal bisection task (as the durations were being represented visually), a simple hallucination task was included. The task involved presenting a series of either static or morphing stimuli (faces and ellipsoids) on a computer screen for 2 seconds. Participants were then prompted to indicate whether the stimulus presented was static or morphing and whether it depicted a face or an ellipsoid. For more information about the visual task, see Vejmola et al. (2025). Even though a moderate significant effect persisted just before the start of the temporal bisection task, it was primarily driven by altered perception of complex images of morphing faces but not ellipsoids. This confirmed that subjects regained sufficient visual perception of simple objects at this point after psilocybin ingestion, and no participants were excluded from completing the temporal bisection task. Blood samples were collected to measure psilocin levels over the course of the session. Psychometric scales assessing psychopathology and the phenomenology and intensity of the experience were also administered (see section Psychometrics below). As the study did not focus on therapeutic outcomes and only healthy volunteers were sampled, we did not offer any therapy before or after sessions. We did, however, offer psychological support to participants who have found their experiences challenging.
Drug and dosage
The psilocybin used in this study was produced in compliance with Good Manufacturing Practice standards at THC-Pharm GmbH in Frankfurt, Germany. Gelatine capsules containing either 1 mg or 5mg psilocybin homogenised with tritici amylum or placebo capsules containing only tritici amylum were produced at the pharmacy of the Institute for Clinical and Experimental Medicine in Prague, Czech Republic. The target dose of 0.26 mg of psilocybin per kilogram of body weight for each participant was achieved by combining 1- and 5-mg capsules to best fit each participant’s weight. Participants weighed from 61 to 92 kg, which resulted in doses ranging from 16 to 24 mg, with a mean dose of 19 mg (SD = 2.47 mg). During placebo trials, participants received the same number of capsules as in their psilocybin trial to maintain blinding. All capsules were ingested with 200 ml of water.
Psychometrics
The Czech translation of the second version of Brief Psychiatric Rating Scale (BPRS; Overall and Gorham, 1988) was administered by the study sitters to track the participants’ psychological states. For the present analyses, only the measurements taken 240 minutes after drug administration were used, as this time point was the closest to the temporal bisection task. For an overview of the timeline of the experiment see Figure 1. below.
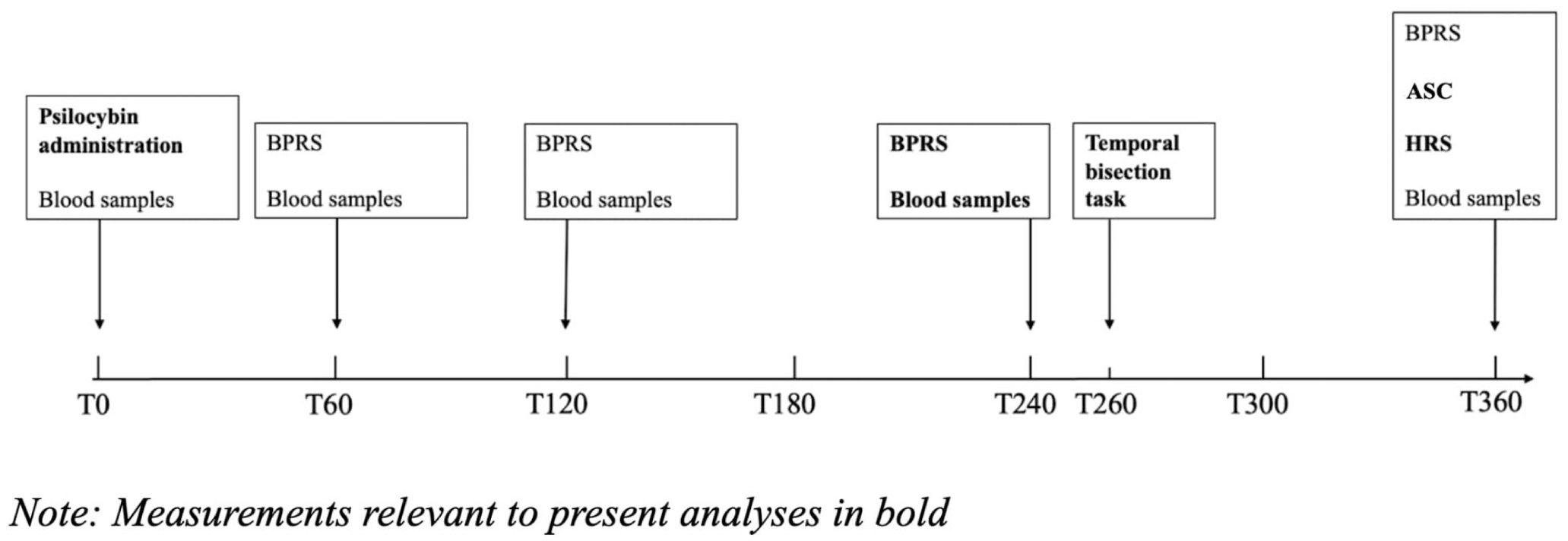
Timeline of the experiment.
The Czech version of the English ASC questionnaire, translated from the original Außergewöhnliche Psychische Zustände by Dittrich (1998), was administered at the end of the session, 360 minutes post psilocybin administration. The questionnaire consists of 72 questions divided into three subscales: Dread of Ego Dissolution (AIA), Oceanic Boundlessness (OBS) and Visionary Restructuralisation (VUS). It also provides a composite score VWB (from the German phrase Veränderter Wachbewusstseinszustand, meaning Altered State of Consciousness), representing the overall degree of consciousness alteration based on the three subscales. VUS captures perceptual alterations, AIA assesses negative experiences related to ego dissolution, while OBS evaluates positive experiences associated with ego dissolution.
In addition to the ASC scale, the HRS (Strassman et al., 1994) was also administered at the end of the session, to retrospectively evaluate participants’ subjective experiences.
Specific questions related to time perception were extracted from both the ASC and HRS scales and analysed in relation to the primary parameters of the temporal bisection task. For the full list of extracted questions, refer to Table 1. below.
Variables and abbreviations.

Temporal bisection task
The task was a computer-based visual test displayed on a standardised screen, with participants seated 70 cm away. Before the intoxication, participants were taught how to complete the temporal bisection task. Afterwards, they were asked to explain what they believed was expected of them, in order to ensure that they understood the instructions correctly. Participants were instructed not to count the time passed in their heads during the task; however, we did not test whether they followed these instructions. The task included training and test phases. During the training phase, participants were presented with a fixation cross for 2000 ms, followed by 200-ms delay. A blue circle, 45 mm in diameter, then appeared at the centre of the screen for either 1000 or 3000 ms, which were chosen as the short and long anchor durations respectively. After the stimulus, participants were prompted to indicate whether they were presented with a short or long stimulus using a left or right mouse click, respectively. Feedback on the correctness of their response was provided after each trial. The test phase consisted of four sequentially alternating trials of short and long stimuli. For a visual representation of the training phase design, see Figure 2.
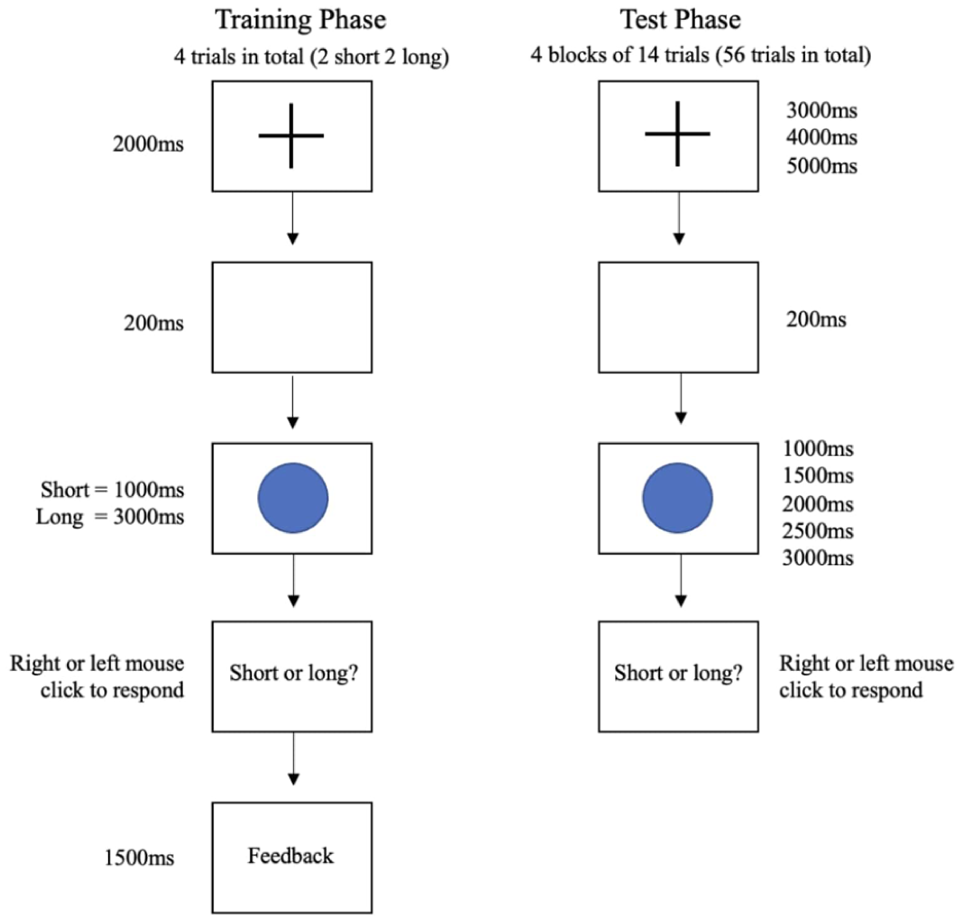
Design of the training and test phases of the experiment.
After participants became familiar with operating the task during the training phase, they proceeded to the test phase. The test phase began with the presentation of a fixation cross for a randomly selected duration (3000, 4000 or 5000 ms). Following a 200-ms delay, the same stimulus as in the training phase—a blue circle—was displayed for one of five possible durations (1000, 1500, 2000, 2500 or 3000 ms), with the anchor durations of 1000 and 3000 ms being presented 16 times each and 1500, 2000 and 2500 ms being presented 8 times each. We acknowledge that the number of trials per interval was lower than ideal. The total of 56 trials was determined based on pilot testing and constraints around participant attention span and task duration. While we aimed to balance duration coverage with practical limits, we recognise that having fewer than 20 trials per interval may impact statistical power, and this is addressed in our discussion of limitations. Participants were then prompted to classify the stimulus as either short or long by clicking the left or right mouse button, respectively. Unlike the training phase, no feedback was provided during the test phase. Each participant completed a total of 56 trials, organised into four blocks of 14 trials each. For a visual representation of the test phase design, refer to Figure 2.
By analysing the proportion of ‘long’ responses for each stimulus duration, we can determine whether – and if so, how – temporal discrimination is affected (Carvalho et al., 2019). Under unaltered and accurate temporal perception, we would expect to see the highest proportion of ‘short’ responses for 1000 ms, followed by 1500 ms stimuli and the highest proportion of ‘long’ responses for 3000 ms, followed by 2500 ms stimuli. For the 2000 ms stimulus, a 50% probability of selecting ‘long’ would be expected, as its duration lies exactly midway between ‘short’ and ‘long’. A shift in the probability of selecting ‘long’ indicates change in subjective time perception: an increased proportion of ‘long’ responses suggests perceived time shortening, whereas a decreased proportion suggests perceived time lengthening.
This is reflected in the two indices we can obtain from the responses to temporal bisection task, BP and JND. BP refers to the interval which a participant is 50% likely to classify as ‘long’. In the case of fully accurate time perception, the BP for our study should lie at 2000 ms, as this is the point that is exactly between ‘short’ and ‘long’ durations (1000 and 3000 ms). By comparing the observed BP of each participant to this ‘ideal’ BP of 2000 ms, we can infer changes in their time perception. For example, BP of 2100 ms would be indicative of subjective time ‘slowing down’, compared to the actual time elapsed, as the participant indicated experiencing 2100 ms, when only 2000 ms have passed, therefore subjectively experiencing extra 100 ms (analogous to subjectively feeling like 70 minutes have passed when in reality 60 minutes have passed). Conversely, BP of 1900 ms would be indicative of subjective time ‘speeding up’, compared to the 2000 ms actually passed (analogous to subjectively feeling 50 minutes have passed when 60 minutes have passed in reality). JND is a measure of the shortest change in duration that the participants can reliably detect.
Blood serum psilocin levels
Blood samples were collected at multiple time points throughout the experiment. The samples were centrifuged at room temperature at 4000 rpm for 10 minutes. The sera were then stored at −80°C until analysed for psilocin content using liquid chromatography–mass spectrometry.
Data processing and statistical analyses
Bisection point and just noticeable difference
The main indices of interest are the BP and JND. In psychophysics, the BP is the stimulus value at which a participant perceives two different stimuli as being equal or at which a variable stimulus is judged by an observer to be equal to a standard stimulus. For the temporal bisection paradigm, the BP is the midpoint at which a participant perceives a test stimulus as equally similar to the two reference stimuli (e.g. short and long durations in a bisection task). It typically represents a point of perceptual bias, and it reflects how closely participants’ responses or perceived values align with the true or objective value of a stimulus, indicating the accuracy of their perception.
JND represents the smallest change in stimulus duration that can be reliably detected. It is typically calculated from the steepness of the psychometric curve, which represents the range over which stimuli are perceived as similar. Here, the JND indicates an observer’s sensitivity to temporal differences – smaller JND suggests better temporal discrimination abilities. It is considered as an indicator of precision, specifically in terms of the variability of responses (van Wassenhove et al., 2008).
JND and BP were calculated based on participants’ responses to stimuli presented at various intervals. To model the relationship between the proportion of long responses and stimulus intervals, a generalised linear model (GLM) was fitted. Specifically, the GLM employed a binomial distribution with a logit link function, chosen for its suitability in modelling binary response data (Vidotto et al., 2019).
The coefficients obtained from the GLM were manipulated to derive the BP and JND using the following formulas:


where a represents the intercept, and b indicates the coefficient for a time interval.
We assessed model fit for the psychometric functions using several standard indices from the logistic regression models (GLMs) fit to each participant’s data. Specifically, for each individual, we modelled the proportion of ‘long’ responses as a function of stimulus interval using a binomial logistic regression. Both intercept and slope terms were statistically significant (p < 0.001), confirming that the psychometric function was well captured by the model.
Statistical analysis
Statistical analyses were done using R version 4.3.2. (Rstudio Team, 2023). We fitted two linear mixed models using the lmer function from the lme4 package (Bates et al., 2015) to separately predict the effects of the predictors on JND and BP. We aimed to quantify the effects of psilocybin on time perception while controlling for the confounding variables. Moreover, we were interested in the relationship between the self-reported changes in time-perception extracted from the ASC and HRS questionnaires and the time-perception indices obtained from the temporal bisection task. Thus, the initial model consisted of all predictors of interest which were iteratively removed based on the significance levels. The best-fitting model was determined through a stepwise elimination process, comparing more complex models to simpler ones based on the Bayes information criterion (BIC) values. Model comparisons were carried out using custom scripts.
Handling missing data
Two participants were missing their psilocin levels at 240 minutes post administration and one participant was missing their response to ASC_Q3. All three instances of missing data were addressed using a multiple imputation algorithm. Having a complete dataset is especially desirable when comparing nested linear mixed models. The imputation algorithm treats each missing value as a dependent variable and predicts it based on the other variables in the data set. A total of 5000 datasets were imputed, pooled together, and the values with the highest certainty were selected. The procedure was done using the library ‘mice’ in R (van Buuren and Groothuis-Oudshoorn, 2011).
Data availability
Raw data can be obtained by request from the corresponding authors.
Results
We plotted the mean responses of participants for each duration of the bisection task on separate graphs and fitted the results with a psychometric curve. Figure 3 shows responses of all participants in the placebo condition, and Figure 4 shows responses of all participants in the psilocybin condition.
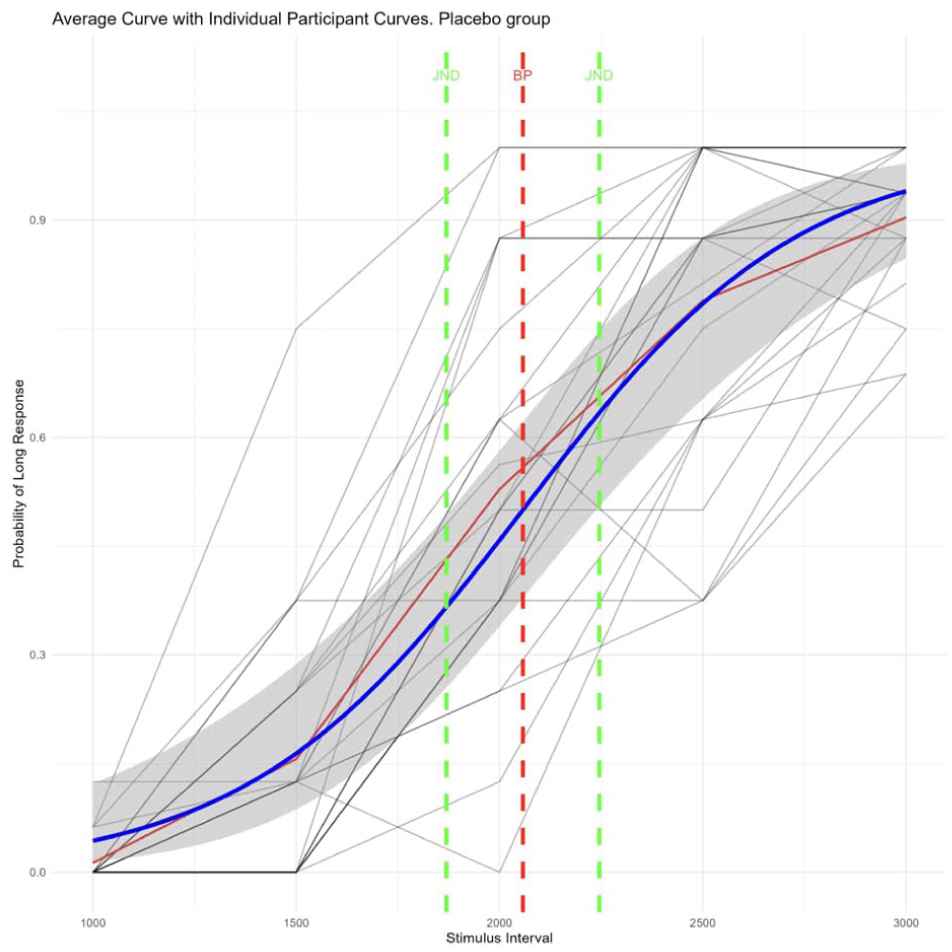
Individual placebo participant responses and average placebo psychometric curve.
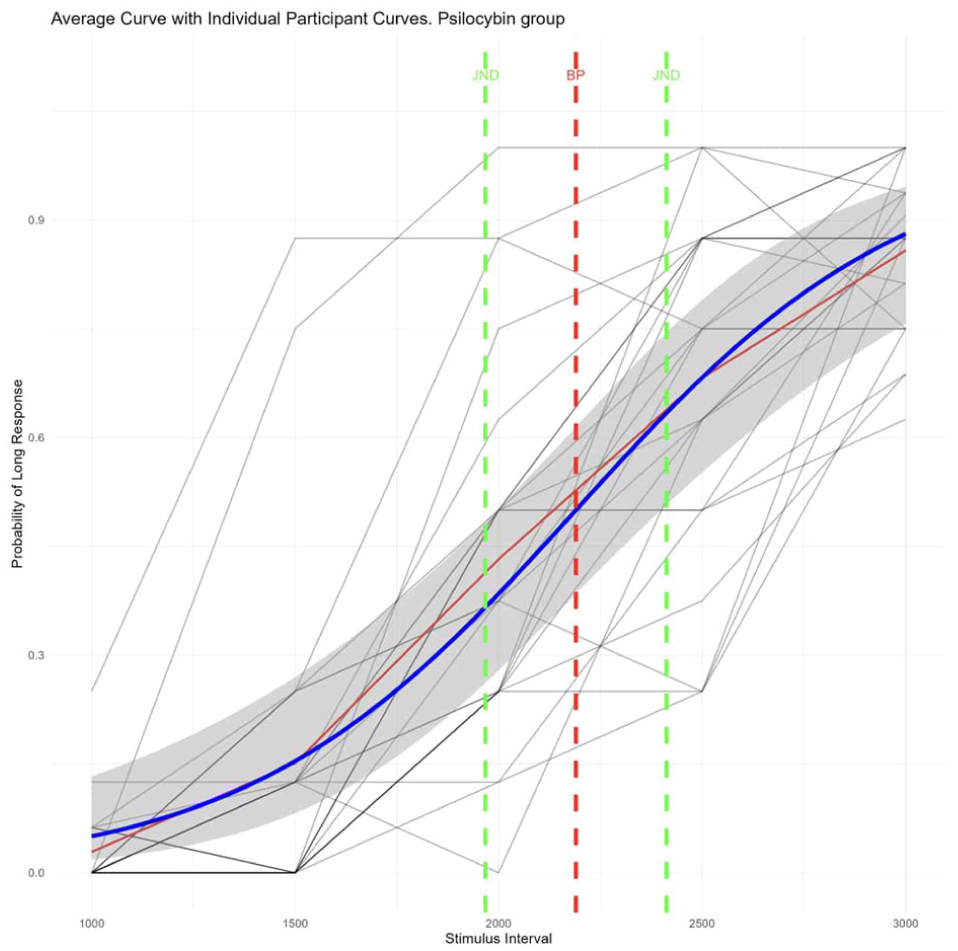
Individual psilocybin participant responses and average psilocybin psychometric curve.
Difference between JND and BP across groups
Initially, we used a paired t-test to compare the average JND and BP under placebo and psilocybin. Both JND and BP were significantly higher under psilocybin compared to placebo when not controlling for any confounding variables (t(23) = 2.48, p = 0.021 and t(23) = 2.27, p = 0.033, respectively). These results reflect a shallower slope (indicating higher JND) and a rightward shift of BP. However, the effect sizes for the t-tests of both JND (g = −0.47) and BP (g = −0.37) were relatively small. For visual representation of the difference between the conditions, see Figure 5.
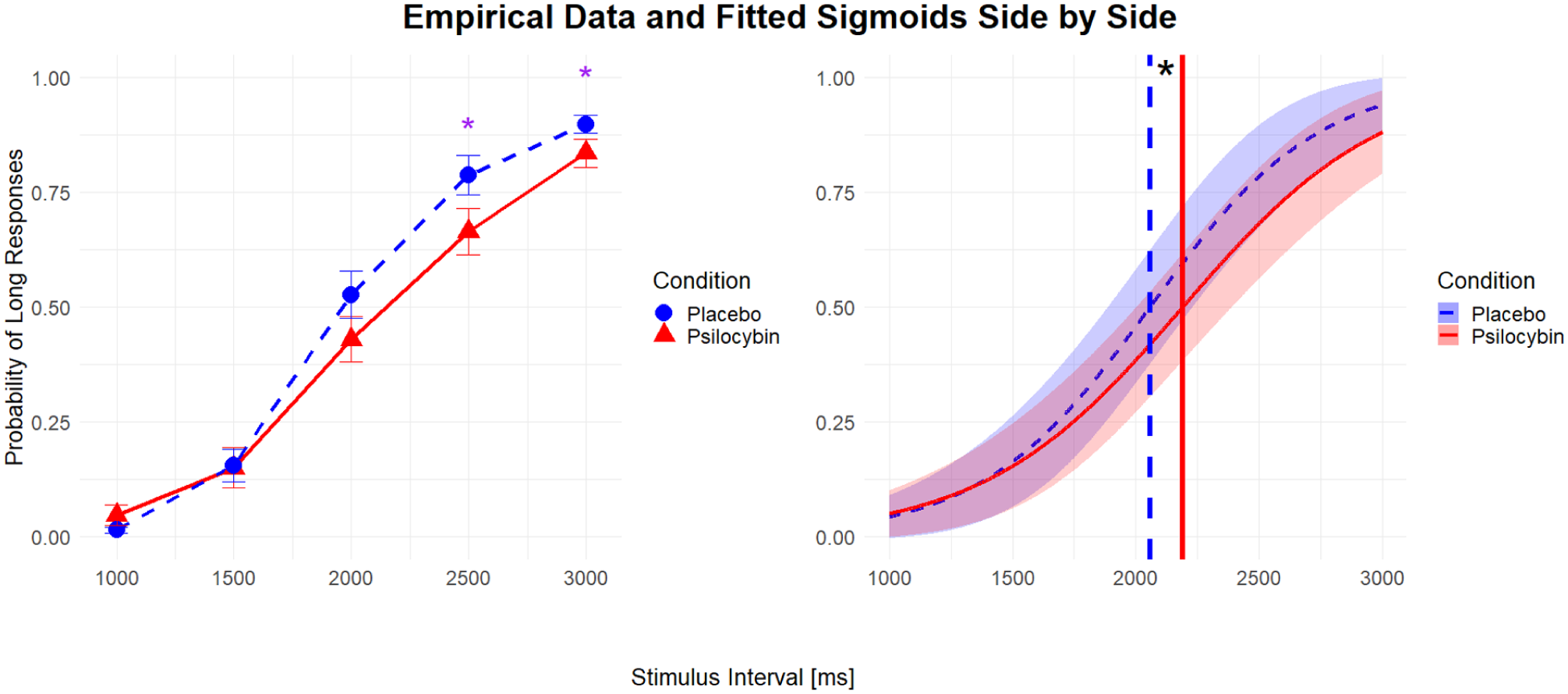
Average fitted response curves from the GLM model across intervals and average observed response proportions across intervals.
These calculations confirmed that time perception was altered under psilocybin. The next step was to identify the source or sources of the time perception distortion and account for the confounding predictors.
Both initial models included predictors:
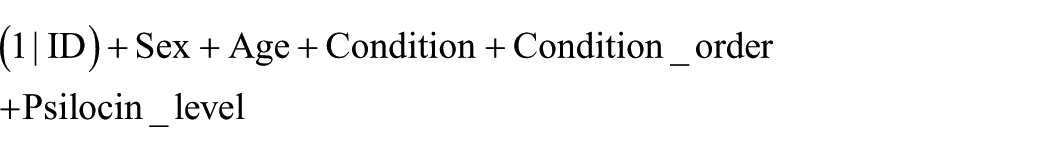
where (1|ID) represents the random intercept.
JND model
We used an iterative model refinement approach based on stepwise elimination to identify the best-fitting model. Starting with all predictors of interest, we removed the least significant predictor at each step and refitted the model. If the simpler model showed a better fit (as assessed by a lower BIC), we proceeded to the next step. This process continued until removing additional predictors no longer improved model fit, or only the main predictor (‘Condition’) remained.
In the final model, the psilocybin condition (ConditionPSI) emerged as the only significant predictor of JND (t(23) = 2.48, p = 0.021; see Table 2).
Summary of the best model predicting JND.
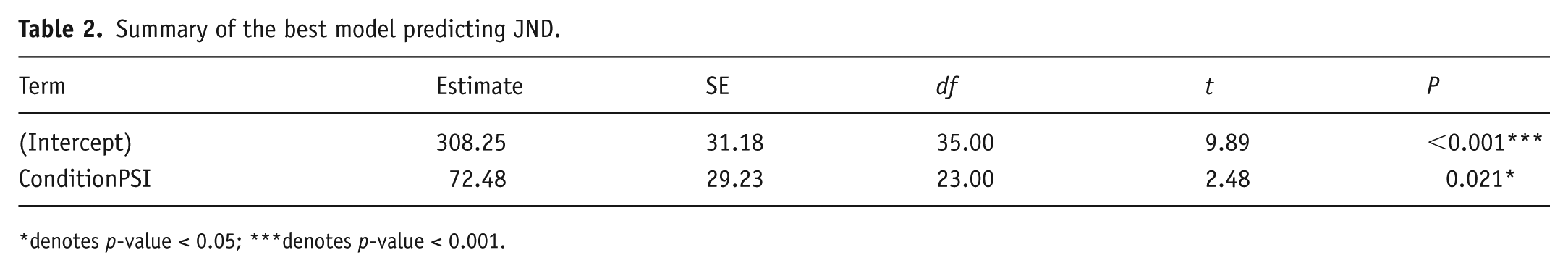
denotes p-value < 0.05; ***denotes p-value < 0.001.
BP model
The BP model was analysed in the same way as the JND model. Following the same process as in the JND model, the final best-fitting BP model retained ConditionPSI as the only surviving predictor (t = 2.27; p = 0.033; see Table 3).
Summary of the best model predicting BP.
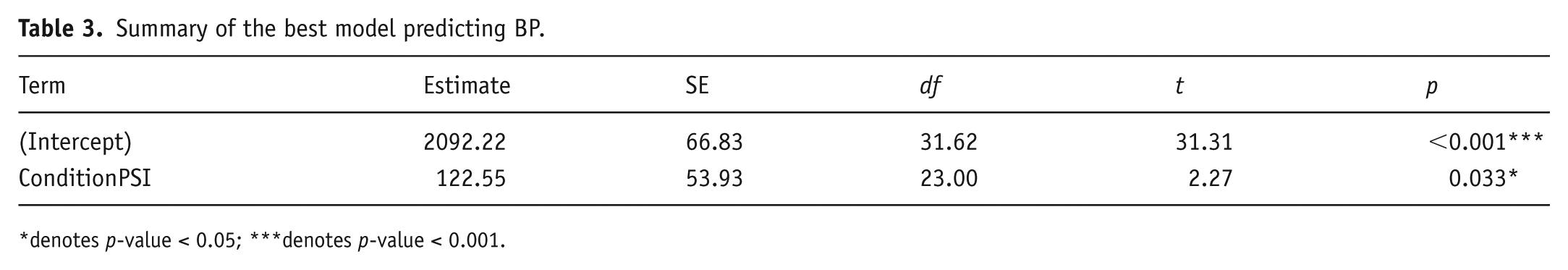
denotes p-value < 0.05; ***denotes p-value < 0.001.
Bootstrapping
Posterior checks revealed that both JND and BP models violated some of the assumptions. To assess the stability and reliability of parameter estimates, bootstrapping was employed using a residual method with 5000 resamples for both the BP and JND models. The 95% confidence interval (95% CI) for the parameter estimate ConditionPSI in the bootstrapped BP model (92.15, 169.65) indicates that the estimate in the original model is reliable. However, 95% CI for ConditionPSI in the bootstrapped JND model includes zero (−2.01, 96.29), suggesting that the original JND model’s estimate for ConditionPSI is not robust and could be statistically insignificant.
Correlation matrix
As models for both JND and BP had condition as the only surviving predictor, meaning none of the other factors improved the model fit, we have decided to run an exploratory correlational analysis between all the factors. We have used Spearman’s correlation coefficient, due to it being a non-parametric method, appropriate for non-normally distributed data. We have corrected for multiple comparisons using the Holm method. The correlation matrix can be seen in Figure 6.
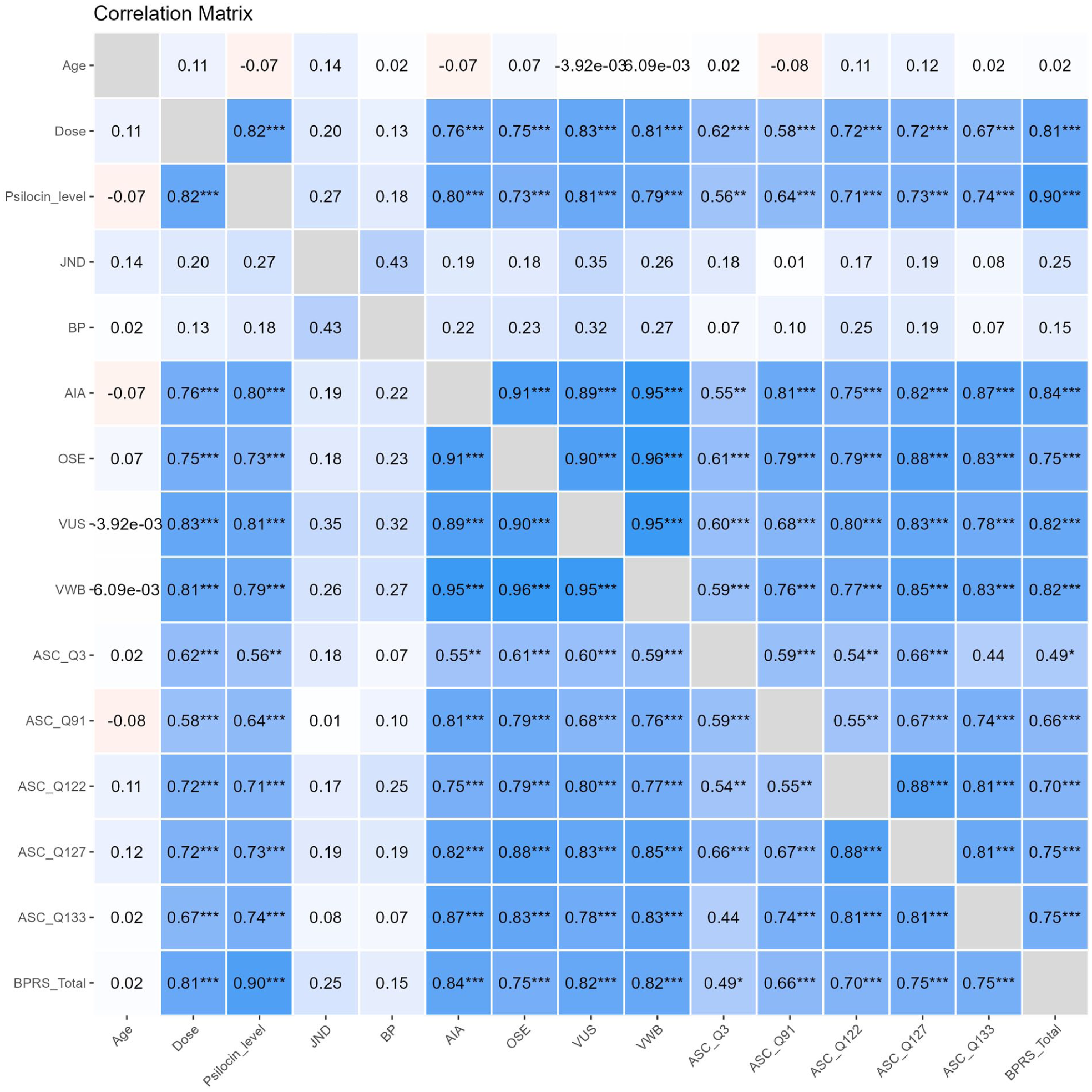
Correlation matrix.
Analysis of subjective perception of time
We used factor analysis to narrow down which questions from the administered questionnaires might be best associated with the changes in time perception. We used items from the ASC questionaire (ASC_Q3, ASC_Q133 and ASC_Q91), as they were specifically prompting participants to report a change of subjective time perception. To see their exact wording, refer to Table 1. We have conducted a factor analysis with Varimax rotation. In a single-factor solution, the variable ASC_Q133 had the highest loading (0.997) and ASC_Q3 had the second highest loading (−0.477). This inverse correlation suggests that the underlying factor relates to ‘the speed of time passing’ as ASC_Q133 asked participants to what extent ‘Time was passing much slower than usual’, as ASC_Q3 asked to what extent ‘Time was passing quicker than usual’. The participants’ responses to ASC_Q133 were significantly higher during the psilocybin compared to the placebo condition (t(23) = 5.72, p < 0.001). Visual representation of this can be seen in Figure 7.
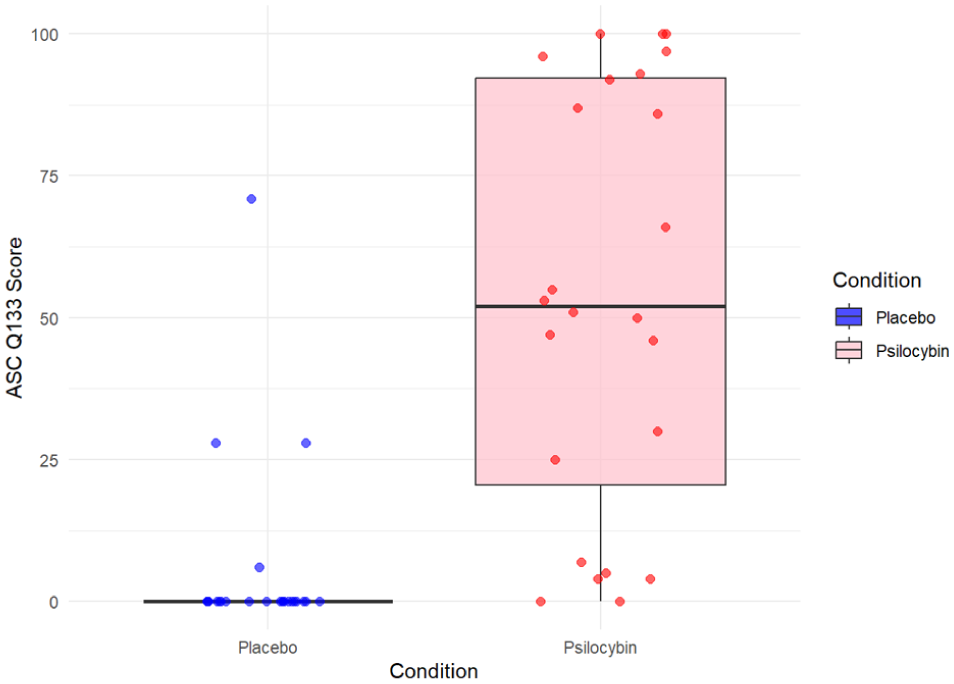
Comparison of responses to ASC_Q133 between placebo and psilocybin conditions.
Furthermore, we examined whether self-reported changes in subjective time perception (ASC_Q133) predicted changes in the parameters measured during bisection task (JND or BP). However, ASC_Q133 did not significantly predict changes in either BP (z = 1.39; Est. = 1.19; p = 0.163) or JND (z = −0.126; Est. = −0.07; p = 0.889).
Analysis of the psilocybin condition
Next, we analysed the psilocybin condition separately, given the significant difference between the groups in their ASC_Q133 responses. The correlation between ASC_Q133 and BP was t(22) = 0.21, r = 0.11, p = 0.838 and the correlation between ASC_Q133 and JND was t(22) = 0.50, r = 0.04, p = 0.620. The correlation between JND and BP was t(22) = 1.10, r = 0.23, p = 0.285.
We were then interested in whether the perceived intensity of time alteration, as measured by HRS_Q86a, could predict either of the parameters JND or BP for the psilocybin group. While HRS_Q86a did not significantly predict JND (t(17) = −1.15; Est. = −20.25; p = 0.265), it did significantly predict BP (t(14) = 2.41, Est. = 74.47; p = 0.030) along with age (t(14) = 2.52; Est. = 17.35; p = 0.024). All p-values in regard to correlation analyses were corrected for multiple comparisons using the Holm method.
Discussion
The main findings of this study are the significant differences between the psilocybin and placebo conditions in both parameters of temporal bisection task, namely BP and JND. Participants under the influence of psilocybin demonstrated a rightward shift in BP, reflecting an underestimation of intervals during the temporal bisection task. Furthermore, JND values were larger under psilocybin, suggesting greater variability in responses and reduced precision in temporal judgement. Moreover, we have also found a significant difference in self-reported changes in subjective time perception between the two groups (as measured by ASC_Q133), with participants in the psilocybin condition reporting more subjective slowing down of time compared to placebo. The self-reported intensity of changes in temporal perception (as measured by HRS_Q86a) significantly predicted the shift in BP for the psilocybin condition. The responses on ASC_Q133 correspond well with the rightward BP shift observed for psilocybin condition, which can be interpreted as being indicative of slowing down of internal time, when interpreting the result via the SET framework.
Our findings partly align with the previous studies on psilocybin’s effects on time perception (Wittmann et al., 2007; Wackermann et al., 2008), with the time intervals at which we observed an effect of psilocybin roughly corresponding to those reported in previous studies. Wittmann et al. (2007) found deficits in interval reproduction for durations exceeding 3 seconds and in motor synchronisation for durations longer than 2 seconds. In our study, group differences emerged from 2 seconds onwards, with the 2500- and 3000-ms intervals reaching statistical significance. These findings support the notion of a threshold around 2–3 seconds (depending on the task used), below which psilocybin does not appear to affect performance on timing tasks. The previous studies on psilocybin and time perception also reported a deceleration of preferred tapping rate under psilocybin. The speed of preferred tapping rate has been suggested to be directly linked to the speed of internal clock (Vanneste et al., 2001), with faster tapping rate corresponding to faster subjective flow of time and slower tapping rate corresponding to subjective time dilation. Therefore, interpreting the rightward shift in BP we observed in the psilocybin group as a slowing down of the internal clock pacemaker is consistent with the previous findings on preferred finger tapping rate and psilocybin. However, due to the motor aspect of the tapping task, the slowing down of preferred tapping rate could have been caused by motor disturbances, which were reported after psilocybin administration by some studies (Velit-Salazar et al., 2024), rather than reflecting the slowing down of internal clock.
However, during temporal reproduction, Wittmann et al. (2007) have reported under-reproduction of intervals after psilocybin. From the perspective of the SET framework, such under-reproduction would imply increased speed of the internal clock. This interpretation contrasts not only with our results, but also with their own findings of decreased finger tapping speed. Reconciling these observations within the pacemaker–accumulator model would require assuming that psilocybin does not affect the pacemaker itself, but rather the reference memory component of the SET model, which would be consistent with the under-reproduction findings and not in contradiction with the rightwards shift in BP. Alternatively, the different timing tasks could impose distinct cognitive, attentional and motor demands, leading to task-specific effects of psilocybin.
However, viewing the results through the Bayesian framework allows us to better reconcile these seemingly contradictory findings. The Relaxed Beliefs Under Psychedelics (REBUS) model (Carhart-Harris and Friston, 2019) proposes that psychedelics decrease the influence of priors, thus increasing the relative weight of bottom-up sensory information. Sadibolova et al. (2023) have re-analysed the data on LSD microdose improving accuracy of temporal reproduction compared to placebo in adults with timing deficits (Yanakieva et al., 2019). Their results are consistent with a reduced influence of priors, which were assumed to underlie temporal biases. However, applying the REBUS framework to our, as well as previous psilocybin results, does not result in satisfactory conclusions. If the influence of top-down priors (leading to biases) decreased, we would expect timing to be more accurate under psilocybin compared to placebo, which was not the case in any of the psilocybin studies. However, the interactions between top-down and bottom-up streams of information are complex, as they are hierarchical, with multiple levels of inferences and feedback loops (Leptourgos et al., 2020). Furthermore, the participants in the Yanakieva et al. (2019) study have only been administered a micro-dose of LSD and, therefore, were expected to experience barely perceptible effects, with no hallucinations or strong perceptual and cognitive alterations. This is in contrast to our and Wittmann et al. (2007) studies, where the psilocybin doses administered were high enough to elicit strong changes in perception and cognition. Contrastingly to REBUS, other researchers have proposed that psychedelic-induced hallucinations (Corlett et al., 2009) as well as hallucinations of non-pharmacological origin (Corlett et al., 2019) are caused by increased top-down processing and stronger priors, that impose a (‘false’) structure on sensory information. Sterzer et al. (2018) have tried to reconcile these opposing views by proposing that different types of hallucinations may be caused by different disruptions to Bayesian interference, with sensory hallucinations being marked by stronger priors, whereas hallucinations related to self-generated phenomena may be the result of weaker priors.
Both the under-reproduction and under-estimation observed in the psilocybin studies can be interpreted as regression to the mean, which has been one of the earliest documented temporal biases (Lejeune and Wearden, 2009). Investigating this effect through the Bayesian framework, Jazayeri and Shadlen (2010) have observed it appears to be stronger for longer durations, due to increase in noise compared to shorter intervals and therefore increased reliance on priors. Psychedelic states have been characterised by increased brain entropy (Carhart-Harris et al., 2014) and changes in functional connectivity between brain networks, as well as decreased inter-connectivity within networks. These changes have been found to lead to lower signal-to-noise ratio (Barnett et al., 2020), which – according to findings by Jazayeri and Shadlen (2010) – should lead to stronger temporal bias. Interestingly, the temporal effects of psilocybin occurred only for longer intervals, suggesting that impairments in cognitive processing may be driving the effect. Meta-analysis of the effects of psilocybin on cognitive domains have found it can cause deficits in working memory, attention and response inhibition (Yousefi et al., 2025), which are all necessary for the completion of the timing tasks. Due to the nature of the psychedelic state, which is characterised by increased noise, we can speculate that longer durations (requiring the use of cognitive processes that are impaired) could lead to such high levels of noise that participants respond early (resulting in under-reproduction or under-estimation), as the cost function would favour an early response.
Although we have found significant differences between psilocybin and placebo in both of the measures of time perception collected (BP & JND), as well as in the frequency of self-reported slowing down of subjective time (as evidenced by ASC_Q133), no significant correlations were found between the magnitudes of these measures. This lack of correlation may stem from the ASC questionnaire (and thus ASC_Q133) being administered at the end of the study and assessing the overall experience, rather than the exact timepoint when BP and JND were measured. While the intensity of subjective changes in time perception (as measured by HRS_Q86a) of participants in the psilocybin condition did not significantly correlate with JND, it did correlate with BP. However, the limited response options offered by the HRS questionnaire, which required the participants to rank the intensity of time alteration on a scale from one to three, slightly detract from the confidence in this finding. Such a scale lacks the granularity necessary to be compared with the variability observed in the BP and JND data. Future studies should consider employing a continuous scale, such as a visual analogue scale, to measure the intensity of subjective time perception changes. This approach would enable a more precise correlation between subjective reports and measures of temporal processing collected via bisection task.
Further studies should also aim to investigate the neural mechanisms underlying the effects psilocybin has on temporal processing. Using both 5-HT2A and 5-HT1A antagonists would allow us to tease apart which 5-HT receptor subtypes are playing a role in the changes in time perception under psilocybin. . As administration of 5-HT2A antagonist ketanserin did not attenuate attentional impairments caused by psilocybin (Carter et al., 2005), testing time perception while pre-treating with different antagonists could help clarify the roles of specific disrupted cognitive processes (as it would allow us to test how much of the effects are driven by impairments in attention).
One possible contributing factor to the rightward shift in BP under psilocybin is the greater JND observed for intervals longer than 2000 ms, compared with shorter intervals. This increase in JND would produce a shallower slope on the right side of the psychometric curve, which, when the curve is fitted, could contribute to the observed shift in BP. However, we believe that the observed pattern still reflects a genuine effect on BP, as a strong trend was present for the 2000-ms interval (see Figure 3.). Given that no significant correlation was found between BP and JND for psilocybin condition, with only a slight positive trend (r = 0.23), it is unlikely that the rightward shift in BP can be fully explained by shallower slope on the right due to increased JND.
The limitations of the current study include the relatively small sample size, as the sample size was being optimised for the fMRI part of the study. This limited sample size reduced the statistical power needed to detect additional nuances in the data. Furthermore, both ASC and HRS questionnaires, used here to assess subjective changes in time perception, were administered at the end of the study. This is problematic for two reasons. Firstly, retrospective reports may be less accurate due to memory bias, as participants were asked to recall and generalise their experiences over the entire session. Secondly, the timing of questionnaire administration was not aligned with the period during which BP and JND were collected, making it challenging to directly correlate the data from the questionnaires and bisection task. Furthermore, using isolated questions from the ASC and HRS is not standard practice and their psychometric properties are unknown, although we have attempted to mitigate this by using multiple questions and running an analysis to choose the one with highest loading. As this was an exploratory study, the questions were used only as a rough estimate of subjective time perception. Further studies should validate the psychometric properties of individual timing questions, should they use them. Finally, the temporal bisection task took place 260 minutes post psilocybin administration to minimise the influence of visual hallucinations on participants’ ability to perceive the target stimulus. However, since this period corresponds to the waning phase of psilocybin’s effects, it is possible that conducting the experiment earlier, during the peak effects, could have yielded more pronounced or different results. Using multiple different doses could have also yielded different results. However, as Wackermann et al. (2008) replicated the results of Wittmann et al. (2007) with a very low dose (12 μg/kg), we have decided against using multiple doses to keep the design straightforward. Future studies should also consider controlling for blinding and devising a way to check whether participants refrained from counting during the bisection task, which is something we have not done.
Conclusion
We have shown that time perception is affected after psilocybin administration, both subjectively reported and measured via temporal bisection task. In addition, we have shown that the alteration of time perception under psilocybin appears to be a universal phenomenon, as we used a different sensory modality to present stimuli and a different type of task compared to previous studies. We have supported the existence of a threshold around 2–3 seconds, below which psilocybin does not seem to have an effect on performance in timing tasks. Finally, we have highlighted the role of cognitive disturbances caused by psilocybin in driving the effects on time perception, rather than psilocybin directly affecting speed of the internal clock.
Footnotes
ORCID iDs
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from Czech Health Research Council (project NU21-04-00307, NW24-04-00413 and NU22-04-00526); Czech Science Foundation (project 20-25349S); Long-term conceptual development of research organization (RVO 00023752), program INTER-EXCELLENCE subprogram INTER-ACTION LUAIZ24146), ERDF-Project Brain dynamics, No. CZ.02.01.01/00/22_008/0004643, project VVI CZECRIN (LM2023049), by the Horizon Europe project PsyPal (Grant Agreement No. 101095146) and COST Action PSY-NET (CA24130), and Charles University research program Cooperatio-Neurosciences and private funds obtained via PSYRES, Psychedelic Research Foundation (![]() ).
).
Declaration of conflicting interests
The authors declared the following potential conflicts of interest with respect to the research, authorship and/or publication of this article: T.P., M.B., F.T. and J.H. declare to have shares in ‘Psyon s.r.o’. T.P., M.B. and J.H. founded the ‘PSYRES – Psychedelic Research Foundation’ and have shares in ‘Společnost pro podporu neurovědního výzkumu s.r.o’. T.P. and M.K. have shares in AVI-X Aviation Experts s.r.o. T.P. reports consulting fees from GH Research and CB21-Pharma outside the submitted work. T.P., V.V., V.A., J.H., M.V. and F.T. were/are involved in Compass Pathways, MAPS, GH-Research, Ketabon clinical trial with psilocybin / MDMA / 5-MeO-DMT (mebufotenin) / ketamine trials and F.T. in MindMed study with LSD outside the submitted work. The remaining authors declare no potential conflicts of interest with respect to the research, authorship and/or publication of this article.