
Abstract
Rationale:
Ketamine, known for its rapid-acting antidepressant effects, is often coadministered with other psychoactive drugs, including opioids and cannabinoids, in both clinical and recreational settings. We investigated whether ketamine (R-, S-, or racemic R, S-) isoforms alter the subjective effects of morphine or Δ9-tetrahydrocannabinol (THC) and whether the interoceptive effects of ketamine overlap with these drugs.
Objectives:
Rats were trained to discriminate morphine (6.4 mg/kg) or THC (3 mg/kg) from their respective vehicle controls. Subsequently, different ketamine isoforms were administered at doses of 5, 10, or 30 mg/kg either prior to or instead of the training drug.
Results:
In drug interaction tests, naltrexone (0.1 mg/kg) fully antagonized morphine’s effects, and rimonabant (0.32, 1, and 3.2 mg/kg) partially inhibited THC cue. However, ketamine isoforms did not fully block the discriminative stimulus effects of morphine or THC. In generalization tests, as expected, fentanyl (0.1 mg/kg) fully substituted for the morphine cue, and cannabidiol did not substitute (10 mg/kg) or partially substituted (3 and 50 mg/kg) for the THC cue. However, none of the ketamine isoforms fully substituted for the morphine or THC cue. Ketamine isoforms did not alter response rates in the morphine experiment, but suppressed responding in the THC experiment.
Conclusions:
These findings indicate that ketamine isoforms do not produce opioid- or cannabis-like subjective effects and are unlikely to fully mask the interoceptive effects of morphine or THC. However, their coadministration with cannabinoids may lead to pronounced sedative-like suppression. This warrants further investigation into the safety and interaction profiles of such drug combinations, particularly in therapeutic settings.
Introduction
Several psychotropic drugs are often coadministered in both therapeutic and recreational settings. Clinically, the dissociative anesthetic and NMDA receptor antagonist ketamine is used to enhance opioid (Cohen et al., 2022; Nielsen et al., 2019) and cannabinoid (Reynolds and Osborn, 2013) analgesic effects and lower total analgesic requirements. Recreationally, ketamine is often taken in combination with opioids, cannabis, and other “club drugs” (Barrios et al., 2025; Lankenau and Clatts, 2005; Palamar et al., 2021). Users report combining drugs to augment or prolong euphoric effects or to modulate adverse effects (Grov et al., 2009; Leri et al., 2003). Such polydrug use episodes can involve concurrent or sequential drug consumption within the same use episode (Lankenau and Clatts, 2005). Importantly, drug mixtures can be used intentionally, but often street-drug supply contains unexpected substances, including ketamine mixed in with cannabis, presumably introduced by dealers to boost potency and other desired effects.
In both clinical and recreational settings, ketamine is either coadministered with opioids or cannabis or used in place of them, posing two critical questions. First, does ketamine modify the subjective effects of opioids or cannabinoids, either by amplifying or attenuating their impact? Second, do the interoceptive (subjective) effects produced by ketamine overlap with those of opioids and/or cannabis. To address these questions, we examined whether the combined administration of ketamine with an opioid morphine or (–)-trans-Δ9-tetrahydrocannabinol (THC) diminishes their subjective (discriminative stimulus) effects, an outcome suggesting ketamine’s potential utility in treating drug misuse or overdose, or instead potentiates those effects or suppresses operant responding, suggesting an added harm. In generalization studies, we evaluated whether ketamine’s subjective effects substitute for those produced by morphine and/or THC, suggesting ketamine’s utility as a pharmacological substitute.
Commercially available racemic ketamine is a mixture of 50% S-(+)-ketamine and 50% R-(−)-ketamine, and has been used as a safe and effective anesthetic since the 1970s. Although (S)-ketamine was approved for use for the treatment of depression in 2019 (see Hashimoto (2019) for review), accumulating data have suggested that (R)-ketamine might produce less subjective side-effects than (S)-ketamine in people (Persson et al., 2002). For instance, the affinity of (S)-ketamine for the mu and kappa opioid receptors is two to fourfold that of (R)-ketamine; for review, see Jelen et al. (2021), and (R)-ketamine produces calmness, rather than psychotic-like reactions characteristic of (S)-ketamine (Vollenweider et al., 1997). While ketamine inhibits various manifestations of substance use disorders, for review see (Jones et al., 2018), recent preclinical data suggest the specific utility of its (R)-isomer (Witkin et al., 2020).
In drug discrimination studies, trained rats report the presence of a drug versus its absence, based on the drug’s interoceptive (subjective) effects. The ability of the tested compound to substitute for the training drug has been used as a high-validity predictor of the subjective effects of drugs in humans (Holtzman, 1985; Schuster and Johanson, 1988). In contrast, a tested compound’s ability to alter the training drug’s discriminative stimulus effects is considered as evidence of interference—either inhibition or enhancement—with the action of the training drug. Drug discrimination assay is well validated for morphine (Gianutsos and Lal, 1975; Walker et al., 1994). Morphine “cue” is readily inhibited by opioid receptor antagonists (Shannon and Holtzman, 1976) and mimicked by other opioids (Miksic et al., 1978). The subjective effects of THC in the drug discrimination test have also been sufficiently characterized (Rowton et al., 2020; Wiley et al., 2021a, 2021b). The purpose of this study was to (a) establish the specificity of morphine and THC subjective cues, (b) examine whether ketamine isomers substitute for, and/or (c) inhibit or enhance morphine and THC subjective cues.
Experimental procedures
Animals and ethics
Forty (morphine experiment) and 42 (THC experiment) male Sprague–Dawley rats (Charles River, Germany) weighing ~250 g upon arrival were group housed in standard laboratory cages, under standard colony-controlled conditions: room temperature 21 ± 2°C, humidity (40%–50%), 12-hour light/dark cycle (lights on: 06:00) with ad libitum access to water and with a mild food restriction for at least 1 week prior to training. Rats could acclimatize for at least 7 days before the start of the experimental procedure; during this time, the animals were frequently handled. At least 1 hour before the start of the experiment, rats were transferred to the experimental room for acclimation.
In this study, 17 g per day per animal of the standard laboratory chow was provided ~1 hour after the training or testing sessions, that is, at ~15:00. The animals were group-fed in their home cages. On most of the weekend days, no sessions were conducted, and the animals were given the same amount of 17 g/rat of food starting at 15:00.
All animals were maintained, and experiments were conducted in accordance with the European Guidelines for animal welfare (2010/63/EU) and Enhancing Quality In Preclinical Data (EQIPD) (https://go-eqipd.org/) guidelines. All experimental procedures were approved by the II Local Ethics Committee for Animal Experiments (Approval 2/2022) at the Maj Institute of Pharmacology, Polish Academy of Science, Kraków, Poland. None of the rats had previously received drugs or behavioral training.
Quality system
At the time of performing this study, the Department of Behavioral Neuroscience and Drug Development has been holding EQIPD (Bespalov et al., 2021); certificate no. PL-INS-DBNDD-11-2021-2 (November 12th, 2021–November 30th, 2024); see https://paasp.net/new-eqipd-certified-research-unit/ and https://go-eqipd.org/about-eqipd/eqipd-certified-research-groups/. Thus, this study followed EQIPD guidelines: https://eqipd-toolbox.paasp.net/wiki/EQIPD_Quality_System. For instance, it was carefully designed in advance, was documented in detail at every stage of progress, and was preceded by randomization and effect size calculations.
Thus, experimental conditions were randomized to ensure a balanced design. Because the number of rats meeting the inclusion criteria varied considerably across different tests and experiments, we applied two randomization approaches: (1) automated randomization using a custom R script, applied when the number of eligible rats allowed for block randomization; and (2) manual randomization, balancing design when the number of test-included rats was insufficient for the block assumption.
The effect size (Cohen’s f, for 1-way ANOVA) was estimated based on the results presented by Wiley et al. (1999). For instance, considering only THC groups, SD was calculated based on SEM and N = 8 and were as follows: THC 1 mg/kg SD = 45.25%, THC 3 mg/kg SD = 2.282%; THC 10 mg/kg SD = 45.25%, pooled SD ~36.97. G*Power software (Statistical Power Analyses for Mac and Windows; https://www.psychologie.hhu.de/arbeitsgruppen/allgemeine-psychologie-und-arbeitspsychologie/gpower) estimated that for 6 groups, power = 0.8 and alpha = 0.05, the total N of 30, resulting in 5 rats/group.
Blinding was not used, because all data were provided by MED Associates Software (https://med-associates.com/).
Apparatus
Eight commercially available operant chambers (model E10-10RF; Coulbourn Instruments, Whitehall, PA, USA) equipped with two nose-poke operanda were used to train the rats and to test drug discrimination responses (Popik et al., 2019). A food dispenser delivering 45 mg Dustless Precision Pellets (Bio-Serv, Flemington, NJ, USA) was mounted between the nose-poke operanda and was equipped with a photocell beam and a sensor. The chambers were illuminated with 5 W white house bulbs, contained an exhaust fan that provided continuous masking noise, and were placed in sound- and light-attenuating cubicles. The Coulbourn boxes were adapted to be controlled by Med Associates, St Albans, VT, USA; MED-PC system. Following every session, the instrumental chambers were cleaned to avoid olfactory cues that may be deposited by animals and serve as discrimination cues for other rats tested in the same chamber (Extance and Goudie, 1981).
General procedure
In two separate cohorts, we trained rats to discriminate morphine and THC from vehicles as described earlier (Popik et al., 2020). As the procedures for morphine and THC did not differ, in this section, they are described as “drug.”
Initial training
Animals were trained for 15 min daily to poke into any operandum on a fixed-ratio 1 (FR1) reinforcement schedule, in which each response delivered a food pellet. After the rats had acquired this behavior, they were trained for 15 minutes daily to poke for the pellet on an FR1 schedule in one operandum, while poking into the other operandum had no scheduled consequences. When responding stabilized, training was shifted to reinforcing responses in the operandum formerly non-reinforced. This way, the rats were trained to poke into both operanda.
Discrimination training
The drug discrimination procedure was based on the prior work of Glennon et al. (1983) and Gresch et al. (2007) as described earlier (Popik et al., 2020). Briefly, the animals were trained every weekday at the same time (~09:00–14:00) to discriminate drug from vehicle under a double alternating sequence, with at least two drug and two vehicle sessions per week (e.g., D-D-V-V-D; Popik et al., 2019). For 50% of rats, the left operandum was designated as the drug operandum, and for the remaining animals, the right operandum was so defined. For all animals, the side assignments remained unchanged throughout the study. To facilitate discrimination learning, at the latter stages of the experiment, occasionally, rats received vehicle training in the morning (09:00) session, and drug training 1 hour later. Administration of either a drug or vehicle 30 minutes prior to a variable interval 15 seconds (VI-15) schedule of reinforcement served as the discriminative cue for the correct (reinforced) nose-poke.
The training sessions, during which correct responding was reinforced for the entire 20 minutes, were interspaced with extinction sessions. These sessions (i) were carried out once or twice per week, (ii) were always preceded earlier with the drug and the vehicle training session, (iii) were used to assess the degree of stimulus control exerted by the drug or the vehicle (Glennon et al., 1983) and (iv) served as internal comparator conditions (see Statistics). They consisted of a 2.5-minute non-reinforced testing period, followed by a “normal” 17.5-minute training period, during which the correct responses were reinforced (Stadler et al., 1999).
Compound testing sessions
Once a stable level of performance was attained at extinction sessions (inclusion criterion: ⩾85% of responses on the drug-appropriate operandum after administration of the drug and ⩽15% of responses on the same operandum after administration of the vehicle), the compound testing sessions had started. These sessions were carried out once a week, usually on Fridays or Saturdays, and were always preceded by extinction sessions with the drug and vehicle given 1–2 days earlier. The compound testing sessions were done exactly as the extinction sessions: they lasted for 2.5 minutes, and were never reinforced to prevent new discrimination learning (Colpaert, 1987).
Immediately after the testing session was terminated, the rat was removed from the testing chamber and placed in his home cage.
Drugs
Doses of morphine used in drug discrimination experiments vary across studies, for instance Shannon et al. (1976) used it at 3.0 mg/kg SC 30 minutes before session, Walker et al. (1994) used it at 3.2 mg/kg SC (with ED50 of 1.5 mg/kg) and Gianutsos et al. (1975) used it at 10 mg/kg IP, 45 minutes before session. We decided to use morphine HCl (Polfa, Kraków, Poland) at a dose of 3.2 mg/kg. Since after several sessions the animals lost the ability to discriminate this dose from vehicle, the training morphine dose was increased to 6.4 mg/kg and remained at this level to the end of the study. Morphine doses are expressed as base: calculated as anhydrous (88.7% of pure morphine); the drug was dissolved in physiological 0.9% saline (vehicle) and given IP 30 minutes before sessions.
Commercially available solution of fentanyl (Polfa, Warszawa, Poland) was used at doses of 0.032 and 0.1 mg/kg, which were selected based on earlier work of Walker et al. (1994) and Koek et al. (1993). Fentanyl was dissolved in sterile water (vehicle) and given IP 15 minutes before the test.
Naltrexone (Tocris, UK) was dissolved in physiological 0.9% saline and was administered SC 30 minutes prior to morphine. The doses (0.032 and 0.1 mg/kg) were selected based on earlier work of Walker et al. (1994) and Koek et al. (1993).
Δ9-THC was obtained from Research Triangle Institute, NC, USA, through the National Institute on Drug Abuse (NIDA; Bethesda, MD, USA) Drug Supply Program. The training and testing (3 mg/kg, IP, 30 minutes before test) dose of THC was based on the Wiley et al. (2021a) study. THC was suspended in a 7.8% polysorbate (Tween) 80/0.1% ethanol and 92.1% (=100%) saline mixture (vehicle; Wiley et al., 2021b).
Cannabidiol (CBD, Sequoya Biotechnologies, Poland) was suspended in Tween 80 (7.8%), 0.9% NaCl (92.1%), EtOH (0.1%; vehicle), and given IP, 40 minutes before the test. CBD doses (3, 10, and 50 mg/kg) were selected based on Vann et al.’s (2008) work. These authors trained Long-Evans rats to discriminate THC (3 mg/kg, IP) and reported that CBD (0.3–30 mg/kg, IP) did not substitute or alter THC discrimination. Because CBD only at doses 30–100-fold higher than the THC dose potentiated and/or prolonged THC discrimination (Hiltunen and Jarbe, 1986), CBD at a dose of 50 mg/kg was also included in this study.
The cannabinoid 1 receptor antagonist/inverse agonist rimonabant (SR141716, Cayman Chemicals, USA) was given IP 10 minutes prior THC, 40 minutes before test (Marusich and Wiley, 2012) at doses suggested by Wiley et al. (2004) (1 and 3 mg/kg) and by Jarbe et al. (2010); (1 mg/kg). Rimonabant was suspended in Tween 80 (7.8%), 0.9% NaCl (92.1%), EtOH (0.1%; vehicle.
Racemic (R,S)-ketamine hydrochloride was purified from the Vetoquinol Biowet racemic ketamine hydrochloride (aqueous solution (115.34 mg/mL), Gorzów Wielkopolski, Poland) preparation by Dr. Ryszard Bugno at the Department of Medicinal Chemistry of the Maj Institute of Pharmacology as described earlier (Popik et al., 2022), because the commercial preparation contains minute amounts of the sedative, weak local anesthetic, antibacterial, and antifungal compound chlorobutanol hemihydrate. The Department of Medicinal Chemistry also provided pure (R)- and (S)-ketamine. Ketamine doses (5, 15, and 30 mg/kg) were selected based on earlier work of Koek et al. (1993) who in male Wistar rats trained to discriminate fentanyl, used racemic ketamine at doses of 5, 10, and 20 mg/kg (the doses of 10 and 20 mg/kg remarkably reduced response rates) and on our previous reports (Malikowska-Racia et al., 2023; Popik et al., 2020, 2022). In drug-interacting studies, ketamine racemate/isomers were given 15 minutes before morphine/THC. In drug substitution studies, ketamine racemate/isomers were given 45 minutes before the test. All ketamine isoforms were dissolved in sterile MilliQ water prior to injection and given IP.
Study design
Morphine
Control experiments with morphine as the training drug started by establishing morphine dose-related effects: morphine was used at 0 (vehicle), 1.06, 2.1, 4.2, and 6.4 (morphine training dose) mg/kg. To examine whether an opioid receptor antagonist affects (inhibits) morphine cue, naltrexone (0, 0.5, or 3.2 mg/kg) was given prior to the training dose of morphine. To observe a generalization of another opioid receptor agonist to the morphine cue, fentanyl (the “positive control”) was used at 0.032 and 0.1 mg/kg.
THC
Control experiments with THC as the training drug started by establishing the THC dose-related effects: THC was used at 0 (vehicle), 0.3, 1, 3 (THC training dose), and 10 mg/kg. To examine whether cannabinoid receptor antagonist affects (inhibits) THC cue, rimonabant (0, 0.32, 1, or 3.2 mg/kg) was given prior to the THC training dose. To check the specificity of THC effect, that is, whether no generalization of the minor cannabinoid occurs to the THC cue, CBD (the “negative control” (Vann et al., 2008)) was used at 3, 10, and 50 mg/kg.
Morphine, THC, and ketamine isomers
To examine whether morphine or THC subjective cues are affected by ketamine isomers, (R)-, (S)-, or (R,S)-ketamine (0, 5, 15, or 30 mg/kg) were given IP 45 minutes before the test and 15 minutes prior to the morphine or THC training doses. To examine whether ketamine isomers could generalize to morphine or THC subjective cues, ketamine isoforms (0, 5, 15, or 30 mg/kg) were given 45 minutes before the session.
Statistics
The dependent measures included the percentage of morphine-like or THC-like responses and the response rate (number of total nose-pokes per minute) exhibited by animals during drug testing sessions, and during extinction sessions preceding drug testing sessions (both lasting for 2.5 minutes).
To estimate the dose of drug required to produce 50% drug response (ED50) with 95% confidence limits, response accuracy data from vehicle and each drug dose tested were log-transformed and fitted using GraphPad Prism’s 9 (https://www.graphpad.com/) sigmoidal dose-response equation: Y = Bottom + (Top-Bottom)/(1 + 10^((LogEC50-X) * HillSlope)). The bottom and the top of the curve were fixed at 0% and 100%, respectively. In addition, one-way ANOVAs were used to examine whether morphine or THC drug-like responses and response rates were altered by increasing doses of training drugs (Figures 1 and 3).
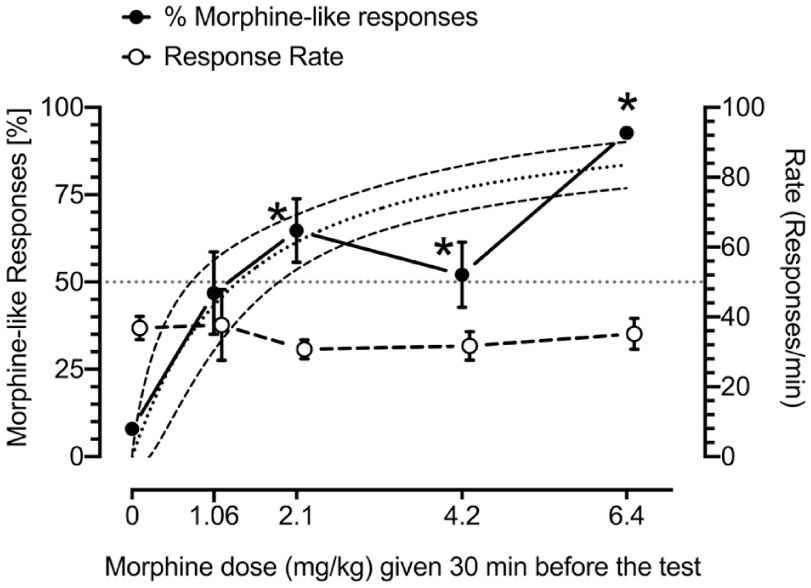
Increasingly higher morphine doses produce increases in morphine-like responses (black circles) and do not affect response rates (open circles) in rats trained to discriminate 6.4 mg/kg of morphine from vehicle. The nonlinear regression with variable slope function on morphine-like responses is presented as the line with large black dots, and confidence intervals (CI) are presented as lines with small black dots. The figure shows mean ± SEM data. The N of rats tested for doses of 0, 1.06, 2.1, 4.2, and 6.4 mg/kg were 25, 7, 8, 10, and 25, respectively, and varied among groups because the analyses included the responses to vehicle (0) and the training dose of morphine (6.4 mg/kg) for all animals the day before testing intermediate doses. Symbols: *p < 0.05 as compared with vehicle (Tukey’s HSD post hoc test followed by one-way ANOVA; see Table 1 for details). The precise P level is not shown for the sake of figure clarity.

Percentage of morphine-like or THC-like responses
For each individual drug interference or substitution study (Figures 2 and 4), statistical comparisons with one-way ANOVAs included data from the given drug and vehicle extinction sessions preceding the test. While responses in these control conditions did not differ statistically, this was done to provide the most accurate individual comparator datasets, because compounds were tested over a long time. Since homogeneity of variance (Levene’s tests) on ANOVA results revealed a violation for some datasets, we decided to use the most conservative Tukey’s HSD post hoc test (Cardinal and Aitken, 2006; Field, 2009). The criteria for drug substitution used here were similar to those originally proposed by Colpaert (1987) and by Fiorella et al. (1995) and were determined by Tukey’s HSD post hoc tests: (1) full substitution/no antagonism: the test compound or combination elicits a level of drug-appropriate responding not significantly different from that elicited by the drug training condition and significantly different from that elicited by vehicle condition; (2) partial substitution/partial antagonism: the test compound or combination elicits a level of drug-appropriate responding significantly different from both the drug and vehicle training conditions; (3) no substitution/full antagonism: the test compound or combination elicits a level of drug-appropriate responding not significantly different from the vehicle training condition and significantly different from that elicited by the drug training condition.
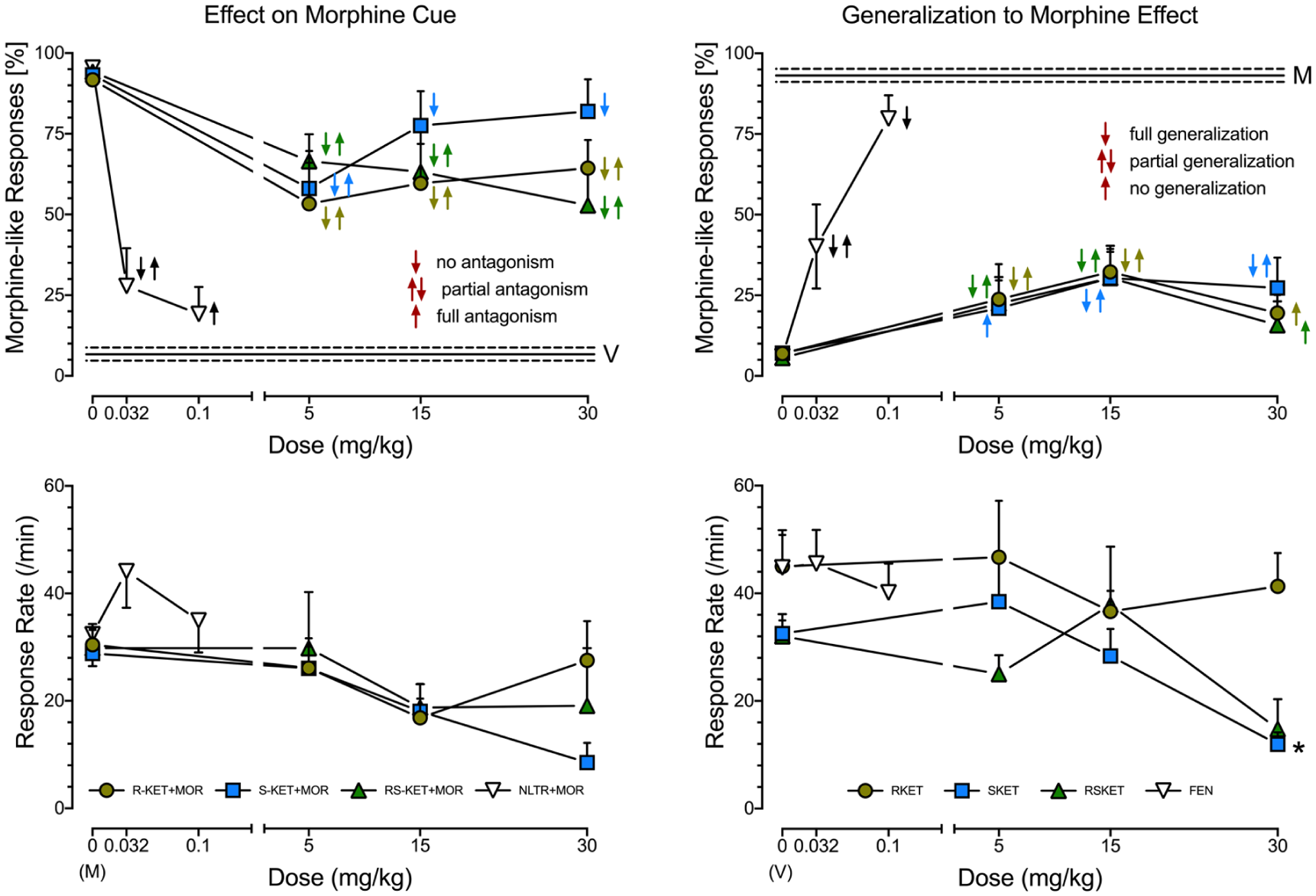
Opioid receptor antagonist naltrexone partially (at 0.032 mg/kg) or fully (at 0.1 mg/kg) inhibits morphine discriminative stimulus (upper left panel) and does not affect response rates (lower left panel). Almost all of the ketamine isomers produced partial antagonism toward the morphine cue. No antagonism was observed for (S)-ketamine at 30 mg/kg. Given with morphine, ketamine isomers did not affect response rates. The N for naltrexone + morphine was 7 rats. For (R)- and (S)-ketamine the Ns were 7-8 rats. For (R,S)-ketamine, the Ns were 4–8 rats. Fentanyl partially (at 0.032 mg/kg) or fully (at 0.1 mg/kg) substitutes for the morphine cue (upper right panel) and does not affect response rates (lower right panel) in rats trained to discriminate saline from 6.4 mg/kg of morphine. (R-), (S)- and (R,S)-ketamine display partial or no generalization for the morphine discriminative stimulus. Only (S)-ketamine (at 30 mg/kg) reduces response rates. For (R)-, (S)- and (R,S)-ketamine, the Ns were 7–8 rats. Symbols: ↓ (no antagonism or full generalization p < 0.05 vs vehicle but not drug training dose); ↑↓ (partial antagonism or partial generalization, p < 0.05 vs vehicle and drug training dose); ↑ (full antagonism or no generalization, p < 0.05 vs drug training dose but not vehicle); *p < 0.05 versus vehicle (Tukey’s HSD post hoc test followed by one-way ANOVA; see Table 1 for details). The figure presents raw mean ± SEM data.
Response rates
Individual analyses of response rates with one-way ANOVAs followed by Tukey’s HSD post hoc test-included data from the given drug (interference) or vehicle (substitution) extinction sessions preceding the test.
Statistical analyses were done with StatSoft, Inc. STATISTICA (data analysis software system), version 12. www.statsoft.com. The alpha level was set at 0.05.
Results
Morphine produced dose-related increases in morphine-like responses with an ED50 of 1.369 mg/kg (CI: 0.7381–1.935 mg/kg, Hill’s slope = 1.052); the doses of 2.1, 4.2, and 6.4 mg/kg significantly (p < 0.05) increased morphine-like responses. No effect was observed on response rate (Figure 1).
Effect on morphine cue
At 0.032 mg/kg partially and at 0.1 mg/kg, naltrexone fully inhibited morphine cue (Figure 2 upper left panel) without affecting response rate (Figure 2 lower left panel). (R)-, (S)-, and (R,S)-ketamine at all doses tested produced only partial antagonism toward the morphine cue, as did (S)-ketamine at 5 mg/kg. (S)-ketamine did not inhibit the morphine cue at the doses of 15 and 30 mg/kg, however (Figure 2 upper left panel). At doses tested, none of the ketamine isomers affected response rates when combined with the morphine training dose (Figure 2, lower left panel). Thus, while the opioid receptor antagonist fully inhibited morphine’s discriminative cue, none of the ketamine isomers produced a similar effect.
Generalization to morphine cue
At 0.032 fentanyl partially and at 0.1 mg/kg completely generalized to morphine cue (Figure 2 upper right panel), without affecting response rate (Figure 2 lower right panel). (R)- and (R,S)-ketamine at 5 and 15 mg/kg only partially generalized to the morphine cue, while both isomers at 30 mg/kg did not generalize to the morphine cue. In contrast, (S)-ketamine at 5 mg/kg did not, and at 15–30 mg/kg partially generalized to the morphine cue (Figure 2 upper right panel). At doses tested, none of the ketamine isomers affected response rates when combined with the morphine training dose, except (S)-ketamine, which, at 30 mg/kg, inhibited response rates (Figure 2, lower left panel). Thus, while opioid receptor agonists fully generalized to morphine’s discriminative cue, none of the ketamine isomers produced a similar effect.
THC produced dose-related increases in THC-like responses with ED50 of 0.2558 mg/kg (CI: 0.08814–0.5038 mg/kg, Hill’s slope = 0.6201). The doses of 3 and 10 mg/kg significantly (p < 0.05) increased THC-like responses. No effect was observed on response rate (Figure 3).
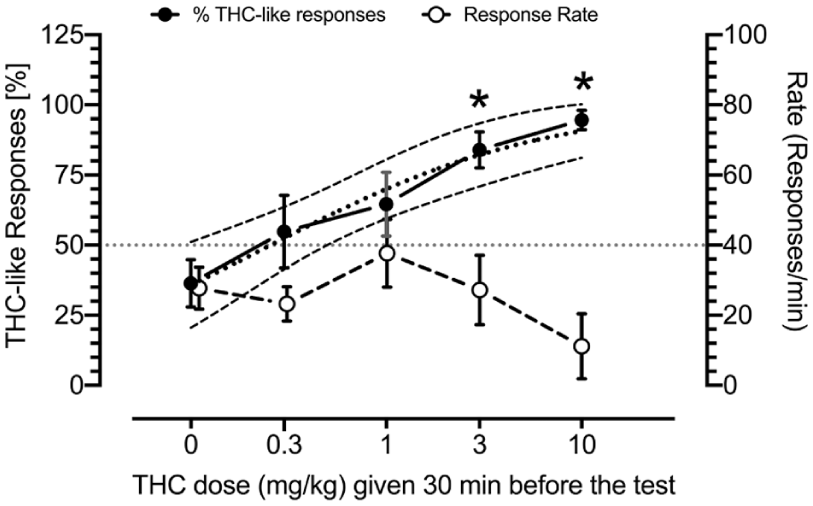
Increasingly higher THC doses produce increases in THC-like responses (black circles) and do not affect response rates (open circles) in rats trained to discriminate 3 mg/kg of THC from vehicle. The nonlinear regression with variable slope function on THC-like responses is presented as the line with large black dots, and confidence intervals (CI) are presented as lines with small black dots. The figure shows mean ± SEM data. The N of rats tested for THC doses of 0, 0.3, 1, 3, and 10 mg/kg were 7, 7, 8, 8, and 6, respectively. Symbols: *p < 0.05 as compared with vehicle (Tukey’s HSD post hoc test followed by one-way ANOVA; see Table 1 for details).
Effect on THC cue
At doses of 0.32, 1, and 3.2 mg/kg, rimonabant only partially inhibited THC cue (Figure 4 upper left panel), and at the dose of 3.2 mg/kg, it inhibited response rate (Figure 4 lower left panel). (R)-ketamine at 5 and 15 mg/kg did not affect, and at 30 mg/kg it partially inhibited the THC cue. Similarly, (S)-ketamine at 5 and 15 mg/kg partially decreased, and at 30 mg/kg did not affect the THC cue. At 15 mg/kg, (R,S)-ketamine did not affect, and at 5 and 30 mg/kg, it partially decreased the THC cue (Figure 4, upper left panel). The effects of ketamine isomers were accompanied by significant decreases in response rates (Figure 4, lower left panel). Thus, in the present experimental conditions, neither cannabinoid receptor antagonist nor ketamine isomers fully inhibited THC’s discriminative cue but reduced response rates.
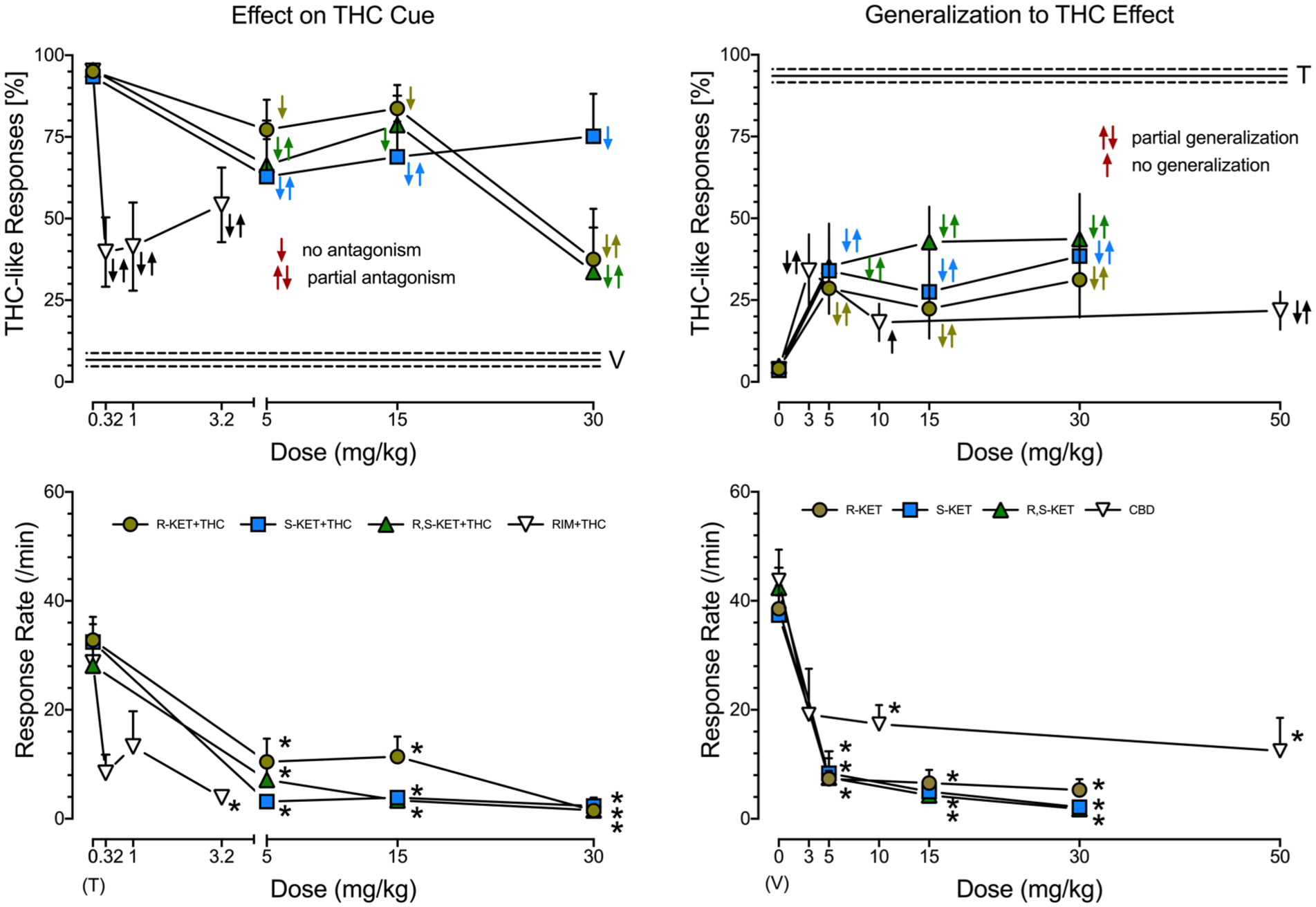
Cannabinoid receptor antagonist rimonabant partially inhibits THC discriminative stimulus (upper left panel) and, at the highest dose of 3.2 mg/kg, inhibits response rate (lower left panel). Almost all of the ketamine isomers produced partial antagonism toward the THC cue. No antagonism was observed for (R)-ketamine (5 and 15 mg/kg), (S)-ketamine (30 mg/kg) and (R,S)-ketamine (15 mg/kg). Given with THC, all ketamine isomers reduced response rates. The N for rimonabant + THC was 7–8 rats. For (R)- and (S)-ketamine the Ns were 9–10 rats. For (R,S)-ketamine the Ns were 10–11 rats. CBD did not (10 mg/kg) or partially (3 and 50 mg/kg) substitute for the THC cue (N = 7 rats per dose; upper right panel) and at 10 and 50 mg/kg reduced response rates (lower right panel) in rats trained to discriminate saline from 3 mg/kg of THC. All of ketamine isomers displayed partial generalization to the THC discriminative stimulus and reduced response rates. For (R)-, (S)- and (R,S)-ketamine, the Ns were 10–13 rats. For Symbols, see Legend to Figure 2.
Generalization to THC cue
At 3 and 50 (but not at 10 mg/kg), CBD only partially generalized to THC cue (Figure 4, upper right panel); in addition, doses of 10 and 50 mg/kg decreased response rate (Figure 4, lower right panel). All of the ketamine isomers at doses tested only partially generalized to the THC cue (Figure 4, upper right panel) and all reduced response rates (Figure 4, lower right panel). Thus, as expected, CBD’s stimulus did not generalize to THC’s cue, though doses of 10 and 50 reduced response rates. Ketamine isomers did not generalize to the THC cue but reduced response rates.
Discussion
Present results confirm earlier observations demonstrating that both morphine and THC may produce reliable interoceptive stimuli, purportedly associated with their psychoactive actions. The ED50 of morphine (1.4 mg/kg, Figure 1) lies in the same dose range as in previous studies (see Introduction), and the ED50 of THC (0.26 mg/kg) is in the same dose range as in Martin et al.’s (1991) seminal work (0.6 mg/kg). Of note, THC produced an insignificant though apparent and expected reduction in response rate at 10 mg/kg, which agrees with its catalepsy ED50 of 6 mg/kg (Martin et al., 1991).
The internal control conditions used in this study provided an expected outcome, though not in all cases. Thus, naltrexone significantly antagonized morphine stimulus, fentanyl fully substituted for morphine’s cue, and the minor cannabinoid CBD did not generalize for THC (Wiley et al., 2024). However, the cannabinoid receptor antagonist rimonabant, purportedly due to the relatively high THC training dose, did not fully antagonize the THC cue and significantly reduced the response rate, like in Solinas et al.’s (2003) work. The absence of full THC antagonism could be explained by the relatively short in vivo half-life of rimonabant’s free base (the form used also in this study; Jarbe et al., 2010).
Discriminative stimulus
To our knowledge, no prior studies tested the effects of pure (S)- or (R)-ketamine on morphine or THC discriminative cues. We failed to identify a ketamine dose or isoform that fully inhibited the discriminative stimulus effect of morphine or THC. In drug interaction studies, ketamine isoforms produced only partial antagonism, with the prominent exception of (S)-ketamine, which at 30 mg/kg did not alter either morphine or THC cue. In this series of experiments, ketamine isomers produced dose-unrelated effects. Ketamine isoforms—induced partial antagonism is consistent with Koek et al. (1993) work, who noted that a low-efficacy agonist can partially antagonize the effects of a full agonist when administered at doses producing partial agonist effects on their own. Also in this study, ketamine enantiomers reduced the drug-like responding produced by morphine and THC to levels comparable to those observed when administered alone.
Ketamine isomers also failed to fully substitute for the discriminative stimulus effects of either morphine or THC. Our findings are consistent with prior reports showing that racemic ketamine produced only partial generalization to the opioid receptor agonist cue in animals trained to discriminate fentanyl (in rats) or morphine (in pigeons) from saline (Herling et al., 1980; Koek and Woods, 1989). From this perspective, the present data showing partial generalization should not be considered as surprising. While the effects of ketamine on THC discriminative cues have not been previously studied in rats, our findings agree with previous reports from C57BL/6J mice demonstrating that (R,S)-ketamine (3–32 mg/kg, IP) partially substitutes for THC in mice trained to discriminate THC (10 mg/kg, IP), producing max. ~25% THC responding (McMahon et al., 2008). The results from rhesus monkeys further support our observations, as (R,S)-ketamine (0.1–1 mg/kg, IV) only marginally generalizes to THC (0.1 mg/kg, IV), producing max. ~25% of THC responding (McMahon, 2006). (R,S)-ketamine (0.032–1 mg/kg (IV) also failed to generalize to non-selective cannabinoid agonist JWH-073 (0.1 mg/kg, IV), mimicking max. of ~20% responding.
Response rates
The interaction between ketamine isoforms and the THC discriminative stimulus is difficult to assess, as all doses of ketamine significantly suppressed response rates, thereby complicating interpretation. The reduction in response rates observed with THC alone at 10 mg/kg (Vann et al., 2008; Wiley et al., 2024), by rimonabant and THC combination (Wiley et al., 2004), and by (R,S)-ketamine with THC in mice (McMahon et al., 2008) resemble the suppressant effects seen with ketamine and THC combinations. As cannabinoid-induced antinociception is one of the four defining characteristics of cannabinoid pharmacology (Martin et al., 1991), a marked suppression of operant responding following co-administration with ketamine may reflect enhanced sedative or motor-impairing effects, which could be relevant in the context of pain management (Reynolds and Osborn, 2013). However, given the absence of a dose-response relationship, the failure of specific combinations, such as (R)-ketamine (5 and 15 mg/kg), (S)-ketamine (30 mg/kg) and (R,S)-ketamine (15 mg/kg), to alter the THC cue should not entirely preclude ketamine’s utility in enhancing, for example, cannabis-induced antinociception. In the present experimental conditions, in animals trained to discriminate morphine, the response rates were not affected. However, in THC-trained animals, all ketamine isomers markedly suppressed response rates. This may suggest a carry-over or a non-specific sedative effect that, to our knowledge, has not been reported previously in THC drug discrimination studies.
To our knowledge, this is the first report investigating the effects of (R)- and (S)-ketamine optical isomers in morphine and THC drug discrimination paradigms. Present results suggest that none of the ketamine isoforms may produce morphine- or THC-like subjective effects. Based on the interaction studies, we hypothesize that ketamine isoforms are unlike to fully alter opioid or THC subjective effects, but may rather produce significant sedative effects when combined with cannabinoids.
Footnotes
Acknowledgements
The authors thank the Department of Medicinal Chemistry, Maj Institute of Pharmacology, Polish Academy of Sciences, for providing (R)- and (S)-ketamine and the National Institute on Drug Abuse (NIDA; Bethesda, MD, USA) NIDA Drug Supply Program, for the supply of Δ9-THC.
Ethical considerations
Author contributions
PP designed research; NM-R and JG performed research; PP analyzed data; PP, AB, and NM-R wrote the paper; KS and PZ provided essential parts of funding.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The study was supported by the statutory activity of the Maj Institute of Pharmacology, Polish Academy of Sciences, and in part by the NCN OPUS 2021/43/B/NZ7/02855 and NCN OPUS 2021/43/B/NZ7/01162 grants.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
The datasets used and/or analyzed during the current study are available from the corresponding author on request.