
Abstract
Background:
Posttraumatic stress disorder (PTSD) and major depressive disorder are often comorbid in humans. Psilocybin reportedly has beneficial therapeutic effects on depression, possibly by promoting neuroplasticity. PTSD is associated with the dysregulation of neuroplasticity in the hippocampus and medial prefrontal cortex (mPFC). We hypothesized that psilocybin might reduce fear memory by promoting neuroplasticity in the hippocampus and mPFC.
Aims:
We investigated the effects of psilocybin on fear memory and explored its underlying mechanisms. We generated a mouse model of PTSD via auditory-cued fear conditioning and treated the mice with either vehicle or psilocybin (2.5 mg/kg, intraperitoneal) on day 0. Fear memory was assessed by the percentage of freezing time in response to conditioned stimuli. Fear memory tests were conducted on days 1, 6, and 7, after which the mice were sacrificed. To investigate the role of neuroplasticity in mediating the effects of psilocybin on fear memory, we assessed structural neuroplasticity and neuroplasticity-associated marker protein levels in the hippocampus and mPFC 7 days after a single dose of psilocybin.
Results:
Psilocybin reduced the cue-induced fear response on days 1, 6, and 7. Psilocybin ameliorated the fear conditioning-induced decreases in neuroplasticity in the hippocampus and mPFC. Through Golgi-Cox staining, Western blotting, and immunofluorescence staining, we found that psilocybin increased dendritic branches and spine density, upregulated GluR1 and synapsin-1, enhanced brain-derived neurotrophic factor and mammalian target of rapamycin signaling, and promoted neurogenesis.
Conclusions:
A single dose of psilocybin reduces both the rapid and sustained fear memory in mice, at least in part by restoring neuroplasticity in the hippocampus and mPFC. These findings indicate that psilocybin has significant potential for use in the treatment of PTSD and other mental disorders characterized by fear memory.
Introduction
Posttraumatic stress disorder (PTSD) is a pervasive and debilitating psychiatric condition that can develop following exposure to traumatic events. It is characterized by intrusive memories, avoidance behaviors, negative alterations in cognition and mood, and heightened arousal (Shalev et al., 2017). PTSD is recognized as a major public health concern, affecting up to 7% of the population worldwide during their lifetime (Shalev et al., 2017). PTSD frequently co-occurs with major depressive disorder, complicating diagnosis and treatment due to overlapping symptoms and mutual symptom exacerbation (Zhang et al., 2022). Current treatments, including cognitive-behavioral therapy and selective serotonin reuptake inhibitors, show limited efficacy, leaving a substantial proportion of patients symptomatic (Du et al., 2023). Consequently, research has focused on developing compounds that modulate the neurobiological substrates of PTSD.
Psilocybin, a naturally occurring serotonin 5-HT2A receptor agonist (NbN: serotonin receptor agonist, 5-HT2A), has attracted attention for its therapeutic potential. Traditionally employed in spiritual ceremonies, psilocybin is now being explored for its psychiatric therapeutic potential (Madras, 2022). Growing evidence indicates that psilocybin, whether administered alone or in conjunction with psychotherapy, can significantly alleviate depressive and anxiety symptoms within the first 24 hours following ingestion. Moreover, these benefits are sustained, lasting up to 18 months after treatment. Psilocybin produces acute, transient psychological and physiological effects—including transient anxiety, nausea, headache, and challenging psychological experiences (e.g., fear, paranoia)—that typically resolve within 24–48 hours (Yerubandi et al., 2024). Despite these acute effects, psilocybin offers a favorable safety profile compared to conventional antidepressants, with no evidence of lasting neurotoxicity or dependence, and serious adverse events being rare in controlled clinical settings with proper screening and supervision (Carhart-Harris et al., 2016; Davis et al., 2021; Goodwin et al., 2022; Griffiths et al., 2016; Gukasyan et al., 2022; Ross et al., 2016). In a phase II randomized double-blind trial, psilocybin had significantly greater antidepressant effects than those of escitalopram, a frontline antidepressant medication (Carhart-Harris et al., 2021). The most recent systematic review further corroborates that psilocybin significantly improves depressive symptoms (Metaxa and Clarke, 2024).
Psilocybin’s antidepressant effects are hypothesized to involve enhanced neuroplasticity in the hippocampus and medial prefrontal cortex (mPFC; Cameron et al., 2021; Du et al., 2023; Raval et al., 2021; Shao et al., 2021; Siegel et al., 2024). Neuroplasticity in these two brain regions is often dysregulated in individuals with PTSD (Bremner et al., 1995; He et al., 2018; Karl et al., 2006; Wang et al., 2021; Yabuki and Fukunaga, 2019). While recent studies have demonstrated that psilocybin can facilitate fear extinction (Catlow et al., 2013; Du et al., 2023; Hagsäter et al., 2021; Woodburn et al., 2024), its long-term effects on cued fear memory persistence (beyond 24 hours) and the underlying structural neuroplasticity mechanisms in PTSD-relevant circuits require further investigation. Specifically, few studies have examined how psilocybin influences structural synaptic plasticity (dendritic branching and spine density) in the hippocampus and mPFC following fear conditioning, or whether these neuroplastic changes persist over extended time periods. This gap limits our understanding of psilocybin’s therapeutic potential for trauma-related disorders. We hypothesize that psilocybin reduces fear memory and enhances neuroplasticity in the hippocampus and mPFC and thus may represent a novel treatment for PTSD.
Auditory-cued fear conditioning is a widely used paradigm to model trauma-induced fear memory dysregulation observed in PTSD, though it represents a form of adaptive emotional learning rather than the full clinical syndrome of PTSD. We aimed to investigate the effects of psilocybin on fear memory in a mouse model of auditory-cued fear conditioning and to explore its underlying mechanisms. In addition, we examined structural neuroplasticity and the expression of neuroplasticity-associated marker proteins in the hippocampus and mPFC to understand the role of neuroplasticity in mediating the effects of psilocybin on fear memory.
Materials and methods
Animals and housing
Adult male C57BL/6J mice aged 10 weeks were sourced from Beijing SPF Biotechnology Co., Ltd., Beijing, China. The mice were housed in plastic enclosures with ad libitum access to water and food. The housing environment was maintained on a 12-hour light-dark cycle, with lights on from 8:00 AM to 8:00 PM, and controlled temperature and humidity levels of 26°C ± 2°C and 50% ± 5%, respectively. Prior to undergoing fear conditioning, the mice were allowed to acclimate to standard conditions for a period of 2 weeks. Behavioral testing was conducted between 9:00 AM and 5:00 PM with counterbalanced group assignment to control for circadian fluctuations in corticosterone. The experimental design aimed to limit the distress and discomfort of the animals and to use the minimum number of mice necessary. The study protocol, including the care and handling of the mice, was approved by the Institutional Animal Care and Use Committee of the Beijing Key Laboratory of Neuropsychopharmacology (approval number IACUC-DWZX-2022-661). Forty-eight C57BL/6J mice were randomly divided into three groups (n = 16 in each group), namely, the non-fear-conditioned group FC (–) Veh, the fear-conditioned group FC (+) Veh, and the psilocybin group FC (+) PSI. All behavioral testing, video analysis, and histological quantification were performed by experimenters blind to treatment group assignment.
Drugs
Psilocybin, a naturally occurring serotonin 5-HT2A receptor agonist, was produced by the Beijing Institute of Pharmacology and Toxicology. Psilocybin was dissolved in physiological saline to prepare a solution for intraperitoneal injection at a dose of 0.1 mL per 10 g of mouse body weight. During our initial trials, a range of doses for psilocybin was tested, including 0.1, 0.5, and 2.5 mg/kg. Upon evaluation, the most effective dose was determined to be 2.5 mg/kg (Du et al., 2023).
Fear conditioning procedure
This study utilized the fear conditioning paradigm (Du et al., 2023; Karpova et al., 2011). The paradigm involved two unique environments: context A, a transparent acrylic chamber, and context B, a triangular chamber crafted from opaque black and white acrylic. Both contexts were fitted with devices for delivering electrical shocks and emitting auditory tones to achieve fear conditioning.
During the fear conditioning session (D −2), the mice were initially introduced to context A for a 120-second adaptation and exploration phase. Subsequently, the mice were subjected to five auditory cues (80 dB, 5000 Hz, each lasting 30 seconds) as the conditioned stimuli (CS) paired with a 0.8 mA foot shock (2 seconds) as the unconditioned stimulus (US). The interval between successive CS-US pairs, known as the intertrial interval (ITI), was set at 120 seconds, with the US and CS ending concurrently. Following the final foot shock, the mice were kept in the fear conditioning chamber for an additional 30 s. The entire fear conditioning process lasted 14.5 minutes.
Fear memory test
Each group was administered either physiological saline or psilocybin at a dosage of 2.5 mg/kg via intraperitoneal injection. All fear memory tests were conducted in the absence of foot shock. On the first (D1) and sixth (D6) days after administration, mice were placed in context B and presented with four CS trials, each separated by a 30-second ITI. On the seventh day (D7), mice were tested in context A under the same CS presentation protocol (Du et al., 2023). Day 7 testing in context A assesses combined contextual + auditory-cued fear, while days 1 and 6 in context B assess auditory-cued fear only. Aggregate freezing was calculated as the sum of freezing time (in seconds) across the four CS trials, expressed as a percentage of the total CS duration (120 seconds; Lonsdorf et al., 2017). After each mouse was tested, their excrement was removed, and the stainless-steel grid and tray were wiped with 75% alcohol (context A) or 0.5% acetic acid (context B) to prevent the influence of odors on the fear memory of subsequent mice.
Freezing behavior definition and analysis
Freezing behavior was defined as the complete cessation of all movement except respiration and heartbeat, serving as a measure of the fear response. Freezing time was recorded and analyzed using an FC video analysis system (Beijing Zhongshidichuang Science and Technology Development Co., Ltd., Beijing, China). Videos were acquired at 30 frames/second under infrared illumination to eliminate visual cue interference. The automated system quantified freezing via frame-by-frame motion detection algorithms. The primary dependent variable was the percentage of freezing time relative to the total duration of CS presentation (4 trials × 30 seconds; 5000 Hz, 80 dB tone).
Open field test
The open field test (OFT) apparatus was supplied by Beijing Zhongshidichuang Technology Development Co., Ltd. (Beijing, China). The OFT was performed on D1, 2 hours prior to the fear memory test. Locomotor activity was assessed by measuring the total distance traveled. The procedure followed the protocol described in our previous study (Du et al., 2023). The mice were positioned at the center of the open field arena (40 cm × 40 cm × 40 cm), which was sectioned into 16 equivalent regions, and permitted to roam freely. Following an initial 2-minute acclimation period, the tracking system captured and analyzed the distance covered by the mice during the subsequent 5-minute interval using an advanced video tracking system (Smart version 3.0; Panlab, Barcelona, Spain).
Western blotting
Following the behavioral assessments on D7, a random selection of five mice was subjected to Western blotting. For Western blotting, mice were deeply anesthetized with 1% sodium pentobarbital (100 mg/kg, i.p.), followed by transcardial perfusion with ice-cold 0.01 M phosphate-buffered saline (PBS; pH 7.4). Hippocampi and mPFC were rapidly dissected, flash-frozen in liquid nitrogen, and stored at −80°C until analysis. The hippocampi and mPFC were extracted from the animals and treated with radioimmunoprecipitation assay buffer and protease inhibitors for tissue lysis. The samples were homogenized and centrifuged, after which the supernatant was harvested for protein concentration determination. Proteins from the total extracts were heated for 5 minutes for denaturation and then applied to a 10% acrylamide gel for electrophoretic separation. Following electrophoresis, the resolved proteins were transferred to a 0.45 μm polyvinylidene fluoride membrane, which was subsequently blocked with 5% milk powder. The membrane was incubated overnight at 4°C with the following primary antibodies: anti-brain-derived neurotrophic factor (BDNF; 1:1000, 108319; Abcam, Cambridge, UK ), anti-mammalian target of rapamycin (mTOR; 1:1000, 2983; Cell Signaling Technology, Danvers, MA, USA), anti-GluR1 (1:1000, 13185; Cell Signaling Technology, Danvers, MA, USA), anti-synapsin-1 (1:1000, 5297; Cell Signaling Technology, Danvers, MA, USA), and anti-β-actin (1:1000 dilution, catalog number 4970; Cell Signaling Technology, Danvers, MA, USA). The membranes were then incubated for 2 hours at room temperature with the appropriate fluorescent secondary antibodies: IRDye 800CW goat anti-rabbit IgG (1:5000 dilution, catalog number 926-32211; LI-COR Biosciences, Lincoln, NE, USA) and IRDye 680RD goat anti-mouse IgG (1:5000 dilution, catalog number 926-68070; LI-COR Biosciences, Lincoln, NE, USA). The blots were visualized using a laser imaging system, with β-actin as a loading control. The band intensities were captured, and the target protein intensity was normalized to that of β-actin by calculating its ratio.
Golgi staining and dendrite analysis
Golgi staining was performed on D 7 post-administration using an FD Rapid Golgi Stain™ kit (FD Neuro Technologies, Ellicott City, MD, USA) according to the manufacturer’s instructions. In brief, mice were deeply anesthetized and transcardially perfused with 0.9% sterile saline. Brains were rapidly removed and cut into 2- to 3-mm-thick tissue blocks containing the hippocampus and mPFC. The tissue blocks were completely immersed in a solution containing potassium dichromate, mercuric chloride, and potassium chromate and placed in a cool, dark, ventilated place for 14 days, with the staining solution replaced every 2 days. After staining, the tissues were soaked in distilled water three times, then incubated overnight in 80% acetic acid. Once the tissues became soft, they were rinsed with distilled water and cryoprotected in 30% sucrose.
The tissues were cut into 100-μm-thick coronal sections on a freezing microtome (CM1860 UV; Leica Microsystems, Wetzlar, Germany), collected on slides coated with a mixture of gelatin, chromium, and potassium sulfate, and dried overnight in the dark. After drying, the sections were rehydrated, incubated with an imaging solution, dehydrated through graded ethanol concentrations, cleared with xylene, and mounted with a resin mounting medium.
The total number of dendritic branches was quantified by Sholl analysis to assess dendritic complexity. Dendritic spine density was expressed as the number of spines per 10-μm dendrite length. To evaluate hippocampal structural plasticity, we selected well-impregnated pyramidal cells from cornu ammonis 1 (CA1), CA2, and CA3, and granule cells from the dentate gyrus (DG) with clearly distinguishable and intact dendrites. For analysis of mPFC structural plasticity, we selected well-impregnated pyramidal cells from the infralimbic cortex, prelimbic cortex, and cingulate cortex area 1. Neuronal tracing and digital reconstruction were performed using ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Immunofluorescence analysis
After the behavioral test, the mice were subjected to anesthesia with 1% sodium pentobarbital intraperitoneally, followed by transcardial perfusion with 0.9% sterile saline and a 4% paraformaldehyde solution. The brains were then fixed in cold paraformaldehyde overnight and cryoprotected in a 30% sucrose solution before sectioning. Using a freezing microtome (CM1860 UV; Leica Microsystems, Wetzlar, Germany), 20 μm-thick coronal brain sections were obtained. The sections were permeabilized with PBS containing Tween 20 (PBST) for 30 minutes and blocked with 5% normal goat serum in PBST for 1 hour at room temperature. The samples were incubated with antibodies against doublecortin (DCX, 1:800 dilution; catalog number 4604S; Cell Signaling Technology, Danvers, MA, USA) at 4°C overnight. After three rinses in PBST, the sections were incubated with secondary antibodies (DCX: Alexa Fluor 594-conjugated goat anti-rabbit IgG, 1:2000 dilution, catalog number 8889S; Cell Signaling Technology, Danvers, MA, USA) for 2 hours at room temperature. After another three rinses in PBST, the sections were mounted with Fluoroshield™ containing DAPI (Sigma‒Aldrich, St. Louis, MO, USA) and examined using an upright fluorescence microscope (BX63; Olympus, Tokyo, Japan). Images of four hippocampal DG sections at 400× magnification were captured. The number of DCX-positive cells in the hippocampal DG was quantified per section using ImageJ software. The aggregate count of DCX-positive cells across the four hippocampal DG sections (from 1.46 to 1.94 mm posterior to bregma) for each mouse was analyzed statistically across five mice per group.
Statistical analysis
The animals were randomly assigned to different treatment groups. Data are presented as mean ± standard error of the mean (SEM) and were analyzed and graphed using GraphPad Prism 9.0 (GraphPad Software Inc., San Diego, CA, USA). The normality of data distribution was assessed using the Shapiro–Wilk test, and homogeneity of variance was evaluated using Levene’s test. For comparisons among the three experimental groups, data were analyzed by one-way analysis of variance (ANOVA) followed by Dunnett’s multiple comparisons test, with the FC (+) Veh group designated as the common control for pairwise comparisons against both FC (−) Veh and FC (+) PSI. In cases where data failed to meet the assumptions of normality or homogeneity of variance, the Kruskal–Wallis test was used, followed by Dunn’s multiple comparisons test. Statistical significance was set at a two-tailed p < 0.05.
Results
Psilocybin mitigates the increased freezing behavior induced by fear conditioning in mice
The experimental procedure is depicted in Figure 1(a). On the day of fear conditioning, the percentage of freezing time during the first sound period was defined as the baseline freezing time. One-way ANOVA revealed no significant difference in baseline freezing time among the three groups (F(2, 45) = 0.536, p = 0.590; Figure 1(b)). The percentage of freezing time during the fifth sound period was defined as the post-conditioning freezing time. One-way ANOVA revealed significant differences among the three groups (F(2, 45) = 65.85, p < 0.001). Dunnett’s posthoc multiple comparisons test showed that this percentage was significantly higher in the FC (+) Veh group than in the FC (–) Veh group (p < 0.001; Figure 1(c)), indicating that the fear conditioning paradigm was successfully established. No significant difference was observed between the FC (+) Veh and FC (+) PSI groups (p = 0.36). Since psilocybin is rapidly cleared from systemic circulation within 6–8 hours (Carhart-Harris et al., 2016), an auditory cue-based fear memory test was conducted 24 hours post-administration to eliminate the influence of acute hallucinogenic effects. Considering that the activity of mice might affect freezing time, we conducted the OFT 2 hours before the fear test to evaluate the activity of the three groups of mice. One-way ANOVA revealed no significant difference in the total distance traveled among the three groups (F(2, 45) = 2.078, p = 0.137; Figure 1(d)), indicating that neither fear conditioning nor psilocybin administration affected the activity of the mice.
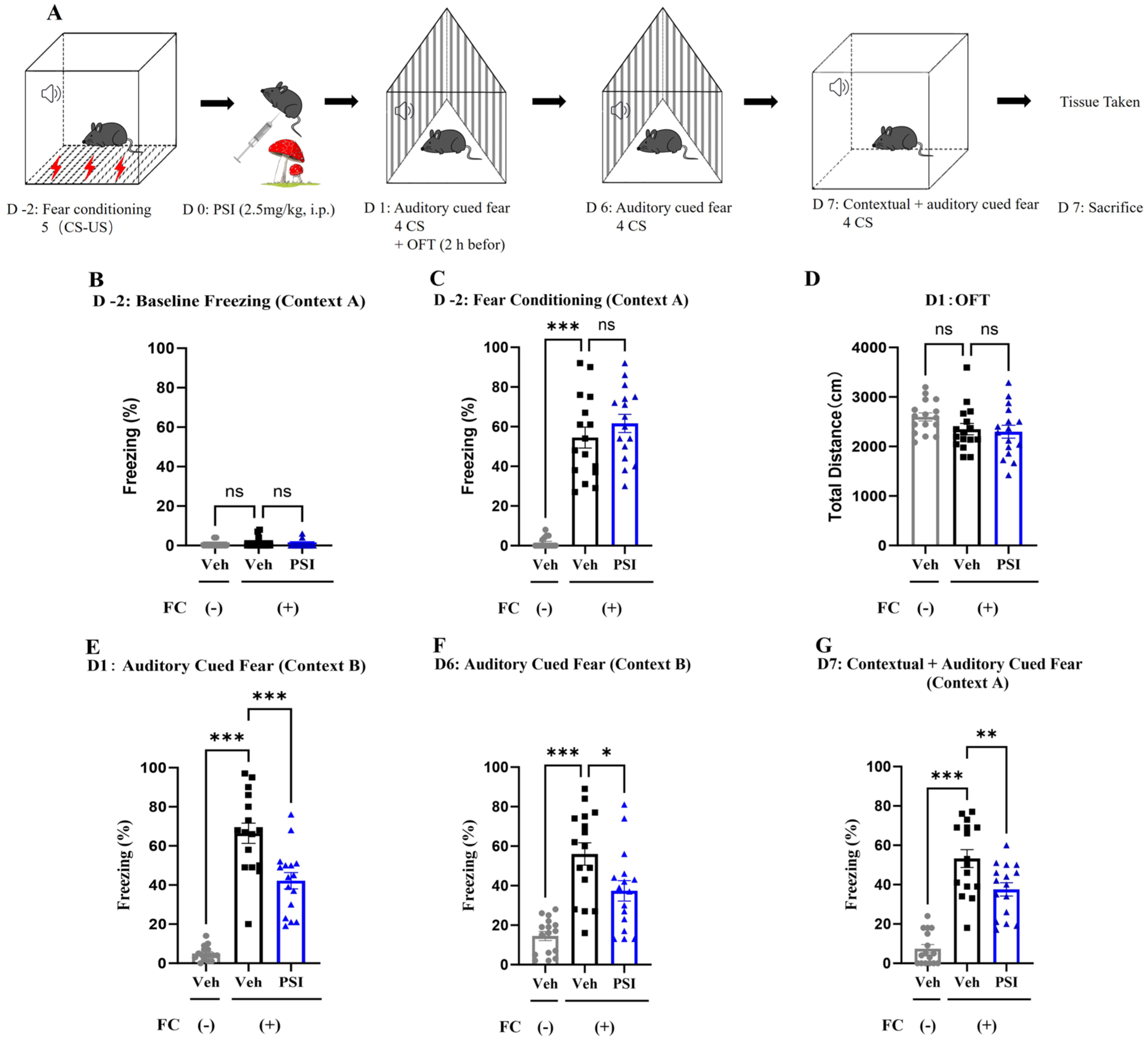
Effects of psilocybin on the percentage of freezing time in fear-conditioned mice. (a) Experimental procedure and timeline. (b) On the day of fear conditioning, no significant difference was observed in the baseline percentage of freezing time (context A) between the fear conditioning and non-fear-conditioned groups during the first sound period. (c) On the day of fear conditioning, the baseline percentage of freezing time (context A) was significantly greater in the fear-conditioned group than in the non-fear-conditioned group during the fifth sound period. (d) In the OFT, a single dose of psilocybin, administered 24 hours prior, did not affect the total distance traveled by the mice, regardless of fear conditioning. (e) A single dose of psilocybin, administered 24 hours prior, significantly reduced the elevated freezing behavior in fear-conditioned mice in context B. (f) Six days following a single psilocybin dose, mice showed a significant reduction in freezing time induced by fear conditioning in context B. (g) On the seventh day after a single dose of psilocybin, the increase in the percentage of freezing time caused by fear conditioning in context A was significantly reduced. Data are presented as mean ± SEM; n = 16 mice/group. Data were analyzed by one-way ANOVA followed by Dunnett’s multiple comparisons test, with the FC (+) Veh group designated as the common control. *p < 0.05, **p < 0.01, ***p < 0.001 versus FC (+) Veh.
The fear memory test, which retained only the auditory cue, was conducted 24 hours after a single dose of psilocybin was administered. The test took place in context B, which differed completely from context A in shape, color, and material. Notably, context A was the environment used during fear conditioning. One-way ANOVA revealed significant differences in the percentage of freezing time among the three groups (F(2, 45) = 64.21, p < 0.001). Dunnett’s posthoc multiple comparisons test showed that the percentage of freezing time was significantly greater in the FC (+) Veh group than in the FC (–) Veh group (p < 0.001; Figure 1(e)), and significantly lower in the FC (+) PSI group than in the FC (+) Veh group (p < 0.001). To investigate whether a single dose of psilocybin could produce a lasting reduction in fear memory, we assessed auditory-cued fear memory on day 6 in context B (distinct from context A in shape, color, material, and odor) and combined contextual plus auditory-cued fear on day 7 in the original conditioning context (context A). On day 6 in the novel context B, one-way ANOVA revealed significant differences among groups (F(2, 45) = 20.45, p < 0.001), with Dunnett’s test showing that freezing time was significantly higher in the FC (+) Veh group than in the FC (−) Veh group (p < 0.001) and the FC (+) PSI group (p = 0.01; Figure 1(f)), indicating that psilocybin reduced auditory-cued fear memory. In contrast, on day 7 in context A, testing of combined fear responses also showed significant group differences (F(2, 45) = 44.50, p < 0.001), with freezing significantly reduced in the FC (+) PSI group compared with FC (+) Veh (p = 0.005; Figure 1(g)). These results demonstrate that psilocybin produces a rapid and lasting reduction in both auditory-cued fear and combined contextual/cued fear memory.
Psilocybin restored structural synaptic plasticity in the hippocampus and mPFC that had been impaired by fear conditioning
On the seventh day after psilocybin treatment, tissue samples were taken for Golgi staining to assess the structural synaptic plasticity of the hippocampus and mPFC. In the hippocampus, one-way ANOVA revealed significant differences among the three groups in both the total number of dendritic intersections (F(2, 12) = 28.52, p < 0.001; Figure 2(a) and (c)) and dendritic spine density (F(2, 12) = 14.92, p = 0.001; Figure 2(b) and (d)). Dunnett’s posthoc multiple comparisons test indicated that the FC (+) Veh group exhibited significantly reduced dendritic intersections (p < 0.001) and lower spine density (p < 0.001) compared with the FC (–) Veh group. Notably, a single dose of psilocybin significantly reversed these reductions, with the FC (+) PSI group showing increased dendritic intersections (p = 0.01) and spine density (p = 0.002) relative to the FC (+) Veh group.
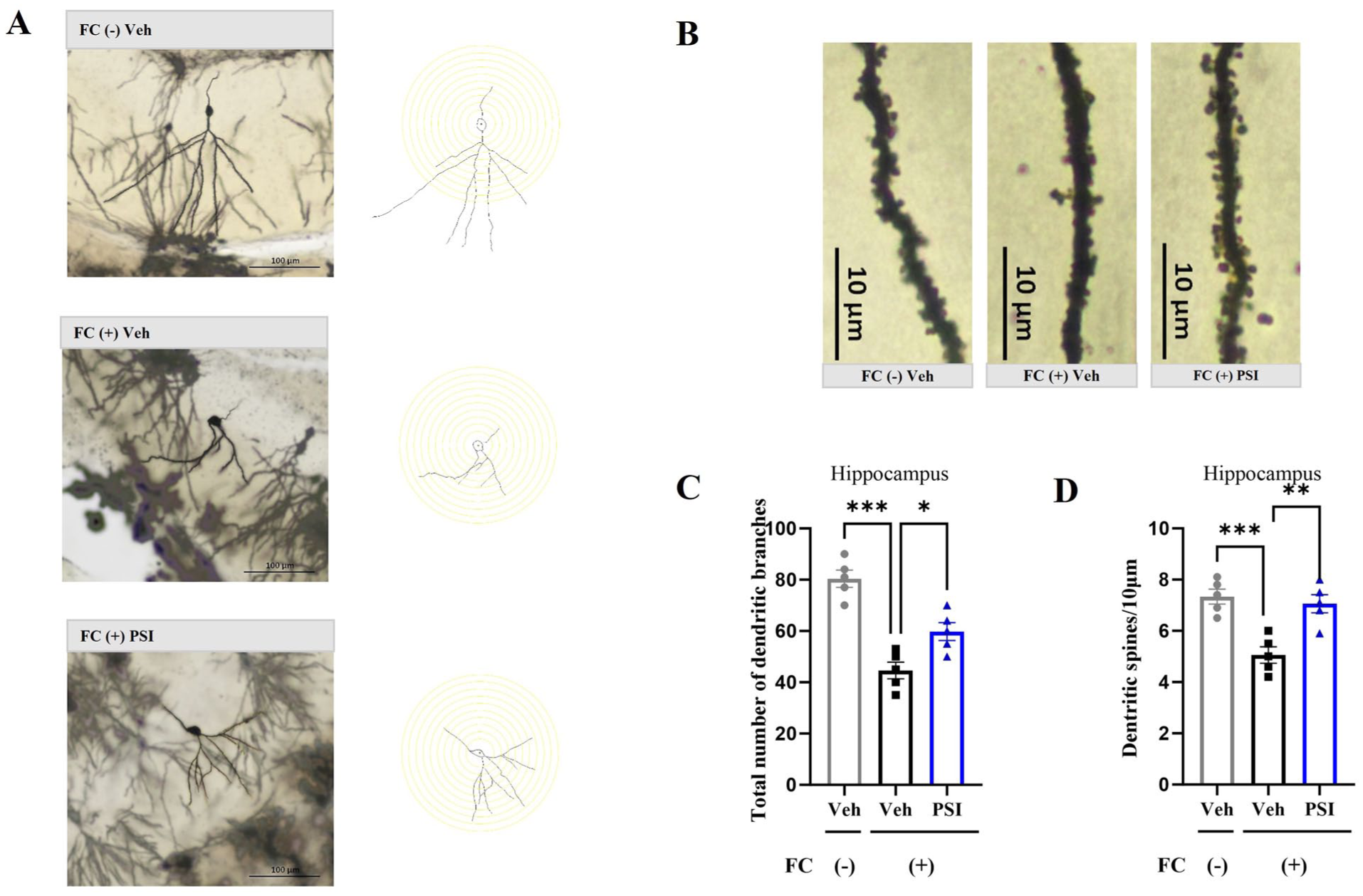
Impact of psilocybin on structural plasticity in the hippocampus. (a) Representative image showing total hippocampal dendritic branching in three groups of mice, alongside a schematic diagram of neuron tracing performed using ImageJ. Scale bar = 100 μm. (b) Representative image of dendritic spine density in the hippocampi of the three groups of mice. Scale bar = 10 μm. (c) Psilocybin significantly reversed the reduction in hippocampal dendritic branching induced by fear conditioning. (d) Psilocybin significantly reversed the reduction in hippocampal dendritic spine density induced by fear conditioning. Data are presented as mean ± SEM. n = 5 mice/group; For each mouse, one section was randomly selected from each of the four hippocampal subregions (CA1, CA2, CA3, and DG), and values from these four sections were averaged to create a single data point per subject. Data were analyzed by one-way ANOVA followed by Dunnett’s multiple comparisons test, with the FC (+) Veh group designated as the common control. *p < 0.05, **p < 0.01, ***p < 0.001 versus FC (+) Veh.
In the mPFC, significant group differences were also observed for both dendritic intersections (F(2, 12) = 8.993, p = 0.004; Figure 3(a) and (c)) and spine density (F(2, 12) = 8.925, p = 0.004; Figure 3(b) and (d)). The FC (+) Veh group displayed significantly fewer dendritic intersections (p = 0.006) and reduced spine density (p = 0.007) compared with the FC (–) Veh group. Furthermore, psilocybin administration significantly reversed these effects, as evidenced by increased dendritic intersections (p = 0.006) and spine density (p = 0.006) in the FC (+) PSI group compared with the FC (+) Veh group. These results suggest that psilocybin alleviated the reduction in structural synaptic plasticity in the hippocampus and mPFC caused by fear conditioning.
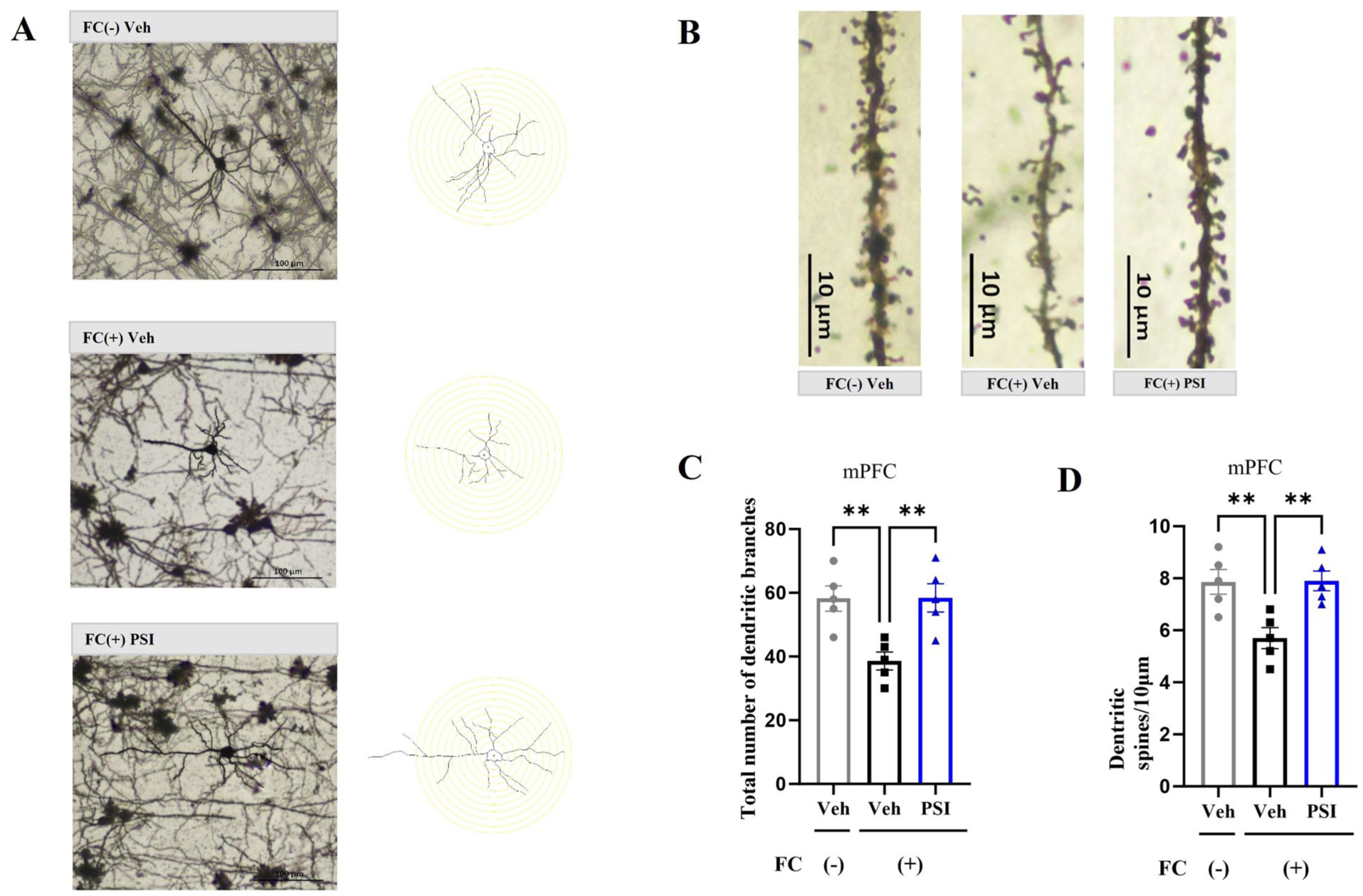
Impact of psilocybin on the structural plasticity of the mouse mPFC. (a) Representative image depicting total dendritic branching in the mPFC across three experimental groups, alongside a schematic representation of neuron tracing conducted using ImageJ software. Scale bar = 100 μm. (b) Representative image of dendritic spine density in the mPFC of the three groups of mice. Scale bar = 10 μm. (c) Psilocybin significantly ameliorated the reduction in total dendritic branching in the mPFC caused by fear conditioning. (d) Psilocybin significantly attenuated the decrease in dendritic spine density in the mPFC caused by fear conditioning. Data are presented as mean ± SEM. n = 5 mice/group; For each mouse, one section was randomly selected from each of the three mPFC subregions (IL, PrL, and Cg1), and values from these three sections were averaged to create a single data point per subject. Data were analyzed by one-way ANOVA followed by Dunnett’s multiple comparisons test, with the FC (+) Veh group designated as the common control. **p < 0.01 versus FC (+) Veh.
Psilocybin ameliorated the downregulation of neuroplasticity marker proteins in the hippocampus and mPFC induced by fear conditioning
The BDNF-mTOR signaling pathway is an important pathway involved in regulating neural plasticity (Pilar-Cuellar et al., 2019). To further investigate the neuroplasticity-modulating effects of psilocybin, we conducted Western blot analysis of the hippocampus and mPFC on day 7 post-administration, focusing on the expression of neuroplasticity markers. One-way ANOVA revealed significant differences among the three groups in BDNF expression in the hippocampus (F(2, 12) = 8.682, p = 0.005; Figure 4(a)) and mPFC (F(2, 12) = 9.742, p = 0.003; Figure 5(a)), as well as in mTOR expression in the hippocampus (F(2, 12) = 8.637, p = 0.005; Figure 4(b)) and mPFC (F(2, 12) = 16.23, p = 0.0004; Figure 5(b)). Dunnett’s posthoc multiple comparisons test indicated that the protein expression levels of BDNF (hippocampus, p = 0.002; mPFC, p = 0.002) and mTOR (hippocampus, p = 0.002; mPFC, p = 0.0003) were significantly lower in the FC (+) Veh group than in the FC (–) Veh group. Notably, a single dose of psilocybin significantly reversed these reductions, as evidenced by increased BDNF (hippocampus, p = 0.014; mPFC, p = 0.014) and mTOR (hippocampus, p = 0.0003; mPFC, p = 0.0003) expression in the FC (+) PSI group compared with the FC (+) Veh group. GluR1 (Jung et al., 2020; Kopec et al., 2007) and synapsin-1 (Zhang et al., 2021) are important proteins related to synaptic plasticity. One-way ANOVA revealed significant differences among the three groups in GluR1 expression in both the hippocampus (F(2, 12) = 31.33, p < 0.001; Figure 4(c)) and mPFC (F(2, 12) = 24.86, p < 0.001; Figure 5(c)), as well as in synapsin-1 expression in the hippocampus (F(2, 12) = 8.35, p = 0.005; Figure 4(d)) and mPFC (F(2, 12) = 42.74, p < 0.001; Figure 5(d)). Dunnett’s posthoc multiple comparisons test indicated that the protein expression levels of GluR1 (hippocampus, p = 0.002; mPFC, p < 0.001) and synapsin-1 (hippocampus, p = 0.007; mPFC, p < 0.001) were significantly lower in the FC (+) Veh group than in the FC (–) Veh group. Notably, a single dose of psilocybin significantly reversed these reductions, as evidenced by increased GluR1 (hippocampus, p < 0.001; mPFC, p < 0.001) and synapsin-1 (hippocampus, p = 0.009; mPFC, p < 0.001) expression in the FC (+) PSI group compared with the FC (+) Veh group. These results suggest that psilocybin improved the downregulation of neural plasticity marker proteins in the hippocampus and mPFC induced by fear conditioning.
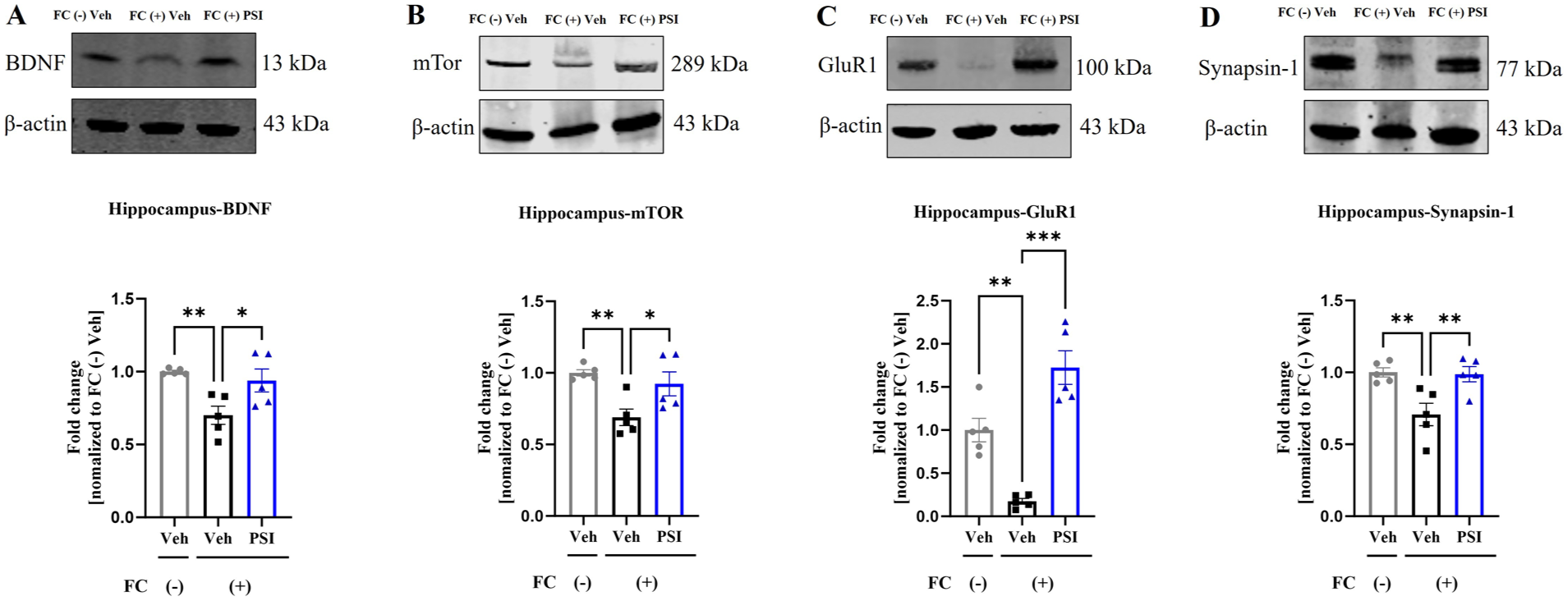
Impact of psilocybin on hippocampal neuroplasticity markers in mice. Psilocybin significantly ameliorated the fear-conditioning-induced decrease in hippocampal expression of BDNF (a), mTOR (b), GluR1 (c), and synapsin-1 (d). Data are presented as mean ± SEM; n = 5 mice/group. Data were analyzed by one-way ANOVA followed by Dunnett’s multiple comparisons test, with the FC (+) Veh group designated as the common control. *p < 0.05, **p < 0.01, ***p < 0.001 versus FC (+) Veh.
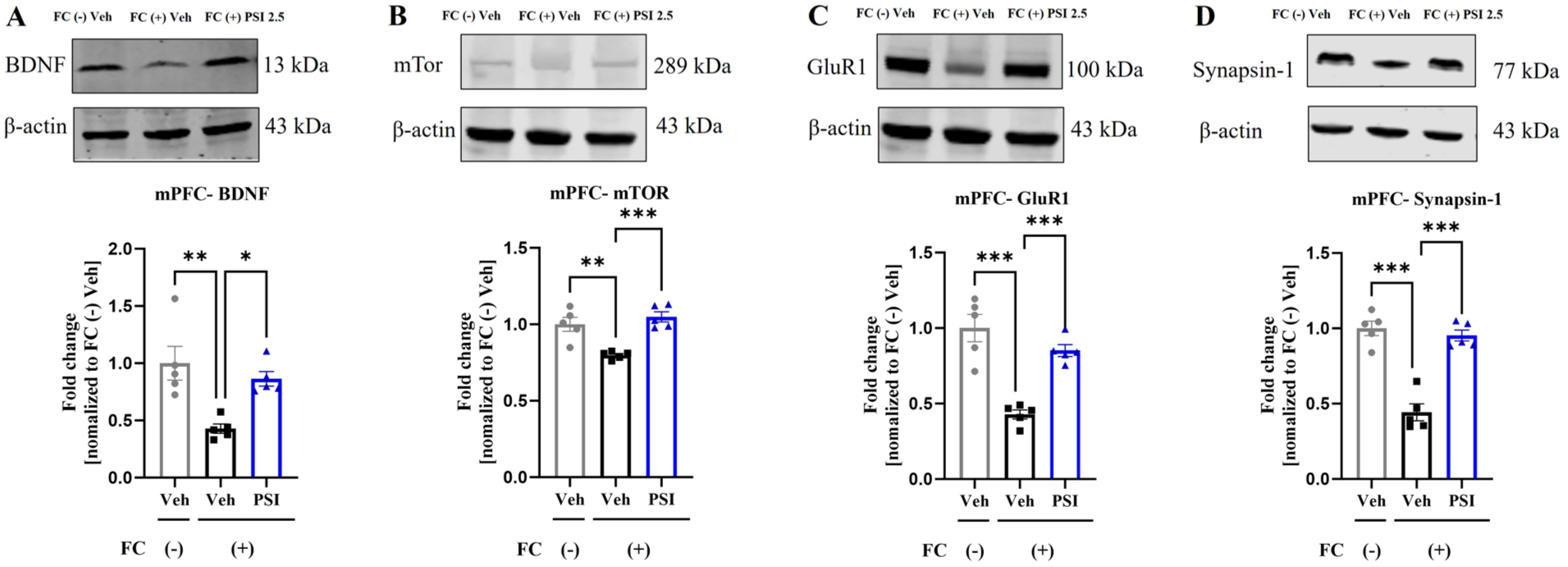
Impact of psilocybin on structural neuroplasticity markers in the mouse medial prefrontal cortex. Psilocybin significantly ameliorated the fear-conditioning-induced decrease in BDNF (a), mTOR (b), GluR1 (c), and synapsin-1 (d). Data are presented as mean ± SEM; n = 5 mice/group; Data were analyzed by one-way ANOVA followed by Dunnett’s multiple comparisons test, with the FC (+) Veh group designated as the common control. *p < 0.05, **p < 0.01, ***p < 0.001 versus FC (+) Veh.
Psilocybin ameliorated the reduction in the number of DCX-positive cells in the hippocampal DG induced by fear conditioning
DCX is a marker of neurogenesis and is associated with neuroplasticity (Maheu et al., 2022). We labeled DCX (red fluorescence)-positive cells and DAPI (blue fluorescence)-positive cells in the hippocampal DG using immunofluorescence staining. One-way ANOVA revealed significant differences among the three groups in the number of DCX-positive cells (F(2, 12) = 8.619, p = 0.005; Figure 6). Dunnett’s posthoc multiple comparisons test indicated that the FC (+) Veh group had significantly fewer DCX-positive cells than the FC (–) Veh group (p = 0.010). Notably, a single dose of psilocybin significantly restored the number of DCX-positive cells (p = 0.005 compared with FC (+) Veh), reaching near-control levels. These results suggest that psilocybin ameliorates the reduction in the number of DCX-positive cells in the hippocampal DG induced by fear conditioning.
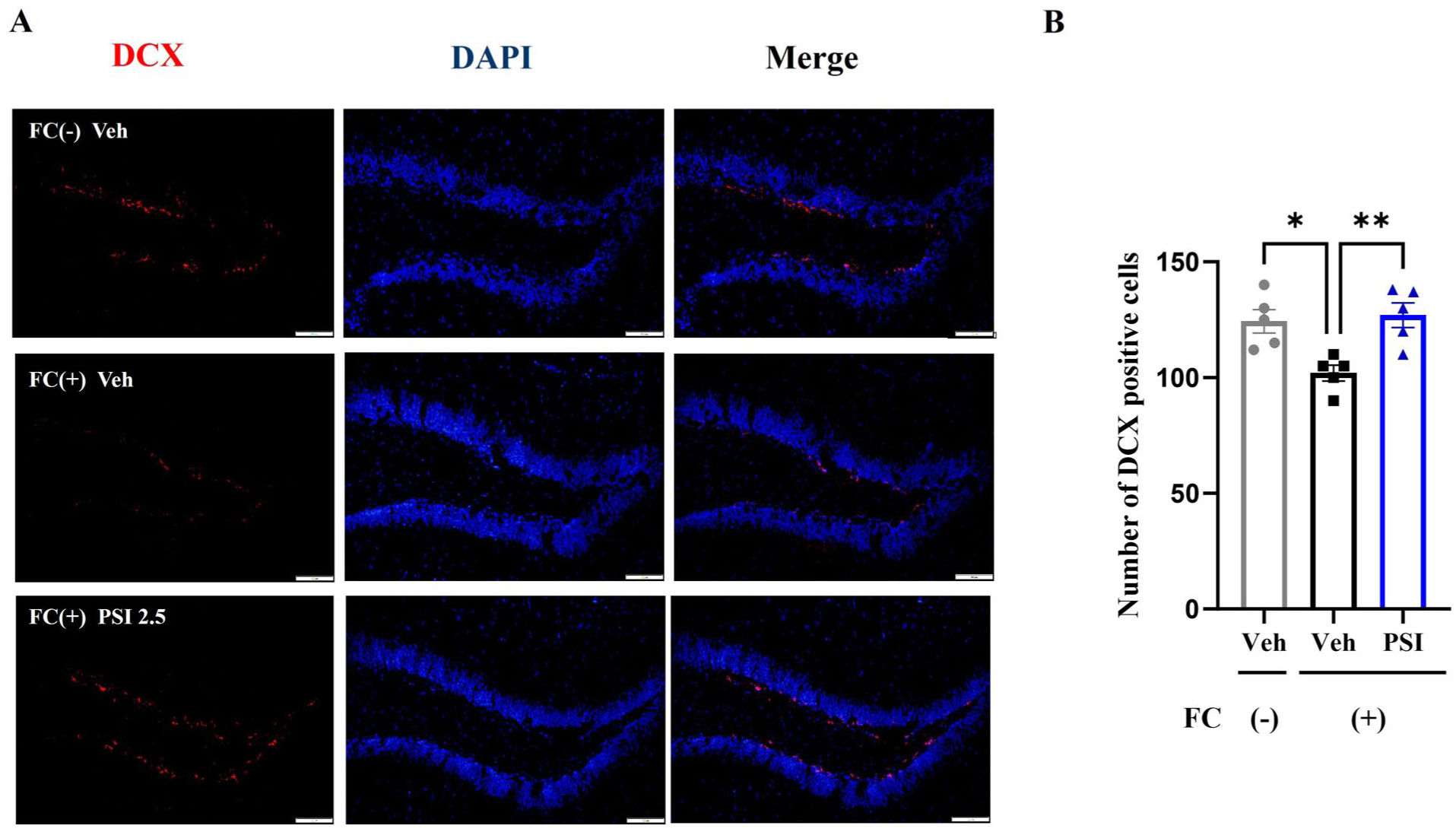
Impact of psilocybin on the number of DCX-positive cells in the mouse hippocampal DG. (a) Typical images of DCX-positive cells in the hippocampal DG of mice from the three groups (red fluorescence). Scale bar = 100 μm. (b) Psilocybin significantly ameliorated the decrease in the number of DCX-positive cells in the hippocampal DG induced by fear conditioning. Data are presented as mean ± SEM. n = 5 mice/group; five mice per group were selected, and four hippocampal brain sections per mouse were used. Data were analyzed by one-way ANOVA followed by Dunnett’s multiple comparisons test, with the FC (+) Veh group designated as the common control. *p < 0.05, **p < 0.01 versus FC (+) Veh.
Discussion
This study investigated the effects of psilocybin on short-term and long-term fear memory and further assessed its role in promoting neuroplasticity in the hippocampus and mPFC. A single administration of psilocybin rapidly and persistently reduced fear memory, with effects manifesting 24 hours after administration and lasting up to the seventh day. This action of psilocybin coincides with the enhancement of neuroplasticity in the hippocampus and mPFC, specifically by increasing structural synaptic plasticity, activating the BDNF-mTOR signaling pathway, upregulating the expression of neuroplasticity marker proteins, and promoting neurogenesis.
On the day of fear conditioning and from day 2 through day 7 post-administration, mice in the fear-conditioned group exhibited a significantly higher percentage of freezing behavior than those in the non-fear-conditioned group, confirming the successful establishment of the fear conditioning model. These results indicate that the fear conditioning model used in our study successfully associated the CS and US without additional stress. The OFT was conducted specifically to exclude locomotor confounds—ensuring that reduced freezing was not due to sedation/hypoactivity or increased freezing due to hyperactivity. The OFT results revealed that the total distance traveled by the mice was unaffected by psilocybin administration, suggesting that the observed reductions in fear memory were not due to changes in general activity caused by psilocybin. Our study demonstrated that a single dose of psilocybin significantly reduced the fear memory of fear-conditioned mice in response to auditory cues. Consistent with previous studies demonstrating psilocybin’s acute effects on fear extinction (Du et al., 2023; Catlow et al., 2013; Hagsäter et al., 2021; Woodburn et al., 2024), our findings extend this work by showing persistent behavioral effects lasting up to 7 days following a single dose. Recent work by Woodburn et al. (2024) similarly demonstrated long-term fear reduction, though the neurobiological substrates remained unexplored. Our study addresses this gap by linking persistent behavioral changes to structural neuroplasticity in the hippocampus and mPFC. Furthermore, a study published in 2021 indicated that administering psilocybin 0.5 hour after fear conditioning reduced the percentage of freezing time and contextual fear memory in mice (Hagsäter et al., 2021). Regrettably, both studies only assessed the impact of psilocybin on fear memory at a single time point, either 0.5 (Catlow et al., 2013) or 24 hours (Hagsäter et al., 2021) after administration, without evaluating the long-term effects on fear memory. In our experiments, to exclude the acute hallucinogenic effects of psilocybin, we conducted auditory-cue fear memory tests 24 hours after psilocybin treatment in context B, which was different from context A in shape, color, texture, and odor. To explore the long-term impact of psilocybin on fear memory, we performed auditory-cue fear memory tests on the sixth day (context B) and seventh day (context A, the fear conditioning context) after administration. Our behavioral tests indicated that a single dose of psilocybin can rapidly and persistently reduce fear memory, with effects manifesting 24 hours after administration and lasting up to the seventh day. This finding is consistent with the latest research findings from 2024 (Woodburn et al., 2024).
Fear conditioning impairs structural plasticity in the hippocampus and mPFC (Du et al., 2023; Lai et al., 2012; Xu et al., 2023). Accordingly, restoring neuroplasticity is an essential goal in PTSD treatment (Yabuki and Fukunaga, 2019). BDNF is a core protein involved in neuroplasticity, and its plasma levels are typically reduced in patients with PTSD (Almeida et al., 2021). Increasing BDNF levels through treatment can inhibit fear memory and exert an anti-PTSD effect. The BDNF-mTOR signaling pathway is an important pathway related to PTSD and fear memory (Notaras and van den Buuse, 2020), and the rapid antidepressant effect of psilocybin is at least partially dependent on the rapid increase in BDNF levels (Duman et al., 2021; Holze et al., 2022). Therefore, we assessed structural plasticity in the hippocampus and mPFC through Golgi staining and the expression of synaptic plasticity marker proteins (BDNF, mTOR, GluR1, and synapsin-1) in these regions using Western blotting. The present study demonstrates that psilocybin reverses these fear conditioning-induced structural deficits, restoring dendritic complexity and spine density. These findings align with prior studies highlighting psilocybin’s neuroplasticity-enhancing properties in the hippocampus (Catlow et al., 2013; Jefsen et al., 2021) and mPFC (Shao et al., 2021), suggesting that its fear memory-reducing effects are mediated by the restoration of structural synaptic plasticity in trauma-related circuits. These results suggest that the fear memory-reducing effect of psilocybin is related to the enhancement of neuroplasticity in the hippocampus and mPFC.
DCX is a microtubule-associated protein transiently expressed in immature neurons, serving as a standard marker for adult hippocampal neurogenesis essential for neuronal migration and differentiation (Klempin et al., 2011). Fear conditioning is known to suppress hippocampal neurogenesis and reduce DCX protein levels, while interventions that promote neuroplasticity restore DCX-positive cell numbers (Maheu et al., 2022). In the present study, we examined DCX to assess whether psilocybin’s effects on fear memory were accompanied by enhanced neurogenesis in the DG. Our study revealed that 7 days after a single administration of psilocybin, the decrease in the number of DCX-positive cells in the hippocampal DG was reversed, which is consistent with our previous findings (Du et al., 2023). These findings indicate that psilocybin promotes neurogenesis in the hippocampal DG, further suggesting that enhancing hippocampal neuroplasticity may be an important mechanism by which psilocybin reduces fear.
The present findings are limited to male mice, and future studies are necessary to determine whether psilocybin produces comparable effects on fear memory and structural plasticity in females, particularly across different estrous cycle phases. The present study employed a single dose of psilocybin based on pilot data; future studies incorporating full dose-response analyses are necessary to determine the minimal effective dose, optimal therapeutic window, and potential non-linear relationships between dose, plasticity markers, and fear extinction. The present study utilized aggregate freezing data across entire test sessions; future studies employing extended CS trial sequences (10–15 trials) are necessary to disentangle psilocybin’s effects on fear memory retrieval/consolidation from potential within-session extinction mechanisms.
Currently, although an increasing number of scientists speculate that psilocybin may serve as an effective treatment for PTSD (Bird et al., 2021; Chi and Gold, 2020; Khan et al., 2022; Krediet et al., 2020; Ly et al., 2018; Murnane, 2019), psilocybin remains a Schedule I-controlled substance in China and most other nations due to its hallucinogenic effects. After ingestion, the parent drug enters the bloodstream and is metabolized into the active metabolite psilocin, which reaches the nervous system and activates serotonin receptors and other receptors, causing hallucinogenic effects. Among all the hallucinogens, psilocybin has a higher safety profile, fewer side effects, and a lower risk of addiction (Lowe et al., 2021), and its use is generally not life-threatening (Carhart-Harris et al., 2016).
In conclusion, a single administration of psilocybin can rapidly and persistently reduce fear memory in fear-conditioned mice, accompanied by restored neuroplasticity in the hippocampus and mPFC. Psilocybin has significant potential for use in the treatment of PTSD and other mental disorders characterized by fear memory.
Footnotes
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the Brain Science and Brain-like Intelligence Technology-National Science and Technology Major Project (2021ZD0202000), the National Natural Science Foundation of China (No. 82270411), the Capital’s Funds for Health Improvement and Research (Nos. CFH2024-2-2058 and 2026-4-2059), and Beijing Natural Science Foundation (No. 7264380)
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.