
Abstract
Background:
Intravenous self-administration (SA) in rodents is widely regarded as the standard approach for studying the neurobiological mechanisms of drug addiction. However, its use in mice is limited by the frequent complications of intravenous catheterization, including catheter failure or occlusion.
Aims:
To overcome these challenges, we developed and validated a noninvasive vapor approach for cocaine SA in mice.
Methods:
In this study, mice were trained to self-administer cocaine vapor under fixed-ratio schedules (FR1 and FR3) and exposure to cocaine SA in both short-access (1 hour/day) and long-access (3 hours/day) sessions.
Results:
Our findings show that the vaporized cocaine SA model induces robust behavioral and physiological effects consistent with cocaine use disorder. Our results demonstrated increased operant responding under higher reinforcement schedules, and the escalation of vaporized cocaine intake during extended access sessions (3 hours/day) emulates features of compulsive drug use in humans.
Conclusions:
Our findings show that the vaporized cocaine SA model offers a translational model to advance preclinical research and develop effective treatments. This vapor-based approach circumvents the need for catheterization, reduces animal stress, and is suitable for longitudinal studies and compatible for use with various modern neurogenetic techniques that require a tether for imaging free-moving, behaving animals (e.g., optogenetics and brain imaging).
Introduction
Substance use disorders represent a major global health challenge. Among them, cocaine use disorder (CUD) is particularly concerning, as it is associated with severe medical, psychiatric, and socioeconomic burdens; yet no FDA-approved pharmacotherapies are currently available (Volkow and Boyle, 2018; Zhu et al., 2025). Thus, advancing our understanding of the neurobiological mechanisms underlying cocaine reinforcement, its escalated use, and relapse after cocaine discontinuation is essential for the development of effective treatments.
Animal models have played a critical role in advancing addiction research. Self-administration (SA) is widely recognized as the “gold standard” for studying the neurobiological mechanisms of addictive drugs in laboratory settings (Panlilio and Goldberg, 2007). Compared to other passive administration models, SA allows animals to actively control their drug intake, thus providing the most direct point-to-point correspondence with addiction-like behaviors as they occur in humans. Moreover, SA models demonstrate strong construct validity by reproducing core behavioral features of addiction, including escalation of drug intake and reinstatement of drug-seeking after abstinence (Ahmed and Koob, 1998; Epstein et al., 2006).
Intravenous cocaine SA in rodents has a long history. Weeks (1962) demonstrated that rats lever-pressed to obtain intravenous cocaine, providing evidence that animals could voluntarily self-administer psychostimulants. Subsequent studies confirmed that cocaine had robust reinforcing effects, such as compulsive-like drug seeking and high reinstatement propensity, which revealed behavioral features closely resembling drug seeking in humans with addiction. With advances in microsurgical techniques, stable cocaine SA procedures were later established in mice in the late 1990s by Sora et al (1998) and refined by Thomsen and Caine (2005). The successful adaptation of SA to mice has become a powerful tool as it can be combined with a wide range of transgenic mouse strains that permit genetic targeting and manipulation using state-of-the-art approaches, including neuroimaging, chemogenetics, and optogenetics (Towers et al., 2019; Zhang et al., 2018).
However, traditional SA procedures in mice (Chistyakov and Tsibulsky, 2006; Kmiotek et al., 2012) require surgical implantation of an intravenous catheter, which is invasive and technically difficult. Maintaining long-term catheter patency is particularly challenging due to their small body size and fragile veins, thereby restricting the duration of cocaine exposure, especially under extended access conditions (Arena et al., 2019; Zhang et al., 2018). In addition, recent advances in non-tethered neuroscience techniques, including wireless photometrical and optogenetic tools, have improved the feasibility of studying cocaine SA in freely moving animals. However, vapor-based SA models remain a useful complementary approach because they avoid the burden of tethering, surgery, and repeated handling stress. Furthermore, although intravenous SA is a widely used preclinical model, it does not fully replicate inhaled or smoked cocaine use in humans, because different routes of administration produce distinct pharmacokinetic profiles (Cone, 1995; Jeffcoat et al., 1989).
To overcome these limitations, foundational nonhuman primate studies demonstrated that vaporized cocaine base reliably supports SA, producing dose-dependent intake, reinforcement, and pharmacokinetic profiles that closely mirror human smoked cocaine use (Carroll et al., 1990; Comer et al., 1994, 1995) and provided a critical translational rationale for inhalational models. Researchers recently developed a vapor-based SA model in rodents that is noninvasive and allows for repeated testing without the need for surgical implantation of catheters for drug deliveries. The vapor SA model has been accomplished in rats for alcohol (de Guglielmo et al., 2017; Vendruscolo and Roberts, 2014), nicotine (Smith et al., 2020), cannabis (Freels et al., 2020; Muthusamy, 2020), and opioids (Marchette and Vendruscolo, 2025), including sufentanil (Vendruscolo et al., 2018), fentanyl (Marchette et al., 2023; McConnell et al., 2021), and heroin (Gutierrez et al., 2021). More importantly, a mouse model of opioid vapor SA using fentanyl was recently reported (Moussawi et al., 2020). Based on our literature search, however, no vapor-based cocaine SA model has been reported in either rats or mice.
In the present study, we report on the development and validation of a noninvasive mouse model of vaporized SA of cocaine that provides a viable alternative to current intravenous approaches. We first established a suitable vaporized cocaine dosage based on locomotor activity and a blood test. Then, we systematically evaluated its behavioral properties, including acquisition, maintenance, and escalation under different fixed reinforcement schedules and both short- and long-access cocaine exposure. We suggest that this noninvasive mouse model of vaporized cocaine SA will be a significant advance for the preclinical field and will facilitate longitudinal within-subjects studies of the neurobiological basis of cocaine addiction.
Methods
Animals
Male and female adult C57BL/6 mice (>10 weeks old) were obtained from The Jackson Laboratory (Bar Harbor, ME, USA). The studies were not designed to directly investigate sex differences, but mice of both sexes were included. Mice were individually housed with free access to food and water. Prior to the start of the experiments, mice were acclimated to the experimental room for at least 1 week and to the vapor chambers for 3 days. Studies were approved by the Institutional Animal Care and Use Committee of Stony Brook University. All experimental procedures were conducted in accordance with the National Institutes of Health (NIH) Guidelines for Care and Use of Laboratory Animals.
Apparatus
Operant vapor SA was carried out in two airtight chambers (14 × 23 × 23 cm; La Jolla Alcohol Research, La Jolla, CA USA; Figure 1) that were placed inside a black Plexiglas enclosure to minimize noise and ambient light. Two holes were mounted side by side on the end wall, and yellow light bulbs were mounted inside the nosepoke holes. A vacuum pump maintained constant airflow in the chambers by creating negative air pressure. The outflowing air was filtered by an in-line HEPA-Cap disposable filter and then disposed of through the facility’s exhaust system. To vaporize cocaine, we used two sets of five interconnected vaporizing tanks that were equipped with an atomizer (Extreme 2.0 Wax Vape Cartridge, HoneyStick, FL, USA), which was filled with free base cocaine. The atomizer was activated by an SVS250 vaporizer (Scientific Vapor, OR, USA). We used Med Associates software and interface (St. Albans, VT, USA) to record nosepokes and control activation of the vaporizers and light cue presentation. The suction system allowed the flow of vaporized drug into the operant chamber when the vaporizer was activated. Drug vapor delivery in the chamber is dependent on drug dose, airflow rate, atomizer output power, and vaporizing time (Marchette et al., 2021). In our experiments, we used an airflow rate of 10 L/minute, atomizer output power of 60 watts, and vaporizing time of 20 seconds. Chamber aerodynamics was calibrated to ensure balanced input and output airflow rates and sufficient O2 level (>20.7%). These parameters allowed drug clearance, defined as the absence of visible vapor in the chamber, within 1 minute after each vapor delivery.
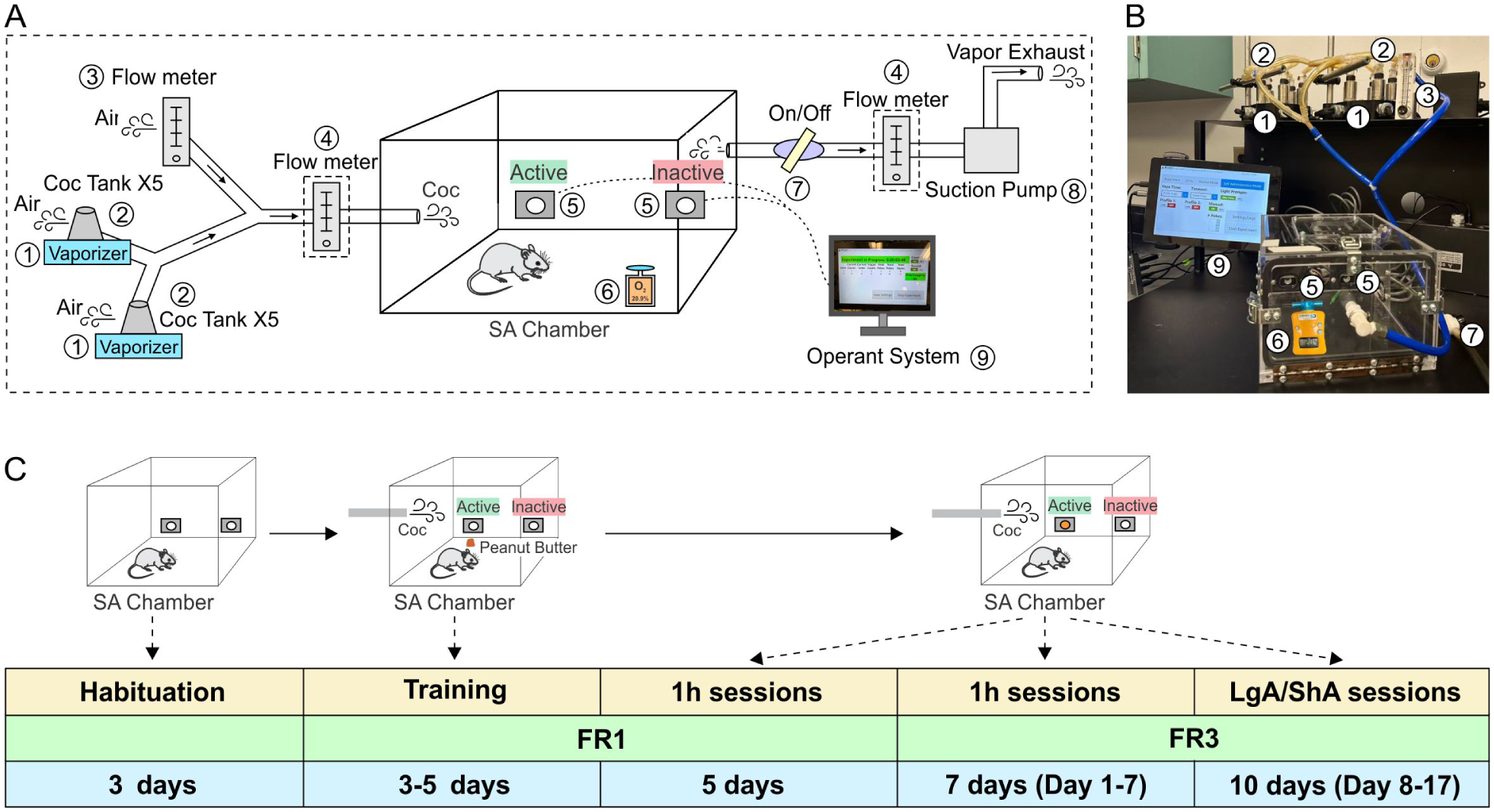
Vapor chamber setup and experimental timeline. (a) Schematic diagram of the vapor chamber set-up. (1) Vaporizer. (2) Two sets of drug loading tanks (5 per set) containing free-base cocaine powder. (3) Flow meter for air regulation. (4) Flow meter for air calibration. (5) Side-by-side nosepoke ports for animal SA. (6) Oxygenation meter for monitoring oxygen level in the chamber. (7) Airflow control switch. (8) Exhaust port connected to a vacuum suction pump. (9) Operating system interface. (4) is used for air flow calibration before the experiments to ensure the balance of the in and out air flow. Inner chamber dimensions are labeled in centimeters. The exhaust system maintains a negative pressure of approximately 10 L/minute. (b) Key components of the vapor chamber apparatus. Component numbers correspond to those in (a). (c) Timeline for training and SA experiment. Mice first underwent 3 days of habituation to the SA chambers, followed by 3–5 days of training with peanut butter to facilitate nosepoke acquisition. Mice first underwent five daily 1-hour sessions under a fixed ratio 1 (FR1) schedule and were then switched to FR3 for seven daily 1-hour sessions (days 1–7). After that, mice were exposed to long-access (LgA; 3 hours) and short-access (ShA; 1 hour) sessions under FR3 conditions (days 8–17), with body weight monitored throughout. The active port triggered cocaine vapor delivery, while the inactive port had no programmed consequences.
Drugs
Free base cocaine in powder form (NIDA Drug Supply Program, Cat# 9041-012) was used to fill the vaporizer tanks. We manually loaded cocaine into the tanks prior to the SA sessions to ensure that sufficient cocaine would be released when the vaporizer was activated. The vehicle vapor (Veh) used in this study contained 60% vegetable glycerin and 40% propylene glycol.
Locomotion test
Different cohorts of mice (n = 24) were used to measure the locomotor effects of different concentrations of cocaine vapor (10, 20, and 40 mg). Mice were habituated in locomotor boxes (22 × 43 × 23 cm) 1 hour/day for 3 days. On the fourth day, mice were placed in the locomotor activity boxes for 10 minutes and then placed in the vapor chambers where they were passively exposed to different dosages of cocaine vapor for a total of five separate vapor deliveries. Each cocaine vapor delivery time was 20 seconds followed by a 3-minute timeout period. A total of five vapor deliveries were evenly spaced and completed within 20 minutes (the last delivery at 20 minutes). Mice were then placed in the locomotor activity boxes for 20 minutes to measure cocaine-induced locomotion patterns.
Locomotion analysis
We employed an artificial intelligence-based approach to conduct locomotion analysis in mice. We utilized the vision foundation model SAM2 (Ravi et al., 2024) to perform precise tracking of the mouse in video recordings, from which we extracted its motion trajectory and subsequently analyzed its movement patterns. The locomotion analysis process consisted of three main steps. First, for each locomotion video, we manually initialized the mouse’s position by annotating a bounding box. Second, the video frames together with the initial bounding box were provided to the SAM2 model to perform continuous tracking of the mouse throughout the sequence. The model generated a mask-based trajectory, which was subsequently converted to a series of bounding boxes. The centroid of each bounding box was taken as the mouse’s position at the corresponding time point. Third, the complete trajectory was used to compute and analyze the mouse’s motion patterns. The real-world displacement and movement metrics were calculated by mapping pixel distances to physical distances, based on the known relationship between the video resolution and the actual object size.
Blood cocaine levels
Different cohorts of mice (n = 15) were passively exposed to different concentrations of cocaine vapor (20 or 40 mg) for a total of five vapor deliveries separately over 20 minutes (i.e., using the same protocol as the locomotor test with a 20-second vapor delivery and 3-minute timeout period) to measure blood cocaine levels. After the last vapor delivery, mice were euthanized by CO2 inhalation; 1 ml of blood was collected by cardiac puncture, placed in vacutainer ethylenediaminetetraacetic acid tubes, and stored at −4°C. Plasma cocaine levels were analyzed by high-performance liquid chromatography/tandem mass spectrometry (NMS Labs, WillowGrove, PA, USA).
Body weight measurements
During the long-access (LgA) cocaine SA experiment, the body weight of each mouse was measured and recorded every 2 days.
Self-administration
One nosepoke hole was active (i.e., resulting in vapor delivery and/or light cue presentation), and the other nosepoke hole was inactive (i.e., no scheduled consequences). An active nosepoke resulted in a 20-second vapor delivery followed by a 3-minute timeout period. The cue light was turned on at the onset of vapor delivery and remained on throughout the timeout period, for a total cue duration of 3 minutes. The vapor was cleared within 1 minute after delivery. All nosepokes were recorded throughout the entire session. The SA session durations were 1 or 3 hours, and these sessions were on a fixed-ratio 3 (FR3) schedule of reinforcement (i.e., every three operant responses on the active operandum resulted in vapor delivery). Mice underwent one SA session per day. The amount of vaporized cocaine powder was 20 mg.
Escalation of cocaine vapor SA
Mice (n = 12) were trained to self-administer cocaine vapor (20 mg per drug vapor delivery) in five 1-hour sessions on an FR1 schedule. They were then switched to seven 1-hour sessions on an FR3 schedule. This was followed by a LgA (3 hours on an FR3 schedule) and a short-access (ShA; 1 hour on an FR3 schedule) SA session every day for 10 sessions. No food or water was provided inside the operant chambers.
Statistics
All data are expressed as the mean and standard error of the mean (SEM). Repeated measure two-way analysis of variance (ANOVA) was used for comparisons between four groups of time-dependent locomotion. One-way ANOVA was used for comparisons between four groups of locomotion parameters and three groups of blood cocaine level. All ANOVA tests were followed by the Holm–Sidak post hoc test. t-tests were used for comparison between two different groups (e.g., FR1 vs. FR3). Pearson correlations were used to compare locomotor behavior and blood cocaine concentration. Linear regression models were applied to analyze escalation data. All statistical tests were performed using SigmaStat software (Systat Software Inc., San Jose, CA), with p < 0.05 considered significant. The following criteria for p-values were used: *p < 0.05, **p < 0.01, and ***p < 0.001.
Results
Vaporized cocaine-induced locomotion
To determine the optimal dosage of vaporized cocaine for mouse SA, the effect of vaporized cocaine at various dosages (10, 20, and 40 mg) and Veh on locomotor activity was investigated (Figure 2). Figure 2(a) illustrates the paradigm of recording locomotor activity. Figure 2(b) shows the representative mouse locomotor activities before (baseline) and after exposure to various doses of vaporized cocaine (Supplemental Movie S1 and Movie S2). Figure 2(c) shows the distance travelled by mice every 5 minutes before and after passive exposure to different doses of cocaine, which revealed a dose-dependent hyperactivity. Specifically, the distance travelled for mice exposed to 40-mg vaporized cocaine (n = 6) increased to 19.36 ± 1.86 m within t = 0–5 minutes and then declined to 10.50 ± 1.26 m at t = 15–20 minutes; for the 20 mg group (n = 9) and the 10 mg group (n = 6), it increased to 14.58 ± 1.78 m and to 11.69 ± 0.96 m and then declined to 8.44 ± 9.36 m and 8.24 ± 1.62 m, respectively. In the Veh group (n = 4), the changes were marginal from a minor increase to 7.34 ± 2.34 m and then a slight decline to 5.37 ± 2.47 m, which was similar to its baseline value of 6.51 ± 0.93 m. Statistical analyses showed a significant difference in locomotion between different doses (dose × time interval, F9,60 = 2.854; p < 0.05). Figure 2(d) compares the locomotor response defined as the average locomotor distance over 20 minutes after different doses of vaporized cocaine exposure (e.g., 11.06 ± 0.32 m for 40 mg, 7.57 ± 1.23 m for 20 mg, 6.43 ± 1.12 m for 10 mg, and 4.15 ± 1.49 m for Veh). Statistical analyses indicated that mice after vaporized cocaine exposure all showed significantly elevated locomotor activity over the Veh (F3,16 = 11.74), among which the 40-mg group (p < 0.001) exhibited a higher elevation than the 20 mg (p = 0.021) and 10 mg (p = 0.008) groups. These results indicate that the level of locomotor behavior increased with the cocaine dosage. Figure 2(e) compares the initial speed within the first 5 minutes after cocaine exposure, which were 7.81 ± 0.61 cm/s, 4.63 ± 0.304 cm/s, 3.90 ± 0.32 cm/s, and 1.55 ± 0.77 cm/s for the 40, 20, and 10 mg, and Veh groups, respectively. The results showed dose-dependent significant increases in locomotor speed except those between the 10 and 20 mg groups. The 40-mg group speed was higher than 20 mg (p = 0.002), 10 mg (p = 0.011), and Veh group (F3,16 = 25.23, p < 0.001). Figure 2(f) summarizes the mobility versus immobility durations (percentage) within a total of 20 minutes, showing a significant increase in mobility durations for different doses compared with Veh (F3,16 = 14.94): 18.32% ± 6.89% (Veh), 38.28% ± 4.92% (10 mg, p = 0.037), 43.12% ± 2.67% (20 mg, p = 0.004), and 59.33% ± 3.18% (40 mg, p < 0.001).
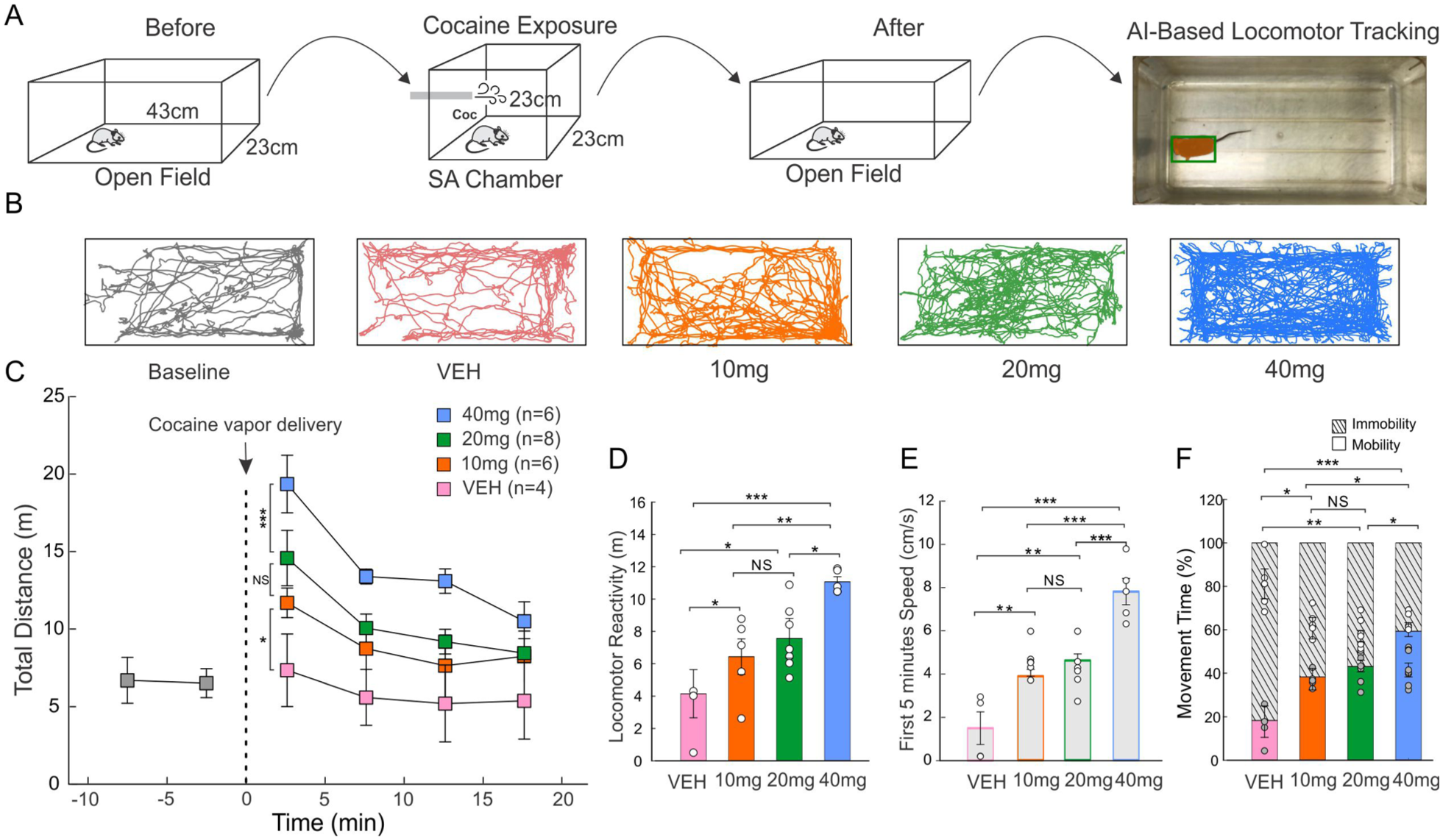
Determination of optimal vaporized cocaine dosage behavioral locomotor activity test. (a) The schematic of the behavioral test setup used to evaluate vaporized cocaine-induced locomotion. (b) Representative tracks and graphs with total distance travelled after exposure to different dosages (10, 20, and 40 mg) of vaporized cocaine versus vehicle vapor (Veh). (c) Time-course analysis of distance traveled (m) following five cocaine vapor deliveries with different dosages over 20 minutes: 40 mg (blue), 20 mg (green), 10 mg (orange), and Veh (pink), respectively. (d) Bar diagram comparing the locomotor reactivity (average distance travelled within 20 minutes) between different cocaine dosages. (e) Bar diagram comparing the average speed of the first 5 minutes for cocaine dosages. (f) Bar diagram comparing the mobility and immobility time percentage within the total 20 minutes among different cocaine dosages; grid lines represent stationary periods.
Dose-dependent blood cocaine levels
Drug-naïve mice were passively exposed to various doses of five vapor cocaine deliveries over 20 minutes (Figure 3[a]), after which their blood was collected for the measurement of blood cocaine levels. Figure 3(b) compares blood cocaine levels among different vaporized cocaine dosages and the Veh vapor. The measured blood cocaine level in the 40 mg group (55.33 ± 7.67 ng/mL) was significantly higher than in the 20 mg group (28 ± 3.17 ng/mL; p = 0.01, n = 6), both of which were significantly higher than in the Veh group (0 ng/mL, F2,12 = 18.59, p < 0.001 for 40 mg, p = 0.03 for 20 mg, n = 3). Figure 3(c) shows a significant correlation between blood cocaine level and initial locomotor activity in Figure 2(c) (r = 0.974, p < 0.001, n = 6).
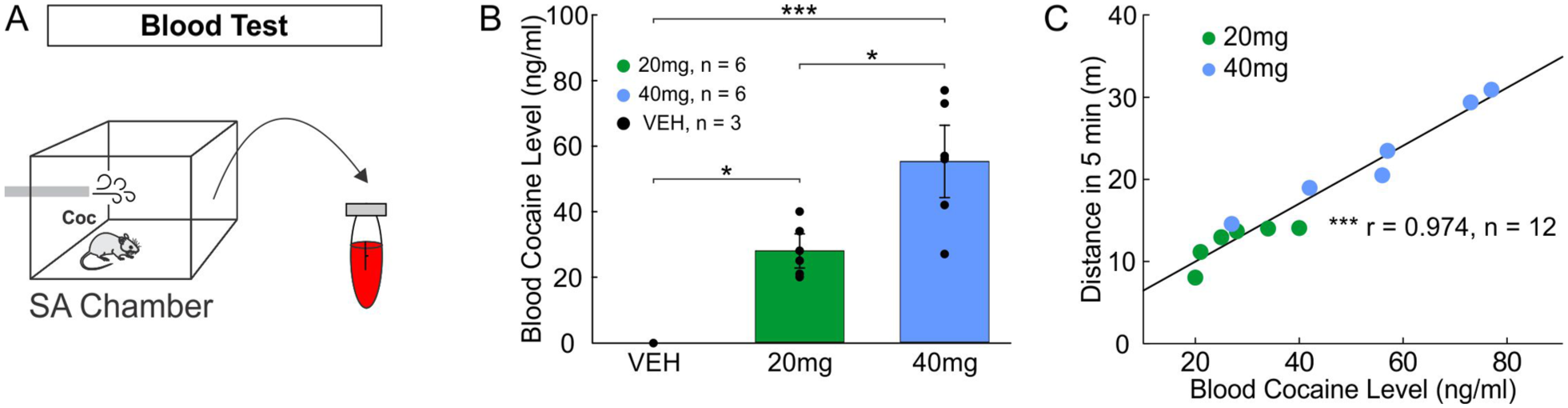
Blood cocaine concentration. (a) Blood samples were collected via cardiac puncture immediately after five passive cocaine vapor deliveries. (b) Bar diagram comparing blood cocaine concentration after five cocaine vapor deliveries at cocaine dosages of 20 mg (n = 6) and 40 mg (n = 6) versus vehicle (Veh) vapor (n = 3). (c) Correlation between blood cocaine level (ng/mL) and distances (m) in 5 minutes after five passive cocaine vapor deliveries (n = 6/each).
Vaporizing cocaine SA under different schedules of reinforcement
To test whether the vaporizing setup and the training paradigm provided sufficient motivation to initiate vaporized cocaine SA, mice underwent the SA of cocaine inhalations under two different schedules of reinforcement—FR1 and FR3 sessions (1 hour) over 5 days. Based on the results from the locomotor activity study (Figure 2) and blood cocaine levels (Figure 3), the 20 mg cocaine dose was selected for protocol development. The 20 mg dose was used, instead of 40 mg, for practical and economic reasons. This amount required less cocaine and generated less waste while still producing robust pharmacological effects. Figure 4(a) and (b) compare the active (solid circle) and inactive (open circle) nosepokes and vapor deliveries (solid triangle) during 1-hour SA sessions under FR1 (n = 9) and FR3 (n = 9) reinforcement schedules, respectively. The results indicate an increase in active nosepokes from FR1 to FR3, whereas inactive nosepokes and vapor deliveries remained unchanged. Quantitative analyses in Figure 4(c) confirmed that active nosepokes increased significantly from 5.74 ± 0.74 for FR1 to 17.70 ± 1.95 for FR3 (p* < 0.001, n = 9), whereas there were no changes in either inactive nosepokes (5.29 ± 0.53 for FR1 vs. 6.84 ± 0.44 for FR3; p = 0.12, n = 9) or vapor deliveries (5.74 ± 0.74 for FR1 vs. 5.85 ± 0.65 for FR3; p = 0.916, n = 9).
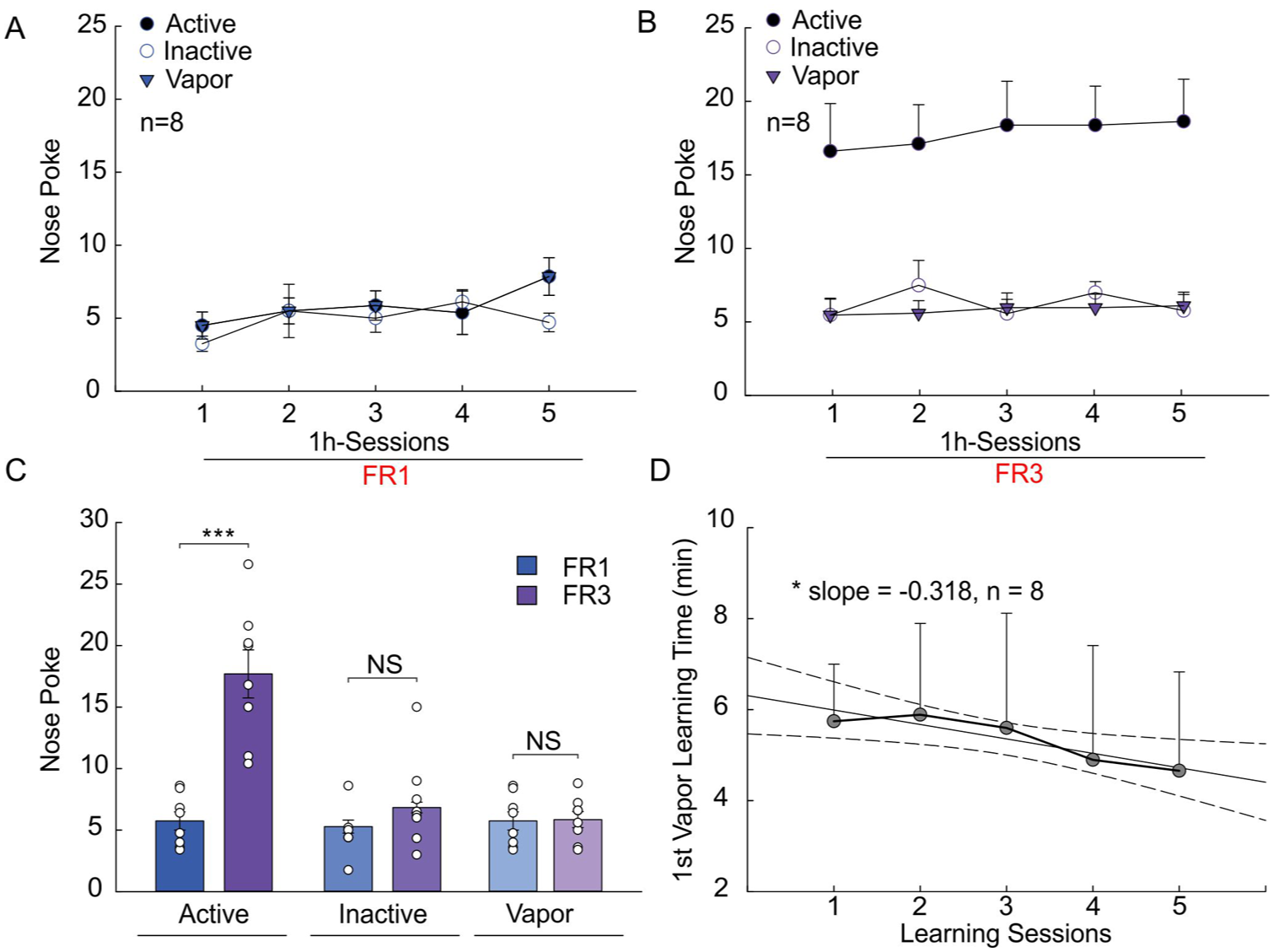
Mouse cocaine vapor self-administration (SA) responses to different reinforcement schedules. (a) Active and inactive nosepokes and cocaine vapor deliveries during 1 hour SA sessions under reinforcement schedule fixed ratio 1 (FR1). (b) Active and inactive nosepokes and cocaine vapor deliveries during 1-hour SA sessions under reinforcement schedule FR3. (c) Average active and inactive nosepokes and cocaine vapor deliveries when the reinforcement schedule was shifted from FR1 to FR3 (n = 8). (d) Linear regression showing a decrease of the time duration for mice to perform first nosepoke during 1-hour SA sessions under reinforcement schedule FR1 (slope = −0.318, CI = [−0.571, −0.064], r2 = 0.841, p* = 0.028, n = 8).
To further evaluate the rate at which mice learned to self-administer cocaine, the interval to initiate the first nosepoke during 1-hour FR1 SA sessions was measured and compared across the 5 days. A linear regression analysis revealed a significant decrease in the 5-day learning sessions (slope = −0.318, confidence interval [CI] = [−0.571, −0.064], r2 = 0.841, p = 0.028, n = 9) (Figure 4[d]).
Escalation of cocaine vapor SA
After demonstrating the efficacy of the vaporizing setup and the training schedules for cocaine SA in mice, we then refined the experimental protocol to ensure reliable escalation of cocaine intake with extended access exposure (Supplemental Movie S3). To do so, the mice after initial 7-day 1-hour FR3 training sessions were divided into two groups: (1) LgA group that self-administered cocaine vapors for 3 hours under FR3 over 10-day sessions (2) ShA group that self-administered cocaine vapors for 1 hour under FR3 over 10-day sessions. Moreover, we included a Veh vapor group to directly test whether active nosepoke responding was maintained by cocaine vapor delivery or vapor without cocaine. As shown in Figure 5(a), active nosepokes in the LgA group escalated (slope = 1.837, CI = [0.820, 2.853], r2 = 0.684, p = 0.003, n = 9) but those in the ShA group (slope = −0.258, CI = [−0.910, 0.394], r2 = 0.094, p = 0.389, n = 4) as well as Veh group (slope = −0.145, CI = [−0.521, 0.230], r2 = 0.091, p = 0.398, n = 4) did not. The significant difference in regression slope was observed from LgA versus ShA groups (F3,16 = 15.99, p = 0.001), as well as LgA versus Veh groups (p < 0.001). Similarly, cocaine vapor deliveries in the LgA group (Figure 5[b]) indicated an escalation over the 10 days of cocaine SA (slope = 0.579, CI = [0.266, 0.892], r2 = 0.695, p = 0.003, n = 9) but not in the Veh groups (slope = −0.079, CI = [−0.233, 0.076], r2 = 0.147, p = 0.273, n = 4), with a significant difference in regression slope (p < 0.001). Concomitantly, active nosepokes (slope = 0.644, CI = [0.272, 1.016], r2 = 0.666, p = 0.004) and cocaine vapor deliveries (slope = 0.185, CI = [0.051, 0.319], r2 = 0.558, p = 0.013) within the first hour in the LgA group escalated (Figure 5[c]). Active nosepokes during the timeout period in the LgA group also escalated over 10 days (slope = 17.092, CI = [10.633, 23.552], r2 = 0.823, p < 0.001, n = 9). In contrast, the timeout responding in the inactive nosepoke hole (slope = 0.801, CI = [−0.426, 2.027], r2 = 0.221, p = 0.171, n = 8) and Veh group (Veh: slope = −1.709, CI = [−4.806, 1.388], r2 = 0.168, p = 0.239, n = 4) remained stable across sessions.
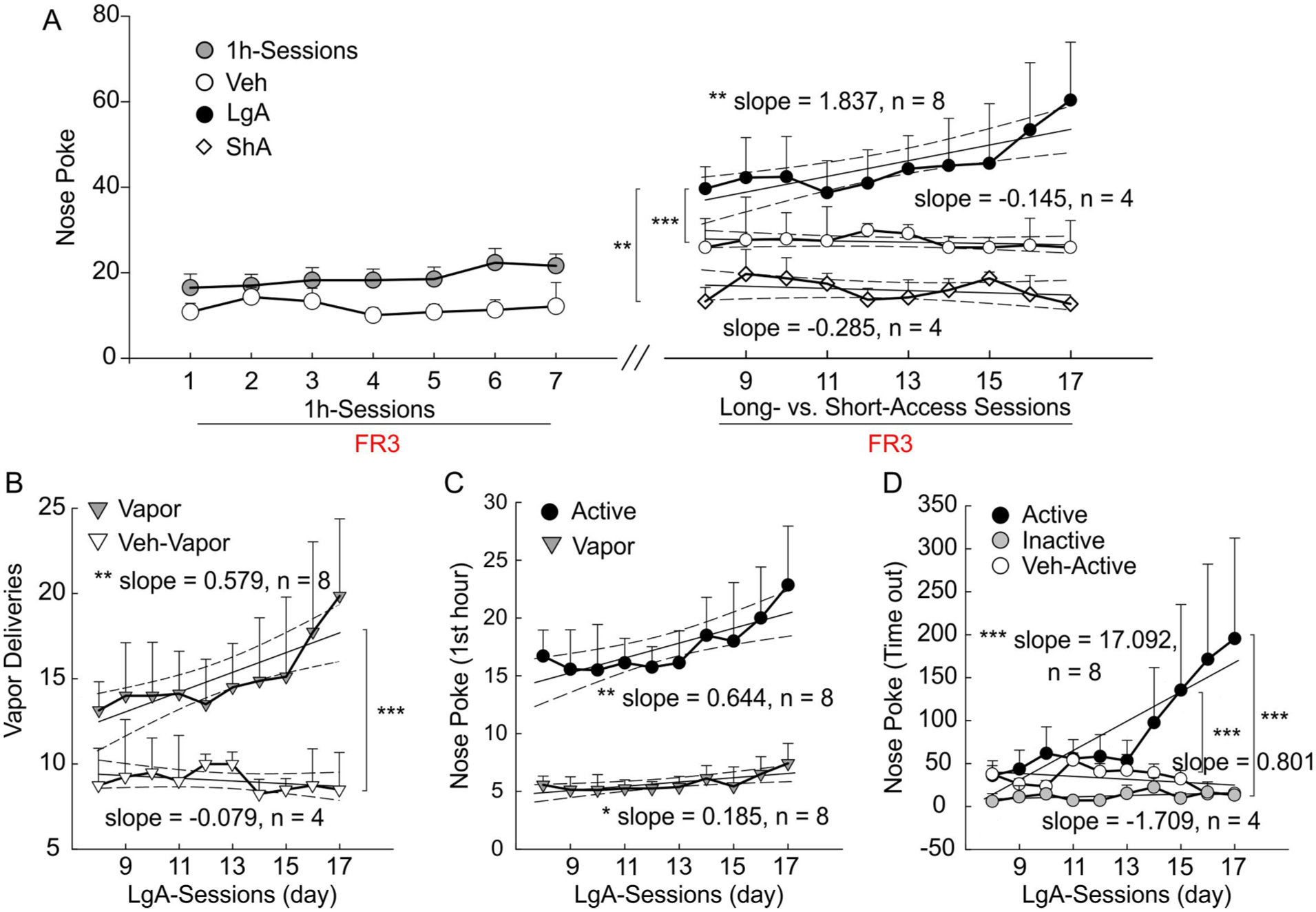
Escalation of cocaine vapor self-administration (SA) in mice under an FR3 reinforcement schedule. (a) Active nosepokes during 1-hour and 3-hour SA sessions with cocaine exposure and vehicle vapor (Veh) separately. Linear regression analysis showed progressive escalation of active nosepokes in the long-access cocaine vapor group (LgA; slope = 1.837, CI = [0.820, 2.853], r² = 0.684, p** = 0.003, n = 8), whereas no significant increase was observed in the short-access cocaine vapor group (ShA; slope = −0.285, CI = [−0.910, 0.394], r² = 0.094, p = 0.389, n = 4) or in the Veh group ( slope = −0.145, CI = [−0.521, 0.230], r2 = 0.091, p = 0.398, n = 4) across the 3-hour SA sessions. The slope of the LgA group was significantly different from that of the ShA group (F3,16 = 15.99, p** = 0.001) and the Veh group (p*** < 0.001). (b) Escalation of cocaine vapor deliveries during 3-hour SA sessions in the LgA group (slope = 0.579, CI = [0.266, 0.892], r2 = 0.695, p** = 0.003), whereas the Veh group remained low and stable (slope = −0.079, CI = [−0.233, 0.076], r2 = 0.147, p = 0.273). The comparison between LgA versus Veh shows significant differences (p* < 0.001). (c) Escalation of active nosepokes (slope = 0.644, CI = [0.272, 1.016], r2 = 0.666, p** = 0.004) and cocaine vapor deliveries (slope = 0.185, CI = [0.051, 0.319], r2 = 0.558, p* = 0.013) during the first hour of 3-hour SA sessions. (d) Escalation of active nosepokes in LgA group (slope = 17.092, CI = [10.633, 23.552], r2 = 0.823, p*** < 0.001); inactive nosepokes (slope = 0.801, CI = [−0.426, 2.027], r² = 0.221, p = 0.171, n = 8) and Veh group (slope = −1.709, CI = [−4.806, 1.388], r² = 0.168, p = 0.239) during the timeout period of 3-hour SA sessions.
To directly assess whether mice are seeking drug versus exhibiting non-specific head entries into the two nosepoke holes, we compared active versus inactive nosepokes between 1-hour and 3-hours FR3 SA sessions. As shown in Supplemental Figure S2A, mice exhibited a clear preference for active nosepokes. Active responses were higher than inactive responses during 1-hour sessions (p < 0.001) and remained elevated during 3-hour sessions (p < 0.001; Supplemental Figure S2B). To examine potential sex differences, we analyzed the SA data in female (n = 5) and male (n = 5) mice separately (Supplemental Figure S3). Both male and female groups escalated responding for cocaine. Specifically, there was no significant difference of regression slope for female versus male mice for active nosepokes (female: slope = 2.997, CI = [1.270, 4.723], r² = 0.667, p = 0.004, n = 5; vs. male: slope = 1.628, CI = [0.470, 2.785], r² = 0.568, p = 0.012, n = 5) as well as vapor deliveries (female: slope = 0.919, CI = [0.309, 1.530], r² = 0.601, p = 0.008; vs. male: slope = 0.561, CI = [0.175, 0.946], r² = 0.585, p = 0.01). The active nosepokes within the first hour also showed no significant difference between females (slope = 1.263, CI = [0.573, 1.953], r² = 0.690, p = 0.003) and males (slope = 0.655, CI = [0.279, 1.03], r² = 0.669, p = 0.004). The number of active nosepokes during the timeout period was higher in females (slope = 25.327, CI = [13.877, 36.777], r² = 0.765, p = 0.001) than in males (slope = 6.307, CI = [3.345, 9.268], r² = 0.751, p = 0.001; p* = 0.002; Supplemental Figure S3D).
LgA vaporized cocaine-induced weight loss
To assess the influence of vaporized cocaine SA on body weight, we recorded the weight of the mice every 2 days during the 3-hour SA sessions (Supplemental Figure S1A). Body weight decreased during the 10 days of 3-hour SA sessions (n = 4; Supplemental Figure S1B). The linear regression analyses showed a significant decrease in averaged body weight from 23.75 ± 1.55 g on day 2 to 21.75 ± 1.80 g on day 10 (slope = −0.263, CI = [−0.45, −0.075], r2 = 0.868, p = 0.021; Supplemental Figure S1C). A minor reduction in body weight (~6%) was observed after LgA exposure to vaporized cocaine. The mean body weight was negatively correlated with cocaine intake during 3-hour SA sessions (r = −0.9, p* = 0.036, n = 4; Supplemental Figure S1D). This is consistent with the well-known anorexigenic effects of cocaine and provides further evidence of cocaine’s pharmacological effects.
Discussion
In this study, we developed and validated a vapor-based cocaine SA model in mice that reflects many important aspects of compulsive cocaine use and CUD in humans. We determined the appropriate concentration range for vaporized cocaine to induce hyperactivity and detectable blood cocaine levels. We also found that mice were motivated to self-administer cocaine vapor as indicated by their elevated responses under higher reinforcement schedules (e.g., FR3) and clear preference for the active versus the inactive nosepoke hole. Moreover, mice showed escalation of cocaine vapor intake when access was extended (to 3 hours). Together, these results highlight vaporized cocaine SA as a novel and reliable model for studying behavioral and physiological effects of cocaine misuse and addiction.
Locomotor activity is an index of cocaine’s psychostimulant effects, reflecting hyperdopaminergic states and serving as a primary readout for drug-induced behavioral activation. Our results show that the hyperactivity behavioral pattern post vaporized cocaine exposure versus Veh (Figure 2) is consistent with the previous reports from intraperitoneal injected cocaine experiments (Runegaard et al., 2018; Sabeti et al., 2003; Yamamoto et al., 2013) and intravenous cocaine SA (Koeltzow and Vezina, 2005; Roberts et al., 2018). This similarity indicates that the cocaine-induced effect is replicated using our inhalation route, highlighting a pharmacological effect that is a shared mechanism of various systemic delivery routes. In addition to more common measurements, such as the total distance travelled, we assessed other locomotor parameters, including distance travelled, average speed, and the proportion of mobility vs. immobility time. Together, these results indicate that the enhanced locomotor activity is likely to reflect cocaine’s effect on mesocorticolimbic dopamine pathways, leading to increased excitatory signaling within the reward circuit, thus validating the pharmacological effects of the new vapor delivery route of cocaine.
In parallel, assessing blood cocaine concentration provides direct evidence of cocaine absorption following vaporized cocaine exposure in mice. Our results showed that blood cocaine levels were in a close range of clinical data (Jenkins et al., 1999, 2002). We observed that blood cocaine concentrations following vapor exposure were modestly lower than those typically reported after intravenous (i.v.) administration (Booze et al., 1997). This difference may reflect route- and formulation-dependent pharmacokinetic properties (Jeffcoat et al., 1989; Jenkins et al., 2002) and may also likely be attributed to delayed blood sample collection (i.e., ~7–10 minutes after the last cocaine vapor exposure). The relatively large variability in plasma cocaine levels observed under passive exposure to 40 mg vapor delivery likely reflects inter-individual differences in respiratory rate, proximity to the vapor inlet port, metabolic clearance (especially for drugs with a short half-life like cocaine) and blood sample collection timing. However, our behavioral results unequivocally indicate pharmacological effects. The robust positive correlation between blood cocaine level and locomotor activity (Figure 3) provides strong evidence that vaporized cocaine is efficiently absorbed by mice and exerts behaviorally relevant effects in a dose-dependent manner.
We employed an FR1 schedule during the initial acquisition phase to ensure that animals rapidly learned to associate nosepokes with cocaine vapor delivery. The poor discrimination between active and inactive nosepoke holes under the FR1 schedule was not surprising. Nose-poking represents a natural exploratory behavior in rodents, which can generate a high baseline level of responding across both nosepoke holes. Consequently, some degree of responding in the inactive hole is expected and does not necessarily reflect a lack of volitional cocaine intake. However, clear discrimination was observed when the reinforcement schedule was increased to FR3. Our results show that shifting reinforcement schedules from FR1 to FR3 significantly increased active nosepokes with responding nearly tripling, whereas the number of vapor deliveries remained unchanged (Figure 4[a]–[c]). This finding suggests that mice maintained active responding when the response requirement increased, indicating that cocaine vapor functioned as an effective reinforcer under a more demanding schedule. Importantly, the unchanged total cocaine vapor intake indicates that the reinforcing efficacy of cocaine vapor was still maintained under higher response requirements, rather than being driven by non-specific increases in activity. Additionally, previous studies showed that responding for Veh SA (i.e., in the absence of drug) or responding for a light cue did not increase when the reinforcement schedule was increased, and the number of Veh or light cue deliveries decreased significantly at higher FRs (Moussawi et al., 2020). These findings suggest that active nosepoke responding reported in the present study was reinforced by cocaine’s effects. These results are consistent with the conventional intravenous cocaine SA models, where higher FR value causes increases in the rate of lever-pressing but drug intake remains stable despite the increased FR value (Desai et al., 2023; Lee et al., 2016). Moreover, similar findings have been observed in vapor-based SA models of other drugs, such as fentanyl (Moussawi et al., 2020) and cannabis (Freels et al., 2020), highlighting the generalizability of inhalation models for studying addictive substances. Furthermore, we also found that the latency to initiate the first nosepoke gradually decreased across training sessions (Figure 4[d]), indicating that mice learned the operant response required to obtain cocaine vapor.
Mice exposed to LgA SA sessions escalated their responding in the active nosepokes compared to those exposed to ShA sessions (Figure 5). Notably, the number of nosepokes during the timeout period also showed escalation across LgA SA sessions, suggesting that the mice continued to seek the drug even when reinforcement was unavailable. Additionally, increased responding during the timeout period reflects persistent cocaine-seeking behavior, but one cannot fully exclude the possibility that incomplete stimulus control also contributed to this pattern. Identical patterns of responding have been reported with traditional intravenous cocaine SA models in rats (Ahmed and Koob, 1998, 1999; Beasley et al., 2023; Mandt et al., 2015). Escalated cocaine vapor intake indicates that mice increase their cocaine-seeking and self-administration behavior over time, suggesting reduced sensitivity to the reinforcing effects of the drug. This pattern of responding is a typical hallmark reflecting the transition from drug misuse to drug addiction (Picetti et al., 2010). A neuropharmacological explanation involves hedonic tolerance, where repeated exposure decreases the drug’s efficacy, leading to increased intake to achieve the desired reward (Bardo et al., 2025; Koob and Volkow, 2016). Much preclinical and clinical data favor a reward deficit hypothesis involving negative reinforcement (Thomsen and Caine, 2007).
Importantly, our data support a pharmacological effect of cocaine because of several reasons: (1) responses in the active hole, but not in the inactive hole, increased when the schedule of reinforcement increased from FR1 to FR3 (Figure 4); (2) mice in the LgA group escalated cocaine SA, whereas Veh SA remained stable (Figure 5); (3) active nosepoke responding during the timeout period also selectively increased in the LgA cocaine group, whereas responding remained stable in the cocaine inactive hole and the active hole for the Veh group (Figure 5); and (4) additional evidence supporting the reinforcing effects of vaporized drugs in similar models has been previously reported (Moussawi et al., 2020).
The escalation in cocaine vapor intake that was observed in the present study might indicate the development of tolerance to cocaine’s reinforcing effects over time with increased responding to maintain optimal intoxication levels. However, because only a single cocaine dose was tested in the present study, these findings may not be definitively interpreted as direct evidence of tolerance. Further studies using multiple self-administered cocaine doses and additional behavioral tests will be needed to determine leftward, rightward, or downward shifts in the cocaine dose-effect function.
An important procedural distinction between the present vapor SA and intravenous SA models is that vapor delivery was slower and more prolonged than the rapid, discrete drug delivery produced by intravenous infusion. As a result, SA rates and intake patterns in the vapor paradigm are likely constrained by vapor generation, chamber exposure, and clearance intervals, and therefore should not be interpreted as directly equivalent to those observed in intravenous models (Kawa et al., 2019). However, the vapor approach offers several practical and translational advantages, including avoidance of catheter implantation and the associated surgical burden, catheter maintenance, and eventual loss of catheter patency, all of which are well-recognized technical challenges in mouse intravenous SA studies (Thomsen and Caine, 2007). These features make the vapor particularly useful for longer-term studies and potentially more compatible with experiments that combine drug exposure with other longitudinal procedures. In addition, vapor delivery may provide greater translational relevance for modeling inhaled/smoked cocaine exposure, as route of administration is known to shape cocaine disposition and pharmacokinetics in humans.
A limitation of this study is that most intravenous cocaine SA studies in rats used 6-hour or longer sessions to elicit robust escalation. However, depending on multiple factors, such as speed of delivery, intermittent access, and early acquisition, some studies have shown escalation in 3-hour, albeit less robust than that with 6-hour access (Allain and Samaha, 2019; Beckmann et al., 2012; Mandt et al., 2015). Very few if any studies have used extended access in mice, but one study with social stress showed escalation with 2-hour access (Han et al., 2015). Our goal in the present study was to distinguish the 3-hour LgA from the 1-hour ShA conditions and to establish a technically feasible mouse cocaine vapor SA paradigm rather than providing a comprehensive SA study. We selected the 3-hour session because it provided substantially greater cocaine vapor exposure and sufficient enhancement in active responding than the ShA (1 hour) group while maintaining experimental feasibility, and in fact, escalation of intake. In addition, at the current stage of development, only two operant chambers were available for the study, and extending to 6-hour sessions was not feasible due to substantially increased experimental burden (e.g., personnel time, drug-use expenditure, overall study duration). Nevertheless, longer sessions would provide valuable features to more closely compare with the intravenous LgA literature. Another limitation is that vaporized cocaine SA model does not allow precise body weight-based dosing (mg/kg) for individual animals. As a result, the actual absorbed dose may vary depending on such factors as body size, respiratory rate, and metabolism. To reduce this variability, we used mice of similar age and exposed all animals under standardized conditions, including the same amount of cocaine used for each vaporization, chamber configuration, airflow rate, vapor delivery duration, and total exposure time. In addition, blood cocaine level measurements and locomotor responses indicated that the selected vapor exposure conditions produced measurable and generally consistent cocaine-related effects across animals; however, individual variability, common in behavioral studies, cannot be fully accounted for when estimating actual systemic exposure.
Furthermore, the low efficiency of the cocaine delivery is also a limitation of this study as most of the cocaine vapor is exhausted by the suction pump rather than inhaled by the mouse. To address this, we regulated the airflow rate, atomizer power, and the vaporizing time to maintain the cocaine vapor within the SA chamber before for 60 seconds was exhausted. This ensured that the mouse in the chamber had sufficient exposure to cocaine vapor and minimized vapor loss. In addition, as the atomizers are designed to vaporize the total amount of cocaine powder in a given tank during each vaporization, 10 tanks were installed to sequentially vaporize cocaine upon nosepokes. Manual cocaine refilling of these tanks was required before the next cycle, which required supervision by an operator that could have potentially disturbed the animals during the LgA sessions. To solve this problem, several tanks were rapidly refilled at fixed time intervals (~30 minutes). Future improvements will focus on optimizing the vapor delivery technology to sustain longer, consistent vaporization without frequent refilling. For instance, purging air from the atomizer rather than all 10 atomizers can substantially increase cocaine concentration in the vapor and thus reduce cocaine use. Such technical advances would minimize drug loss, reduce animal disturbance, and further enhance the reliability of vapor-based SA models. Vapor clearance from the chambers was verified by visual inspection in the present study. A more rigorous assessment of vapor clearance will require dedicated air-sampling and analytical methods and may be used in the future.
Here, we developed a noninvasive vapor-based SA model of cocaine misuse and addiction in mice. Compared to traditional intravenous SA models, our vaporized cocaine model offers several advantages. The vapor delivery mimics the inhalation route commonly used by human cocaine users, which produces rapid delivery of the drug to the brain (Volkow et al., 2000). Similarly, the vapor model reproduces the pharmacokinetic effects and behavioral responses associated with human cocaine use, thereby enhancing its translational relevance. In contrast, to intravenous SA models that require catheter implantation and constant maintenance, the vapor model does not require surgery, thus eliminating potential complications of infection, vascular damage, and catheter blockage. It also reduces chronic pain and stress to animals that may confound behavioral and physiological responses to cocaine. These features are particularly advantageous for long-term experiments that investigate the neuropharmacological effects of chronic drug exposure. Together, the vapor model better reflects human patterns mimicking smoked cocaine use, simplifies long-term experimental design, and may avoid some surgery related burdens for catheterization. These advantages make vaporized cocaine SA a robust model, especially for mouse models, to study cocaine addiction and to test interventions aimed at reducing cocaine use.
In summary, by demonstrating dose-dependent increases in blood cocaine levels and locomotor activity, enhanced drug seeking behavior under higher reinforcement schedules, and escalation of cocaine vapor intake, our vaporized cocaine SA model captures key hallmarks of CUD and provides a foundation for future investigations into the neurobiological mechanisms and potential therapeutic interventions for cocaine addiction. Such work may help bridge the gap between preclinical research and clinical efforts to develop effective treatments for CUD.
Supplemental Material
sj-docx-1-jop-10.1177_02698811261453842 – Supplemental material for A noninvasive approach to modeling cocaine self-administration in mice
Supplemental material, sj-docx-1-jop-10.1177_02698811261453842 for A noninvasive approach to modeling cocaine self-administration in mice by Yanzuo Liu, Qiaochu Wang, George F. Koob, Leandro F. Vendruscolo, Nora D. Volkow, Congwu Du and Yingtian Pan in Journal of Psychopharmacology
Footnotes
Acknowledgements
The authors thank Maury Cole for his advice and help in setting up the vapor delivery mechanism, Wensheng Cheng for help with locomotor tracking, and Michael Arends for help with editing the manuscript.
Author contributions
Y.P., G.F.K., N.V., L.F.V., and C.D. designed the research. Y.L. and Q.W. conducted animal studies and data analysis (contributed equally, listed alphabetically). All authors contributed to data interpretation, result discussions, and manuscript writing. The contributions of the National Institutes of Health authors were made as part of their official duties as National Institutes of Health federal employees, follow agency policy requirements, and are considered Works of the United States Government. However, the findings and conclusions that are presented in this article are those of the authors and do not necessarily reflect the views of the National Institutes of Health or the U.S. Department of Health and Human Services.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported in part by National Institutes of Health grants 2R01 DA029718 (C.D., Y.P.), RF1DA048808 (Y.P., C.D.), and R21DA042597 (C.D., Y.P.) and the NIH’s Intramural Research Programs of the National Institute on Drug Abuse (G.F.K., L.F.V.) and of National Institute on Alcohol Abuse and Alcoholism (N.V.). The operant vapor self-administration station was purchased from La Jolla Alcohol Research for modifications. Freebase cocaine was provided by the National Institute on Drug Abuse Drug Supply Program.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.