
Abstract
Background:
Major depressive disorder is one of the most debilitating psychiatric disorders worldwide. First-line treatments such as selective serotonin reuptake inhibitors have significant limitations, including delayed onset of therapeutic effects and treatment resistance in about 30% of patients. Increasing evidence suggests that acute administration of serotonergic psychedelics, such as psilocybin, produces rapid and long-lasting antidepressant effects, including in treatment-resistant patients. However, it remains unknown which specific 5-HT receptor subtype mediates psilocybin’s antidepressant activity.
Methods:
We examined in Wistar rats whether pretreatment with the 5-HT2B receptor (5-HT2BR) antagonist RS-127445 (0.32, 1.0, or 3.2 mg/kg) blocked the rapid (day 1) and sustained (day 21) behavioral effects of a single psilocybin administration (0.32 mg/kg) in the forced swim test (FST), a test with predictive validity for antidepressant efficacy. We also measured the impact of RS-127445 on psilocybin-induced head-twitch response (HTR), a behavioral proxy in rodents for psychedelic properties.
Results:
Our data showed that psilocybin produced both a rapid and sustained decrease in immobility and an increase in climbing behavior in the FST and significantly increased HTR counts. Although RS-127445 did not affect HTR counts at any tested dose, it dose-dependently reversed both the rapid and sustained psilocybin-induced reductions in immobility and increases in climbing behavior.
Conclusion:
These findings indicate that 5-HT2BRs are required for psilocybin’s behavioral effects in the FST, but are not required for its HTR. The results add to evidence that psilocybin’s predictive validity in the FST can be dissociated from its 5-HT2A-mediated psychedelic effects.
Introduction
Major depressive disorder (MDD) is a debilitating psychiatric disorder affecting over 330 million individuals globally. Characterized by persistent low mood, anhedonia, hopelessness, and significant somatic disturbances (i.e., fatigue, weight changes, and sleep disturbances), MDD imposes a staggering personal and socioeconomic burden (Malhi and Mann, 2018). While first-line pharmacological interventions, such as selective serotonin reuptake inhibitors (SSRIs) and serotonin–norepinephrine reuptake inhibitors, remain the standard of care, their clinical utility is frequently undermined by a delayed therapeutic onset and burdensome side-effects such as sexual dysfunction and gastrointestinal disturbances (Lorenz et al., 2016; Oliva et al., 2021). Most critically, approximately 30% of patients fail to achieve an adequate clinical response to at least one trial of conventional antidepressants (drugs for depression; Cipriani et al., 2018; Henssler et al., 2018). This condition, commonly referred to as treatment-resistant depression (TRD), represents a major public health burden and underscores the urgent need for novel, rapid-acting, and durable antidepressant interventions. Among them, psilocybin, a psychedelic compound, has emerged as a promising therapeutic option for both MDD and TRD (Nichols, 2016).
Psilocybin is a tryptamine alkaloid found naturally in numerous species of Psilocybe mushrooms. The psychedelic properties of psilocybin, like other classical psychedelics, are attributed to activation of the serotonin 5-HT2A receptor (5-HT2AR). Human studies have indeed shown that psilocybin produced substantial brain 5-HT2AR occupancy, which correlated positively with subjective psychedelic intensity (Madsen et al., 2019). Further evidence for 5-HT2AR involvement comes from a study showing that psilocybin’s effects on positive mood and on the attenuation of recognition of negative facial expressions were blocked by the 5-HT2AR antagonist ketanserin (Kometer et al., 2012). Preclinical studies provided additional support with consistent reports of psilocybin induced increase in the frequency of the head-twitch response (HTR; Erkizia-Santamaría et al., 2022; Seillier et al., 2025; Shahar et al., 2022), a behavioral correlate of 5-HT2AR activation widely adopted as a proxy in rodents for human hallucinogenic effects (Halberstadt et al., 2020). These increases in the HTR were blocked by the 5-HT2AR antagonists M100907 and MDL11939 (Erkizia-Santamaría et al., 2022; Shahar et al., 2022) and were absent in 5-HT2AR knockout mice (González-Maeso et al., 2007). Collectively, these findings support the idea that 5-HT2AR stimulation is a key determinant of the psilocybin-induced psychedelic experience.
Until recently, it was assumed that psilocybin’s therapeutic effects required the activation of 5-HT2ARs and the associated psychedelic and/or alteration of consciousness effects (Cameron et al., 2023; Miranda, 2025; Vargas et al., 2023). Although psilocybin’s antidepressant-like effects are often studied within this 5-HT2AR agonism framework, emerging data suggest that its psychedelic effect may not be required for its antidepressant-like actions (Hesselgrave et al., 2021; Sekssaoui et al., 2024). Specifically, Hesselgrave et al. (2021) showed that psilocybin-induced reversion of anhedonia (as measured by the sucrose preference test (SPT) in mice exposed to a chronic multimodal stress paradigm) was not blocked by the 5-HT2AR antagonist ketanserin, unlike its psychedelic-like action (as assessed by the HTR). Similarly, Sekssaoui et al. (2024) demonstrated that psilocybin’s antidepressant-like effects in the SPT and the novelty-suppressed feeding (NSF) test in a chronic despair mouse model persisted in 5-HT2AR knockout mice. By contrast, the two 5-HT2AR agonists, DOI and lisuride, had their antidepressant-like effects suppressed in 5-HT2AR knockout mice. In line with these observations, a recent clinical case study confirmed the preserved efficacy of psilocybin when combined with a potent 5-HT2AR antagonist in an adult with TRD (Rosenblat et al., 2023). As described by the authors, despite the absence of psychedelic effects, the patient experienced rapid, robust, and sustained antidepressant effects. Together, these data indicate that psilocybin, unlike other psychedelics, exerts its antidepressant effects independently of its 5-HT2AR-mediated psychedelic effects.
Given the proposed involvement of 5-HT1AR in the mechanism of action of conventional antidepressant drugs, such as SSRIs (Artigas, 2015; Celada et al., 2013), there has been growing interest in their potential role in psilocybin’s antidepressant-like effects. However, Sekssaoui et al. (2024) recently showed that psilocybin’s antidepressant-like effects, assessed across several behavioral tests in the chronic despair model, were not inhibited by the 5-HT1AR antagonist WAY-100635, thereby questioning the involvement of these receptors in psilocybin’s antidepressant-like effects. In contrast, WAY-100635 blocked psilocybin-induced hypothermia (Erkizia-Santamaría et al., 2022), suggesting that 5-HT1AR activation mediated psilocybin’s hypothermic effect. These observations have increased recognition that psilocybin’s therapeutic profile and side effects (e.g., 5-HT1AR-mediated hypothermia) likely reflect a complex interplay among multiple serotonin receptor subtypes, allowing a possible dissociation between its psychedelic and antidepressant actions (Hesselgrave et al., 2021; Sekssaoui et al., 2024). However, the pharmacological mechanism underlying psilocybin’s antidepressant-like effects remains unknown.
Importantly, psilocybin, or more specifically its active metabolite psilocin, can bind to and activate many other 5-HT receptors: 5-HT2BR > 5-HT1DR > 5-HT1ER > 5-HT1AR > 5-HT5AR > 5-HT7R > 5-HT6R > 5-HT2CR > 5-HT1BR > 5-HT2AR (ordered based on psilocin’s affinity; Ray, 2010). Paradoxically, despite psilocin’s highest binding affinity for 5-HT2BRs over the other receptors, including 5-HT2AR (Ray, 2010), psilocybin research has focused on the latter, and the role of 5-HT2BRs in its antidepressant-like effects has been largely overlooked. Interestingly, 5-HT2BR activity might contribute, alongside their action on the serotonin transporter, to the therapeutic effectiveness of SSRIs (Hertz et al., 2012). In particular, the antidepressant effect of fluoxetine was abolished in 5-HT2B knockout mice or after pharmacologic inactivation of 5-HT2BRs in the NSF test (Diaz et al., 2012). Moreover, the behavioral and biochemical antidepressant-like effects of fluoxetine and paroxetine were mimicked by administration of the selective 5-HT2BR agonist BW723C86 in mice (Diaz et al., 2012; Diaz and Maroteaux, 2011). Given these premises, the present study was designed to test whether blocking 5-HT2BR prevents the behavioral effects of psilocybin in the forced swim test (FST), a behavioral assay with predictive validity for antidepressant efficacy in otherwise healthy rodents (Commons et al., 2017). Importantly, while the ability of conventional antidepressants to promote active coping and reduce immobility in the FST in normal rodents has been shown to predict their antidepressant efficacy (Bianchi et al., 2002; Fernández-Guasti et al., 2017), these observations were extended to psilocybin and other fast-acting antidepressants like ketamine (Carhart-Harris et al., 2016; Hibicke et al., 2020; Kolasa et al., 2024; Murrough et al., 2013; Sekssaoui et al., 2024). Specifically, we compared in Wistar rats the effect of pretreatment with the selective 5-HT2BR antagonist RS-127445 (at doses of 0.32, 1.0, or 3.2 mg/kg) on psilocybin (0.32 mg/kg) induced HTR and behavioral effects in the FST. We also examined whether this pretreatment would affect the rectal temperature response to psilocybin. The identification of specific, non-psychedelic receptor mechanisms is indeed critical for developing novel therapeutics that harness psilocybin’s potent antidepressant potential without inducing hallucinogenic or other side effects (e.g., hypothermia). Such an advance would significantly increase the clinical utility and patient accessibility of this promising class of compounds.
Materials and methods
Animals
Male Wistar rats (180–200 g, n = 64) were purchased from Charles River Laboratories International, Inc. (Germany) via a local supplier, Velaz (Prague, Czech Republic). Rats were housed two per cage at 22 ± 1°C under a 12-hour light-dark cycle with free access to food and water (ad libitum). Animals were habituated to these housing conditions (i.e., standard cages without environmental enrichment) for 2 weeks before the start of the experiments. All experiments involving animals were conducted with the approval of the Committee for Animal Care and Use of the Third Faculty of Medicine, Charles University, Prague, the Ministry of Education, Youth and Sports of the Czech Republic (protocol code: MSMT-25258/2025-4), and the Directive 2010/63/EU for the protection of animals used for scientific purposes (European Parliament and Council of the European Union; as amended on June 9, 2019).
Treatment schedule
Behavioral testing was performed in the morning during the light phase. Tests were conducted from day 0 (pretreatment with either vehicle or the 5-HT2BR antagonist RS-127445, followed 30 minutes later by acute saline or psilocybin administration) to 3 weeks post-treatment (Figure 1). Animals were weighed weekly.
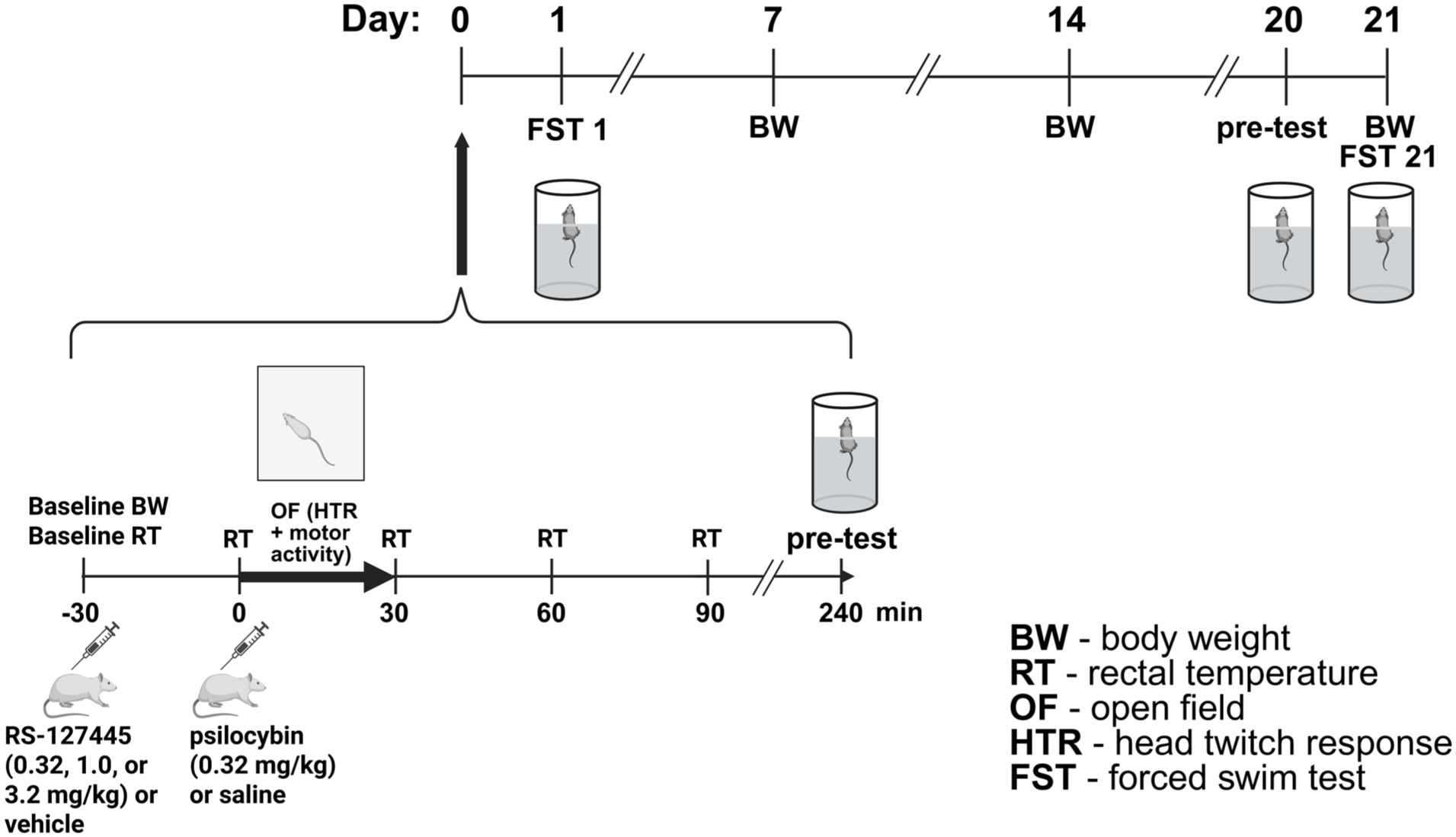
Treatment schedule and behavioral battery. Behavioral procedures spanned 21 days. Baseline body weight (BW) was measured on day 0 (before baseline rectal temperature (RT) measurement and drug administration), and subsequently measured weekly. On day 0, baseline RT was measured before pretreatment with RS-127445 or vehicle (−30 minutes), immediately before psilocybin or saline injection (0 minute), and then every 30 minutes for 90 minutes post-administration. Immediately after injection (0 minute), rats were placed in the open field (OF) arena to assess the head-twitch response (HTR) and locomotor activity. The FST was conducted 1 and 21 days after psilocybin administration (FST 1 and FST 21, respectively), with a pre-test performed 24 hours earlier (day 0 and day 20, respectively).
Drugs and treatments
Psilocybin, purchased from Cayman Europe OÜ (Tallinn, Estonia, No. 14041), was dissolved at a concentration of 1 mg/ml in sterile saline solution (0.9% NaCl) and administered intraperitoneally (i.p.) at 0.32 mg/kg of body weight. Control animals received an i.p. injection of the sterile saline solution (0.9% NaCl). This dose was chosen based on our previous study as the lowest effective dose to produce robust behavioral effects in the FST, as well as pro-social and hedonic effects (Seillier et al., 2025).
The selective 5-HT2BR antagonist RS-127445, purchased from Sigma Aldrich (Schnelldorf, Germany, No. R2533), was dissolved in a vehicle consisting of Tween80:polyethylene glycol:physiological saline (0.9% NaCl) in a ratio of 5:5:90. The antagonist or its vehicle was administered i.p. in a volume of 2 ml/kg of vehicle solution. To establish a detailed dose–response curve, we used 3 doses of RS-127445 (0.32, 1.0, or 3.2 mg/kg); this choice was based on a previously published in vivo dose–response study in rats (Cathala et al., 2020; Devroye et al., 2016).
On day 0, animals received a single i.p. injection of either the 5-HT2BR antagonist (RS-127445 at 0.32, 1.0, or 3.2 mg/kg) or its vehicle, as pretreatment. This was followed, 30 minutes later, by a second i.p. injection of either saline (saline-treated group) or psilocybin (0.32 mg/kg, psilocybin-treated groups). Accordingly, the experiment utilized 8 experimental groups (n = 8 per group): vehicle-saline (VEH-SAL), 0.32 mg/kg RS-127445-saline (RS0.32-SAL), 1.0 mg/kg RS-127445-saline (RS1.0-SAL), 3.2 mg/kg RS-127445-saline (RS3.2-SAL), vehicle-psilocybin (VEH-PSI), 0.32 mg/kg RS-127445-psilocybin (RS0.32-PSI), 1.0 mg/kg RS-127445-psilocybin (RS1.0-PSI), and 3.2 mg/kg RS-127445-psilocybin (RS3.2-PSI). To control for time-of-day and weekly variations, treatment groups (n = 8 per group) were allocated to testing slots using an 8 × 8 Cyclic Latin Square design. This ensured that each treatment was represented in every procedural time slot across the 8-week testing period.
A sensitivity power analysis for the mixed 2 × 4 × 2 design (α = 0.05, power = 0.80) indicated that a sample size of eight rats per group provides sufficient sensitivity to detect effects as small as f = 0.25, consistent with the variability observed in our previous study (Seillier et al., 2025).
Rectal temperature measurement
Rectal temperature was measured as previously described (Seillier et al., 2025). Briefly, a probe was inserted 2 cm into the rectum, and a steady temperature readout was obtained within 10 seconds. Importantly, animals were acclimated to the procedure by taking rectal temperature measurements several times before the study began. On the testing day (day 0), animals were placed in the testing room, with the temperature maintained at 24 ± 1°C, 1 hour before any recordings. Rectal temperature was measured immediately before the pretreatment with vehicle or the 5-HT2BR antagonist, 30 minutes later (i.e., immediately before the saline or psilocybin injection), and then every 30 minutes for 90 minutes. All temperature measurements were performed in the morning (starting at 7:00 AM) to avoid baseline temperature differences caused by circadian rhythms.
Locomotor activity and HTR
Immediately after the single injection of psilocybin or saline—that is, 30 minutes after the pretreatment with RS-127445 or vehicle—rats were placed in an Open Field arena (45 cm × 45 cm × 45 cm), and their behavior was recorded with a camera device for 30 minutes. The arena was thoroughly cleaned with 70% ethanol and dried between test sessions. The rats’ movements were quantified as mobility time, distance traveled (cm), and velocity (cm/s), using EthoVision XT 15 tracking software (Noldus Information Technology).
In addition to the locomotor activity, we also assessed the HTR as previously described (Seillier et al., 2025). HTRs, commonly used in rodents to assess 5-HT2AR activation (Halberstadt and Geyer, 2013), were manually scored by an experimenter blind to the study as a mixture of head shakes and/or whole-body shakes, similar to the behavior of dogs shaking off water (Halberstadt and Geyer, 2018).
Forced swim test
The FST is a rodent paradigm often used to assess the efficacy of antidepressants in normal rats (i.e., not depressed-like animals). It consists of 2 sessions, 24 hours apart: the pre-test and the test session (Porsolt et al., 1977). The test and its pre-test session were performed twice: on day 1 (FST 1; with the pre-test on day 0, 4 hour after injection) and day 21 (FST 21; with the pre-test on day 20) to assess the rapid and sustained behavioral effects, respectively. A 15-minute pre-exposure session to the paradigm 1 day before the test session is used to elicit a consistent passive coping strategy (immobility) in otherwise healthy rats (Slattery and Cryan, 2012). As previously described (Seillier et al., 2025), during the pre-test session, rats were placed into a glass cylindrical tank (48 cm × 24 cm) containing 30 cm of water at 24 ± 1°C. The water depth was such that the rats could not support themselves with their hind paws, and their tails did not touch the bottom of the tank while keeping their noses above water. After a 15-minute swim, the rats were removed, towel-dried, and placed in a specialized warming environment (e.g., a recovery cage under an infrared lamp) for 20 minutes, then returned to their home cages. Fresh water was used for each animal. On the testing day (24 hours later), the rats were exposed to a 5-minute swim under the same conditions, followed by the same recovery period. Unlike the pre-test, which was not recorded, the 5-minute test session was recorded for later scoring for immobility, climbing, and swimming. The FST behaviors were scored manually by an experimenter blind to the study, using the modified sampling technique by scoring the predominant behavior in each 5-second period of the 5-minute test (Slattery and Cryan, 2012). Immobility was defined as no active attempts to escape while maintaining a floating posture, with only the movements necessary to keep their heads above water. Swimming was defined as actively attempting to escape with motions directed outward against the wall of the cylinder. Climbing was defined as actively attempting to escape with upward motions against the cylinder walls.
Statistics
Outliers were predefined using Carling’s median rule, using the recommended ideal fourths and constant k2 adjusted for sample size [k2 = (17.63n–23.64)/(7.74n–3.71)] (Carling, 2000), ensuring inclusion criteria remain objective and reproducible. This procedure was applied uniformly to each dataset before statistical analysis, with exclusion based solely on this predefined criterion. In each dataset, the Gaussian distribution was verified using the Shapiro–Wilk normality test, and the homogeneity of variances using Levene’s test. Data were analyzed by two-way or three-way Analysis of Variance (ANOVA) followed by Tukey’s Honestly Significant Difference (HSD) or Unequal N Tukey’s HSD post hoc tests for groups with normal distribution, or by Kruskal–Wallis test with Dunn’s post-hoc adjustment for groups with nonparametric distribution and/or violation of homoscedasticity. The Tukey’s HSD (or Unequal N Tukey’s HSD) post hoc tests were chose to maintain a conservative threshold for significance across all possible pairwise comparisons and to strictly control the family-wise error rate. This approach was chosen to rigorously validate our a priori hypotheses regarding treatment effects while simultaneously monitoring for any potential independent effects of the pharmacological agents. Body weight gain and locomotor activity were analyzed by a two-way ANOVA with Antagonist (four levels) and Agonist (two levels) as between factors. For the time-course temperature measurements, data were analyzed with a three-way mixed ANOVA with Antagonist (four levels) and Agonist (two levels) as between factors, and Time (four levels) as a repeated measure. HTR was analyzed by a nonparametric Kruskal–Wallis test. FSTs (FST 1 and FST 21) were analyzed by a three-way mixed model ANOVA with Antagonist (four levels) and Agonist (two levels) as between factors, and Time (two levels) as a repeated measure. The threshold for statistical significance was set at 0.05. Statistical analysis was performed using GraphPad Prism (v. 9.5.1) and Statistica (version 10, StatSoft, Inc. (2011)).
Results
Neither psilocybin nor RS-127445 altered body weight or rectal temperature
Although our previous work (Seillier et al., 2025) showed that psilocybin at 0.32 mg/kg did not alter body weight or rectal temperature (effects observed only at 1 and 3.2 mg/kg), given the possibility that RS-127445 could shift psilocybin’s dose–response profile, we nevertheless assessed whether pretreatment with the 5-HT2BR antagonist RS-127445 might modify these physiological measures in saline- or psilocybin-treated animals.
As shown in Figure 2(a), neither RS-127445 nor psilocybin affected body weight gain over the 21-day observational period compared to their respective saline-treated rats. The ANOVA did not reveal any effect of Antagonist (F3, 56 = 0.18, p = 0.91), Agonist (F1, 56 = 0.63, p = 0.43), nor of the interaction between these two factors (F3, 56 = 0.30, p = 0.83).
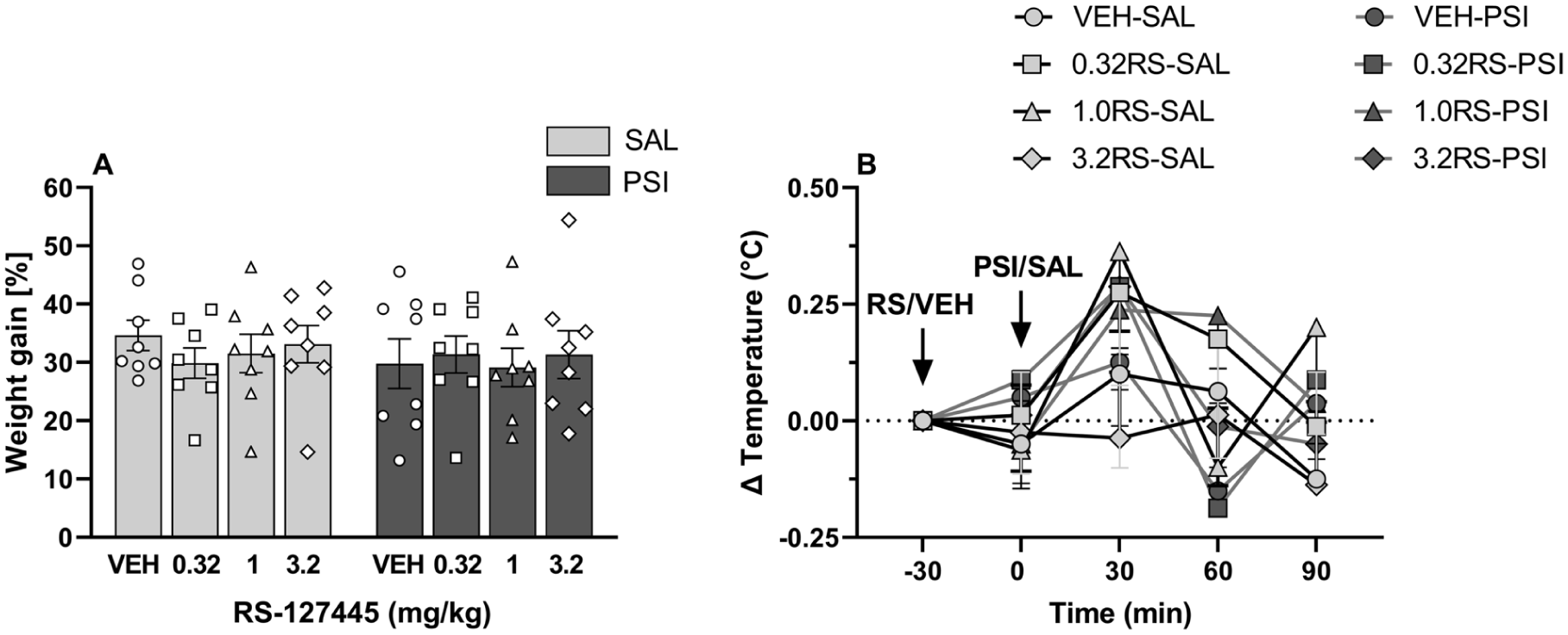
Psilocybin and RS-127445 did not alter body weight or rectal temperature. Neither psilocybin (0.32 mg/kg) nor pretreatment with RS-127445 (0.32, 1.0, or 3.2 mg/kg) affected (a) body weight as expressed in weight gain over 21 days (%) or (b) rectal temperature (°C). Data are plotted as means ± SEM (n = 8 per group). For the body weight, the individual values (for each rat) are reported as open circles for vehicle, open squares for 0.32 mg/kg RS-127445, open triangles for 1.0 mg/kg RS-127445, and open diamonds for 3.2 mg/kg RS-127445. The raw data are reported in the Supplemental file (S1 Raw data).
As shown in Figure 2(b), neither RS-127445 nor psilocybin produced any change in rectal temperature. Three-way repeated measures ANOVA revealed only a significant effect for Time (F3, 168 = 6.60, p < 0.001), but not for Antagonist (F3, 56 = 0.49, p = 0.69), Agonist (F1, 56 = 0.11, p = 0.74), nor for the interaction Antagonist × Agonist × Time (F9, 168 = 1.80, p = 0.07).
Neither psilocybin nor RS-127445 altered locomotor activity
Locomotor activity was also measured immediately after psilocybin or saline administration, with 30 minutes pretreatment with RS-127445 or vehicle in the Open Field Test (OFT). Data showed no significant differences between groups on mobility time (Antagonist (F3, 55 = 0.47, p = 0.71); Agonist (F1, 55 = 3.95, p = 0.052); Interaction (F3, 55 = 0.26, p = 0.85); Figure 3(a)), distance traveled (Antagonist (F3, 55 = 0.58, p = 0.63); Agonist (F1, 55 = 2.11, p = 0.15); Interaction (F3, 55 = 0.82, p = 0.49); Figure 3(b)) and velocity (Antagonist (F3, 55 = 0.58, p = 0.63); Agonist (F1, 55 = 2.12, p = 0.15); Interaction (F3, 55 = 0.82, p = 0.49); Figure 3(c)).
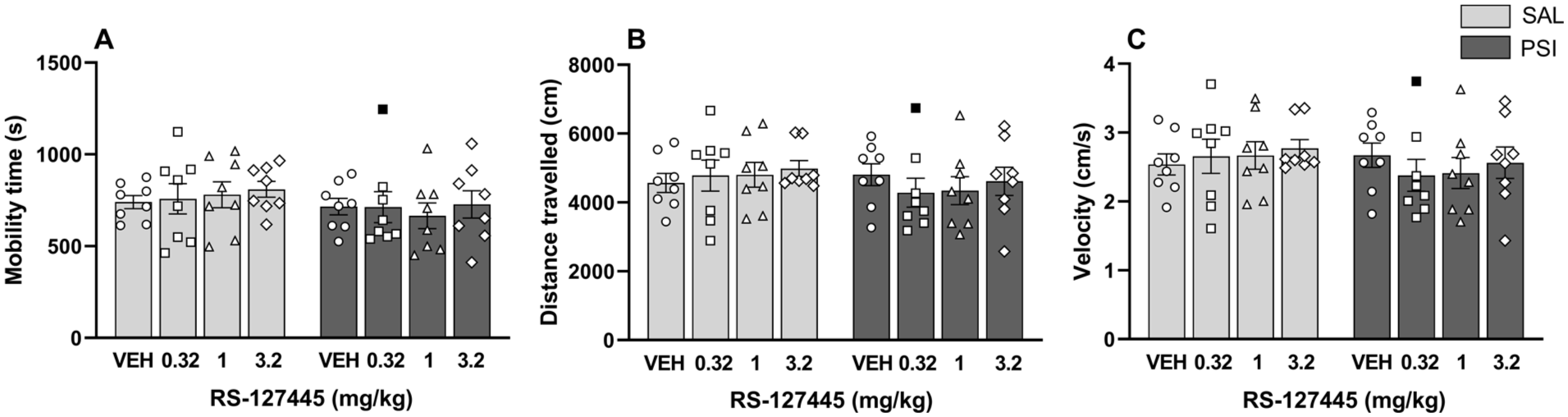
Psilocybin and RS-127445 did not alter motor activity in the OFT. Neither psilocybin (0.32 mg/kg) nor pretreatment with RS-127445 (0.32, 1.0, or 3.2 mg/kg) affected (a) mobility time (s), (b) distance traveled (cm), and (c) velocity (cm/s). Data are plotted as means ± SEM (n = 8 per group). The individual values (for each rat) are reported as open circles for vehicle, open squares for 0.32 mg/kg RS-127445, open triangles for 1.0 mg/kg RS-127445, and open diamonds for 3.2 mg/kg RS-127445. Outliers are depicted as black symbols. The raw data are reported in the Supplemental file (S1 Raw data).
RS-127445 did not block psilocybin-induced HTR
We investigated the effect of the 5-HT2BR antagonist, RS-127445, on psilocybin-induced HTR counts (Figure 4). A Kruskal–Wallis test revealed a significant overall effect of treatment on the number of HTR across the eight groups (H = 44.63, p < 0.0001). Mean ranks of HTR were 15.13, 11.64, 15.93, 21.25, 45.0, 42.50, 48.0, and 46.14 for the VEH-SAL, RS0.32-SAL, RS1.0-SAL, RS3.2-SAL, VEH-PSI, RS0.32-PSI, RS1.0-PSI, and RS3.2-PSI, respectively. Post hoc comparisons showed that all psilocybin groups, regardless of RS-127445 pretreatment dose, had significantly higher mean HTR ranks than the VEH-SAL group (p < 0.05, p < 0.05, p < 0.01, and p < 0.05, respectively). No significant differences were detected among the RS-127445 pretreatment doses within either the saline or psilocybin conditions.
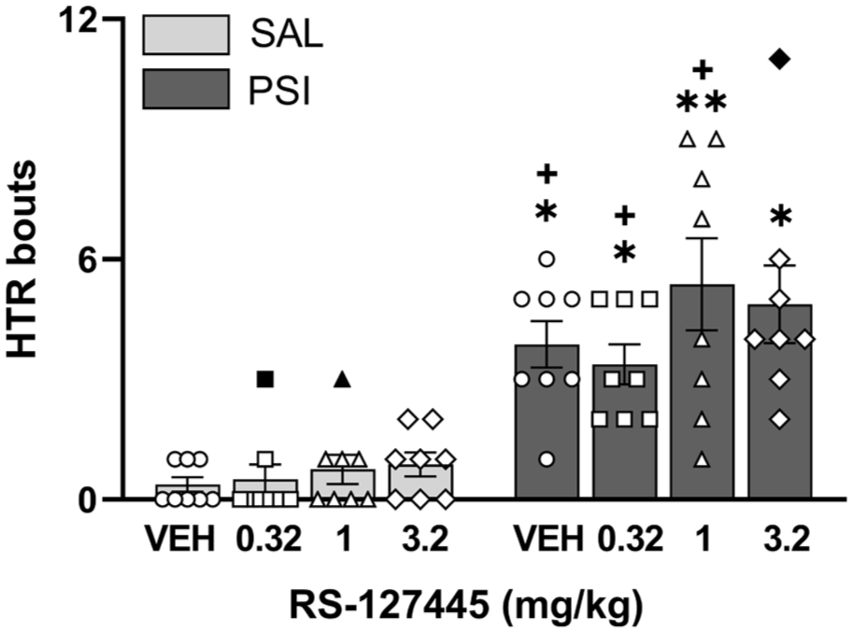
RS-127445 did not block psilocybin-induced HTR. Psilocybin (0.32 mg/kg) significantly increased HTR (expressed as the number of bouts) compared to saline-treated rats. This effect was not reversed by the 5-HT2BR antagonist RS-127445 (0.32, 1.0, or 3.2 mg/kg). Data are plotted as means ± SEM (n = 8 per group). The individual values (for each rat) are reported as open circles for vehicle, open squares for 0.32 mg/kg RS-127445, open triangles for 1.0 mg/kg RS-127445, and open diamonds for 3.2 mg/kg RS-127445. Outliers are depicted as black symbols. *p < 0.05, **p < 0.01 compared to saline-treated rats with vehicle pretreatment; +p < 0.05 compared to the respective saline-treated rats (i.e., the same pretreatment). The raw data are reported in the Supplemental file (S1 Raw data).
RS-127445 blocked both the rapid and the sustained psilocybin-induced behavioral effects in the FST
The impact of various doses of RS-127445 was assessed on both psilocybin-induced rapid (i.e., 1 day post-treatment; Figure 5(a)–(c)) and long-lasting (i.e., 21 days post-treatment; Figure 5(d)–(f)) behavioral effects in the FST. Due to the lack of interaction with three-way mixed ANOVA (Antagonist × Agonist × Day: F3, 52 = 1.10, p = 0.36 for immobility; F3, 51 = 1.25, p = 0.30 for climbing; F3, 54 = 0.40, p = 0.75 for swimming), indicating that the combined influence of Antagonist and Agonist did not differ across the two timepoints, the data were analyzed by two-way ANOVA separately for Days 1 and 21.
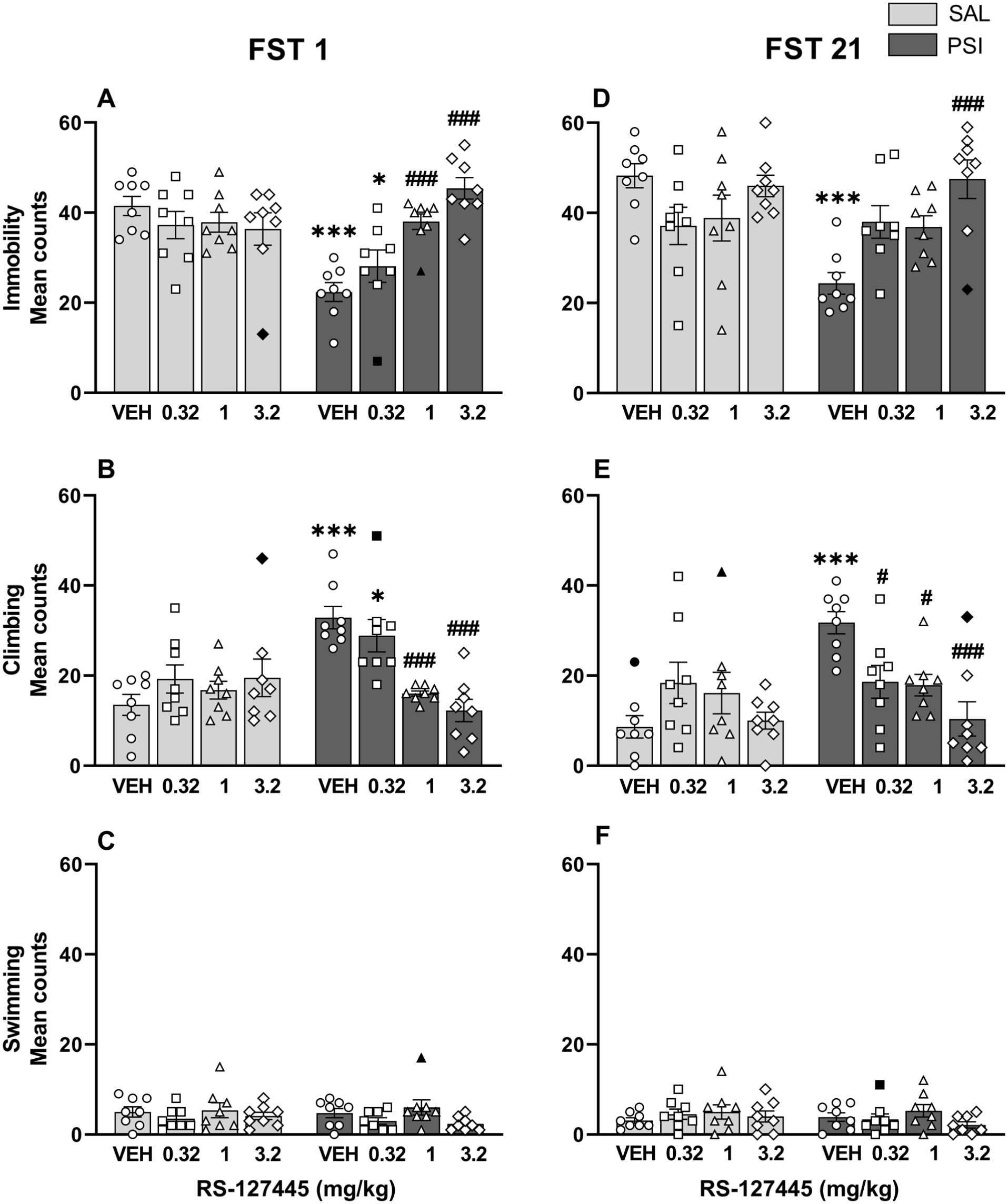
RS-127445 blocked both the rapid and sustained psilocybin-induced behavioral effects in the FST. Psilocybin produced both rapid (FST 1; a–c) and long-lasting (FST 21; d–f) decreases in immobility (a, d) and increases in climbing behavior (b, e). Pretreatment with RS-127445 dose-dependently reversed psilocybin-induced decrease in immobility (a, d) and increase in climbing (b, e) behaviors. No changes were observed in swimming behavior (c, f). Behavioral scores are presented as the number of 5-second intervals (bins) in which immobility, climbing, or swimming occurred during the 5-minute test. Data are plotted as means ± SEM (n = 8 per group). The individual values (for each rat) are reported as open circles for vehicle, open squares for 0.32 mg/kg RS-127445, open triangles for 1.0 mg/kg RS-127445, and open diamonds for 3.2 mg/kg RS-127445. Outliers are depicted as black symbols. *p < 0.05, ***p < 0.001 compared to saline-treated rats with vehicle pretreatment; #p < 0.05, ###p < 0.001 compared to psilocybin-treated rats with vehicle pretreatment. The raw data are reported in the Supplemental file (S1 Raw data).
As illustrated in Figure 5(a) and (b), psilocybin (0.32 mg/kg; vehicle pretreatment) produced rapid behavioral effects (FST 1) as shown by a decrease in immobility (Figure 5(a); p < 0.001) and an increase in climbing behavior (Figure 5(b); p < 0.001) compared to saline-treated animals with vehicle pretreatment. Both effects were dose-dependently reversed by pretreatment with RS-127445. Specifically, RS-127445 at 0.32 mg/kg did not block the psilocybin-induced effect in immobility or climbing; rats remained significantly less immobile (p < 0.05, Figure 5(a)) and exhibited more climbing behavior (p < 0.05, Figure 5(b)) than the saline-treated rats (vehicle pretreatment). At both 1.0 and 3.2 mg/kg, RS-127445 reversed psilocybin-induced reduction in immobility (Figure 5(a)) and enhancement of climbing behavior (Figure 5(b)). These observations were supported by the ANOVA, which revealed a main effect of Antagonist (Immobility: F3, 53 = 9.42, p < 0.0001; Climbing: F3, 54 = 8.05, p < 0.001), Agonist (Immobility: F1, 53 = 8.34, p < 0.01; Climbing: F1, 54 = 11.50, p < 0.01), and the interaction between the factors Agonist and Antagonist (Immobility: F3, 53 = 12.77, p < 0.0001; Climbing: F3, 54 = 10.44, p < 0.0001). No significant changes were observed in swimming behavior (Antagonist: F3, 55 = 1.83, p = 0.15; Agonist: F1, 55 = 1.52, p = 0.22; Antagonist × Agonist: F3, 55 = 0.23, p = 0.88; Figure 5(c)).
A similar pattern was observed for FST 21 (Figure 5(d) and (e)), with pretreatment with RS-127445 dose-dependently reversing the sustained behavioral effects of psilocybin, that is, reduced immobility (Figure 5(d); p < 0.001) and increased climbing behavior (Figure 5(e); p < 0.001). The ANOVA showed main effects of Antagonist (Immobility: F3, 55 = 5.50, p < 0.01; Climbing: F3, 53 = 5.33, p < 0.01), Agonist (Immobility: F1, 55 = 4.43, p < 0.05; Climbing: F1, 53 = 11.33, p < 0.01), and of the interaction between these factors (Immobility: F3, 55 = 7.41, p < 0.001; Climbing: F3, 53 = 8.96, p < 0.001). No significant changes were observed in swimming behavior (Antagonist: F3, 55 = 1.46, p = 0.24; Agonist: F1, 55 = 1.01, p = 0.32; Antagonist × Agonist: F3, 55 = 0.93, p = 0.43; Figure 5(f)).
Discussion
In this study, we examined the effect of pretreatment with the selective 5-HT2BR antagonist RS-127445 (0.32, 1.0, or 3.2 mg/kg) on both psilocybin (0.32 mg/kg)-induced HTR (a behavioral proxy of human psychedelic properties) and behavioral effects in the FST. Our findings confirmed that psilocybin increased HTR and produced a rapid and sustained decrease in immobility and an increase in climbing behavior in the FST. However, while RS-127445 pretreatment did not influence HTR counts, it dose-dependently (fully at the highest dose) blocked both the rapid and sustained psilocybin-induced reductions in immobility and increases in climbing behavior. Not only did this pharmacological dissociation add to the growing evidence that psilocybin can produce antidepressant-like effects independently of its 5-HT2A receptor activation (Hesselgrave et al., 2021; Sekssaoui et al., 2024), but it also revealed that 5-HT2BR agonism is required for psilocybin’s rapid and long-lasting predictive validity in the FST.
Over the past few years, psilocybin has emerged as a promising treatment for MDD. Unlike traditional antidepressants, which can take 4–6 weeks to show effects (Machado-Vieira et al., 2010), psilocybin demonstrated rapid antidepressant effects after a single administration in both preclinical (Kolasa et al., 2024; Seillier et al., 2025) and clinical settings (Goodwin et al., 2022, 2023). More importantly, the antidepressant effect of psilocybin has also been observed in patients with TRD (Agin-Liebes and Davis, 2021; Goodwin et al., 2022, 2023), making it a “breakthrough therapy” for TRD according to the U.S. Food and Drug Administration (Reiff et al., 2020). Remarkably, psilocybin’s therapeutic effects—unlike those of ketamine, another fast-acting antidepressant—can persist for at least 4–6 weeks (Carhart-Harris et al., 2016; Davis et al., 2021; Hibicke et al., 2020). Moreover, psilocybin may offer a more favorable safety profile than ketamine (Johnson et al., 2018; Morgan et al., 2012). However, it is not devoid of side effects beyond its hallucinogenic properties (Holze et al., 2021; Passie et al., 2002; Yerubandi et al., 2024). In a dose-dependent manner, psilocybin can induce physical discomfort, elevated blood pressure, nausea, headache, and dizziness, as well as psychiatric adverse events such as heightened anxiety and hallucinogen-persisting perception disorder (Perez et al., 2023; Yerubandi et al., 2024). Consistent with these clinical observations, our previous preclinical work showed that only the 0.32 mg/kg dose of psilocybin produced behavioral effects in the FST without the adverse physiological changes observed at higher doses (Seillier et al., 2025). In the current study, we confirmed that a single 0.32 mg/kg dose of psilocybin produced a robust decrease in immobility and an increase in climbing behavior in the FST at 24 hours and 21 days post-administration. Although some studies have reported no effect of psilocybin on immobility in control animals (Erkizia-Santamaría et al., 2025), our findings align with several preclinical reports demonstrating reduced immobility following psilocybin treatment (Kolasa et al., 2024; Sekssaoui et al., 2024). As discussed in our previous dose–response study (Seillier et al., 2025), such discrepancies may reflect the use of higher, behaviorally ineffective doses in studies reporting null results. However, an intriguing aspect of our findings is that psilocybin produced a climbing-dominant active coping profile without changes in swimming. According to the framework proposed by Detke et al. (1995), increased swimming is typically associated with serotonergic agents (e.g., SSRIs), whereas increased climbing is more characteristic of noradrenergic modulation. This counterintuitive pattern has, however, been reported by others. Across several recent studies, psilocybin has consistently been shown to increase climbing behavior, with some reports also noting concurrent increases in swimming (García-Cabrerizo et al., 2025; Hibicke et al., 2020; Kolasa et al., 2024; Seillier et al., 2025; Wang et al., 2025). Interestingly, the opposite pattern may occur in mice, where psilocybin has been reported to increase swimming rather than climbing (Erkizia-Santamaría et al., 2025), suggesting that species-specific differences may shape the precise active coping profile observed. Notably, this climbing-dominant pattern raises the possibility that, at this intermediate dose, psilocybin preferentially engages downstream noradrenergic circuits. This interpretation is supported by recent work showing that psilocin modulates extracellular noradrenaline (NA) in a non-linear, dose-dependent manner (Kościuk et al., 2026). Specifically, lower concentrations produced larger NA increases than higher concentrations, and these increases exceeded those produced by the 5-HT2A agonist 25I-NBOMe, which did not significantly alter NA levels relative to baseline. Together, these findings suggest that psilocybin/psilocin recruit neuromodulatory systems differently from prototypical 5-HT2A agonists and that small dose changes can shift which downstream systems are engaged. In line with our prior observations of a narrow therapeutic window, an intermediate dose may preferentially activate circuits that bias active coping toward climbing rather than swimming.
The long-standing assumption that 5-HT2AR activation is essential for the therapeutic benefits of psychedelics may have hindered the identification of other receptor systems involved in psilocybin’s antidepressant action. Psilocybin is rapidly dephosphorylated after oral administration, forming psilocin in the acidic environment of the stomach or by alkaline phosphatase in the intestine and kidney. Importantly, psilocin is a high-affinity ([Ki] = 49 nM) ligand with moderate intrinsic efficacy at the 5-HT2BR (Rudin et al., 2026), thus often described as a “partial agonist” at these receptors. It is also worth stressing that, while psilocybin per se has a very low affinity for most 5-HT receptors, including 5-HT2AR ([Ki] < 10,000 nM; as opposed to psilocin: [Ki] = 339.6 nM), it has a moderate affinity for 5-HT2BR ([Ki] = 99.6 nM; (Ling et al., 2022) and can act as a weak 5-HT2BR agonist comparably to psilocin (Sard et al., 2005). In the present study, blockade of 5-HT2BRs left the acute, 5-HT2A-mediated HTR intact, yet eliminated psilocybin’s effects on immobility and climbing in the FST. This pattern indicates that 5-HT2BR activation is required for the FST behavioral effects of psilocybin. Remarkably, this 5-HT2BR involvement is not unprecedented. A similar dependence on 5-HT2BR signaling has been reported for classical antidepressants; notably, fluoxetine fails to produce behavioral responses in the FST in 5-HT2B-knockout mice, despite being effective in wild-type animals (Diaz and Maroteaux, 2011). Importantly, the 5-HT2BR agonist BW723C86 has been reported to decrease immobility in the FST in wild-type control mice but not 5-HT2BR knockout (Diaz and Maroteaux, 2011), suggesting that 5-HT2B activation may be sufficient—at least for rapid behavioral effects measured acutely. However, in their study, Diaz and Maroteaux administered the drug 30 minutes before the test (unlike our protocol, in which psilocybin was administered the day before or 20 days before the test). Whether 5-HT2BR agonism per se proves sufficient to mimic both the rapid and sustained antidepressant-like effects of psilocybin warrants further exploration, especially given the polypharmacology of this compound. Building on these observations, and while limited to the FST in normal rats, our data identify a non-5-HT2A pathway—mediated by 5-HT2BRs—that contributes to rapid and sustained behavioral effects in the FST and highlight the importance of considering multiple serotonergic targets and dose-dependent receptor recruitment when developing psychedelic-inspired therapeutics.
One plausible locus for this 5-HT2BR-mediated effect is non-neuronal glial cells—astrocytes and microglia—which express 5-HT2BRs and are central regulators of neuroimmune function implicated in MDD pathophysiology (Deng et al., 2020). Emerging evidence links psilocybin/psilocin to both immunomodulation and neuroplasticity. For example, psilocybin reduced circulating tumor necrosis factor-alpha (TNF-α) in healthy volunteers (Mason et al., 2023), and in vitro work showed that psilocybin/psilocin suppresses bacterial lipopolysaccharide (LPS)-induced TNF-α expression in microglia in a 5-HT2B-dependent manner while potentiating LPS-induced brain-derived neurotrophic factor (BDNF) expression (Laabi et al., 2025). Notably, the BDNF effect was sensitive to 5-HT2A antagonism rather than 5-HT2B blockade, consistent with prior data linking the role of 5HT2ARs in BDNF regulation (Vaidya et al., 1997) and our previous observations that psilocybin-induced increases in BDNF levels in both the hippocampus and prefrontal cortex were dissociated from its behavioral effects (Seillier et al., 2025). Separately, psilocybin promotes synaptic plasticity in the prefrontal cortex and hippocampus (Jefsen et al., 2021; Shao et al., 2021; Zhao et al., 2024), and microglia are key mediators of synaptic remodeling (Deng et al., 2020). In animal models of depression, astrocytic 5-HT2BR expression is reduced and is restored by chronic fluoxetine (Dong et al., 2015), further linking glial 5-HT2B signaling to antidepressant mechanisms.
These insights, together with the potential contribution of astrocytic 5-HT2BR to the pathophysiology of MDD (Verkhratsky et al., 2021), indicate that non-neuronal 5-HT2BR may play a more significant role than previously thought in the pharmacological mechanism of classic antidepressants (Hertz et al., 2012), but also psilocybin.
Conclusion
Overall, our results identify a critical and previously underappreciated role for 5-HT2BRs in mediating the rapid and sustained predictive validity of psilocybin for antidepressant effects in the FST in otherwise healthy Wistar rats. Pretreatment with the selective 5-HT2B antagonist RS-127445 produced a complete abolition of psilocybin’s reductions in immobility and increases in climbing, strongly indicating that 5-HT2B signaling is necessary for these behavioral outcomes. Although necessity does not imply sufficiency, these data emphasize the importance of non-5-HT2A targets and implicate 5-HT2BRs as key contributors to psilocybin’s efficacy in this paradigm.
Crucially, we observed a pharmacological dissociation between the acute HTR and the rapid, enduring behavioral effects in the FST, suggesting that distinct receptor pathways underlie psilocybin’s psychedelic and therapeutic actions. This dissociation raises the prospect of developing therapeutics that retain antidepressant efficacy while minimizing psychedelic effects by selectively targeting 5-HT2BR-linked mechanisms. More broadly, our findings contribute to a more nuanced understanding of psilocybin’s polypharmacology and highlight the importance of multi-receptor approaches in elucidating its mechanisms of action.
Supplemental Material
sj-xlsx-1-jop-10.1177_02698811261458349 – Supplemental material for Blocking 5-HT2B receptors abolishes psilocybin’s efficacy in the rat forced swim test
Supplemental material, sj-xlsx-1-jop-10.1177_02698811261458349 for Blocking 5-HT2B receptors abolishes psilocybin’s efficacy in the rat forced swim test by Lenka Seillier, Alexandre Seillier, Morgan A. Zvolska and Romana šlamberová in Journal of Psychopharmacology
Footnotes
Acknowledgements
We thank Zuzana Ježdíková, who assisted with in vivo experiments. Figure 1 was created in BioRender (![]() ) under the agreement number CJ29LQUX5P.
) under the agreement number CJ29LQUX5P.
Ethical considerations
All experiments involving animals were conducted with the approval of the Committee for Animal Care and Use of the Third Faculty of Medicine, Charles University, Prague, the Ministry of Education, Youth and Sports of the Czech Republic (protocol code: MSMT-25258/2025-4), and the Directive 2010/63/EU for the protection of animals used for scientific purposes (European Parliament and Council of the European Union; as amended on June 9, 2019).
Author contributions
LS and AS conceptualized and designed the study. LS conducted the investigations, collected the data, and conducted the statistical analysis. RŠ acquired funding for the experiment. LS and MZ wrote the first draft of the manuscript, and AS and RŠ revised it. All authors revised and contributed to the consecutive versions of the manuscripts and approved the final manuscript.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the research program Cooperatio Neurosciences from Charles University.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
All relevant data are within the article and its Supplemental file.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.