
Abstract
MicroRNAs (miRNA) are endogenously produced small, non-coded, single-stranded RNAs. Due to their involvement in various cellular processes and cross-communication with extracellular components, miRNAs are often coined the “grand managers” of the cell. miRNAs are frequently involved in upregulation as well as downregulation of specific gene expression and thus, are often found to play a vital role in the pathogenesis of multiple diseases. Central nervous system (CNS) diseases prove fatal due to the intricate nature of both their development and the methods used for treatment. A considerable amount of ongoing research aims to delineate the complex relationships between miRNAs and different diseases, including each of the neurological disorders discussed in the present review. Ongoing research suggests that specific miRNAs can play either a pathologic or restorative and/or protective role in various CNS diseases. Understanding how these miRNAs are involved in various regulatory processes of CNS such as neuroinflammation, neurovasculature, immune response, blood-brain barrier (BBB) integrity and angiogenesis is of empirical importance for developing effective therapies. Here in this review, we summarized the current state of knowledge of miRNAs and their roles in CNS diseases along with a focus on their association with neuroinflammation, innate immunity, neurovascular function and BBB.
Keywords
Introduction
MicroRNAs (miRNAs, miRs) are endogenously produced small, non-coded, single-stranded RNAs which are about 19–24 nucleotides long. They can be referred to as ‘grand managers’ because of their involvement in various cellular processes, and cross-communication with extracellular components along with different cell types.1,2 The role of miRNAs in regulating protein-coding genes has been annotated in human genome studies. 3 Though the multifaceted role of miRNAs in different diseases is not yet fully explored, the recent discoveries of new miRNAs and their roles attract much attention.
Neurological or central nervous system (CNS) disorders possess significant global health concerns. CNS diseases can emerge from a multifaceted range of factors, including neurodegenerative processes, ischemic events, hemorrhages, and inflammatory responses. When the CNS is prone to any infectious or abnormal conditions, resident glial immune cells induce an inflammatory response via chemokines, pro-inflammatory cytokines, and reactive oxygen species. This induced neuroinflammation promotes tissue repair processes and protects against pathogens and neurotoxins or may lead to apoptosis and cytotoxicity of the target brain cells.
There has been a considerable amount of research on the role of miRNA in CNS-related diseases. Increasing evidence underscores the crucial role of miRNAs in governing both the injury/damage and repair/recovery process of CNS diseases. They are approached as biomarkers for diseases 4 and potential therapeutic interventions in various CNS diseases. 5 The regulatory role of miRNA in innate immunity leading to the involvement in neuroinflammation is vastly studied.6,7 In the present review, we summarized the current findings regarding miRNA influence on the systemic functioning of the CNS including innate immune response and neuroinflammation, neurovascular integrity and angiogenesis, and blood-brain-barrier (BBB), as well as the role of miRNA in specific CNS diseases such as stroke, vascular dementia, Alzheimer’s disease, Parkinson’s disease, traumatic brain injury, and brain tumors.
MicroRNA
miRNA history
The discovery of the first miRNA, lin-4, in 1993 can be marked as an evolutionary era of molecular biology. Lee and colleagues discovered the mechanism and importance of transcript gene lin-4 in the downregulation of lin-14 protein for initiation of progression of the first larval stage (L1) to L2 of the nematode Caenorhabditis elegans.8,9 Later a collaborative study between Lee and Wightman group uncovered the pairing of complementary regions of the 3’untranslated region (3’UTR) of lin-4 gene and lin-14 messenger RNA (mRNA), leading to translational repression of lin-14. 10 More extensive studies using the basis of this research discovered homologs of this gene later in humans. 11 Various studies following these findings have contributed to the concept of miRNA, which continues to grow.
miRNA biogenesis
The biogenesis of miRNAs is a multi-step process that initiates from the transcription of genes via RNA polymerase-II into typical hair-pin loop-shaped primary miRNAs. This structure is recognized and converted into percussor miRNA by microprocessor complex subunit – DiGeorge syndrome critical region 8 (DGCR8) protein and Drosha ribonuclease-III enzyme by forming a microprocessor complex. The precursor miRNA is then transported from the nucleus to the cytoplasm via Exportin-5 and then converted into double-stranded miRNAs by the DICER enzyme. Argonaute protein AGO-2 unwinds the miRNA to form an RNA-indued silencing complex (RISC) which can cause post-translational suppression or transcript degradation of mRNA by attaching to 3′UTR of target genes. 12
miRNA function/mechanism
The mechanism of action of miRNA begins by binding of miRNA sequence with 3′UTR of mRNA, which leads to repression of mRNA translation. On the contrary, an increase in translation can occur due to the binding of miRNA sequence to 5′UTR of mRNA, 13 proving the binding-specific action of miRNA. Moreover, an interaction between miRNA and protein can occur via the proteic receptor, also called a miRceptor. 14
miRNA in innate immune response and neuroinflammation
Innate immunity can be termed ‘immunity by birth’. Monocytes, dendritic cells, macrophages, natural killer cells, and granulocytes are the various cells of the innate immune system working as a first line of defense. The influence of diverse miRNAs and their function in governing these cells underscores the foundational importance of miRNAs in innate immunity regulation. The imbalance in levels and function of miRNA leads to neuro-inflammation. 7 There has been proof of the involvement of miRNA in various steps of the innate immune network such as feedback regulation of immune homeostasis, expression of costimulatory and adhesion molecules, and mainly the release of cytokines/chemokines. 15 miR-223 is proven to have a regulatory role in myeloid differentiation, neutrophil and macrophage functions. 16 One of the studies found that deletion of the Dicer-1 protein used in miRNA biogenesis affected the miRNA levels in macrophages, causing downregulation in inflammatory cytokines upon innate immunity activation. 17 This identifies a link between miRNAs, immune responses and inflammation.
Research shows that there exist pro-inflammatory and anti-inflammatory miRNAs, suggesting these transcription regulators can be either pathological or protective in the setting of CNS dysfunction. As an anti-inflammatory regulator, miR-193b-3p inhibits histone deacetylase 3 expression and thus reduces inflammation. 18 miR-23a, miR-23b, and miR-19 act on inhibitors of nuclear factor-κB (IκB) kinase-alpha (IKK-α), TAB2, TAB3, and nuclear factor-kappa beta (NF-κB) immune genes to suppress the inflammatory activity.19,20 miRNAs have been demonstrated to be strict modulators of toll-like receptor (TLR) expression and thus can increase or decrease immune response. For example, Let-7b and miR-146b act on TLR4 to induce anti-inflammatory activity stimulated by innate immunity.21,22 miR-93 has been shown to be one such miRNA that upregulates the expression of TLR2 and TLR4, leading to an increase in neuroinflammation. miR-181c is another miRNA that functions on TLR4. 23 By inhibiting this TLR, miR-181c has been shown to hinder the production of inflammation. miR-181c-3p also serves as a post-infarct anti-inflammatory regulator. 14 In addition to reducing damaging inflammation, miR-181c also upregulates the expression of tripartite motif 2 (TRIM2) in the setting of cerebral hypoxia. TRIM2, in turn, prevents the ubiquitination and subsequent destruction of neurofilament light (NF-L). NF-L is a significant contributor to neural remodeling and dendritic branching. Through this complex pathway, miR-181c protects from continually escalating cognitive impairment by promoting neuroplasticity.
Macrophages are types of white blood cells involved in causing immune responses in the CNS. Microglia polarization from phenotype M1 (pro-inflammatory) to phenotype M2 (anti-inflammatory) is advantageous as it leads to an anti-inflammatory effect 24 and a neuroprotective effect after ischemic insult. 25 Several miRNAs play a positive or negative role in the polarization of macrophages in turn contributing to and regulating the production of pro-inflammatory cytokines. 15 For example, miR-155, and miR-9 regulate M1 macrophage which has a pro-inflammatory effect15,26 whereas, miR-21 and miR-146 regulate M2 macrophage which has an anti-inflammatory effect. 15 MiR-155 has been implicated as a significant contributor to neuroinflammation due to its additional functions in the regulation of cytokines production via macrophage coordination. 27 Recent research has also shown that miR-3473b acts as an anti-inflammatory at the beginning of stroke and as another pro-inflammatory with stroke progression by exaggerating neuroinflammation. 28 At the beginning of the stroke, miR-3473b suppresses macrophage-mediated inflammation. As the stroke insult progression continues, miR-3473b negatively regulates the suppressor of cytokine signaling 3 to induce the progression of stroke via pro-inflammatory factors such as tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6). This duality in the functioning of miRNAs as pro- or anti-inflammatory in the context of CNS disorders is portrayed as a double-edged sword when it comes to their correlation with neuroinflammation.
Role of miRNAs in modulating neurovascular unit integrity and angiogenesis
Neurovasculature refers to the network of blood vessels that supply nutrients and oxygen to the brain and spinal cord. The neurovascular unit (NVU) is an important target of neurovasculature and it is comprised of neurons, astrocytes, microglia, pericytes, myocytes, endothelial cells, and blood-spinal cord barrier. 29 The miRNAs released by several components of the NVU have been observed to maintain NVU integrity by participating in angiogenesis, modulation of neurotropic factors (NTFs), and neuronal coupling. 30
In the context of angiogenesis and vascular integrity, miR-126 is found to regulate the response of endothelial cells to vascular endothelial growth factor (VEGF) by repressing negative regulators of the VEGF pathway, including the Sprouty-related protein, SPRED1, and phosphoinositol-3 kinase regulatory subunit 2 (PIK3R2). 31 Current research suggests that endothelial-derived miRNA, miR-126-3p may upregulate the growth of neurites, directly contributing to neuroplasticity. 35 miRNAs released by NVUs also play a role in the regulation of cellular function in neurovasculature. For example, miR-181c-3p could directly downregulate platelet-activating factor receptors and thus, hinders microglial apoptosis. 32 An astrocyte-derived miRNA, miR-190b, downregulates neuronal death via autophagy-related gene-7, and further neuroprotection is offered by microglial-derived miRNA. 33 miR-124, expressed in the penumbra post-infarction, actively inhibits the accumulation of astrocytes through the downregulation of signal transducer and activator of transcription 3 (STAT3), thus reducing glial scar development and improving post-stroke outcomes. 34 Furthermore, miR-26a was also implicated in influencing the expression of hypoxia-inducible factor-1α and VEGF via the PI3K/AKT pathway, further bolstering its role in angiogenesis. miRNAs like – miR-126, miR-137, miR-145, and miR-493 are found to modulate neurovascular integrity and function such as apoptosis, neuroinflammation, neurogenesis, and angiogenesis through regulation of mitogen-activated protein kinase (MAPK) signaling pathway. 37
The NTFs are endogenous peptides or small protein biomolecules mainly belonging to three families: glial cell line-derived neurotrophic factor (GDNF) family ligands, neuropoietic cytokines, and neurotrophins. NTFs such as brain-derived neurotrophic factor (BDNF), neurotrophin 3 (NT-3), neurotrophin 4 (NT-4), and nerve growth factor (NGF) are responsible for deciding the fate of the neuron by regulating neurite survival and differentiation, synaptic plasticity and neurotransmitter synthesis. 38 miRNAs are found to cause modulation of different NTFs and hence affect the NVU integrity to control neurovasculature function. For instance, miR-15a, -26a/b, -134, -206, and -210 are found to directly regulate BDNF activity and expression. 39 miR-132 and -182 are found to downregulate BDNF protein levels via activating cAMP-response element binding protein transcriptional factors in human neuronal cell models and depression patients. 40 Whereas miR-155 has been found to increase NGF protein expression and its downregulation inhibits cytokine signaling-1 expression and NF-κB activation in myocardial infraction mice model. 41 miR-21 downregulates NT3 mRNA levels 42 and miR-17/20 43 downregulates NT4, while -21-5p upregulates NT4. 44 Apart from this, miR-144 downregulates mesencephalic astrocyte-derived neurotrophic factor, and miR-134 and miR-141 downregulate cerebral dopamine neurotrophic factor to cause neurodegeneration in Parkinson’s disease. 45 Hence, miRNAs affect NTFs which in turn have a role in neurovasculature and CNS disease progression.
Role of miRNA in blood-brain barrier
The BBB is a highly specialized and selective barrier that separates the bloodstream from the brain's extracellular fluid, regulating the passage of molecules and cells into and out of the central nervous system. BBB disruption-induced increased permeability is the major factor for brain edema after brain injury in CNS diseases. Loss of BBB integrity can also expose neuronal tissue to circulation-derived inflammatory cytokines and other injurious molecules. Therefore, modulating BBB function and integrity is a very important pathway to change the disease progress and outcome. 46 One of the pivotal roles of miRNA in regulating neurovascular function is via the maintenance of the integrity of the BBB.47,48 miRNA can target endothelial junctions and tight junctions to affect the integrity of BBB positively or negatively. For example, miR-132 decreases the degradation of tight junction proteins vascular endothelial-cadherin and β-Catenin to offer BBB protection in ischemic stroke mice via inhibiting the matrix metallopeptidase (MMP) 9 gene. 49 miR-132 in neurons blocks eukaryotic elongation factor-2 (eEF2K) expression which upregulates the cadherin-5 (Cdh5) expression in endothelial cells of BBB promoting the brain vascular integrity. 50 Whereas miR-210 disrupts junction proteins such as occludin and β-catenin, thereby damaging the integrity of BBB. Hence, inhibition of mR-210 significantly decreases the BBB damage. 51 Similarly, inhibition of miR-15a/16-1, 52 and miR-182 53 can restore the integrity of BBB by rescuing Zonula Occludens-1 expression. Inflammatory cytokines play an important role in mediating the disruption of BBB and increasing BBB permeability. Targeting inflammation is one of the roles of miRNA in modulating BBB function, such as miR-21-5p maintains the integrity of BBB by decreasing the inflammation via downregulating proinflammatory cytokines such as TNF-α and IL-6. 54 Endothelial cells, as one of the key cellular components of the BBB, are also regulated by miRNAs. miR-182 inhibition attenuates apoptosis of endothelial cells in BBB via mammalian target of the rapamycin (mTOR)/forkhead box O (FOXO1) pathway, 53 while miR-23 stimulates endothelial cell survival by inhibiting inflammatory factors like TNF-α, IL-6, and NF-κB. 55 Inhibition of miR-181c has been reported to increase BBB integrity via upregulation of phosphoinositide-dependent kinases-1 maintaining actin cytoskeleton. 56 Besides endothelial cells, pericytes play a vital role in maintaining BBB homeostasis in CNS. 57 miRNAs like miR-27b, and miR-150 can modulate the functioning of pericytes by targeting semaphorin 6 A/D and endothelial receptor tyrosine kinase Tie-2 respectively, leading to the promotion of interaction of endothelial cells with pericytes and leading to inhibition of claudin-5 expression and endothelial cell survival respectively.58,59 Thus, miRNAs play a role in CNS homeostasis by maintaining BBB integrity and modulating BBB function.
miRNAs in the diseases of CNS
CNS diseases comprise of a broad spectrum of disorders that affect the brain and spinal cord. CNS diseases often result from complex interactions between genetic predispositions, environmental factors, and molecular dysregulations within the nervous system. miRNAs have emerged as key regulators of various physiological processes within the CNS, including neuronal development, neurovasculature and neuroinflammation. Figure 1 illustrates detailed information regarding miRNA biogenesis along with their cellular targets and pathways involved in the regulation of various CNS diseases. Dysregulation of miRNA expression has been implicated in the pathogenesis, diagnostic markers and treatment of CNS diseases. The most common CNS diseases are discussed as follows –
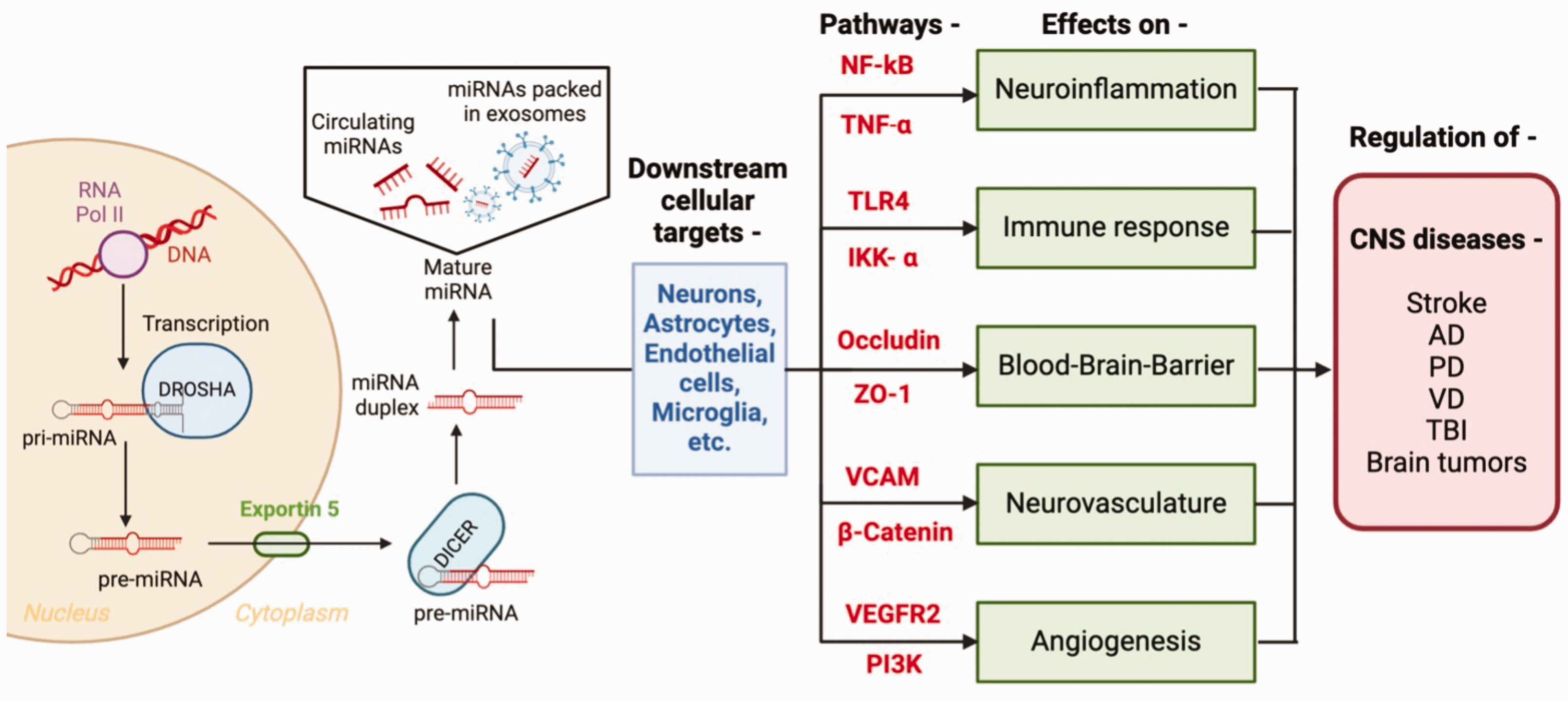
The biogenesis, downstream cellular targets and targeted pathways of miRNAs in stroke, Alzheimer’s Disease (AD), Parkinson’s Disease (PD), Vascular Dementia (VD), Traumatic Brain Injury (TBI) and brain tumor. The biogenesis of miRNA begins with gene transcription by RNA polymerase-II, forming hairpin-shaped primary miRNA. It is processed into precursor miRNA by Drosha enzyme. Precursor miRNA is transported to the cytoplasm via Exportin-5 and converted into double-stranded pre-miRNA by DICER enzyme. Pre-miRNA (circulating or packed in EXs) gets converted to mature miRNA and acts on various cells. In CNS, miRNAs act on cellular targets such as neurons, astrocytes, endothelial cells, and microglial cells to affect neuroinflammation, immune response, BBB, neurovasculature and angiogenesis via various pathways such as NF-kB, TNF-a, TLR4, IKK-a, occluding, ZO1, VCAM, b-catenin, VEGFR2 and PI3K. With these actions, miRNAs have a regulatory role in CNS diseases such as stroke, AD, PD, VD, TBI, and brain tumors.
Stroke
Stoke is one of the current leading causes of death worldwide with high morbidities. Amongst the two types of strokes, ischemic stroke is more prevalent than hemorrhagic stroke. Ischemic stroke occurs when the brain is deprived of blood due to artery blockage. On the other hand, hemorrhagic stroke is when the blood vessels are ruptured inside the brain. Both of these types lead to significant disability as it is known to cause the death of all types of brain cells such as neurons, astrocytes, pericytes, microglia, endothelial cells, etc. Stroke pathophysiology is a complex and interconnected process, culminating in brain cell death through a cascade of events. The planning of stroke recovery involves a complex interplay of processes, such as angiogenesis, neurogenesis, synaptogenesis, and neuronal and synaptic plasticity. These dynamic mechanisms collaborate to promote the brain's healing and functional rehabilitation after a stroke. 60 Numerous miRNAs have been studied and consequently implicated in the pathogenesis or repair of cerebral ischemia through a variety of functions. miR-191 has been hypothesized to contribute to the pathogenesis of stroke via its inhibition of vascular endothelial zinc finger 1-induced angiogenesis to promote ischemic brain injury. 61 Conversely, miR-21 has been shown to have a pro-angiogenic effect via activation of AKT and extracellular signal-regulated kinase (ERK) pathway. 62 miR-182 has been shown to inhibit the mTOR/FOXO1 pathway, downregulating apoptosis and maintaining BBB integrity following stroke. 53 miR-3552 has been shown to regulate well-established stroke biomarker Caspase-3 which functions to promote cell apoptosis in the setting of stroke. 63 miR-497 is another miRNA contributing to stroke pathology via selectively targeting and downregulating antiapoptotic genes B cell lymphoma-2 (Bcl-2) and Bcl-w. 64 miR-223 provides a protective role following reperfusion of ischemic tissue by downregulating the expression of glutamate receptors – Glutamate receptor 2 and NR2B. 65 Some miRNAs protect the integrity of BBB by reducing endothelial cell apoptosis through different pathways. For example, miR-126-3p downregulates vascular cell adhesion molecule-1 to attenuate intracranial stroke-induced BBB disruption. 66 miRNAs also play a crucial regulatory role in the process of post-stroke recovery, such as angiogenesis. 67 For example, miR-21 62 and miR-31 68 have pro-angiogenic effects. Expression of miR-26a has also been shown to be upregulated in the context of cerebral infarct. 36 miR-26a expression was correlated to endothelial lumen proliferation, suggesting a role in post-stroke angiogenesis. In astrocytes, miRNAs like miR-29a, miR-29b, miR-130a, and miR-210 regulate apoptosis, and angiogenesis and exert anti-inflammatory effects. 69 miRNAs such as miR-126a, miR-297, miR-140, and miR-17-5p in pericytes act as proangiogenic and also maintain BBB stability. 69 There is an involvement of miRNAs such as miR-124 in the production of IL-10, miR-106, miR-132, and miR-181c in mediating the anti-inflammatory effect via microglial cells. 69 In summary, the role of multiple miRNAs in the regulation of stroke pathogenesis calls for their implementation in treatment strategies.
Alzheimer’s disease
Alzheimer's disease (AD) is a neurodegenerative disorder recognized as the most common cause of dementia which typically transpires with ageing, leading to memory impairment and cognitive decline. The pathogenesis of AD is characterized by the buildup of amyloid β and tau proteins, along with neuroinflammation, which ultimately leads to the degeneration and demise of brain cells. The current therapies for AD include the use of Acetylcholinesterase enzyme (AChE) inhibitors and N-methyl-D-aspartate (NMDA) inhibitors. Some future therapies are currently under study, such as immunotherapy, anti-amyloid, and anti-tau therapy for reduction, prevention, and clearance of amyloid β and tau protein, respectively. 70 miRNAs are wildly used as very efficient biomarkers of AD, such as – miR-26b, miR-30e, miR-34a, miR-34c, miR-107, miR-125b, miR-146a, miR-151, miR-200c, miR-210, and hsa-miR-485 have been hypothesized to predict the early onset of AD nearly 20 years before the onset of disease. 71 Several miRNAs have been investigated as significant regulators of Aβ and tau expression and metabolism. For example, miR-210 is found to dysregulate the synaptic functioning in AD to cause cognitive impairment through the synaptosomal-associated protein of 25 KDa (Snap25). 72 miR-124-3p is found to cause attenuation of apoptosis of neuroblastoma cells in vitro through inhibition of abnormal hyperphosphorylation of Tau protein by directly targeting caveolin-1. 73 miR-339-5p 74 has been hypothesized to be a modulator of Aβ metabolism. Current research suggests miR-339-5p downregulates the expression of β-site APP-cleaving enzyme 1 (BACE1), an enzyme that plays a crucial role in cleaving the initial Aβ precursor protein into insoluble Aβ filaments. As expected, researchers discovered miR-339-5p was decreased in AD brain specimens along with corresponding high BACE1 protein levels. Researchers fortified this proposed relationship by demonstrating that miR-339-5p transfection in primary human brain cultures resulted in declining BACE1 levels. While more research is required to further elucidate this mechanism, it is worth noticing that the pathogenesis of AD likely hinders and/or downregulates this miRNA early in the disease process to accomplish the characteristic accumulation of Aβ filaments. Additionally, restoring miR-339-5p levels in AD patients may also present promise in slowing the accumulation of Aβ filaments and thus progression of cognitive impairment. Further investigations are needed to thoroughly explore and grasp the functions of diverse miRNAs, given their potential as biomarkers to address their potential as therapeutic agents for AD.
Parkinson’s disease
Parkinson's disease (PD) stands as the second most widespread neurodegenerative disorder globally, following closely behind AD. It is characterized by the progressive degeneration of dopaminergic neurons in the substantia nigra pars compacta, accompanied by the formation of Lewy bodies, which consist mainly of misfolded α-synuclein. The current treatment for PD involves various pharmacological therapies such as anticholinergics, anti-glutamatergic, monoamine oxidase inhibitors, dopamine agonists, catechol-o-methyl transferase inhibitors, and adenosine A2 receptor antagonists. 75 The role of miRNAs in PD is studied in diagnosis as biomarkers as well as regulators. Amongst various studied miRNAs, miR-30, miR-29, let-7, miR-485, and miR-26 have been implicated in PD pathogenesis. 76 miRNAs - let-7, miR-128, miR-433, miR-485-5p, miR-132, and miR-212 were found to be dysregulated in cerebrospinal fluid (CSF) of PD patients hence proving their function as a biomarker and their potential in the therapy. 77 miR-124 have been proven to modulate several pathways in PD such as – NF-κB, STAT3, calpain 1/p25/cyclin-dependent kinases 5 (CDK5), ERK-mediated, 5′ adenosine monophosphate-activated protein kinase (AMPK), and Bcl-2-interacting mediator of cell death (Bim) pathways to regulate neuroinflammation, apoptosis, autophagy, oxidative damage, mitochondrial dysfunction and cell survival.78,79 miR-26a is found to act on death-associated protein kinase 1 (DAPK1) which is a widely distributed serine/threonine kinase to formulate cellular and molecular pathologies of PD. 80 In one of the studies of the rotenone-induced PD rat model, miR-211-5p was found to augment autophagy and relieve endoplasmic reticulum stress by affecting Beclin-1, catalase, IL-1β, α-synuclein, E74-like factor 2/new Ets-related factor (ELF2/NERF) and protein kinase RNA-like ER kinase (PERK)/eukaryotic initiation factor 2-alpha (eIF2α)/C/EBP homologous protein (CHOP) pathways. 81 Immune cells such as macrophage and microglia play a regulatory role in PD as they can be either pro- or anti-inflammatory depending on the polarization state. miRNAs are found to modulate the immune cells in PD. miR-132-3p is found to regulate the population of microglial cells via glutaredoxin (GLRX) to regulate dopaminergic neurodegradation and neuroinflammation in PD. 82 miR-150-5p is found to be a target of rhabdomyosarcoma 2-associated transcript (RMST), a key regulator of neural stem cells to regulate the release of inflammatory cytokines and neuronal apoptosis which is developed in PD. 83
Vascular dementia
Vascular dementia (VD) stands out as one of the most prevalent causes of dementia. Hence, early diagnosis of VD assumes paramount significance in averting disease progression and vascular cognitive impairment. In VD, myelinated brain tissue is frequently impacted, and the primary pathologic manifestation involves the calcification of arterioles. The molecular mechanism behind the adverse effects of VD is oxidative stress, neuroinflammation, and compromised BBB integrity, mainly followed by endoplasmic reticulum stress, changes in homeostasis of cholesterol, and various lipoproteins. 84 The current treatment for managing VD includes managing risk factors, non-pharmacological treatment for preventing the progression of the disease, and therapeutics such as anti-dementia and cerebrovascular problems. 85 The biomarker and regulatory roles of miRNAs in VD is recently studied. In VD patients as compared to healthy human controls, circulating miRNAs such as miR- 376a-3p and -409-3p were decreased, and miR-363-3p, -32-5p, -451a, -486-5p, and -502-3p were increased. 86 Apart from the diagnostic function of miRNAs in VD, they are studied to have a regulatory role via different mechanisms. For example, miR-26b protects the brain from dementia by alleviating microglia inflammatory response and targeting IL-6. 87 Zhang et. al and team have combined the data for various functions of miRNAs in VD and the use of various miRNA mimics and antagomirs in treating VD. 84 miR-93 demonstrates a strong presence in the serum of VD patients, and treatments aiming to attenuate serum miR-93 have shown promise in alleviating pathologic neuroinflammation. Because loss of endothelial and neurovascular integrity is closely associated with ongoing VD pathogenesis, miR-126 has also been implicated as an important factor in the development of VD. 88 miR-126 has been demonstrated to modulate endothelial cells, platelets, and inflammatory molecules to promote vascular integrity following VD by contributing to angiogenesis, and maintenance of vascular patency via modulation of the ERK and AKT pathways. 89 A recent inquiry into the association between miR-126 and VD found that infarction and ischemia associated with VD significantly decrease miR-126 expression and thus contribute to the loss of vascular integrity. Furthermore, injections of miR-126 were demonstrated to improve vascular functioning and angiogenesis. miR-126’s crucial role in maintaining vessel patency presents promise as a therapeutic target for patients with suspected vascular dysfunction and/or cerebral hypoperfusion. Recent research has proposed that miR-26b binds to the mRNA of IL-6 to reduce the production of this inflammatory cytokine. 87 Reduction of IL-6 in turn reduces both neuroinflammation and neural apoptosis. Reduced levels of miR-26b were found in the hippocampus of experimental ischemia models, leading credence to the potential link between miR-26b and VD pathology. Loss of important cytokine modulators such as miR-26b may comprise one of the first steps in the pathogenesis of VD. The role of miR-26a in generating greater levels of angiogenesis proves beneficial in VD as well as in stroke by bolstering collateral circulation and providing greater perfusion in areas of hypoperfusion. 90 Further inquiry into the relationship between miRNAs and cerebrovascular health presents a novel opportunity to develop more efficient treatments, particularly those that harness endogenous repair mechanisms.
Traumatic brain injury
Traumatic brain injury (TBI) refers to a form of acquired brain injury resulting from an external force applied to the head. It can result in a spectrum of physical, cognitive, and emotional impairments, the duration of which hinges on the severity of the injury. These effects may manifest over a short or extended period, making it crucial to assess and manage TBI cases comprehensively for better outcomes and recovery. Recent research has shown that miRNAs are involved in the pathophysiology of TBI and may serve as biomarkers for diagnosis, prognosis, and treatment response. These miRNAs are involved in secondary brain damage induction or prohibition via the regulating inflammatory response, neuronal cell regeneration, regulation of intracellular transport and apoptosis, and controlling the BBB leakage to regulate neuroplasticity. 91 Studies have identified specific miRNAs that are altered following TBI, including miR-21, miR-146a, miR-155, and miR-124. 92 miR-21-5p is found to regulate the neuroinflammation post-TBI via acting on the microglial M1/M2 polarization. 93 In the TBI mice model, miR-23a is found to reactivate the AKT/mTOR pathway and hence modulate neurological apoptosis and inflammation response. 94 miR-212-5p is found to play a protective role against ferroptosis which causes neuronal death in controlled cortical impacted TBI mice model by partially targeting prostaglandin-endoperoxide synthase-2 (Ptgs2). 95 In repetitive mild TBI upregulating miR-124-3p is found to improve cognitive outcomes by Rela/ApoE signaling pathway. 96 miR-351-5p is found to alleviate lipid peroxidation which in turn relieves the stress on the endoplasmic reticulum and exerts anti-ferroptotic effects by acting on 5-lipoxygenase. 97 miR-873a-5p improves neurological deficits and attenuates microglia-mediated neuroinflammation induced by TBI by inhibiting the NF-κB signaling pathway. 98 Additionally, miRNAs have been implicated in the regulation of genes involved in neurodegeneration and neuroinflammation, which are critical features of TBI. This emphasizes the necessity for further comprehensive research into the role of miRNAs in regulating TBI and its potentially fatal consequences.
Brain tumor
Brain tumors represent a diverse group of abnormal growths within the brain or its surrounding tissues, including benign (non-cancerous) or malignant (cancerous). They can originate from various cell types, leading to a wide range of tumor types with distinct characteristics and behaviors. Brain tumors can arise from both primary brain tissue and from cells that have migrated from other parts of the body (metastatic tumors). Gliomas are the most common type of primary brain tumors that originate from glial cells, which provide support and protection for neurons. The profiling of miRNA expression has been shown to impact the activity of oncogenes (tumor suppressor genes) implicated in gliomagenesis, tumor growth, proliferation, apoptosis, and the posttranscriptional regulation of anti-oncogenes.99,100 For example, miR-93 101 and miR-296 102 are found to cause angiogenesis of glial tumors in human glioma cells in vitro via inducing suppression of integrin-β8 expression and overexpression of VEGF2 respectively to increase tumor cell survival, growth and neurosphere formation. As a major angiogenesis regulator, miR-126 enhances the formation of glioma cancer stem cells via down-regulation of insulin receptor substrate-1 in a neurotrophin signaling pathway. 103 miR-132 is found to promote the formation of glioblastoma-initiating cell phenotypes by downregulating tumor suppressor candidate-3 104 and polypyrimidine tract-binding protein 2 (PTBP2) 105 to cause cell proliferation and self-renewal of U87 glioblastoma cells in vitro. Whereas Chen et al. and group found that hepatic leukemia factor-mediated miR-132 can directly suppress threonine tyrosine kinase protein expression, thus exerting inhibitory effects on cancer cell proliferation, metastasis and radioresistance in U87 glioblastoma cells in-vitro. 106 These studies demonstrated that miRs could exert different functions by targeting different downstream pathways. Other study by Wang et al. and group found that miR-132 targets and downregulates the expression of Glioma-associated oncogene protein 1 to inhibit the invasion and proliferation of the U251 glioma cell line. 107 A discrepancy in the outcome of miR-132 could be due different cell lines were used. Moreover, various miRNAs are used to enhance the sensitivity of tumor cells to anti-cancer treatments and outcomes of radiation and chemotherapies. For instance, miR-21 amplifies the cytotoxic impact of temozolomide, doxorubicin, paclitaxel, and sunitinib in U87 glioblastoma cells in vitro. 108 miR-203 has been shown to enhance radio and chemosensitivity by suppressing epithelial-mesenchymal transition in U87 glioblastoma cells by targeting Snail Family Transcriptional Repressor 2. 109
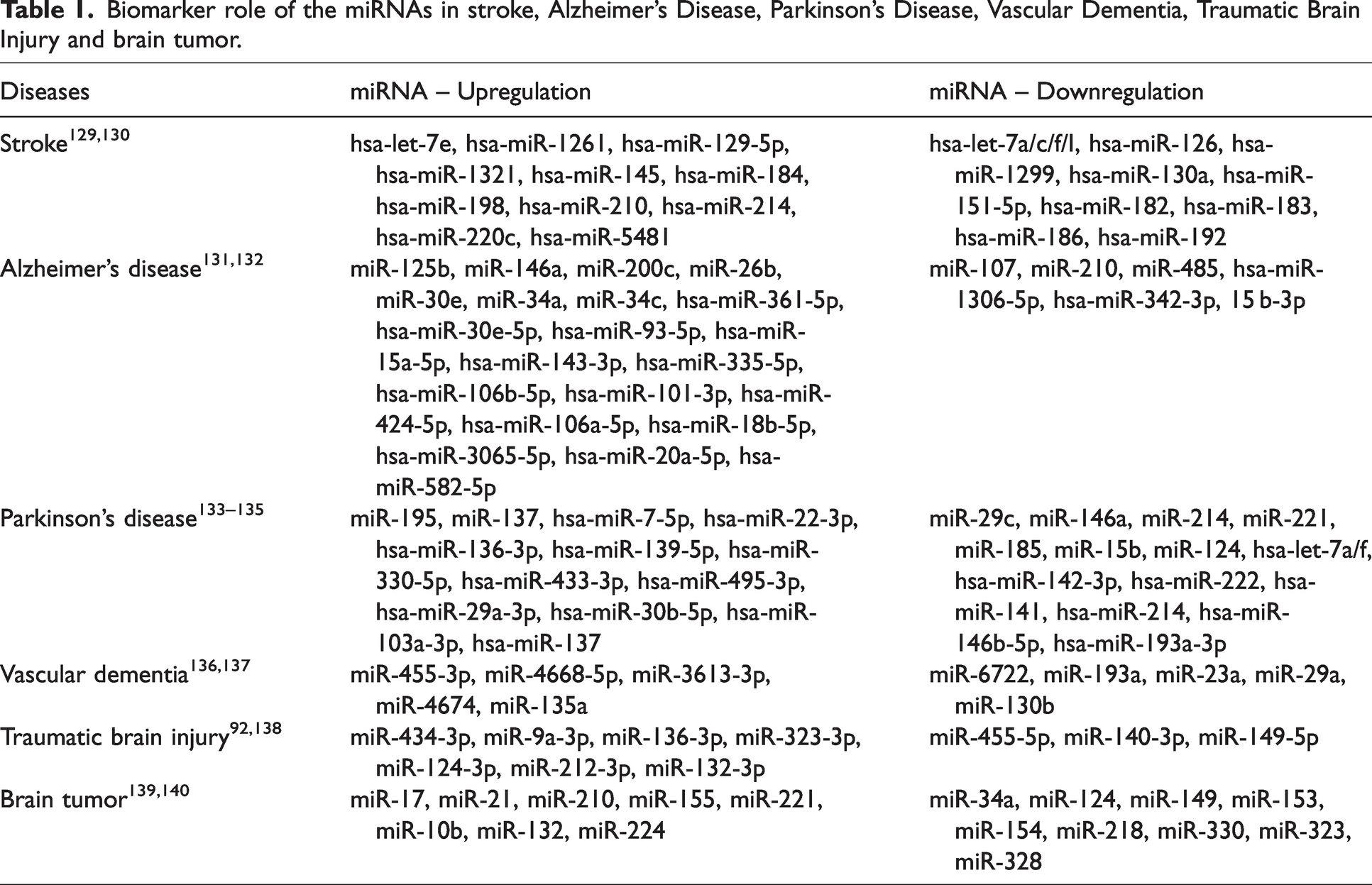
Taken all together, the biomarker role of miRNAs has been widely studied and we summarized the biomarker role (either up or downregulation) of miRNAs in CNS diseases in Table 1. Moreover, we have summarized the role of specific miRNA and its downstream pathway in CNS diseases in Table 2.
Biomarker role of the miRNAs in stroke, Alzheimer’s Disease, Parkinson’s Disease, Vascular Dementia, Traumatic Brain Injury and brain tumor.
Different promising miRNAs with downstream target pathways involved in a major regulatory role in stroke, Alzheimer’s Disease, Parkinson’s Disease, Vascular Dementia, Traumatic Brain Injury and brain tumor.
A novel delivery approach of miRNAs through exosomes
EXs as natural carriers of miRNAs
As we have discussed the different roles of miRNAs in various CNS diseases, the next questions are how do we use miRNAs or deliver miRNAs systemically or locally? Traditionally, we administer exogenous miRNAs by intravenous or local injections, such as brain or heart injections. 110 The advancement in the field of miRNA delivery occurred with the use of viral vectors such as retroviral, lentiviral and adenoviral vectors. However, there are some issues reported with these delivery systems such as the degradation of the miRNAs in the circulation and low efficiency of the penetration to the brain. 110 To overcome these issues, researchers from other groups and our group found a great vehicle for miRNAs, exosomes (EXs), originating from the multivesicular endosomal cell compartment. EXs are nanosized extracellular vesicles, typically ranging from 30–150 nm in size. 111 Once discharged, EXs can either remain in the extracellular vicinity near the point of discharge or traverse into biological fluids like plasma, urine, and CSF. 112 EX-carried miRNAs are increasingly being used in various CNS diseases as excellent diagnostic markers and therapeutic agents. 113 Deep-sequencing technology has presented a reliable approach to detect deregulated exosomal miRNAs in CSF of PD and AD patients enabling fair robustness regarding specificity and sensitivity in the diagnosis. 114
EX-miRNAs mediate cell-specific communications in CNS
Research has found that EXs can deliver miRNAs to various cells to mediate a cross-communication. 115 Because of the nanosize, EXs were reported to be able to cross the BBB which provides an excellent option for treating/managing CNS diseases. Our research has shown that intravenously injected stem cell-released EXs could cross the BBB and merge with the brain ECs, neurons, astrocytes and glial cells. More importantly, the injected EXs could deliver their carried miR-126 to the brain by increasing the level of miR-126 in the brain. 116 Previous studies from other groups further showed that intravenously injected EXs could deliver their carried miR-124-3p to the brain. 96 Additionally, EXs are excellent delivery systems for endogenous miRNAs as they naturally encapsulate and protect miRNAs from degradation by RNases in the extracellular environment. 117 For instance, the delivery of miRNAs in cartilage repair through EXs is reported to be more efficient than direct injections in the management of osteoarthritis. 118 Apart from improvement in efficacy, EXs can also reduce the harmful effects of miRNAs via site-specific action. Shimbo et al. introduced miR-143 into MSCs, and upon delivery to osteosarcoma cells, the miR-143 present within the EXs was able to significantly inhibit the migration of the cancerous cells. 119 After delivered, EXs could be rapidly taken up by recipient cells to mediate cell-specific function. Our in vitro time-dependent experiments showed that the endothelial progenitor cells (EPC)-released EXs could deliver their carried miR-126 116 or miR-210 122 to the neurons, ECs and astrocytes as early as 6 hours after co-incubation. The incorporated miR-126 into the ECs could modulate the level of angiogenic pathways (VEGFR2/PI3k) in ECs to enhance EC function and cell viability. More importantly, EXs can deliver miRNAs to specific cell types via receptor-mediated binding, thus being rapidly taken up by recipient cells, minimizing off-target effects exhibited through systemic circulation. 120 A study by Mathiyalagan et. al demonstrated cell-specific delivery of miRNAs through EXs via in vitro monitoring of human CD34+ stem cell-derived EX-mediated delivery of miRNA precursors in human umbilical vein endothelial cells. 121 Pusic et. al found that nasal administration of IFNγ-stimulated rat dendritic cells secreted EXs accumulate in the brain specifically oligodendrocytes and to a lesser extent in microglial cells to promote the myelination. 122 Deng et al found that bone marrow-derived mesenchymal stem cells-derived EXs deliver miR-138-5p to the astrocytes to offer neuroprotection by downregulating neutrophil gelatinase-associated lipocalin and inflammatory factors in IS mice. 123
EX-miRNAs as promising therapeutic approach for CNS diseases
Since the EX-miRNAs are functional, the therapeutic role of EX-miRNAs has been recently studied. In mice with repetitive mild TBI, the intravenously injected microglial-EX was taken up by neurons in the injured brain. Besides, miR-124-3p in the EXs was found to get transferred into hippocampal neurons and alleviate neurodegeneration by targeting an inhibitory transcription factor of ApoE, Rela and hence affecting Rela/ApoE signaling pathway to promote the β-amyloid proteolytic breakdown, thereby inhibiting β-amyloid abnormalities. 96 Our lab also has found that in IS, EPC-EXs mediated miR-210 have a protective role in the brain against oxidative stress and cell apoptosis as well as promote cell viability both in vitro and in vivo partially though VEGFR2/PI3k and TrkB/PI3k pathway to recover neurological function by increasing angiogenesis and neurogenesis.124,125 To overcome the limitations of EX applications such as rapid clearance, low loading capacity and low target specificity, engineering EXs provide excellent delivery systems and therapeutic agents. According to the recent advancements of methods of engineering EXs, either surface modification or loading modification of EXs can be achieved by various techniques such as plasmid transfection, sonication, electroporation, membrane fusion, co-incubation etc. 126 In CNS diseases, effectively targeting the brain is challenging as EXs tend to localize in the liver and spleen. Whereas engineered EXs have shown target specificity to the brain. For example, blood-derived EXs when loaded with dopamine to treat PD, showed less toxicity and better therapeutic impact on PD mouse model. 127 Tang et al confirmed the therapeutic potential of engineered EX-based miRNA in CNS disease. In this study, they used rabies virus glycoprotein peptide–modified EXs and loaded with miR-25 and miR-181a. After peripheral injection in the transgenic SCA3 mice model of spinocerebellar ataxia type 3, a substantial reduction of ATXN3 aggregation and cytotoxicity was observed. 128 This evidence strongly demonstrates that EXs could be a powerful vehicle or avenue to facilitate the role of miRNAs in managing CNS diseases.
Conclusion and perspectives
In conclusion, miRNAs play an important role in modulating the function of neurovascular via multiple pathways, which leads to regulating the progression of CNS diseases. Given that miRNAs exist widely in the circulation as well as other body fluids, miRNAs have advanced biomarker and therapeutic roles in treating CNS diseases. Nonetheless, the mechanisms that underlie the regulation of neurovascular inflammation and repair through miRNAs deserve further investigation. This pursuit is crucial for the development of miRNA-based therapeutic strategies for CNS disease that lead to specific cell targets more effectively.
Here are some important miRNAs in neurovascular injury and repair –
Modulation of immune cell activation: miR-9, miR-124, and miR-155 have been shown to regulate the activation of immune cells, such as microglia and T cells. Regulation of neuroinflammation: miR-9 and miR-155 have been shown to regulate pro-inflammatory cytokines. miR-21 and miR-146 have been shown to regulate anti-inflammatory cytokines. The duality of the role of miRNAs in neuroinflammation opens up a passage to investigate and modulate the use of miRNAs as therapeutic agents to treat neurovascular repair after injury. Regulation of neurovascular unit integrity and angiogenesis: miR-132, miR-124, and miR-29 have been shown to regulate the expression of NTFs, such as BDNF, which play critical roles in neurovascular repair and plasticity. miRNAs, such as miR-124 and miR-132 have been shown to promote angiogenesis and endothelial cell proliferation, which can contribute to neurovascular repair after injury. Regulation of BBB integrity: miR-124, miR-155, and miR-146a have been shown to regulate BBB integrity by targeting the tight junction complex and basement membrane genes.
Footnotes
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Institute of Neurological Disorders and Stroke (1R01NS102720), Chronic Disease Research Program (WV-INBRE grant, P20GM103434), and West Virginia Clinical and Translational Science Institute (WV-CTSI) grant (2U54GM104942).
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Authors’ contributions
H.S., B.S., and E.M. searched the literature and drafted the manuscript. J.C.B. critically revised the manuscript. All authors have made contributions to the work and approved it for publication.