
Abstract
Aging and metabolic syndrome, common risk factors for ischemic stroke, are associated with collateral rarefaction. We investigated the effects of aging and diet-induced obesity on cerebral collateral arteriogenesis and the restoration of cerebrovascular reserve following proximal arterial occlusion, using a right common carotid artery occlusion (RCCAO) model. Male and female C57BL/6J mice from three age groups (young: 4–6, aging: 16–20, aged: 24–28 months) and a high-fat diet group underwent either RCCAO or sham surgery. The diameters of the circle of Willis (COW) arteries and pial collaterals were measured 4 weeks after RCCAO. Resting cerebral blood flow (CBF) and CBF reserve during induced hypotension were assessed by laser speckle flowmetry at 3 hours and 4 weeks after RCCAO. In young mice, RCCAO induced a significant increase in the diameters of the COW and pial collaterals, leading to complete recovery of baseline CBF at 4 weeks. In contrast, aging progressively diminished arteriogenesis, resulting in partial (aging) and absent (aged) CBF recovery. Cerebrovascular reserve was diminished with advancing age. Diet-induced obesity had milder effects on arteriogenesis. Our study revealed that cerebral collateral arteriogenesis is a strong determinant of impaired cerebrovascular reserve following proximal arterial occlusion and progressively diminishes with aging.
Keywords
Introduction
Collateral development plays an important role in maintaining cerebral blood flow (CBF) in patients with critical stenosis or occlusion of large extracranial and intracranial cerebral arteries.1–5 Moreover, the presence of collaterals governs how long focal ischemic brain tissue survives and the success of reperfusion therapies both clinically and experimentally.6–17 Therefore, it is critical to understand the determinants of collateral formation. In the adult brain, the most important mechanism of collateral development is arteriogenesis, that is, the growth and remodeling of preexisting collateral vessels, driven by increased endothelial shear stress via a complex cascade of molecular and cellular mechanisms that involve endothelial activation and upregulation of adhesion molecules, monokines, vascular endothelial, fibroblast, and platelet-derived growth factors, monocyte infiltration, and smooth muscle cell proliferation.5,18–24
Age is the most common and non-modifiable risk factor for ischemic stroke.25–27 In humans and rodents with acute ischemic stroke, aging is associated with inadequate leptomeningeal collateral perfusion.28–31 Aging is also associated with fewer collateral channels at rest, suggesting rarefaction.5,28,32,33 However, whether age-related rarefaction at rest also reflects a diminished capacity for cerebral arteriogenesis upon chronic proximal artery stenosis or occlusion is poorly understood.34,35 Similarly, vascular risk factors such as diabetes may also affect the arteriogenesis response.
Here, we examined the influence of age and diet-induced obesity on the angiographic circle of Willis (COW) and pial collateral arteriogenesis response (i.e., collateral numbers, outward remodeling) to chronic cerebral hemodynamic insufficiency upon unilateral common carotid artery occlusion (CCAO) in mice,34,36,37 which mimics acute dissections, cardioembolic events occluding the carotid artery, or ruptured carotid plaques in humans. Moreover, we linked collateral arteriogenesis with resting CBF and cerebrovascular reserve in various arterial territories using full-field laser speckle imaging. Our data reveal a profound and progressive age-dependent impairment in arteriogenesis to CCAO, leading to diminished cerebrovascular reserve.
Methods
Animals
Experiments were carried out in accordance with the Guide for Care and Use of Laboratory Animals (National Institute of Health Publication No. 85-23, revised 2011), were approved by the Institutional Animal Care and Use Committee (Massachusetts General Hospital Subcommittee on Research Animal Care), and reported according to the ARRIVE 2.0 guidelines. 38 In the absence of clear guidance on which mouse strain best represents human cerebral circulation, we selected the C57BL/6J mouse strain (Jackson Laboratory, Bar Harbor, ME) because it is the most common background strain in genetic models and has variable presence of posterior communicating arteries, which allowed us to test whether arteriogenesis (i.e., growth of existing arteries) is accompanied by new artery formation. In the aging cohort, a total of 148 mice (n = 80 male, 68 female) of three age groups were used (young: 20±2 weeks; aging: 71±9 weeks; aged: 109±6 weeks). The oldest age group corresponded to over 70 years in human age. 39 In the diet-induced obesity cohort (age 21 ± 0 weeks), a total of 32 mice (n = 16 male, 16 female) were randomly allocated to a regular diet or high-fat diet (HFD; 60% of calories from fat; TD.06414, Teklad, Madison, WI) 12 weeks before surgery and continued on the same diet until sacrifice. This diet is well documented to cause hyperglycemia, hyperlipidemia, and obesity, mimicking metabolic syndrome.40–45 The control cohort received a regular diet for the same period. Mice were kept in an American Association for Accreditation of Laboratory Animal Care (AAALAC)-accredited animal facility in cages of two to four mice with a 12-h light/dark cycle, room temperature of 22 ± 5°C, and humidity of 45–65%.
Experimental timeline
All mice underwent either right common carotid artery ligation (RCCAO) or the sham procedure (Figure 1(a)). A subset of mice was studied 3 hours after the procedure (i.e., acute) to examine the resting CBF and CBF reserve during hypotension using full-field laser speckle flowmetry (LSF). The remaining mice were followed for 4 weeks (i.e., chronic) with weekly neurological exams and body weight measurements. At the end of this period, a subset of these mice underwent an angiogram to measure the diameters of major arteries of COW and pial collaterals. In the remaining mice, CBF was measured in the dorsal cortex using LSF.
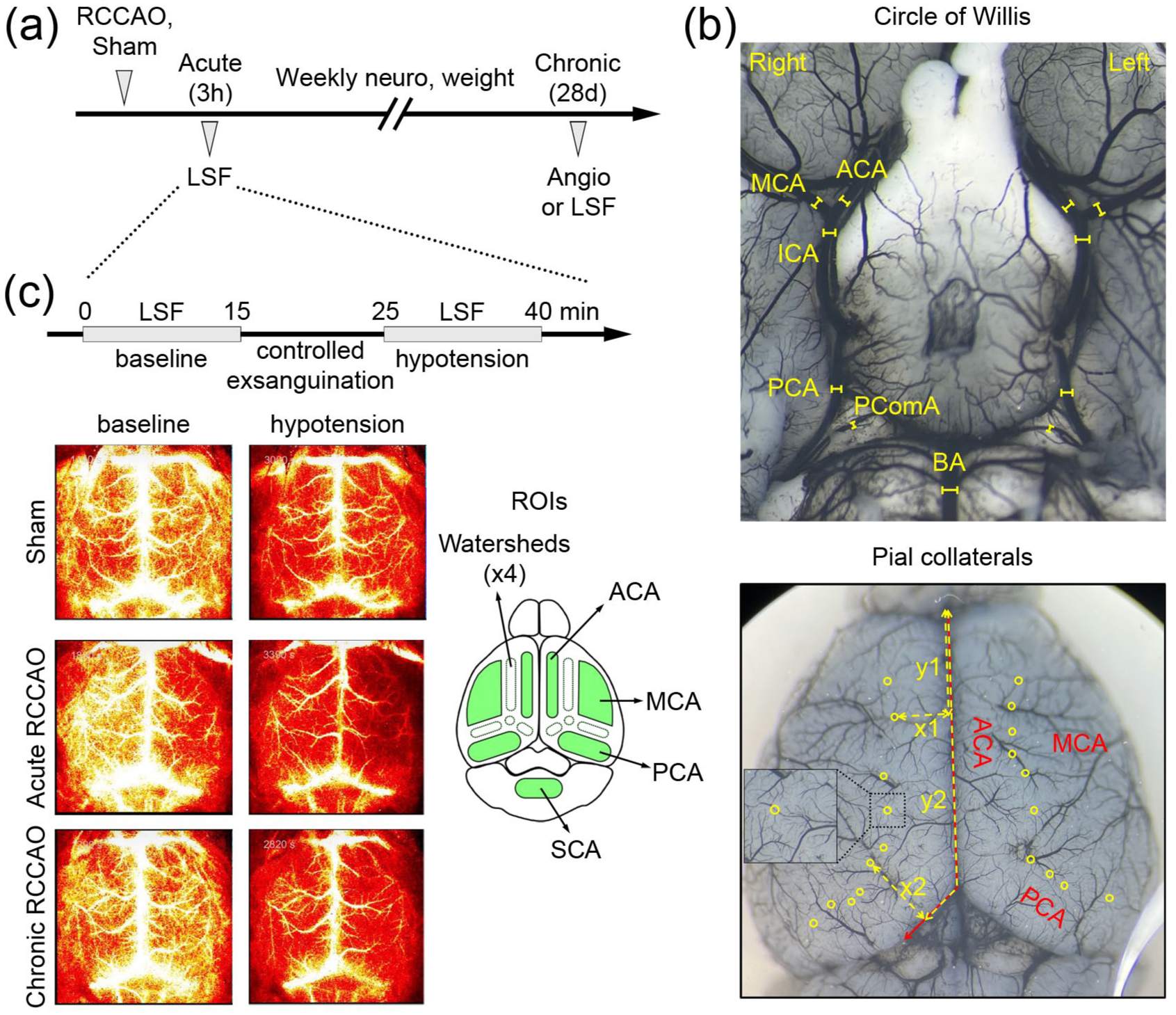
Experimental protocols. (a) The timeline shows the experimental protocol. The timing of the full-field laser speckle flowmetry (LSF) and angiogram are shown after the right common carotid artery ligation (RCCAO) or sham procedure. (b) The upper panel shows a representative image of the diameters of major arteries of the COW (internal carotid artery, ICA; anterior cerebral artery, ACA; middle cerebral artery, MCA; posterior cerebral artery, PCA; posterior communicating artery, PComA; basilar artery, BA). The lower panel shows a representative image of the pial collaterals (yellow circles). The anteroposterior distance from the frontal pole along the medial and posterior hemispheric edge (y1, y2), the mediolateral distance of MCA/ACA collaterals to the midline (x1), and the perpendicular distance of MCA/PCA collaterals to the posterior hemispheric edge (x2) were measured as shown. (c) The upper panel shows the timeline of an LSF imaging experiment. Baseline CBF was measured for 10–15 min after anesthesia was completely switched to avertin. Cerebrovascular reserve was measured after controlled exsanguination. The lower panel shows representative LSF images in different occlusion time points and blood pressure (BP) stages (sham, young female; acute RCCAO, young female; chronic RCCAO, young female). Regions of interest (ROI) were placed on both hemispheres in the major arterial territories and watersheds (lower inset), and CBF was expressed relative to the young sham baseline using inverse correlation time values.
Unilateral common carotid artery occlusion
Mice were anesthetized with isoflurane (3% induction, 1.5% maintenance in 70% N2O and 30% O2). Rectal temperature was maintained at 36°C–37°C by a thermostatic heating pad (FHC, Bowdoinham, ME). The RCCA was exposed by a midline neck skin incision, separated from connective tissue, preserving the vagal nerve, and ligated permanently with a 6-0 silk suture. Sham procedures were performed in the same way without ligation. Skin was sutured with 6-0 silk.
Angiograms
We examined the COW and pial collaterals using black ink angiograms 4 weeks after RCCAO or sham procedure (young sham: n = 24; young RCCAO: n = 23; aging sham: n = 16; aging RCCAO: n = 15; aged sham: n = 9; aged RCCAO: n = 9; HFD sham: n = 8; HFD RCCAO: n = 8). Mice were anesthetized with 4.0% isoflurane and transcardially perfused with 1.2 mL Black India Ink (0.03 mL/s; Chartpak Inc, One River Road Leeds, MA) after clamping the descending aorta. We maintained the infusion rate constant among all animals to ensure consistent filling across groups. However, we did not use a vasodilator because perfusions were successful without a vasodilator, and because pharmacologically dilated diameters may not be the best indicator of collaterals under physiological conditions, in vivo. Brains were carefully removed, and care was taken to preserve the pial arteries and COW branches. The ventral and dorsal brain surfaces were imaged under a stereomicroscope. A blinded observer took the images and performed the measurements. The diameters of major branches of the COW (internal carotid, ICA; anterior cerebral, ACA; middle cerebral, MCA; posterior cerebral, PCA; posterior communicating, PComA; basilar, BA) were measured bilaterally using Image J (NIH, Bethesda, MD) (Figure 1(b), upper panel). Pial collaterals were defined as the middle or the thinnest point between a distal MCA branch and a distal ACA branch or a distal MCA branch and a distal PCA branch (Figure 1(b), lower panel). The number and diameters of pial collaterals were measured. To examine the spatial distribution of pial collaterals, we also measured for each collateral the anteroposterior distance from the frontal pole along the medial and posterior hemispheric edge (Figure 1(b), y1, y2, etc.), the mediolateral distance of MCA/ACA collaterals to the midline (Figure 1(b), x1), and the perpendicular distance of MCA/PCA collaterals to the posterior hemispheric edge (Figure 1(b), x2). In uncommon cases when a segment of the pial arteries was damaged during the brain removal, collaterals in the preserved regions of the same brain were measured and normalized for the entire anteroposterior length to represent the pial collateral numbers; the spatial distribution or diameters of such damaged segments were excluded from the analyses.
Because we used intravascular ink as a contrast agent, all measurements reflected luminal diameters rather than the outer vessel diameters. The optical resolution of the angiograms was 3.33 micrometers/pixel. We did not use digital magnification. To examine the sensitivity of our measurements, we randomly picked five animals from the entire cohort. In each animal, we measured the diameter of five randomly selected pial vessels, as well as the right and left MCA and ACA. We repeated each diameter measurement five times independently for each artery and calculated the coefficient of variation. The median [interquartile range] diameters of pial and COW vessels were 20 [17–23] and 114 [105–125] μm, respectively. The median [interquartile range] coefficient of variation of five repeated diameter measurements from the pial and COW vessels was 7.6 [4.3–10] and 2.8 [1.9–4.2] %, respectively, suggesting methodological reproducibility, high sensitivity, and good statistical power.
Laser speckle flowmetry
Laser speckle contrast imaging provides a measure of CBF changes in the dorsal cortex and, when all optical parameters and surgical preparation are identical, can be used to estimate and compare the resting CBF among animals or experimental groups by using the inverse correlation time values (1/τc, sec−1), as reported previously.46–50 We measured CBF in the dorsal cortex using LSF 3 hours (acute) and 4 weeks (chronic) after RCCAO or sham procedure (young sham: n = 6; young acute: n = 6; young chronic: n = 6; aging sham: n = 6; aging acute: n = 6; aging chronic: n = 6; aged sham: n = 9; aged chronic: n = 7). Continuous full-field LSF was conducted using a near-infrared laser diode (785 nm, 75 mW) and a CCD camera (Cohu 4600, 640 × 480 pixels). Mice were anesthetized with isoflurane (3% induction, 1.5% maintenance in 70% N2O and 30% O2), and rectal temperature was maintained at 36°C–37°C by a thermostatic heating pad (FHC, Bowdoinham, ME). After femoral artery cannulation for mean arterial blood pressure (BP) measurement, mice were placed on a stereotaxic frame, and the skull was exposed by a midline skin incision. Anesthesia was switched to 2,2,2-tri-bromoethanol (Avertin, 250–400 mg/kg intraperitoneally in 20 mg/ml containing 1.25% tert-amyl alcohol, bolus and continuous infusion; Sigma Aldrich, St. Louis, MO) to avoid cerebrovascular effects of isoflurane.46,51,52 Baseline CBF was measured 10–15 min after anesthesia was completely switched to avertin (Figure 1(c)). To evaluate cerebrovascular reserve, we induced hypotension by controlled exsanguination (target BP: 40–50 mmHg). To evaluate CBF changes in different vascular territories, regions of interest (ROI) were placed on both hemispheres in the major arterial territories and watersheds. Average CBF values at baseline and during hypotension were calculated in each ROI using custom-written scripts in MATLAB (Math Works, Natick, MA, USA). In addition to speckle contrast images from individual animals, color maps were generated showing average CBF values for each cohort and ROI relative to young sham baseline 1/τc values (i.e., 100%).
Exclusions
In the cohort allocated to the angiogram experiment, one mouse died on day 1 after RCCAO in the 24–28-month-old group. In the LSF experiment, two mice in the 4–6-month-old sham group died during imaging before baseline CBF measurements, one mouse in the 16–20-month-old acute RCCAO group was excluded because cortical spreading depolarization occurred before baseline CBF measurement, and one mouse each in the 4–6-month-old sham and 16–20-month-old acute RCCAO groups died because of femoral artery cannulation failure. We also excluded the diameters of absent PComA from all analyses (i.e., not imputed).
Statistical analyses
All statistical analyses were performed using GraphPad Prism version 10 (GraphPad Software Inc., San Diego, CA) and SPSS (IBM, Armond, NY, USA). Data were analyzed using two-way ANOVA, followed by post hoc Sidak’s multiple comparisons. In addition, we performed multivariable analysis for each dependent variable using a linear mixed model. The initial model for the angiographic readouts included categorical variables age, sex, occlusion status, artery and hemisphere, and full interactions as fixed-effect predictors. The initial model for the perfusion readouts included categorical variables occlusion status, hemisphere, BP status, and ROI, numeric variable age, and full interactions as fixed-effect predictors. In both models, we performed stepwise elimination based on the Akaike Information Criterion (AIC), continuously adding and removing variables both forward and backward until no further improvement in fit to select the optimal model. All statistical tests are indicated in the text, tables, or figure legends where data are presented. Normality was checked by Shapiro-Wilk test. Nonparametric tests were used for data sets that failed normality. P<0.05 was considered statistically significant.
Results
Aging progressively diminishes COW and leptomeningeal collateral arteriogenesis upon RCCAO
Our a priori hypothesis was that RCCAO induces COW and pial collateral arteriogenesis and that aging impairs this response. Given the large dataset (total n=96), we first performed a multivariable analysis to determine the independent contributions of categorical variables occlusion status (sham, chronic RCCAO), artery (MCA, ICA, ACA, PCA, and PComA), hemisphere (left, right), age (young, aging, aged), and sex (male, female), on the COW diameters and their plasticity. Artery, hemisphere, occlusion status, and age, but not sex, significantly predicted the vessel diameters in the main effects (Supplemental Table 1). We also found a two-way interaction between occlusion status and age, suggesting that the effect of RCCAO on COW diameters was affected by age.
Given the multivariable analysis results, we pooled males and females and plotted the COW diameters to compare the sham and RCCAO groups side-by-side in the three age groups, looking at each artery and hemisphere separately (Figure 2). RCCAO induced a statistically significant increase in the diameters of right but not left PComA and, to a smaller extent, bilateral ACAs; ICA diameters also showed a minor albeit statistically significant increase bilaterally. More importantly, this COW plasticity diminished with advancing age. The aged cohort showed no statistically significant change in COW diameters upon RCCAO. Altogether, these data indicated a progressive defect in COW arteriogenesis with aging. Interestingly, in the sham cohorts, right ACA, ICA, PCA, and bilateral MCAs showed larger diameters in the 16–20-month-old cohort than in the 4–6-month-old cohort. This age-related diameter increase persisted only in the right ACA at 24–28 months of age.
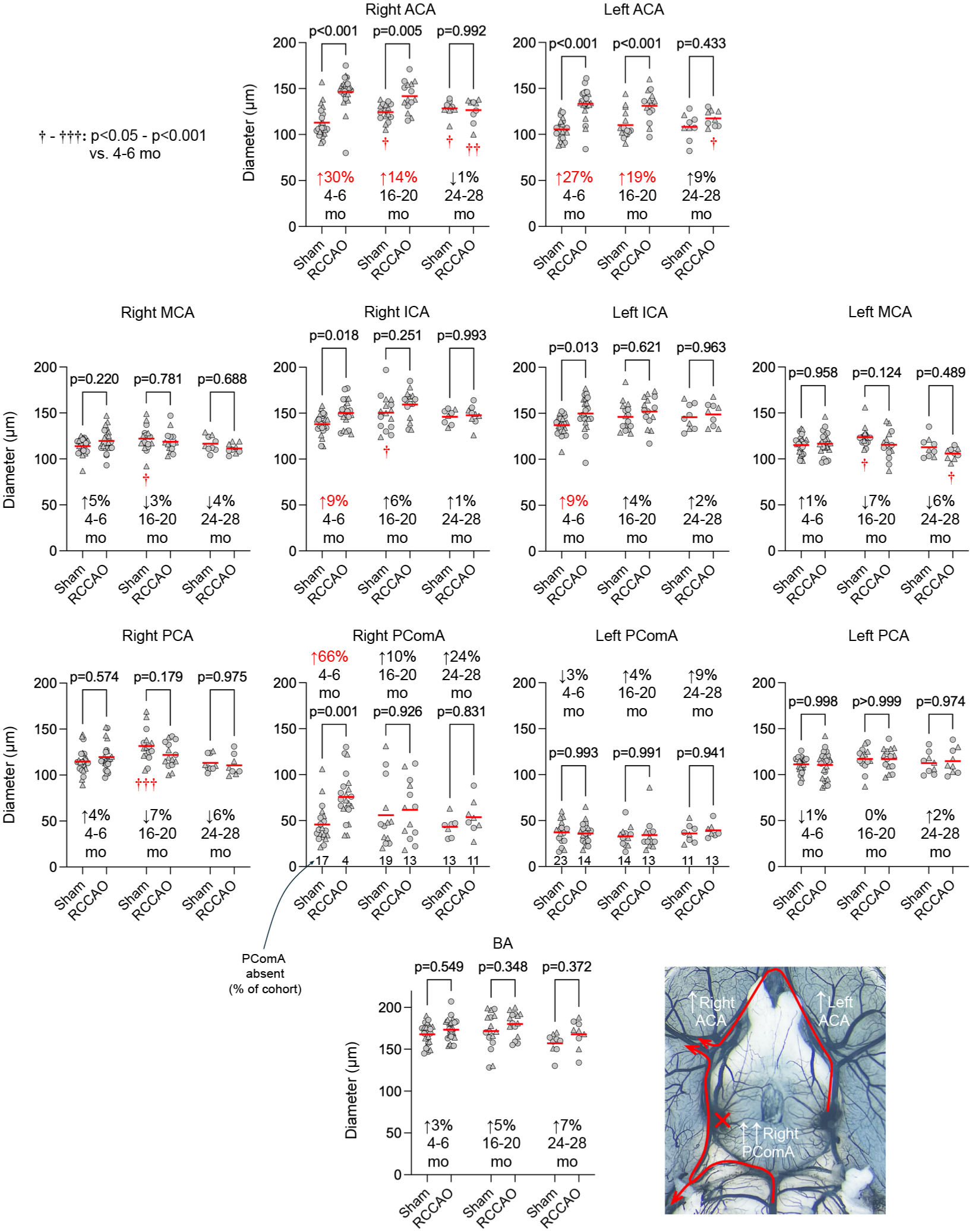
Diameters of major arteries of the circle of Willis in the aging cohort. Individual diameters of the COW are shown in each age group and hemisphere to compare RCCAO with sham. Each symbol represents one mouse. Circles males, triangles females. Statistically significant average magnitudes of diameter change comparing sham versus RCCAO are shown in red on each panel. The percentage of animals with absent PComA is also shown at the bottom of the PComA graphs (Two-way ANOVA followed by Sidak’s multiple comparisons within each graph panel using independent variables occlusion status and age. The lower right panel shows the flow diversion after RCCAO.
Next, using a similar multivariable approach, we examined the predictors of dependent variables pial collateral diameters, numbers, and distance from the midline (independent categorical variables: occlusion status, hemisphere, age, and sex; Supplemental Table 2). Age and occlusion status interacted to predict pial collateral diameters. Direct examination of the data showed that pial collateral diameters in the 4–6-month-old cohort were larger in the RCCAO arm compared with shams on both hemispheres, and this arteriogenesis to RCCAO was completely absent in aging and aged cohorts (Figure 3), mirroring the findings with COW diameters. Moreover, bilateral growth of pial collaterals suggested that the contralateral (i.e., left) hemisphere was also hemodynamically affected by RCCAO. Sex also independently predicted pial collateral diameters, which appeared 9% larger in females (male 20.3 ± 3.5 vs female 22.1 ± 3.7 μm, p < 0.001, unpaired t test, n = 96 each; mean ± SD).
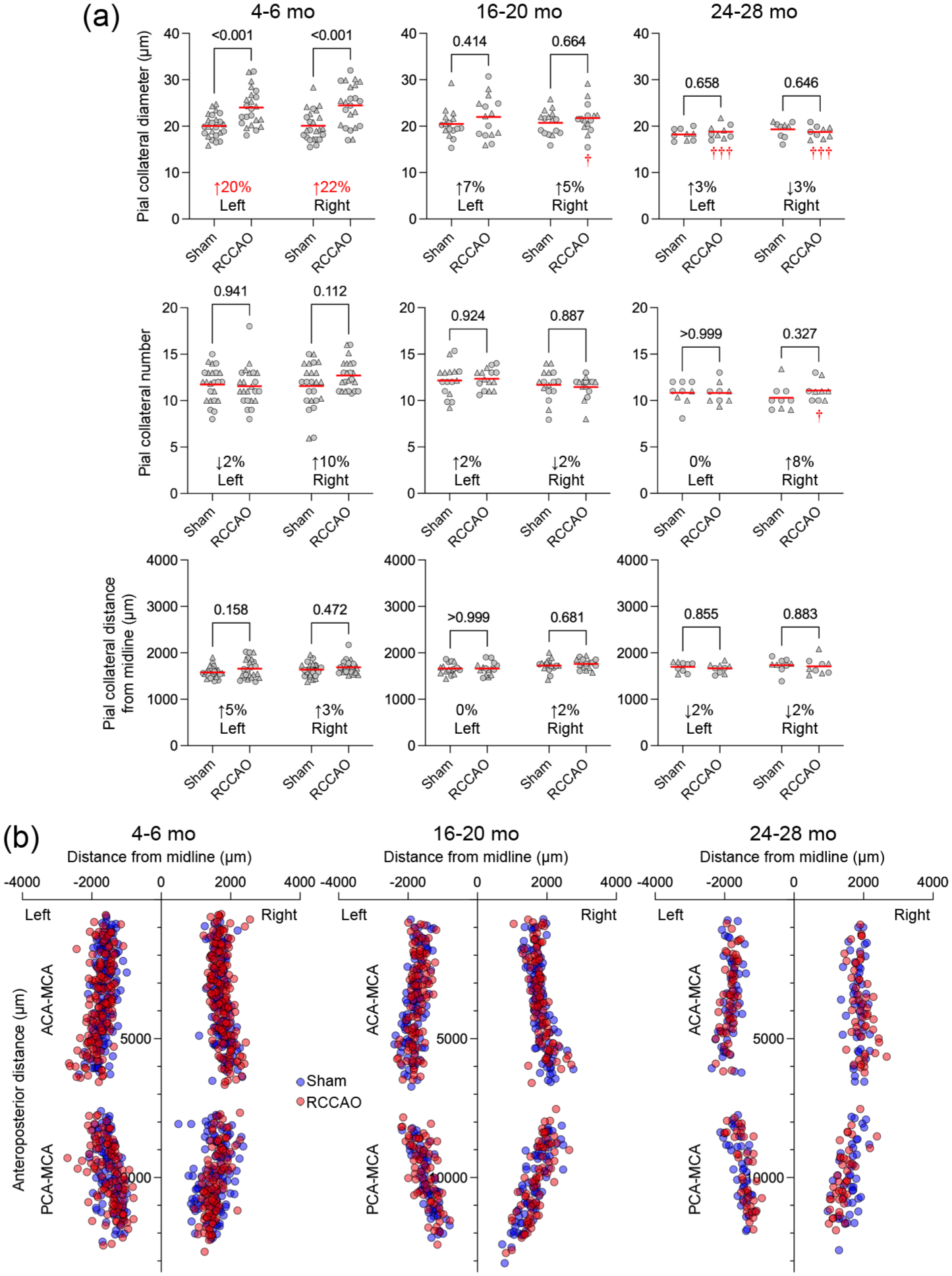
Pial collateral diameters, numbers, and distributions in the aging cohort. (a) The pial collateral diameters, numbers, and distances from the midline are shown in each age group and hemisphere to compare RCCAO with sham. Each symbol represents the average of all pial collaterals in one mouse. Circles males, triangles females. The average percent change in the diameters, numbers, and distances after RCCAO are shown on each panel. Statistically significant magnitudes of diameter change between sham versus RCCAO are shown in red (Two-way ANOVA followed by Sidak’s multiple comparisons within each graph panel using independent variables occlusion status and hemisphere). (b) The lower panel shows the spatial distribution of pial collaterals (red RCCAO, blue sham).
Pial collateral numbers did not respond to RCCAO or differ between the hemispheres or sexes and were predicted only by age without interactions (Supplemental Table 2). Direct examination of the data showed smaller pial collateral numbers in the aged group (β-coefficients: −0.8 collaterals vs young, −1.4 collaterals vs aging; Figure 3).
The distance of pial collateral midpoints from the medial or posterior hemispheric edge was also affected by age; both aging and aged cohorts were closer to the MCA than ACA and PCA territories (β-coefficients: 61 and 60 μm, aging and aged vs young, respectively; Figure 3). In contrast, occlusion status did not affect the distance of pial collateral midpoints from the medial or posterior hemispheric edge. Interestingly, the multivariable model also showed that the distance of pial collateral midpoints was larger on the right versus the left hemisphere (β-coefficient: 55 μm); the biological underpinnings or the significance of this finding is unclear. These data showed that the age-dependent progressive defect in arteriogenesis also involves smaller caliber pial collaterals.
Diet-induced obesity diminishes COW and leptomeningeal collateral arteriogenesis upon RCCAO
To evaluate the influence of diet-induced obesity on the COW and pial collateral arteriogenesis, we examined the diameters of the COW and pial collaterals, the number of pial collaterals, and their distance from the midline, using the same statistical approach as in the aging cohort. In the control cohort on a regular diet, RCCAO induced a statistically significant increase in the diameters of the right PComA and, to a lesser extent, the bilateral ACAs; the left ICA also showed a minor but statistically significant increase, similar to that observed in the young control mice (4–6 months) in the aging experiment. In contrast, the HFD cohort responded to RCCAO with relatively weaker arteriogenesis; statistically significant changes were noted only in the diameters of the right PComA and left ACA (Figure 4). Pial collateral diameters in the regular diet group were larger after RCCAO compared to the shams in the right hemisphere, which was again diminished in the HFD cohort (Figure 5). Pial collateral numbers did not change in response to RCCAO in either the control or HFD cohorts. Overall, these data indicated that diet-induced obesity attenuated arteriogenesis in the COW collaterals, particularly the right ACA, left ICA, and, to a lesser extent, the left ACA, as well as in pial collaterals, though not to the degree observed in the aging cohort.
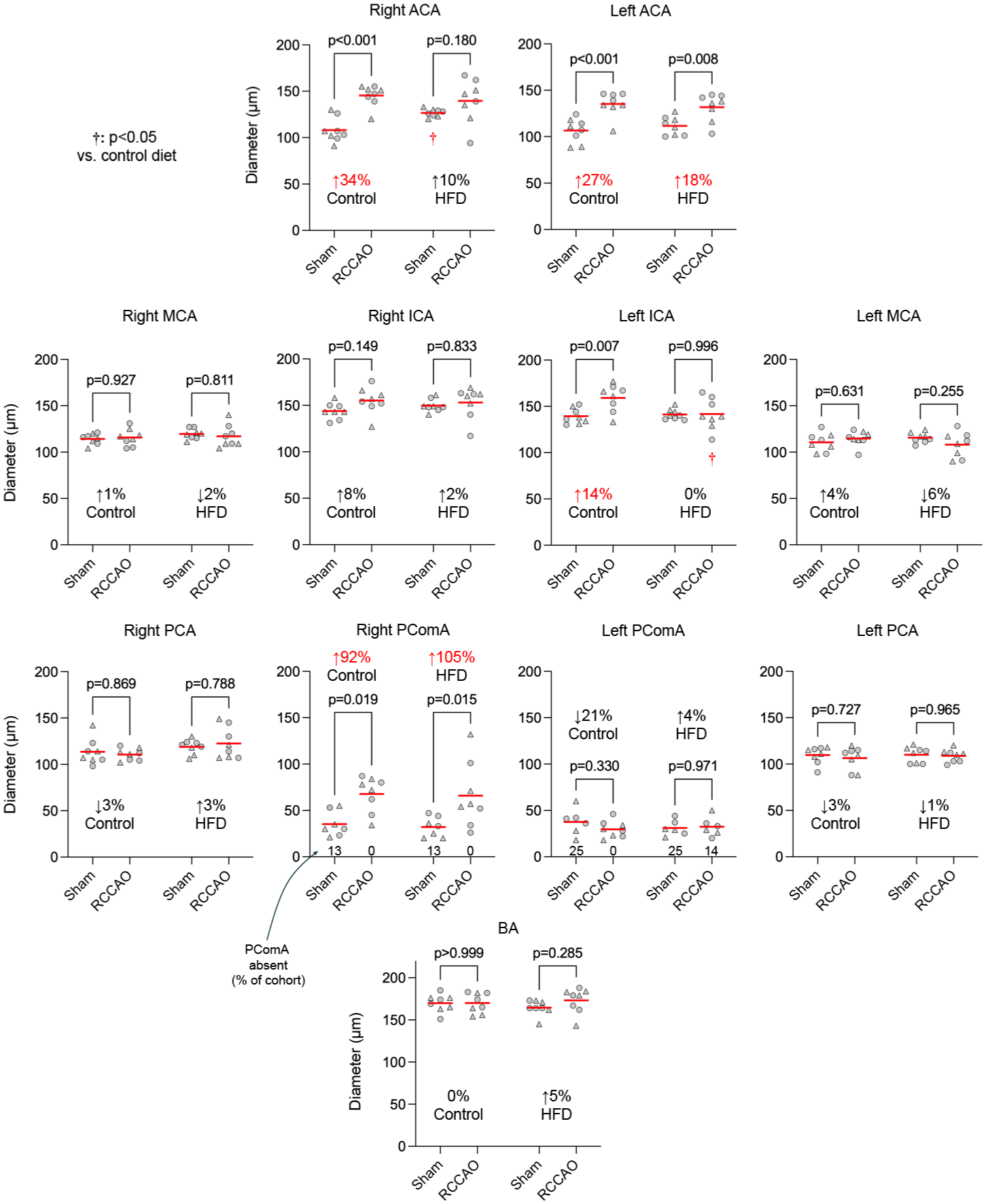
Diameters of major arteries of the circle of Willis in the diet-induced obesity cohort. Individual diameters of the COW are shown in each group and hemisphere to compare RCCAO with sham. Each symbol represents one mouse. Circles males, triangles females. Statistically significant average magnitudes of diameter change comparing sham versus RCCAO are shown in red on each panel. The percentage of animals with absent PComA is also shown at the bottom of the PComA graphs (Two-way ANOVA followed by Sidak’s multiple comparisons within each graph panel using independent variables occlusion status and high-fat diet).
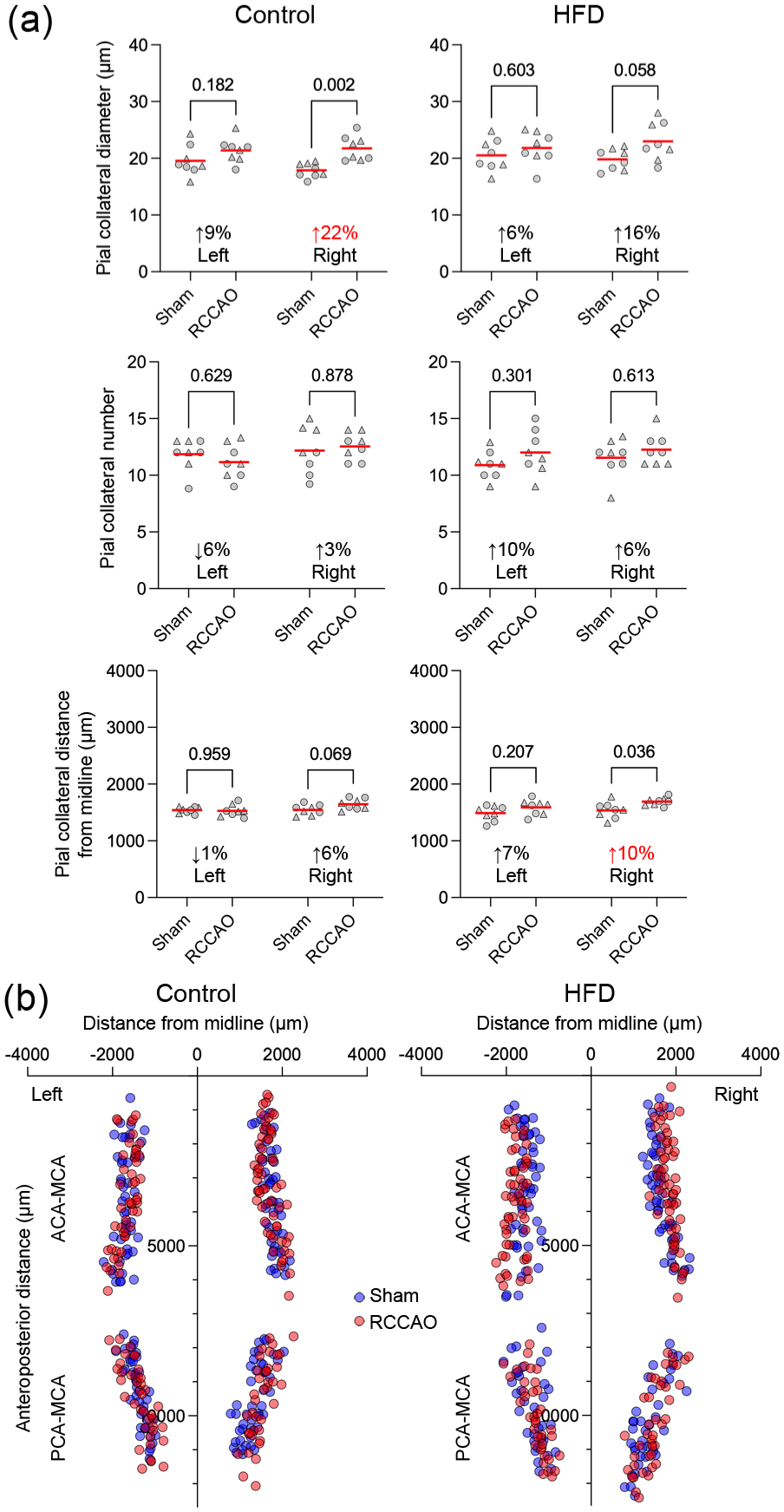
Pial collateral diameters, numbers, and distributions in the diet-induced obesity cohort. (a) The pial collateral diameters, numbers, and distances from the midline are shown in each group and hemisphere to compare RCCAO with sham. Each symbol represents the average of all pial collaterals in one mouse. Circles males, triangles females. The average percent change in the diameters, numbers, and distances after RCCAO are shown on each panel. Statistically significant magnitudes of diameter change between sham versus RCCAO are shown in red (Two-way ANOVA followed by Sidak’s multiple comparisons within each graph panel using independent variables occlusion status and hemisphere). (b) The lower panel shows the spatial distribution of pial collaterals (red RCCAO, blue sham).
Aging progressively diminishes resting CBF and CBF reserve upon RCCAO
Given that aging exerted a more profound influence on the COW and pial collateral arteriogenesis compared with diet-induced obesity, our investigation of CBF reserve focused on the effects of aging. To determine the impact of diminished COW and pial collateral arteriogenesis on resting CBF and the cerebrovascular reserve, we employed LSF, a full-field minimally invasive optical imaging technique, to measure CBF in the dorsal cerebral cortex through the intact skull with high spatial and temporal resolution. We initially placed ROIs in the three main arterial territories (i.e., ACA, MCA, PCA) and four watersheds (Figures 1 and 6). The latter were placed because we expected watersheds to be more vulnerable to perfusion pressure drop. However, contrary to our expectation, watersheds did not show a particular predilection for hypoperfusion; they simply reflected a transition between (i.e., average of) the adjacent main territories (Figure 6). Therefore, further statistical analyses focused on the main territories (i.e., MCA, ACA, PCA).
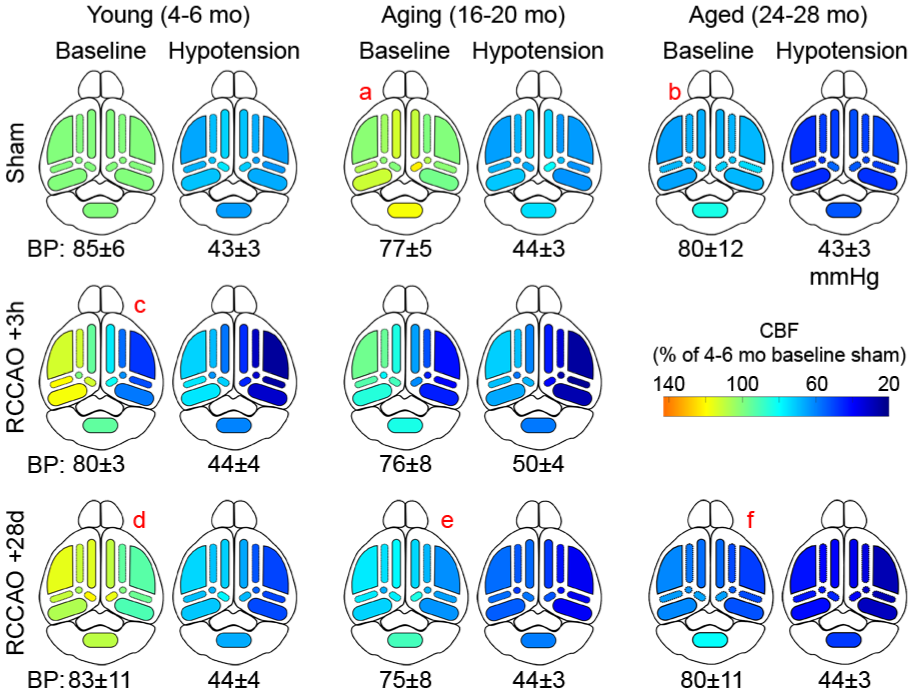
Color maps of CBF changes as an overview. Color maps are shown based on the average CBF values in each occlusion status, BP status, and ROI relative to young sham baseline (i.e., 100%) inverse correlation time (1/tc). BPs during baseline and hypotension are shown under each color map (mean±SD). Labels “a-f” are described in the Results.
The multivariable linear mixed effects model showed a highly significant contribution of all independent variables, including age, occlusion status, BP status, hemisphere, and ROI (Supplemental Table 3). Age interacted with all other variables. Anticipated interactions also emerged among the other variables, lending confidence to the model. Based on these results, we examined the effect of age on resting CBF (see Methods) and cerebrovascular reserve (average perfusion maps and individual data points are shown in Figures 6 and 7, respectively).
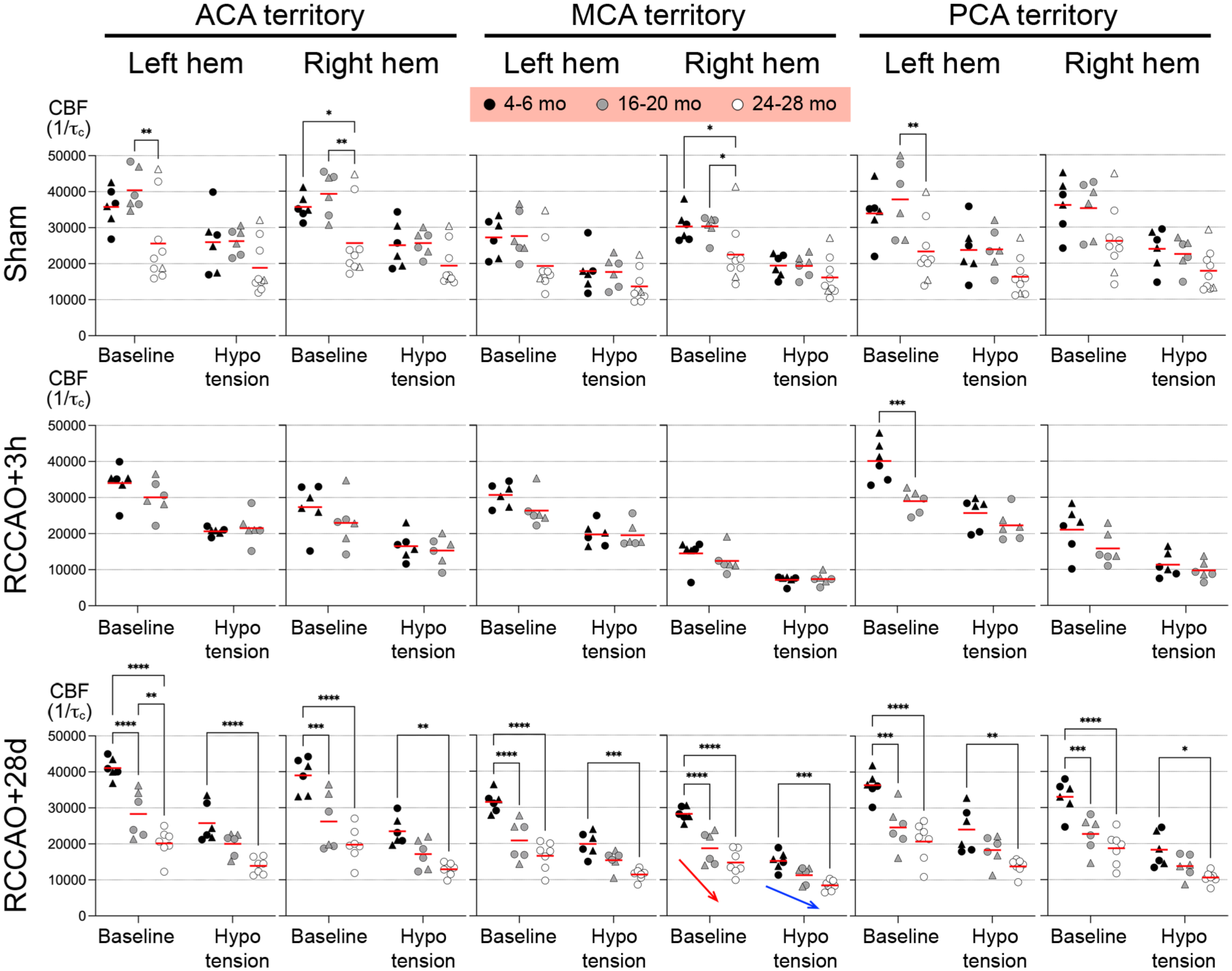
Inverse correlation time values as a surrogate for absolute CBF. The average CBF change is shown in each occlusion status, BP stage, ROI, and hemisphere, comparing the three age groups represented with different gray tones. Each symbol represents the CBF in one mouse. Circles males, triangles females. Red and blue arrows show age-dependently diminished recovery of resting CBF and cerebrovascular reserve after RCCAO, respectively; only the right MCA ROI is marked for brevity, but the observation applies to all ROIs (Two-way ANOVA followed by Sidak’s multiple comparisons within each graph panel using independent variables age and BP stage.
In the absence of RCCAO (i.e., sham), baseline CBF was comparable between young and aging cohorts (Figure 6(a)). In contrast, the aged cohort had ~30% lower resting CBF in all ROIs and both hemispheres (Figure 6(b)). In the acute (3-h) RCCAO cohort, young mice showed hypoperfusion in the right hemisphere, which was most severe in the MCA territory (~50% of baseline; Figure 6(c)), likely because it is the farthest from the COW and the ACA confluence and PComA collaterals. PCA was also severely affected (~40% reduction), probably due to the often hypoplastic or absent PComA in this mouse strain. The CBF drop in the right ACA was milder (~25% reduction). Aging mice showed a similar pattern of hypoperfusion to young mice, albeit more severe, especially in the right MCA territory (~60% reduction). We could not test acute RCCAO in the aged cohort because of concern for high mortality during the second round of anesthesia for imaging and superimposed hypotension 3 hours later.
In the chronic (28-day) RCCAO cohort, CBF completely recovered in young mice to or above baseline in all territories (Figure 6(d)). In contrast, recovery of CBF was partial in aging mice (Figure 6(e)) and completely absent in the aged group (Figure 6(f)), mirroring the age-dependent loss of arteriogenesis (Figure 7, red arrow). Superimposed hypotension exacerbated the territorial hypoperfusion (Figure 6; BPs indicated under each map; mean ± SD). In the sham cohort, hypotension caused a symmetric and relatively even reduction in CBF in all ROIs and age groups, paralleling the baseline CBF (Figures 6 and 7). In the acute (3-h) RCCAO cohorts, hypotension further reduced CBF in the right MCA, PCA, ACA, and left ACA. In the young chronic (28-day) RCCAO cohort, hypotension exacerbated the hypoperfusion in all ROIs (Figure 6), which was worse in the right hemisphere (Figure 7, blue arrow). The same pattern was observed in the aging and aged groups, albeit more severe. Altogether, perfusion imaging suggested diminished cerebrovascular reserve after RCCAO that was partially compensated by age-dependent collateral arteriogenesis over time.
A total of five animals developed a spreading depolarization emerging from the right MCA territory during the hypotension phase, clearly visible on LSF; four of these were in the acute RCCAO group (n = 2, 4–6 months of age; n = 2, 16–20 months of age), and 1 was in the 16–20 months chronic CCAO group. None of the other animals developed SD (n = 28). The laser speckle CBF values (1/τc in artificial units) were 6778 (4937–8186) and 8659 (7687–13016) in animals that developed an SD versus those that did not, respectively (median [interquartile range]; p = 0.03). Therefore, SD occurrence was more likely in animals with lower residual CBF during hypotension after RCCAO.
Aging does not affect neurological function or weight loss upon RCCAO
We did not observe any overt neurological deficits or weight loss during the follow-up after the RCCAO or sham procedure, except for one female mouse in the 24–28-month-old cohort that displayed leaning to the right 2 days after the RCCAO for the next 3 weeks.
Discussion
Age is the most common and non-modifiable risk factor for ischemic stroke.25–27 Aging is associated with diminished leptomeningeal collateral circulation, an important determinant of ischemic stroke outcomes.28–31,53 Here, we show that aging progressively diminishes resting CBF, cerebrovascular reserve, and, most importantly, collateral arteriogenesis upon unilateral CCAO in a simple but powerful mouse model34,36,37 that mimics acute dissections, cardioembolic events occluding the carotid artery, or ruptured carotid plaques in humans. Collateral growth was evident 4 weeks after RCCAO in both the circle of Willis and leptomeningeal collaterals and corresponded well with the restoration of cerebrovascular reserve in different arterial territories in an age-dependent manner. We believe our study is the most comprehensive to date of aging-induced impairment in cerebral collateral arteriogenesis after carotid occlusion, examining both the COW and the pial collaterals and linking these to cerebral hemodynamic reserve in all three major arterial territories using laser speckle imaging in three age groups ranging from 4 to 28 months.
Metabolic syndrome is also recognized as a risk factor for ischemic stroke.54–58 In rodent models, metabolic syndrome is associated with impaired collateral growth following major arterial occlusion in various organs.59–63 In the present study, we hypothesized that diet-induced obesity, as a model of metabolic syndrome, diminishes arteriogenesis in the COW and leptomeningeal collaterals. Although some degree of collateral impairment was observed 4 weeks after RCCAO, its impact appeared to be less pronounced compared with that seen in aging. Chronic inflammation and endothelial dysfunction are key features of metabolic syndrome,64–66 however, their contribution on cerebral collateral arteriogenesis remains unclear. Further studies using diet durations longer than 12 weeks and other animal models of vascular risk factors (e.g., hyperlipidemia, hyperhomocysteinemia) will be important to further elucidate the relationship.
The circle of Willis arteriogenesis upon RCCAO revealed a unique pattern of flow diversion (Figure 2, angiographic inset). The robust increase in ipsilateral PComA diameter indicated flow redirected from the superior cerebellar artery (SCA) to the PCA, presumably assisting the MCA and PCA perfusion. This flow diversion from the SCA was evident in the small drop in cerebellar perfusion in the aging cohort after acute RCCAO (Figure 6, RCCAO +3 h). Note that rodents typically have a “fetal” PCA originating from the ICA, and the PComA, when present, connects the SCA to the PCA. The symmetric growth in bilateral ACAs indicated flow diversion from the contralateral ICA, which was also apparent in the small drop in left hemispheric perfusion after acute RCCAO (Figure 6, RCCAO +3 h). The leptomeningeal collaterals also responded to RCCAO by increasing their diameters, which, interestingly, was bilateral (Figure 3). In contrast, the number of leptomeningeal collaterals was unaffected by RCCAO, although the aged group had lower numbers regardless of the occlusion status (Supplemental Table 2). Similarly, the distance of leptomeningeal collateral midpoints from the medial or posterior hemispheric edge did not respond to RCCAO but varied with age.
These data are generally consistent with prior studies examining similar readouts.5,34,36,37,67–77 A previous study showed diminished collateral growth and recovery of cerebrovascular reserve in aged C57/BL6 mice after right ICA occlusion (RICAO). 34 Despite the overall conclusions being consistent, there were conflicting findings as well. For example, the previous study found only a 24% and 11% increase in PComA diameters after RICAO in 4–6-week to 18-month-old mice, respectively, which did not reach statistical significance. In contrast, we found a 66% increase in PComA diameters after RCCAO in 4–6-month-old mice. The previous study found increased diameters in MCA, PCA, and BA after RICAO in 4–6-week-old mice, while we found virtually no change in these vessels after RCCAO in any age group. Baseline CBF was reduced in the 18-month-old group compared with the 4–6-weeks-old cohort in the previous study, while 18-month-old mice had normal baseline CBF compared with 4–6-months-old mice in our study. Surprisingly, the previous study found no acute or chronic effect of RICAO on MCA perfusion in any age group, while RCCAO caused a significant drop in MCA perfusion in all age groups in our study, as previously reported. 78 However, there were notable differences between the study designs and readouts (previous study vs ours): RICAO versus RCCAO; age range from 4 to 6 weeks to 18 months versus 4–6 months to 24–28 months; acute post-occlusion imaging at 30 min versus 3 h; chronic post-occlusion follow up 2–3 weeks versus 4 weeks; angiograms performed via aortic perfusion after systemic papaverine versus intracardiac perfusion alone; cerebrovascular reserve studied using systemic acetazolamide versus systemic hypotension; and ketamine/xylazine versus Avertin anesthesia. Moreover, we reported all angiographic readouts on the right and left separately; the previous study reported a single set of data likely representing an average of right and left. Any number of these differences could explain the conflicting findings between the studies. Another study in mice found a progressive decrease in pial collateral numbers and diameters between 3 and 31 months of age. 28 This is consistent with our data. The same study suggested diminished pial collateral diameter increase after permanent distal MCA occlusion in the 24-month-old group, although the percent change compared with the control (i.e., non-ischemic) group was not that different from 3-month-old mice. Lastly, a previous study in mice found no effect of sex on pial collaterals, 79 but in our multivariable analyses, female sex independently predicted larger pial collateral diameters. These data suggest that the magnitude of the effect of age and sex on baseline morphology and arteriogenesis may depend on the species, occlusion model, and experimental technique.
Mechanistically, aging-induced impairment of collateral arteriogenesis is likely linked to endothelial dysfunction and dysregulation of nitric oxide (NO) signaling. Endothelial cells play an important role in sensing shear stress and initiating vascular remodeling through eNOS-derived NO production, which promotes vasodilation, endothelial activation, recruitment of monocytes and proliferation of smooth muscle cells.18–24 Aging is associated with oxidative stress, inflammation, and eNOS uncoupling, leading to reduced NO bioavailability and impaired endothelial shear stress responsiveness.80,81 Recent studies confirmed that aging is associated with endothelial senescence, loss of shear-sensing pathways, and decreased collateral vessel plasticity, consistent with our findings.5,82 Altogether, age-related decline in endothelial NO signaling could represent a central mechanism underlying the progressive loss of cerebral collateral plasticity but molecular mechanisms are likely multifactorial and warrant further investigation.
Contrary to our expectations, watersheds did not appear more susceptible to hemodynamic impairment upon RCCAO. The most likely reason for this was that the ipsilateral MCA territory was the most compromised because of its distance to the COW, and that ipsilateral ACA and PCA were receiving collateral flow from the contralateral ACA and PComA, creating a linear CBF gradient from ACA and PCA toward MCA. Interestingly, however, even in the sham group, hypotension did not cause more severe hypoperfusion in the watersheds compared with the large arterial territories in any age group, suggesting that the reduced pial collateral numbers and diameters in aged mice did not affect watershed perfusion. Of note, we placed the watershed ROIs guided by the typical distribution of pial collaterals in the angiographic study; therefore, we are confident that the measurements were indeed from the watersheds.
Our study has limitations as well. First, we studied only a single mouse strain (i.e., C57BL/6J). Although this is the most common mouse strain used in cerebrovascular studies and displays inherent variations in the COW akin to humans, there are numerous mouse strains with more or less robust COWs in which the impact of aging might differ. Second, we examined only aging and diet-induced obesity. Although these are major risk factors for ischemic stroke,25–27,54–58 other common comorbidities may also influence collateral arteriogenesis. Third, aging is associated with increased skull thickness between 3 and 20 months. 83 Although this might, in theory, affect the LSF measurements of resting CBF, we did not find a difference in resting CBF between 4 and 6-month-old and 16–20-month-old cohorts, suggesting that skull thickness did not significantly contribute to the perfusion values obtained from LSF. Fourth, we did not measure blood flow directly within the main cerebral arteries given the relatively low resolution of full-field transcranial LSF. Nevertheless, we believe our measurements of CBF within individual arterial territories and the watersheds actually reflect the blood flow in the main arteries as surrogates. There are other non-invasive CBF measurement methods as well (e.g., color-coded pulsed Doppler ultrasonography, photoacoustic imaging), that provide complementary information when coupled to LSF.84–86 Fifth, we selected CBF autoregulation as our main vascular reactivity index since our focus was RCCAO as a model of hemispheric ischemia, where blood pressure fluctuations are often responsible for transient neurological deficits. Obviously, there are other commonly used paradigms to interrogate cerebrovascular reflexes (e.g., functional neurovascular coupling, hypercapnic hyperemia), and the pattern of loss and recovery of these upon RCCAO may differ from autoregulation and remains to be studied.
In summary, our study shows that cerebral collateral arteriogenesis is progressively diminished with aging and is associated with impaired cerebrovascular reserve. This is a simple yet powerful experimental model to interrogate the determinants and mechanisms of arteriogenesis in response to proximal perfusion pressure drop upon occlusion of a major cervical artery and to test interventions. We did not examine the molecular mechanisms of age-induced impairment, but given the breadth of gene expression changes associated with aging, the molecular underpinnings of diminished arteriogenesis in aged mice are likely to be many and highly complex.28,80,81,87
Supplemental Material
sj-docx-1-jcb-10.1177_0271678X251411837 – Supplemental material for Cerebral collateral arteriogenesis and cerebrovascular reserve in aged and diet-induced obese mice after unilateral common carotid occlusion
Supplemental material, sj-docx-1-jcb-10.1177_0271678X251411837 for Cerebral collateral arteriogenesis and cerebrovascular reserve in aged and diet-induced obese mice after unilateral common carotid occlusion by Yuichi Sasaki, Sanem A. Aykan, Joanna Yang, Fumiaki Oka, Xuyan Jin, Andreia Morais, Takahiko Imai, Elyssa Alber, Sava Sakadžić, David Y. Chung and Cenk Ayata in Journal of Cerebral Blood Flow & Metabolism
Footnotes
Authors’ contributions
YS and CA designed and coordinated the research. YS performed all experiments and analyzed data with support from SA. JY, FO, XJ, AM, TI, EA, and DC supported the experiments. SS provided aged mice essential for the experiments. YS and CA wrote the manuscript. All authors reviewed the manuscript and accepted submission.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: grants from the National Institute of Health (R01NS129192, R01NS115401).