
Abstract
Objective
To explore the effects and mechanisms of metandienone abuse on hepatotoxicity.
Methods
HepG2 cells were treated with 1 μg/mL metandienone for 24 h. Cell viability and apoptosis were detected via Cell Counting Kit-8 and terminal deoxynucleotidyl transferase dUTP nick end labeling assays, respectively. Cellular morphology was assessed via transmission electron microscopy. Genetic changes were analyzed using transcriptome sequencing. Meanwhile, metandienone (0.1 mg·g−1·d−1) was administered via gavage to mice for 14 days. Liver pathology was examined via hematoxylin and eosin, periodic acid–Schiff, and transmission electron microscopy. Serum alanine aminotransferase, aspartate aminotransferase, alkaline phosphatase, albumin, total cholesterol, triglyceride, lactate dehydrogenase, and creatine kinase were measured.
Results
Cell Counting Kit-8 assay confirmed that cell viability decreased to 88.01% after 24 h of metandienone treatment. Terminal deoxynucleotidyl transferase dUTP nick end labeling assay showed that the apoptosis rate increased to 25.08%, compared with 4.94% in the NC group. RNA transcriptome data showed that the expression levels of 221 genes in the metandienone group altered significantly; these genes were closely related to liver metabolism, oxidative stress, inflammatory response, and immune regulation. Meanwhile, exposure to metandienone-induced liver injury was characterized by hepatocyte steatosis and inflammation, accompanied by significant increases in serum alanine aminotransferase and aspartate aminotransferase.
Conclusion
The abuse of anabolic hormones leads to liver function damage, and the mechanism may be related to inflammatory responses and metabolism.
Introduction
Metandienone (MA) is an androstane derivative of anabolic androgenic steroids (AAS), with strong anabolic effects and weak androgen-like effects. 1 MA has been listed as a prohibited doping substance by the International Olympic Committee since 1976 and remains a key controlled substance by the World Anti-Doping Agency. 2
MA increases protein synthesis, inhibits protein breakdown, and promotes muscle growth. It is clinically used to treat patients with chronic wasting diseases, osteoporosis, dwarfism, and aplastic anemia and can also accelerate wound repair and improve lipid metabolism. It has been reported that oral administration of 0.05 mg/kg body weight (BW) of dehydrotestosterone aqueous suspension to 8-week-old white stone chickens daily significantly promoted growth and increased nucleic acid levels in pectoral muscles. 3 A study has also shown additive effects of AAS and exercise on body, tibia, and reproductive organ weights in male Wistar rats. 4 However, abuse of MA has become a significant risk factor endangering public health and social security. In particular, the illegal use of food-derived stimulants in sports competitions 5 and their use as growth promoters in livestock and poultry feed have led to prominent hormone residues in animal-derived foods,6,7 posing a threat to food and drug safety.
Studies show that MA use within therapeutic doses is relatively safe. Short-term abuse can cause digestive system symptoms (e.g. nausea, vomiting, indigestion, and diarrhea), 8 while long-term and excessive abuse can lead to liver function damage, endocrine and reproductive disorders,9,10 collagen fiber dysfunction, increased cardiovascular disease risk, and mental disorders.11–13 Research reports indicate that AAS abuse by amateur and professional athletes leads to complex regulation of gene expression involved in skeletal muscle metabolism, hypertrophy, inflammation, and regeneration. 4 Patients with more than three courses of AAS show abnormal liver and cardiac enzyme levels, and more than ten courses may cause acute liver infection or inflammation. 14 Long-term use of synthetic androgen steroid stimulants at supraphysiological doses reduces learning and memory abilities. 15 In addition, prolonged or excessive use of synthetic androgen steroids such as MA is likely to cause irreversible damage to the nervous system and psychological development in adolescents; these effects are often overlooked and may contribute to drug dependence. 16
In this study, human hepatocyte HepG2 cells were used as the research model.1,17 At the cellular and molecular levels, genetic changes after MA treatment of HepG2 cells were analyzed using bioinformatics approaches, including transcriptome sequencing, to elucidate the damaging effects and molecular mechanisms of MA abuse on the liver. In parallel, pathological examination of mouse liver tissues and blood biochemical analyses were conducted. Combined cellular and animal experiments were used to explore the effects of MA on liver cells and its potential mechanisms of action, accurately identify the characteristics of MA-induced liver injury, and provide a basis for the prevention and treatment of drug abuse.
Materials and methods
Reagents and instruments
MA (M77900, Jizhi Bio, Shanghai, China); Dulbecco modified Eagle’s medium (DMEM; C11995500BT, Gibco, USA); fetal bovine serum (FBS; 10099-141, Gibco, USA); trypsin (25200-072, Gibco, USA); penicillin–streptomycin (15070063, Thermo Fisher Scientific, MA, USA); TRIzol reagent (T9424, Sigma, Germany); Cell Counting Kit-8 (CCK-8) reagent (A311-02, Vazyme, Nanjing, China); alanine aminotransferase (ALT) assay kit (S03030, Rayto, Shenzhen, China); aspartate aminotransferase (AST) assay kit (S03040, Rayto); alkaline phosphatase (ALP) assay kit (S03038, Rayto); albumin (ALB) assay kit (S03043, Rayto); total cholesterol (CHO) assay kit (S03042, Rayto); triglyceride (TG) assay kit (S03027, Rayto); lactate dehydrogenase (LDH) assay kit (S03034, Rayto); urea nitrogen (UREA) determination kit (S03036, Rayto); creatinine (CREA) determination kit (S03076, Rayto); creatine kinase (CK) determination kit (S03024, Rayto).
Animals
Twenty 8-week-old male C57BL/6J mice were randomly divided into two groups (normal control (NC) and MA treatment), with 10 mice in each group. All animals were purchased from Jiangsu Jicui Yaoke Biotechnology (Nanjing, China) and housed in a specific pathogen-free environment (license number: SCXK (Su) 2024-0016). All protocols were approved by the Institutional Review Board of Nanjing University. Referring to the results of multiple studies and after sorting out the dosage and modeling time of MA in mice, this dose selection was based on published classic animal models used to study steroid hepatotoxicity. Therefore, this study chose a high sublethal dose and short-term exposure. The MA treatment group received 0.1 mg·g−1·d−1 via gavage daily for 14 consecutive days. After 14 days, all animals were euthanized and either injected with 30 μL/g afferden (JT0791, Jitian, Beijing, China) or exposed to high levels of carbon dioxide.
All animal care and procedures aimed at minimizing suffering were performed in accordance with the policies of the National Institutes of Health – Office of Laboratory Animal Welfare. The animal experiments were reviewed and approved by the Animal Ethical and Welfare Committee (IACUC) of the School of Life Sciences, Nanjing University (Nanjing, China) (approval number: IACUC-2009003; dated 16 September 2020). The methods were conducted in accordance with the Helsinki Declaration of 1975, as revised in 2024. The reporting of this study conforms to the Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) guidelines. 18
Cell culture and cell treatment
Human hepatocyte HepG2 cells were subjected to short tandem repeats (STR) analysis and tested for mycoplasma contamination. Cells were cultured in DMEM containing 10% FBS, 100 U/mL penicillin, and 0.1 mg/mL streptomycin and maintained in a humidified incubator at 37°C with 5.0% CO2. Referring to the experimental procedures of Zschiesche et al., 17 MA was added to the culture medium at a final concentration of 1 μg/mL (final solvent concentration of 0.1% EtOH/H2O), dissolved in EtOH/H2O (1:1, v/v), and the influence of endogenous steroid production in HepG2 cells was excluded.
CCK-8 analysis
HepG2 cells were evenly seeded into 96-well plates at a density of 6 × 10³ cells per well, with eight replicates per sample. When cell confluence reached 60%–90%, 1 μg/mL MA was added for treatment, and ethanol/water (1:1) was used as the NC group. After 12, 24, 36, 48, and 60 h, 100 μL of CCK-8 reagent prepared in serum-free DMEM was added and incubated for 2 h in the dark. The absorbance (OD) of each well was measured at 450 nm using an enzyme-labeled instrument.
Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay
HepG2 cells in the logarithmic growth phase were grouped and treated. The MA treatment group received MA at a final concentration of 1 μg/mL, whereas the NC group received an equal volume of solvent (0.1% EtOH/H2O). After 24 h, the culture medium was removed, and the cells were washed with PBS, fixed in 4% paraformaldehyde for 30 min, and permeabilized with 0.1% Triton X-100 in PBS for 15 min. TdT enzyme and fluorescently labeled dUTP were added according to the manufacturer’s instructions. Samples were incubated at 37°C for 1 h, followed by three washes with PBS to remove unbound dUTP and TdT. After mounting with an antifluorescence quenching agent, samples were observed under a fluorescence microscope and images were captured. Apoptotic cells showed positive fluorescence signals.
Transcriptome sequencing and analysis
Cells in the NC and MA treatment groups were pipetted and homogenized at a ratio of 1 mL TRIzol per 5 × 106 cells and then transferred to RNase-free centrifuge tubes. Samples were rapidly frozen in liquid nitrogen for 0.5 h and stored at −80°C. Transcriptome sequencing was performed by Majorbio (Shanghai, China). Gene Ontology (GO) enrichment analysis of differentially expressed genes (DGEs) was conducted using the online tool Metascape (http://metascape.org) in the molecular function (MF) category. RNA sequencing (RNA-seq) data quality was assessed using FASTQC. Transcripts per million were calculated by calculating the expression value of each gene. Sequencing was performed on the Illumina NovaSeq 6000 platform using paired-end sequencing (PE150), with a read length of 2 × 150 bp. Data filtering criteria required a Q30 base percentage >95.67%. Adapter sequences and low-quality reads were removed using fastp v0.23. The alignment rate exceeded 95%. Clean reads were aligned to the Homo sapiens GRCh38 reference genome using HISAT2 v2.2.1. The expression levels of genes and transcripts were quantitatively analyzed using RSEM. Gene set enrichment analysis (GSEA) was performed using GO (MF) gene sets with GSEA v4.1.0. As this study did not involve patients with clinical data, the requirement for patient information was waived.
Transmission electron microscopy (TEM) analysis
After completion of the cell and animal experiments, liver tissues were collected and stored at −80°C. Three mice from each group were randomly selected for electron microscopy. Liver samples (∼1 mm³) were excised, immediately immersed in 2.5% glutaraldehyde fixative at 4°C, and sent to Zhongke Baice Technical Service (Beijing, China) for TEM analysis. Samples underwent dehydration, immersion, embedding, sectioning, and staining. Observations and radiography were performed using TEM at a magnification of 6000×.
Hematoxylin and eosin (H&E) staining
Liver tissue samples were fixed in 4% paraformaldehyde for 48 h, rinsed with PBS, and dehydrated stepwise in ethanol. Samples were made transparent with xylene, embedded in paraffin, sectioned into 5-μm thick slices, and dewaxed to water. 19 After H&E staining, pathological changes in liver tissue were observed under a microscope.
Periodic acid–Schiff staining (PAS)
Based on the above procedure, tissue sample sections were placed in periodic acid solution for 5 min and then immersed in Schiff’s reagent for 15 min, followed by hematoxylin staining for 2 min. Sections were differentiated using hydrochloric acid–ethanol solution and dehydrated stepwise in ethanol. Slides were mounted with neutral resin, and pathological changes in liver tissue were observed under a microscope.
Detection of blood routine parameters in serum
After anesthesia, blood was collected from the abdominal aorta of mice, allowed to stand for 30 min, and centrifuged at 4°C at 2500 r/min for 10 min. The separated serum was stored at −80°C. Serum ALT, AST, ALP, ALB, CHO, TG, LDH, CREA, UREA, and CK were measured according to the manufacturers’ instructions.
Statistical analysis
All experiments were independently repeated at least three times. Data were analyzed using GraphPad Prism 8.0. Results were presented as mean ± SEM. Comparisons between two groups were performed using Student’s t-test. Multiple-group comparisons were analyzed using one-way analysis of variance followed by Tukey’s test. A p value <0.05 (*) was considered statistically significant; p value <0.01 (**) significant; and p value <0.001 (***) highly significant.
Results
Effect of MA on the proliferation and apoptotic activities of HepG2 cells
HepG2 cells were incubated with 1 μg/mL MA for 24 h to assess proliferation and apoptosis, as shown in Figure 1(a). Cell proliferation was measured using the CCK-8 assay. The results showed that cell proliferation significantly decreased to 88.01% after 24 h of MA treatment and further decreased to 62.37% after 60 h (p < 0.001) (Figure 1(b)). Apoptosis was evaluated using TUNEL staining. The fluorescence signal in the NC group was weak, with sparse and scattered distribution of green fluorescent dots, and the proportion of TUNEL+ cells was 4.94%. After MA treatment, the density of green fluorescent signals increased, and the proportion of TUNEL+ cells increased to 25.08% (Figure 1(c) and (d)). At a concentration of 1 μg/mL for 24 h, MA stably and reproducibly induced significant cytotoxicity (viability decreased to 88.01%) and early apoptotic signals (TUNEL positivity rate increased to 25.08%) while avoiding excessive short-term cell death that may obscure adaptive responses and molecular mechanisms. This condition is consistent with the simulation of “high sublethal dose residual exposure.”

Analysis of cell proliferation and apoptosis after metandienone (MA) treatment of HepG2 cells. (a) Flowchart of the experimental design. After incubating 1 μg/mL MA with HepG2 cells for 24 h, the proliferation and apoptosis of the cells were assessed. Control cells were treated with ethanol/water (1:1) (EtOH). (b) Effect of CCK-8 detection on the proliferation of HepG2 cells. (c, d) Effect of TUNEL staining on the apoptosis of HepG2 cells (c, scale = 100 μm) and quantitative analysis (d). HepG2 cells were treated with 1 μg/mL MA for 24 h, fixed in 4% paraformaldehyde, and labeled according to the TUNEL assay protocol. Nuclei were stained with DAPI (blue). Three independent fields were quantified using ImageJ. Values are presented as mean ± SEM. Statistical significance was determined using Student’s t-test in (b) and (d). *p < 0.05, **p < 0.01, ***p < 0.001. TUNEL: terminal deoxynucleotidyl transferase dUTP nick end labelling; CCK-8: Cell Counting Kit-8.
Morphology of HepG2 cells treated with MA via TEM
As shown in Figure 2, compared with the NC group, the overall ultrastructure of HepG2 cells after MA treatment was abnormal. The accumulation of lipid droplets and glycogen particles in the cytoplasm increased, the endoplasmic reticulum was dilated or ruptured, and apoptotic bodies or necrotic regions were observed. In the MA treatment group, mitochondrial morphology was abnormal, including swelling, increased volume, matrix dilution, blurred internal structure in some mitochondria, cristae rupture or disappearance, disordered arrangement, and even vacuolation. These changes indicate that MA causes damage to mitochondrial structure and metabolism in hepatocytes, which may further affect normal cellular metabolism and function.

TEM analysis of HepG2 cells after MA treatment (6000×, scale = 500 nm). MA: metandienone; TEM: transmission electron microscopy.
Analysis of transcriptome sequencing of HepG2 cells treated with MA
The volcano plot and scatter plot showed the statistical results of the classification of DGEs (Figure 3(a); Supplementary Figure 1). Compared with the NC group, there are a total of 221 significantly DGEs in the MA treatment group, among which the expression of 133 genes was differentially upregulated and that of 88 genes differentially downregulated. These significantly different genes formed distinct clusters in the cluster analysis, facilitating the identification of key biological modules and regulatory networks. GO classification statistical bubble analysis showed that DGEs were significantly enriched in biological processes such as oxidative stress response, inflammatory response, and lipid metabolism, suggesting that MA-induced liver injury is related to combined disturbances in redox balance and metabolic homeostasis (Figure 3(b)). Kyoto Encyclopedia of Genes and Genomes (KEGG) classification statistics column clarified that DGEs showed significant enrichment in inflammatory and immune regulation, lipid metabolism, and cancer-related pathways, suggesting broad effects on the neuro-immune-metabolic network. Interactions among these pathways may constitute the complex mechanistic network underlying MA-induced liver dysfunction (Figure 3(c)). mRNA expression levels of DGEs showed that after MA treatment, genes involved in inflammatory responses (PTGS2 and EGR1), oxidative stress (LAMP3, CXCL2, and CXCL3), and immune-related processes (FUT1, STX11, IL4R, IL11, IL10RB, and IL20RB) were significantly upregulated. In contrast, genes involved in inflammatory regulation (PLCD4), cell signal transduction (EDN2 and KCNJ3), and cell proliferation (GUSBP13) were significantly downregulated (Figure 3(d)). The changes in the expression of genes related to liver metabolism, inflammatory response, and oxidative stress after MA treatment were analyzed using RNA-seq data (Table 1). Upregulated genes were associated with certain pathways of enhanced or activated functions, while downregulated genes may reflect functional inhibition or suppression of metabolic pathways. Enrichment analysis of the GSEA gene set showed that genes associated with autophagy, endoplasmic reticulum stress, mitochondrial dysfunction, and oxidative stress (e.g. LAMP3) were significantly enriched in the MA treatment group. In contrast, genes involved in mRNA splicing, translation, and post-transcriptional regulation (e.g. APOBEC3H and CSDC2) were significantly inhibited (Figure 3(e) to (g)).

Transcriptomic analysis of HepG2 cells after metandienone (MA) treatment. (a) Scatter plots of HepG2 cells treated with MA. (b) Gene Ontology (GO) analysis of differentially expressed genes in HepG2 cells treated with MA. (c) KEGG pathway analysis of differentially expressed genes in HepG2 cells treated with MA. (d) Relative mRNA expression levels of enriched genes. (e–g) Gene set enrichment analysis (GSEA) of core enriched genes. Values are presented as mean ± SEM. Statistical significance was determined using Student’s t-test in (d). *p < 0.05, **p < 0.01, ***p < 0.001.
Changes in the expression levels of enriched genes.
Effect of MA on the morphological indicators of the mouse liver
Mice were randomly divided into the NC group and MA treatment group. MA (0.1 mg·g−1·d−1) was administered once daily for 14 consecutive days. The histological morphology of the liver and blood routine parameters were assessed (Figure 4(a)). H&E staining showed that the hepatocytes in the NC group were normally arranged with a clear structure. However, in the MA treatment group, the arrangement of hepatocytes was disordered; the hepatocytes were swollen; nuclei were constricted and fragmented; and obvious hepatocyte steatosis, necrosis, and inflammatory cell infiltration were observed. PAS staining showed that the glycogen distribution was uniform in the liver of the NC group, while in the MA treatment group, there was abnormal accumulation of liver glycogen, with increased glycogen granules, appearing as deep purple-red blocks or granules. This indicated that MA interfered with glycogen metabolism in the liver, resulting in impaired glycogen decomposition and utilization (Figure 4(b)). TEM showed that hepatocyte structure in the MA treatment group was abnormal; mitochondria were swollen; and apoptotic bodies and necrotic areas were observed (Figure 4(c)). These changes indicated that MA caused structural damage to the hepatocytes and affected their normal functions.

Effect of MA on liver morphology in mice. (a) Flowchart of the experimental design. Mice were randomly divided into the NC and MA treatment group. MA (0.1 mg·g⁻¹·d⁻¹) was administered intragastrically once daily for 14 consecutive days. (b) H&E staining and PAS staining of liver tissue (scale = 100 μm). (c) TEM analysis of liver tissue (×6000, scale = 500 nm). H&E: hematoxylin and eosin; MA: metandienone; PAS: periodic acid–Schiff; TEM: transmission electron microscopy.
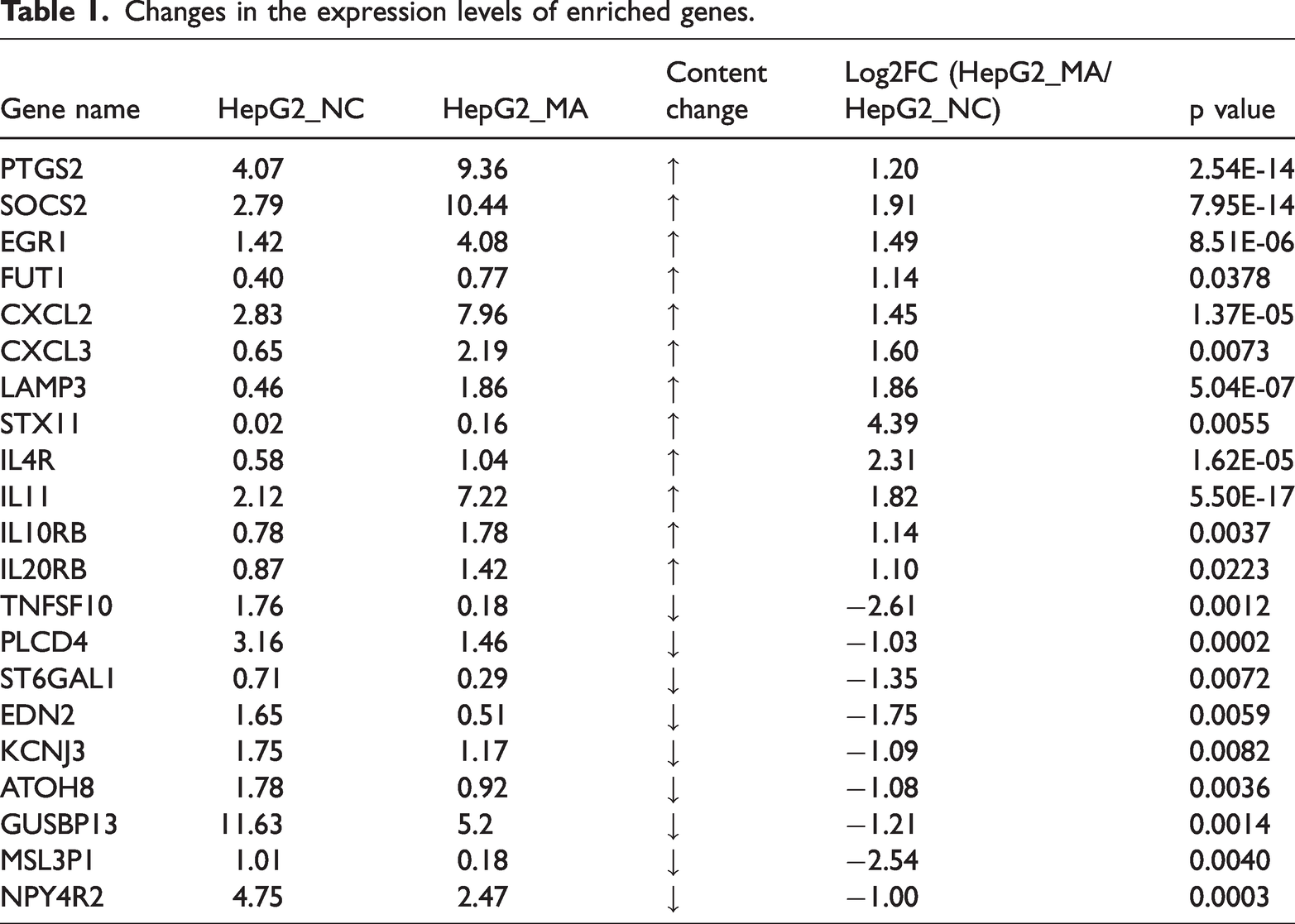
Effect of MA on serum biochemical indicators of liver function in mice
Serum liver function indicators revealed that the MA treatment group had higher ALT and AST activities than the NC group (Figure 5(a) and (b)), with a more pronounced increase in AST, suggesting hepatocyte injury and inflammation. The ALP level in the MA treatment group was not higher than that in the NC group (Figure 5(c)). The results suggested that synthetic steroid hormones such as MA may directly inhibit the synthesis or release of hepatocyte enzymes (e.g. ALP), and the overall liver function was inhibited. It has been reported that selective androgen receptor modulators (SARMs) cause cholestatic liver injury, and the biochemical pattern shows a mixed type of injury with normal or only slightly elevated ALP and bilirubin levels. Histological evidence has shown predominantly cholestatic injury with canalicular bile plugs and mild hepatocellular damage without significant fibrosis. 20 There was no significant difference in ALB levels between the MA treatment and NC groups (Figure 5(d)). This may be because ALB is synthesized by hepatocytes and has a longer half-life (∼20 days). Short-term or mild liver injury may not be sufficient to significantly reduce serum ALB levels. In chronic liver diseases, the decrease in ALB is more obvious. It is suggested that MA may partially counteract the inhibition of ALB synthesis caused by liver injury by promoting protein synthesis. Lipid function indicators showed that the MA treatment group had higher levels of CHO and TG compared with the NC group (Figure 5(e) and (f)), suggesting that the use of synthetic steroids such as MA caused abnormal liver function, leading to lipid metabolism disorders, reduced cholesterol excretion, accumulation of TG in the liver, and increased serum CHO and TG levels. The MA treatment group had a higher LDH level than the NC group (Figure 5(g)), suggesting that hepatocyte necrosis or increased membrane permeability led to increased release of LDH into the blood.
Effect of MA on serum biochemical indicators of liver function in mice. (a) ALT activity in serum. (b) AST activity in serum. (c) ALP activity in serum. (d) ALB levels in serum. (e) CHO levels in serum. (f) TG levels in serum. (g) LDH activity in serum. Values are presented as mean ± SEM. Statistical significance was determined using Student’s t-test in (a–g). *p < 0.05, **p < 0.01, ***p < 0.001. ALT: alanine aminotransferase; AST: aspartate aminotransferase; ALP: alkaline phosphatase; ALB: albumin; CHO: total cholesterol; TG: triglyceride; LDH: lactate dehydrogenase; MA: metandienone.
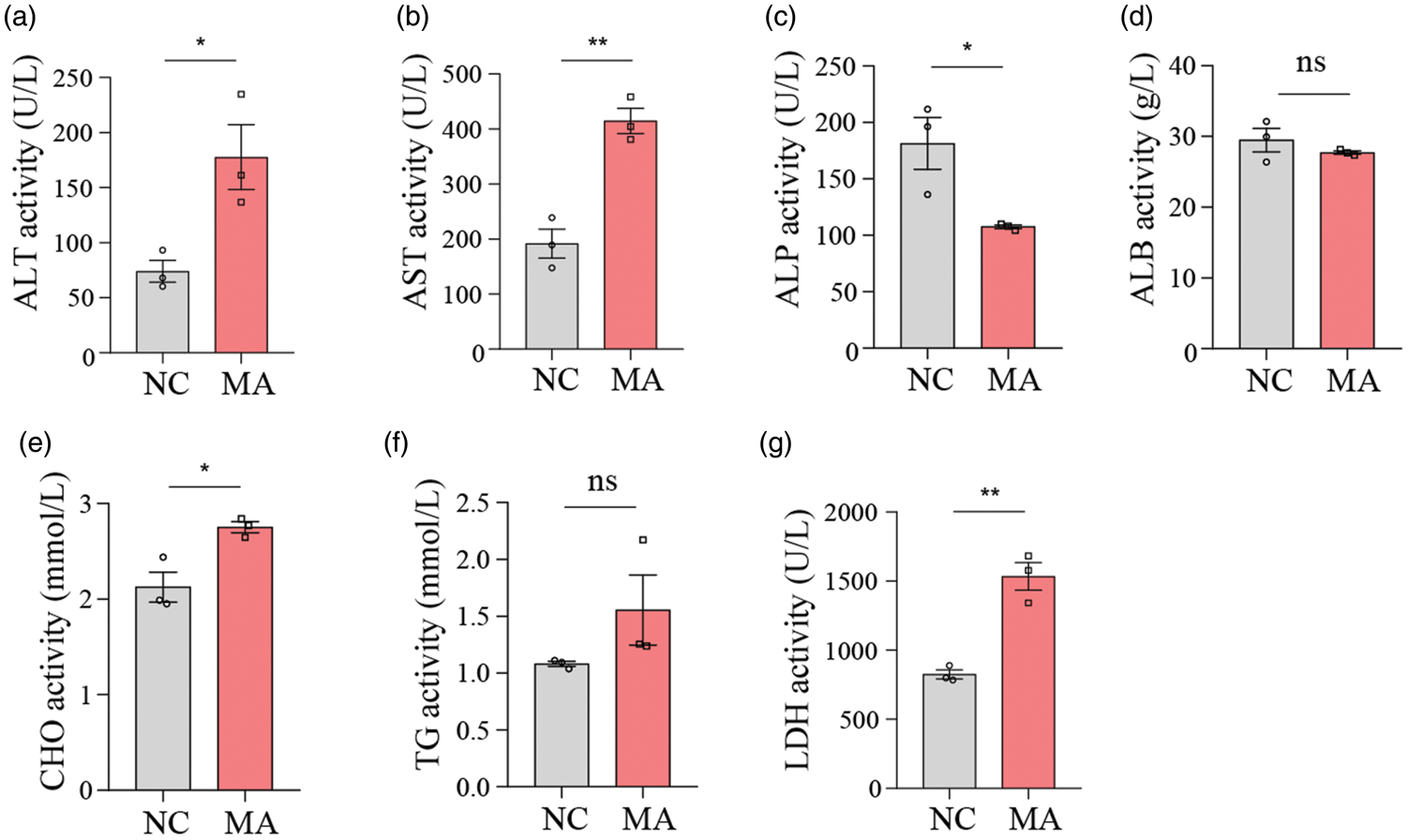
Effect of MA on other serum biochemical indicators in mice
Renal function indicators showed that the MA treatment group had higher levels of CREA and UREA than the NC group (Figure 6(a) and (b)). Myocardial enzyme spectrum functional indicators showed that there was no difference in CK activity between the MA treatment and NC groups (Figure 6(c)), suggesting that short-term or mild exposure to MA may not be sufficient to significantly change CK levels, and skeletal muscle was not damaged in the short term.
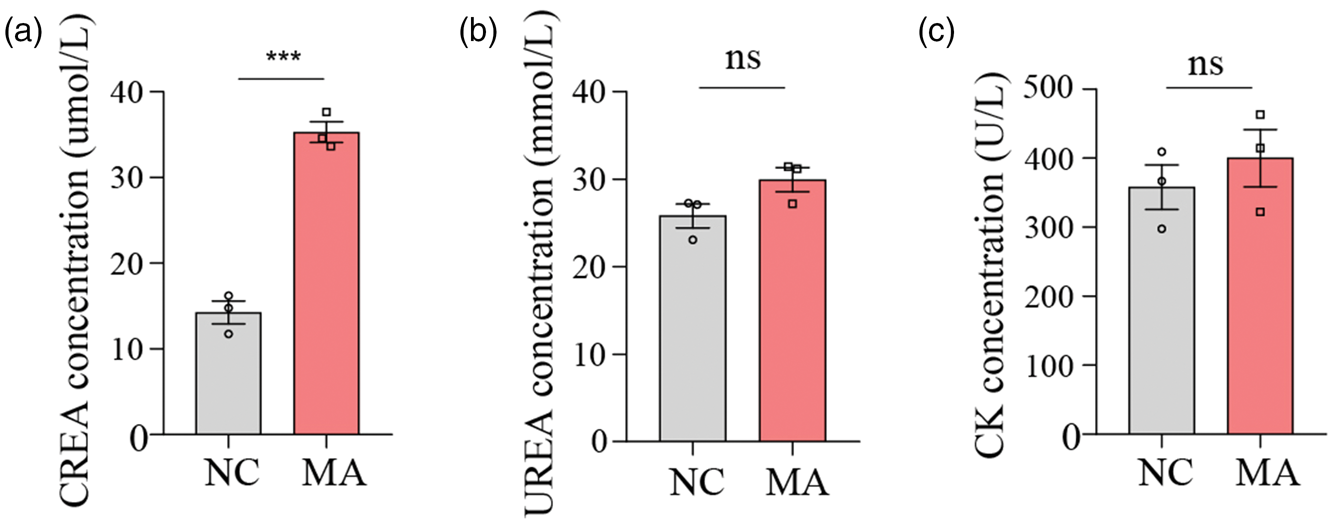
Effect of MA on additional serum biochemical indicators in mice. (a) CREA levels in serum. (b) UREA levels in serum. (c) CK levels in serum. Values are presented as mean ± SEM. Statistical significance was determined using Student’s t-test in (a–c). *p < 0.05, **p < 0.01, ***p < 0.001. MA: metandienone; CREA: creatinine; UREA: urea nitrogen; CK: creatine kinase.
Discussion
This study was conducted primarily at the cellular and mouse experimental levels. By comprehensively applying various experimental methods, such as CCK-8 assay, TUNEL staining, RNA transcriptome sequencing analysis, TEM, H&E and PAS staining, and serum biochemical index detection, the effects of MA abuse on the liver were systematically evaluated. The results showed that MA abuse decreased proliferation activity, increased apoptosis, and significantly changed the morphology of liver cells (e.g. vacuolar degeneration and abnormal accumulation of liver glycogen). Meanwhile, at the animal level, exposure to MA-induced acute liver injury is characterized by hepatocyte steatosis and inflammatory cell infiltration, accompanied by significant increases in serum ALT and AST, confirming the substantial hepatotoxicity of MA. It is worth noting that although typical liver injury markers were elevated, ALP activity did not increase simultaneously. A study reported that a 26-year-old male bodybuilder developed liver damage after taking high doses of AAS. After 12 weeks of discontinuation, bilirubin and ALP values were similar to those reported in the literature, while AST and ALT values were markedly higher than those reported in other studies. This was the first reported case of toxic hepatitis caused by AAS, mainly characterized by hepatocyte necrosis rather than intrahepatic cholestasis. 21 Case reports have also indicated that substances in SARM (ligand alcohol or ostarine) and/or “post-cycle therapy” may cause cholestatic liver injury in amateur athletes engaged in muscle training with prolonged recovery. These findings suggest an atypical biochemical pattern of liver injury associated with AAS: liver function tests may show only mild-to-moderate transaminase elevation, while ALP and bilirubin levels may remain normal or only slightly elevated. 20 The abovementioned studies suggest that the hepatotoxic mechanism of MA may differ from that of traditional cholestatic injury and may be related to the direct interference of drugs with the synthesis and secretion functions of hepatocyte enzymes (such as ALP). In addition, the half-life of ALB in humans is approximately 20 days, while in mice it is ∼1–2 days. In the 14-day subacute and sublethal injury model, the liver’s strong synthetic reserve is sufficient to maintain stable serum ALB levels. 22 Unless extensive (>70%–80%) acute liver necrosis occurs, ALB levels usually do not decrease significantly. However, our experiment did not measure serum bilirubin levels, which is a limitation of this study. This parameter will be included in future studies. Furthermore, transcriptome sequencing provides a systematic perspective for analyzing complex mechanisms. MA also caused abnormal changes in the expression of related genes in liver cells. The underlying mechanisms involve multiple processes, including oxidative stress injury, inflammatory responses, metabolic disorders, and mitochondrial dysfunction (e.g. cytochrome P450 enzyme system). 23 First, regarding oxidative stress, MA inhibited the activity of antioxidant enzyme systems (such as superoxide dismutase and catalase), leading to excessive accumulation of reactive oxygen species and triggering lipid peroxidation and DNA damage in liver cells. 24 Second, in terms of inflammatory responses, the accumulation of ROS activated inflammatory pathways such as NF-κB and TNF-β, increased the secretion of proinflammatory cytokines (such as TNF-α, IL-6, and IL-17), and induced immune cell infiltration, thereby exacerbating inflammatory damage in liver tissue. 4 In addition, regarding lipid metabolism, synthetic steroid stimulants can be absorbed through the skin. They cause abnormal accumulation of triglycerides in liver cells and hepatic steatosis by inhibiting fatty acid oxidation pathways while simultaneously activating lipid synthesis pathways. 25 These metabolic abnormalities further aggravate oxidative stress and inflammatory responses, eventually forming a vicious cycle leading to steatohepatitis, liver fibrosis, and even cirrhosis. 14 Therefore, these mechanisms are interrelated and influence each other, jointly constituting the complex pathological process of liver injury caused by MA abuse. The novelty of this study lies in the combination of in vitro and in vivo approaches to elucidate the hepatotoxic effects of MA, providing insights into its impact on liver metabolism, oxidative stress, and inflammatory responses. The hepatotoxic mechanism of MA is anchored in the interaction network among inflammation, metabolism, and oxidative stress, providing a clear target map for subsequent research.
Although cell and animal experiments have revealed some of the basic mechanisms underlying MA-induced liver injury, their relevance in humans requires further clinical validation. Because humans have more complex physiological systems and greater interindividual variability, factors such as genetic background, lifestyle, and underlying diseases may affect MA metabolism. Findings on the reversibility of steroid hormone–related side effects (such as those of MA) remain controversial: on the one hand, some studies suggest that organs can return to their initial state after discontinuation of androgen use; on the other hand, alterations in different biochemical parameters may lead to irreversible organ damage. 8 Therefore, future research should incorporate clinical cases 26 and advanced detection techniques—such as multiomics approaches (transcriptomics, proteomics, and metabolomics), 23 gene-editing technologies (e.g. clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated protein 9 (Cas9)), 27 and clinical imaging—to further elucidate the mechanisms by which MA affects the human liver. Meanwhile, epidemiological investigations should be conducted to examine the dose–effect relationship of MA abuse and identify risk factors for hepatotoxicity, thereby providing a stronger theoretical basis for developing more effective prevention and control strategies.
Although this study has systematically evaluated the effects of MA abuse on the liver and provides important evidence for understanding its hepatotoxic mechanisms, several limitations still inevitably exist. As a discovery study, the key pathways and genes identified by transcriptome analysis still require in-depth validation at the protein level and through functional experiments. Furthermore, the current model is subacute and does not reflect the consequences of long-term abuse. In practice, MA abuse may involve long-term low- or high-dose exposure. Long-term low-dose exposure may cause chronic and progressive liver injury, and short-term changes within a short period may not be fully manifested. Under high-dose exposure, acute and severe liver damage may occur. However, this study only involved short-term treatment and established a subacute and sublethal (14-day) toxicity model, without addressing the effects of long-term high-dose exposure. A study reported that 24 male Wistar rats were randomly divided into 3 groups: control, nandrolone (10 mg/kg), and nandrolone + N-acetylcysteine (150 mg/kg daily). After 6 weeks, nandrolone treatment led to increased levels of triglycerides, cholesterol, low-density lipoprotein, AST, ALT, and ALP, along with decreased HDL levels compared with the NC group. 28 It has been reported that mice were treated daily with 15 mg/kg nandrolone (ND) or testosterone (T) or oil (vehicle) for 19 days. Supraphysiological doses of AAS may cause long-term functional abnormalities, particularly in the heart and liver. ND and T increased H2O2 levels in the heart and liver relative to controls, and both AAS decreased glutathione peroxidase activity in the heart and liver. 29 Another study reported intraperitoneal administration of physiological solution, Deposteron (testosterone cypionate, 0.8 mg/kg), or Winstrol Depot (stanozolol, 1.8 mg/kg), with treatment lasting 1 month and injections performed twice weekly. 30 These data suggest that supraphysiological doses of steroids can cause considerable damage to nervous tissue, with associated ultrastructural and behavioral impairments. Other studies have reported that high doses of AAS are effective for increasing muscle mass but pose a risk of cardiovascular side effects. In a study, 40 female mice were treated with testosterone or sham pellets for 14 days, and echocardiography was used to evaluate vascular function and morphology. 31 Long-term users of AAS are at increased risk of severe cardiovascular events. In another study, 40 adult male mice were divided into 5 groups (n = 8 per group) and treated weekly with Deca-Durabolin (nandrolone decanoate) at 30 g/kg BW for 1 month (GI), 2 months (GII), or 3 months (GIII). The results showed significant degenerative changes in cardiac and gonadal tissues after 1 month of androgen abuse. These changes worsened with prolonged treatment and further deteriorated 6 weeks after treatment cessation in cardiac tissue, while gonadal tissue did not fully recover during this period. 32 Furthermore, previous experimental studies from the same research group showed that in the long-term treated GIII group, significant degenerative changes and structural disorders occurred in the heart, kidneys, and testicles. 33 The dosage used in humans cannot be directly translated to mice. Mahmood reported that it is important to consider strategies to reduce extrapolation errors, which may substantially affect animal safety and therapeutic responses. Interspecies scaling represents a rational approach with considerable potential in drug development. 34 Isenmann et al. 26 reported that a daily dose of 12 mg ecdysterone (0.15 mg/kg BW in an 80-kg volunteer) was administered in the normal-dose group (Ec1), while the high-dose group (Ec2) received 48 mg daily (0.6 mg/kg BW in an 80-kg volunteer). Therefore, long-term toxicity experiments are needed in the future. Experimental groups with different doses and exposure durations should be established to observe the pathological changes, functional alterations, and dynamic changes in gene expression in the liver under long-term MA exposure to comprehensively understand the development process and potential mechanism of MA-induced liver toxicity.17,35 However, these limitations do not diminish the discovery value of this study; instead, they highlight directions for future research.
This study provides important experimental evidence for an in-depth understanding of the mechanisms of MA-induced liver injury. The findings help raise public awareness of the harms of MA abuse, strengthen supervision of the use of performance-enhancing drugs in sports events, and strictly control their illegal addition in livestock and poultry breeding, thereby reducing abuse at the source. Moreover, clarifying the toxicity mechanisms provides potential targets for the development of targeted therapeutic drugs and regimens, improving liver function in patients with abuse and reducing the occurrence and progression of liver diseases. Furthermore, the results of this study may provide a scientific basis for formulating and improving antidoping testing standards for athletes to maintain the fairness and integrity of sports events. There remain many unknown aspects regarding the effects of MA on the liver, which require further in-depth investigation. Future studies should focus on addressing the limitations of this research. It is necessary to continuously expand the depth of research to provide more robust data for safeguarding public health, maintaining fairness in sports events, and regulating the healthy development of the livestock and poultry breeding industry.
Conclusion
As a typical 17-alkylated androgen anabolic steroid, hepatotoxicity caused by MA abuse has long been recognized as a clinical risk; however, its precise molecular mechanisms have not been fully elucidated. By integrating in vitro cell models with in vivo animal experiments and high-throughput transcriptomic analysis, this study revealed the characteristics and potential action networks of MA-induced liver injury from multiple perspectives.
In conclusion, this study suggests that hepatotoxicity induced by MA abuse results from the combined effects of metabolic dysfunction and an excessive inflammatory response. The mechanism may begin with the direct interference of MA with hepatocyte metabolism (particularly mitochondrial function and lipid metabolism), which subsequently triggers a strong inflammatory response and ultimately leads to liver damage. These findings provide new insights into the pathophysiology of AAS-induced hepatotoxicity and suggest that interventions targeting specific metabolic pathways or inflammatory processes may represent potential strategies for the prevention and treatment of drug-induced liver injury in the future. Further studies are needed to perform in-depth molecular and functional validation of the key hub genes and pathways identified by this transcriptomic analysis.
Supplemental Material
sj-pdf-1-imr-10.1177_03000605261448774 - Supplemental material for Metandienone abuse induces hepatotoxicity via dysregulated inflammation and metabolic dysfunction
Supplemental material, sj-pdf-1-imr-10.1177_03000605261448774 for Metandienone abuse induces hepatotoxicity via dysregulated inflammation and metabolic dysfunction by Yangyang Ye, Jing Li, Zuodong Chen, Xiaoming Xue and Jinzhong Xi in Journal of International Medical Research
Footnotes
Acknowledgments
Not applicable.
Authors’ contributions
Yangyang Ye was responsible for study design, experimental implementation, data collection and analysis, and manuscript drafting. Jing Li was responsible for technical guidance and study conception. Jinzhong Xi and Zuodong Chen were responsible for the literature review and manuscript revision. Xiaoming Xue was responsible for funding acquisition and final manuscript review.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Data availability statement
The study data will be made available upon reasonable request to the corresponding author.
Declaration of conflicting interest
The authors declare no potential conflicts of interest with respect to the research, authorship, and publication of this article.
Ethical considerations
Not applicable.
Funding
The study received funding from the Special Fund Project for Basic Scientific Research Business Expenses of Central Universities (LGZD202402, LGYB202504) and the “14th Five-Year Plan” Jiangsu Province Key Construction Discipline “Public Security Technology” (Su Teaching Research Letter 2022 No. 2, 235).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.