
Abstract
Rustrela virus (RusV), a recently discovered pathogen for domestic and wildlife animals, was identified as the causative agent of meningoencephalomyelitis in domestic cats and various zoo animals including lions. To analyze a past outbreak of increased mortality in lions and to reveal its possible etiological relationship with an RusV infection, this retrospective study re-evaluates 20 cases of lions originating from a zoo in Western Germany using archived formalin-fixed paraffin-embedded (FFPE) tissues. Animals with different neurologic signs were submitted for necropsy between December 1970 and April 1971. Eight lions (40%) suffered from non-suppurative meningoencephalomyelitis with RusV RNA and antigen detectable in the central nervous system (CNS). Twelve animals (60%) were negative for RusV. Eleven animals had an etiologically undetermined degenerative encephalomyelopathy characterized by dilated myelin sheaths, myelinophages, and spheroids. Eight of these 12 lions suffered from an erosive, lymphohistiocytic enteritis with nuclear inclusion bodies in enterocytes associated with parvoviral antigen and nucleic acid in the intestines, lymph nodes, and spleen, but not in the CNS. Five of the parvovirus-infected animals had a granulomatous inflammation in mesenteric lymph nodes that was also the only detectable lesion in one other lion. Acid-fast bacilli and Mycobacterium bovis DNA confirmed the diagnosis of tuberculosis. In summary, this study provides convincing evidence of the usefulness of long-term stored FFPE material for further investigations using immunohistochemistry, reverse transcription quantitative polymerase chain reaction, and in situ hybridization for resolving past disease outbreaks. It provides further insights into the epidemiology of infectious agents like RusV and parvovirus.
Keywords
Lions (Panthera leo) are classified as a vulnerable species according to the Red List of Threatened Species, as the number of adult individuals is decreasing in the wild. 68 For such a species, which is already under pressure from external anthropogenic factors such as habitat destruction and over-exploitation, infectious diseases pose a potential threat for their survival. An outbreak of canine distemper virus (CDV) in lions in the Serengeti, during which approximately a third of the population died, represents a well-studied and publicized example of the devastating effect of infectious diseases on this species. 76 Therefore, any additional knowledge about infectious agents that cause disease in free-ranging and captive lions is of great value regarding conservation efforts.45,50,61,66,67,71,83,105
CDV is regarded as the most important cause of encephalitis in lions, which was reported in both captive and free-ranging lions.2,7,33,64,65,76 Other important agents that cause neurologic signs in lions include rabies virus and rustrela virus (RusV, species Rubivirus strelense, family Matonaviridae), which was first described in lions in 2023.12,21
RusV is closely related to the human rubella virus and was identified in 2020 as the cause of a non-suppurative meningoencephalitis in a wide range of zoo animals in central and northern Europe and North America.10,29,73,99 Moreover, previous studies revealed RusV to represent the most likely cause of a neurologic disease entity called “staggering disease” in domestic cats (Felis catus).58,90,103 In a retrospective study of 3 lions suffering from non-suppurative meningoencephalitis from the 1980s, the newly discovered RusV was identified by using reverse transcription quantitative polymerase chain reaction (RT-qPCR), immunohistochemistry (IHC), and in situ hybridization. 21 Therefore, the present study was conducted to further investigate the etiologic role of RusV in an even older outbreak from the 1970s and to identify any associated or concurrently occurring disease patterns to gain further knowledge on the epidemiology of RusV. The study also aimed to prove the usability of formalin-fixed paraffin-embedded (FFPE) material to investigate and resolve past disease outbreaks.
To date, little is known about the pathogenesis and epidemiology of RusV. The role of RusV-infected animals with non-suppurative meningoencephalitis in the transmission of the virus to other animals remains undetermined, and they are considered dead-end hosts.10,73,99 Interestingly, RusV RNA was detected in the brains of yellow-necked mice (Apodemus flavicollis) that had been caught near a zoo in Germany in which RusV-infected animals including a donkey, capybara (Hydrochoerus hydrochaeris), and a red-necked wallaby (Macropus rufogriseus) were identified.10,73,69 In addition, the investigation of brain tissue from wood mice (Apodemus sylvaticus) examined for monitoring purposes, which were collected in the vicinity of RusV-infected cats in Sweden, yielded similar results. 58 Notably, these rodents did not have any histopathologic lesions in the brain, suggesting that they might serve as a virus reservoir.10,58,69
Descriptions of degenerative lesions in the central nervous system (CNS) in lions are rare and limited to a few captive animals. Changes in the spinal cord and brain stem, which were particularly pronounced in the white matter, were reported in 1 case of vitamin A deficiency. 55 Other investigations of hypovitaminosis A in lions reported hyperostotic changes of the cranium associated with cerebellar herniation.55,82,96 Idiopathic, non-compressive myelopathy has been described recently in a retrospective study of a captive population of Panthera species comprising axonal degeneration or focal myelomalacia. 61 In both young and adult cheetahs (Acinonyx jubatus), as well as in a few cases in mountain lions (Puma concolor) and leopards (Panthera pardus), encephalo- and myelomalacia were reported.16,100
Viral enteric infections in lions include norovirus and feline parvovirus (FPV) and canine parvovirus.11,25,56,60 FPV was described in the 1930s as the cause of panleukopenia in cats, a disease of high morbidity and mortality predominantly affecting juvenile cats.34,87,93 The first report of FPV in lions dates back to 1973. Since then, several cases of FPV infections have been described and the virus has been identified in both captive and free-ranging lions.25,36,60,86 In wild lions, there has been serologic evidence of canine parvovirus infections since the mid-1980s.6,11 The described lesions are similar to those in domestic cats.25,36,60,86 First evidence of canine parvovirus in dogs was reported in 1978 in Ontario, Canada, which is assumed to be the result of an evolutionary adaptation of FPV following spillover infection from cats.24,75,91,94
The first cases of Mycobacterium bovis (M. bovis) infections in lions were reported in both wild and captive individuals in the 1990s. 97 Respiratory tract-associated and presumably alimentary infections with lesions in the intestine and mesenteric lymph nodes were described.40,41,62,97 In large felids, reports of Mycobacterium avium infections that may cause a histopathologic indistinguishable picture are rare. 18
Cases of encephalitis of unknown etiology characterized by histopathological lesions indicative of a viral cause have also been described in various species.8,39,52,80 Despite further investigations using viral screening tools targeting virus-specific structures, the majority of these cases remain unresolved.8,39,52,80 This underlines the need for the development of pan-viral sensing tools that (1) are not based on the detection of already known viral structures and (2) enable the identification of new, previously unknown viruses. Therefore, this study additionally investigated the usability of anti-double-stranded RNA (dsRNA) antibodies as an alternative for the detection of virus infection. This is based on the hypothesis that dsRNA occurs during the replication of most viruses, offering an alternative approach to detect unknown viruses without targeting specific viral proteins or nucleic acids.22,23,38,46,49,57,84,101
The animals included in this study initially raised our attention due to the clustered occurrence of neurologic impairment and meningoencephalomyelitis of undetermined cause in lions originating from the same zoological gardens more than 50 years ago. Aiming to retrospectively clarify this outbreak of increased morbidity and mortality and its possible etiologic link to RusV, this study also provides further evidence for the usefulness of long-stored FFPE material to elucidate past disease outbreaks. Other diseases found during this re-evaluation were also further described and characterized, and the usability of dsRNA as a viral sensing tool was investigated.
Materials and Methods
Animals
This study investigated archived records including medical history, macroscopic and histologic findings, microbiologic reports, and the respective FFPE material from 20 lions (Supplemental Table S1; cases 1-20) originating from a zoo in Western Germany that were submitted in 2 groups. The first group, consisting of 8 animals (cases 1-8), was examined in December 1970; the second group, consisting of 12 animals (cases 9-20), was examined in April 1971.
Histology
In this study, all organs that had been archived as FFPE material were re-examined. The organ samples had been obtained during necropsy and subsequently fixed in formalin. It is noteworthy that the spectrum of organs differed considerably between animals due to non-standardized sampling protocols. Taken together, samples from the brain [n = 19: cerebrum (n = 19), cerebellum (n = 14), medulla oblongata (n = 17), mesencephalon (n = 8), pons (n = 5)], spinal cord (n = 19); kidney (n = 17); liver (n = 16); small intestine (n = 14); lung (n = 11); mesenteric lymph node (n = 7); spleen (n = 6); heart (n = 4); skin (n = 4); adrenal gland (n = 3); pancreas (n = 2); and eye, nose, thyroid gland, stomach, and large intestine (n = 1, respectively) were examined. A detailed list of histologically investigated organs for each animal is provided in Supplemental Table S1. Specified anatomic localizations of the brain and spinal cord samples were not recorded. Histologic examination was performed on hematoxylin and eosin-stained sections that had been previously processed through standard procedures. Ziehl-Neelsen stain was performed on selected slides for visualization of acid-fast bacteria. Luxol fast blue-Nissl stain was applied on selected CNS sections to demonstrate demyelination and chromatolysis.
Immunohistochemistry
IHC was performed to elucidate potential infectious etiologic agents, to characterize degenerative changes, and to investigate the expression of dsRNA, as described previously. 31 Primary antibodies, targeted proteins, the supplier, clonality, host species, performed pretreatment, and dilution of primary antibodies are summarized in Table 1. Screening for viral antigens was performed by the use of antibodies specific for RusV capsid antigen, 58 parvovirus capsid antigen cross-reacting with viruses causing enteritis in various species, 77 CDV nucleoprotein, 43 and feline leukemia virus (FeLV) envelope protein. 42 As previous studies on RusV and its distribution in various infected animal species, including lions, have shown that the virus appears to be restricted to the CNS in dead-end hosts, investigation for RusV antigen was restricted to FFPE samples from the CNS of all animals.10,21,73,99 In contrast, immunohistochemical screening for the presence of parvovirus antigen was performed on all available tissues from all animals. Furthermore, immunohistochemical testing for CDV and FeLV antigens was performed on CNS tissues of all 20 animals. In addition, lungs (cases 1, 3, 5, 7, 8, 12, and 16-20), lymph nodes (cases 5, 9-11, 15, 16, and 20), spleens (cases 1, 4, and 7), and small intestines (cases 1-3, 5, 8, 10-12, 14, and 16-20) were tested for CDV antigen. IHC for the presence of FeLV was performed on lymph nodes (cases 5, 9-11, 15, 16, and 20) and spleens (cases 1, 4, and 7).
The expression of viral dsRNA was investigated by the use of anti-dsRNA antibodies J2, 13 K1, 13 and 9D5, 13 on spleens, lymph nodes, intestines, and CNS tissues from all animals included in this study. To confirm the specificity of dsRNA labeling, CNS sections from 1 RusV-positive lion with a positive dsRNA signal (case 8) and 1 RusV-negative lion without dsRNA labeling (case 13) were pretreated with RNase III (Ambion RNase III, Invitrogen, Thermo Fisher Scientific, Vilnius, Lithuania), that specifically degrades dsRNA molecules, prior to anti-dsRNA antibody incubation.
Antibodies used for immunohistochemistry.
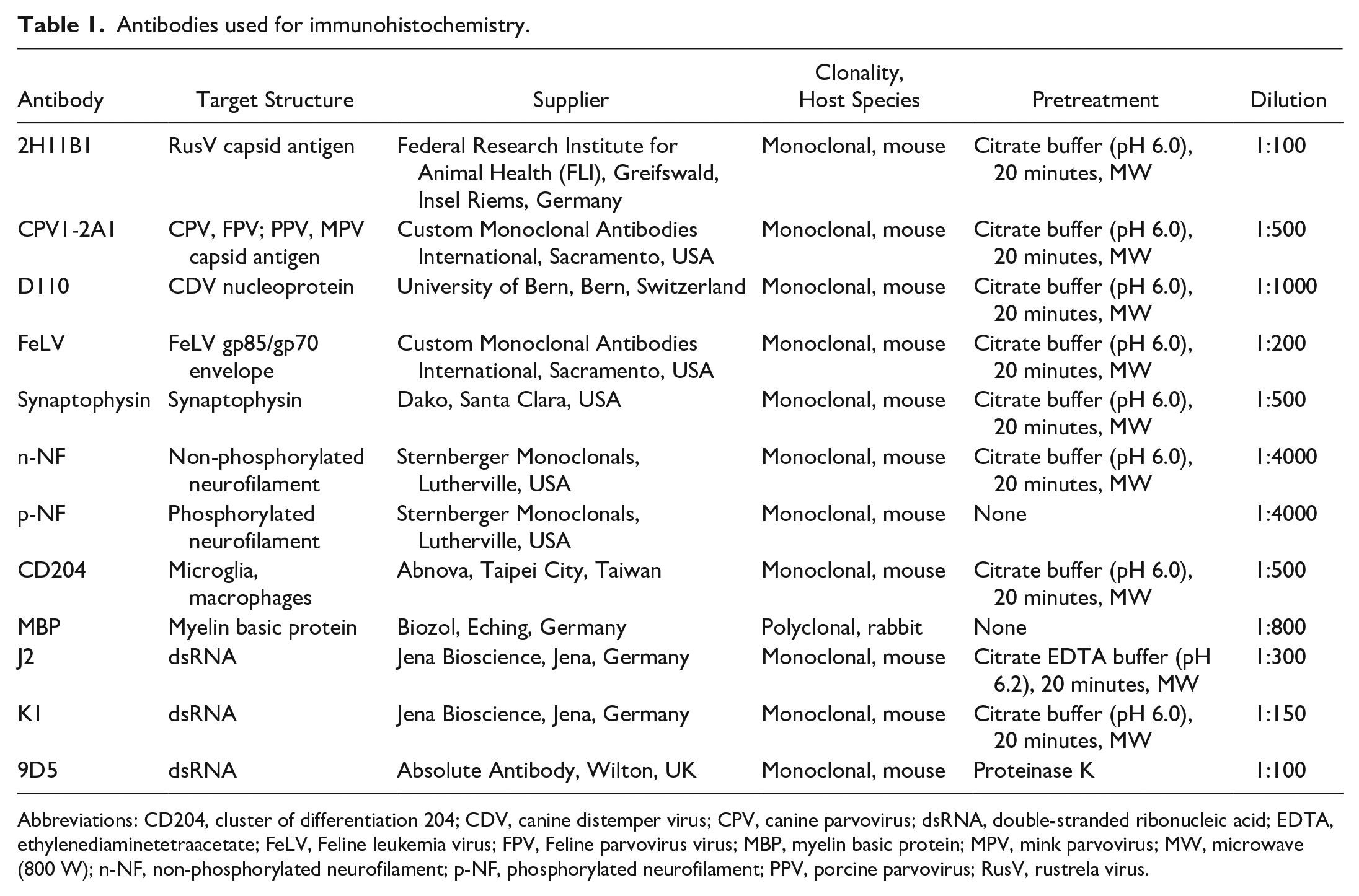
Abbreviations: CD204, cluster of differentiation 204; CDV, canine distemper virus; CPV, canine parvovirus; dsRNA, double-stranded ribonucleic acid; EDTA, ethylenediaminetetraacetate; FeLV, Feline leukemia virus; FPV, Feline parvovirus virus; MBP, myelin basic protein; MPV, mink parvovirus; MW, microwave (800 W); n-NF, non-phosphorylated neurofilament; p-NF, phosphorylated neurofilament; PPV, porcine parvovirus; RusV, rustrela virus.
For the characterization of degenerative lesions of lions, which were diagnosed with encephalomyelopathy, CNS tissues were examined using an anti-phosphorylated neurofilament (p-NF), 47 an anti-non-phosphorylated neurofilament (n-NF) antibody, 47 an anti-myelin basic protein antibody, 27 an anti-synaptophysin antibody, 31 and an anti-cluster of differentiation 204 48 antibody for visualization of microglia and/or macrophages.
After deparaffinization and rehydration, blocking of endogenous peroxidase activity was achieved by incubation in 85% ethanol and 0.5% hydrogen peroxidase for 30 min at room temperature. Heat-induced or proteolytic antigen retrieval was performed depending on primary antibodies (Table 1). The sections were either pretreated with the respective buffer for 20 minutes in a microwave (800 W) or incubated with Proteinase K (0.3 mg proteinase K [Roche Diagnostics, Mannheim, Germany]) diluted in 1000 µL phosphate-buffered saline (PBS, pH 7.1) for 7 minutes at room temperature. No antigen retrieval was performed for the anti-myelin basic protein and anti-p-NF antibodies. To reduce non-specific binding, sections were blocked with inactivated normal goat serum (diluted 1:5 in PBS) for 30 min at room temperature. For dsRNA degradation, CNS tissues from 2 lions were pretreated with 15 units of RNase III per section for 60 minutes at 37°C prior to the application of dsRNA antibodies. Thereafter, primary antibodies were diluted to their respective concentration (Table 1) in PBS with added bovine serum albumin, applied to the slides, and incubated overnight at 4°C. Negative controls were incubated either with Balb/c mouse ascitic fluid (diluted 1:1000 in PBS) or with rabbit serum (diluted 1:3000 in PBS) instead of the primary antibody. After application of the secondary biotinylated goat anti-mouse antibody (Vector Laboratories, Newark, USA) or biotinylated goat anti-rabbit antibody (Vector Laboratories) (diluted 1:200 in PBS) and incubation for 60 minutes at room temperature, the avidin–biotin-peroxidase complex (Vectastain ABC Kit Standard, Vector Laboratories, Newark, CA, USA) was applied for 30 minutes at room temperature. By using 3,3’-diaminobenzidine tetrahydrochloride (0.05%, Sigma Aldrich Chemie, Taufkirchen, Germany) with the addition of 0.03% hydrogen peroxide, immunopositive reactions were visualized. Finally, sections were counterstained with Mayer’s hematoxylin (Roth, Karlsruhe, Germany). Slides were analyzed semiquantitatively, and immunoreactivity was categorized as mild (<33% immunopositive cells), moderate (33%-66% immunopositive cells), and marked (>66% immunopositve cells), independent of the investigated tissues and antibodies. The used positive controls included CNS tissues from an RusV-infected lion from a previous study, 21 the intestine from a cat infected with parvovirus, a persistently CDV-infected DH82 cell pellet with more than 90% virus-positive cells, lymphoid tissue from a cat infected with FeLV, brain tissue from a dog with degenerative changes for the demonstration of p-NF, n-NF, and myelin basic protein, adrenal gland from a dog for visualization of synaptophysin, a lymph node from a dog for labeling microglia and/or macrophages using CD204, and a cell pellet from experimentally Umatilla virus-infected quail fibroblasts as a positive control for dsRNA. Furthermore, archived small intestine samples from 3 lions without histopathologic lesions (cases 21-23) were included as negative controls for the investigation of dsRNA expression in the small intestine.
RNA Extraction and Reverse Transcription Quantitative Polymerase Chain Reaction of the Central Nervous System
Nucleic acids for the detection of RusV RNA were extracted from FFPE scrolls using the RNeasy FFPE Kit (Qiagen, Hilden, Germany) with Deparaffinization Solution (Qiagen) in accordance with the manufacturer’s instructions. Detection of RusV RNA was performed with the RT-qPCR assay panRusV-2a, 90 which is a modification of the previously published assay panRusV-2 with an altered TaqMan probe (RusV_257_P; 5’-[FAM]TGAGCGACCACCCAGCACTCCA[BHQ1]-3’). 58 Primers and polymerase chain reaction (PCR) conditions were identical to those described previously. 58
Sequencing and Phylogenetic Analysis of Rustrela Virus
Generation of partial RusV p150-encoding sequences was attempted for all RusV-positive animals by Sanger sequencing of 4 overlapping conventional RT-PCR amplicons of 142 to 191 base pair lengths, as described previously. 21 Each amplicon was sequenced in both directions. Raw sequences were trimmed for quality and primer sequences and assembled to result in a sequence of 409 nucleotides (nt) in length (representing genome positions 100-508 of RusV reference genome MN552442.2). The final sequences are deposited in GenBank under accession numbers PP910114 to PP910120.
Phylogenetic analysis of these sequences was performed together with all RusV sequences derived from GenBank for which the respective sequence stretch of the 409 nt length was available (n = 68). Following nucleotide sequence alignment using MUSCLE 3.8.425 (available in Geneious Prime 2021.0.1, Auckland, New Zealand), a neighbor-joining phylogenetic tree was calculated using MEGA (version 11.0.13; Tamura-Nei method with a gamma distribution; shape parameter = 1; 1000 bootstrap replicates). 63
Metagenomic Sequencing and Sequence Analysis of Intestine
For the detection of unexpected/unknown viruses in FFPE scrolls from the intestine, RNA was extracted from samples of 9 lions with enteritis (cases 10-12, 14, and 16-20) using the miRNeasy FFPE Kit after application of Deparaffinization Solution (both Qiagen, Hilden, Germany) in accordance with the manufacturer’s instructions. After quality control with an RNA 6000 pico assay on an Agilent 2100 Bioanalyzer (both Agilent, Waldbronn, Germany), the 2 best-suited RNAs (originating from cases 11 and 14) were chosen for library preparation and sequencing according to the protocol described previously with a few modifications. 104 Briefly, after concentrating RNA with the aid of Agencourt RNAClean XP Beads (Beckman Coulter, Krefeld, Germany), complementary DNA (cDNA) was prepared with the SuperScript IV First-Strand cDNA Synthesis System (Invitrogen, Darmstadt, Germany) followed by second strand synthesis with an NEBNext Ultra II Non-Directional RNA Second Strand Synthesis Module (New England Biolabs, Frankfurt am Main, Germany). The obtained double-stranded DNA was fragmented to a target size of 200 base pairs with a Covaris M220 Focused ultrasonicator (Covaris, Woburn, MA, USA) and subsequently purified using Agencourt AMPure XP Beads (Beckman Coulter, Brea, CA, USA). The fragmented DNA was then used as input for library preparation with the NEBNext Fast DNA Library Prep Set for Ion Torrent (New England Biolabs). Finally, the sequencing libraries were size selected with Agencourt AMPure XP Beads, their quality was verified with a High Sensitivity DNA Kit on the Bioanalyzer 2100 (both Agilent) and quantitated with the QIAseq Library Quant Assay Kit for Ion Torrent (Qiagen). The sequencing-ready libraries were prepared for sequencing with the Ion Chef and sequenced on an Ion Torrent S5 XL using Ion 540 chips and respective reagents (all Thermo Fisher Scientific, Darmstadt, Germany).
For taxonomic assignment of the reads, the obtained data sets were analyzed using a modified RIEMS workflow. 78 For further characterization, viral and unclassified reads were assembled using the 454 software suite (v3; Roche Diagnostics), and the taxonomic assignments of the sequences were confirmed by BLASTn against the complete GenBank database. 1 The obtained sequence data sets are available from ENA under project accession number PRJEB77040.
DNA Extraction and Polymerase Chain Reaction for Mycobacterium bovis
DNA extraction from FFPE lymph node scrolls followed a modified protocol from Weiss and colleagues. 102 In brief, the paraffin was extracted using xylol (1 incubation for 1 hour at 37°C, 2 incubations for 30 minutes at 60°C), and the tissue pellet was washed twice with 96% ethanol. After drying the pellet, the DNA was extracted using the Blood and Tissue Kit (Qiagen). Subsequently, 2 real-time PCRs were conducted detecting (1) the insertion element IS1081 to identify members of the Mycobacterium tuberculosis-complex and (2) the deletion of the RD4 region to identify M. bovis. 30
Chromogenic In Situ Hybridization for Rustrela Virus
To confirm the immunohistochemical results for RusV, chromogenic in situ hybridization (CISH) was used as described. 74 Three RusV-positive lions with immunopositive signals for RusV capsid antigen (cases 2, 5, and 6) as well as 3 RusV-negative lions lacking RusV antigen expression (cases 13, 15, and 19) were investigated using CISH. As indicated above, only CNS tissues were examined. CISH was performed using the commercially available ViewRNA ISH Tissue Core Reagents and ViewRNA ISH Tissue Fast Red Module (Invitrogen by Thermo Fisher Scientific, Vienna, Austria) and a commercially produced ViewRNA Type 1 Probe Set (Invitrogen by Thermo Fisher Scientific) according to partial p150-encoding RusV sequences (GenBank accession numbers OQ319479 and OQ319480), generated in a previous study. 21 The day before CISH, the slides were heated at 60°C for 1 hour. On the following day, the sections were first deparaffinized and rehydrated. This was followed by heat pretreatment with Pretreatment Solution at 85 to 90°C for 10 minutes. For subsequent proteolytic digestion and fixation, the slides were incubated for 10 minutes at 40°C with Protease QF diluted 1:100 in PBS. Prior to probe set hybridization, fixation in 10% neutral-buffered formalin at room temperature for 4 minutes was followed by an additional pretreatment with 0.2 M hydrochloric acid for 10 minutes at room temperature. The probe was applied in a dilution of 1:50 in Probe Set Dilution and incubated for 4 hours at 40°C. The application of Probe Set Dilution without the addition of the probe was used to generate negative controls. The visualization of positive signals was achieved by incubation with Pre-amplifier Mix, Amplifier Mix, and Label Probe 1-AP Solution at 40°C. This was followed by the application of AP Enhancer Solution for 5 minutes at room temperature. Then, Fast Red Substrate was added to the sections, and these were incubated for 60 minutes at room temperature. Finally, counterstaining was achieved by the use of Mayer’s hematoxylin (Roth).
Chromogenic In Situ Hybridization for Parvovirus
For confirmation of histologic and immunohistochemical findings indicating a parvovirus infection, CISH was applied to 2 lions that tested positive for parvovirus by IHC (cases 10 and 11) and to screen CNS FFPE tissues from animals with degenerative encephalomyelopathy (cases 10-20) for possible parvovirus infection. CISH was performed using a 222 base pair long digoxigenin-labeled RNA antisense probe targeting the canine parvovirus viral protein 2 (VP2) gene, as previously described. 77 Briefly, slides were deparaffinized, rehydrated, and rinsed in diethyl pyrocarbonate-treated water and PBS. Proteolytic digestion was performed by applying 1 μg/ml proteinase K (Roche Diagnostics) at 37°C for 15 minutes. Following fixation, acetylation, and prehybridization, hybridization was performed in a humid chamber with a probe concentration of 100 ng/ml at 52°C overnight. Afterwards, slides were rinsed twice in 6× standard saline citrate with 45% formamide for 15 minutes at 42°C, followed by washing twice in 2× standard saline citrate for 5 minutes at room temperature. Subsequent RNase treatment was followed by several washing steps in 2× standard saline citrate at room temperature and in 0.2× standard saline citrate at 50°C. Digoxigenin-labeled probes were identified with an anti-digoxigenin antibody conjugated to alkaline phosphatase (Merck, Isernhagen, Germany). Visualization of positive reactions was achieved by using nitroblue tetrazoliumchloride (Merck, Beijing, China) and 5-bromo-4-chloro-3-indolyl phosphate (X-Phosphate, Merck, India). Negative controls were generated by incubation with hybridization buffer without probe application.
Transmission Electron Microscopy, Pop-Off Technique
Material from the small intestine from 1 lion (case 11) with suspected virus inclusion bodies was further investigated using transmission electron microscopy (TEM), as previously described. 51 A 5-µm thick section was cut from the FFPE block, mounted on a glass slide and stained with hematoxylin and eosin without subsequent covering. The area chosen for TEM was marked on the backside. First, the slide was washed twice with absolute ethanol for 5 minutes each and incubated for 2 minutes in propylene oxide and absolute ethanol (ratio of 1:1). Afterwards, the sample was coated with pure propylene oxide for a few seconds, subsequently covered with propylene oxide and epoxy resin (ratio 1:1) for 20 minutes, and finally with pure epoxy resin. A gelatine capsule filled with epoxy resin was placed upside down on the area of interest. Polymerization was performed by incubating the slide for 1 hour at 35°C, 1 hour at 45°C, and overnight at 85°C. Thereafter, the epoxy block was removed by dipping the slide in liquid nitrogen. Under vision control, the block was further trimmed for ultra-sectioning. Afterwards, the 70 nm thin sections were mounted on 200 mesh copper grids and investigated by TEM (EM 906, Zeiss, Oberkochen, Germany). Images were taken with a CCD camera (sharp:eye, Tröndle, Moorenweis, Germany).
Results
The investigation of archived FFPE tissue of lions yielded 4 different disease entities, including viral and bacterial infections as well as degenerative alterations, which are presented and discussed in detail in the following sections. The overall results are summarized in Table 2. Furthermore, Supplemental Tables S1 and S2 provide detailed findings with respect to the histopathologic findings and investigated infectious agents.
Overview of disease pattern in investigated lions.
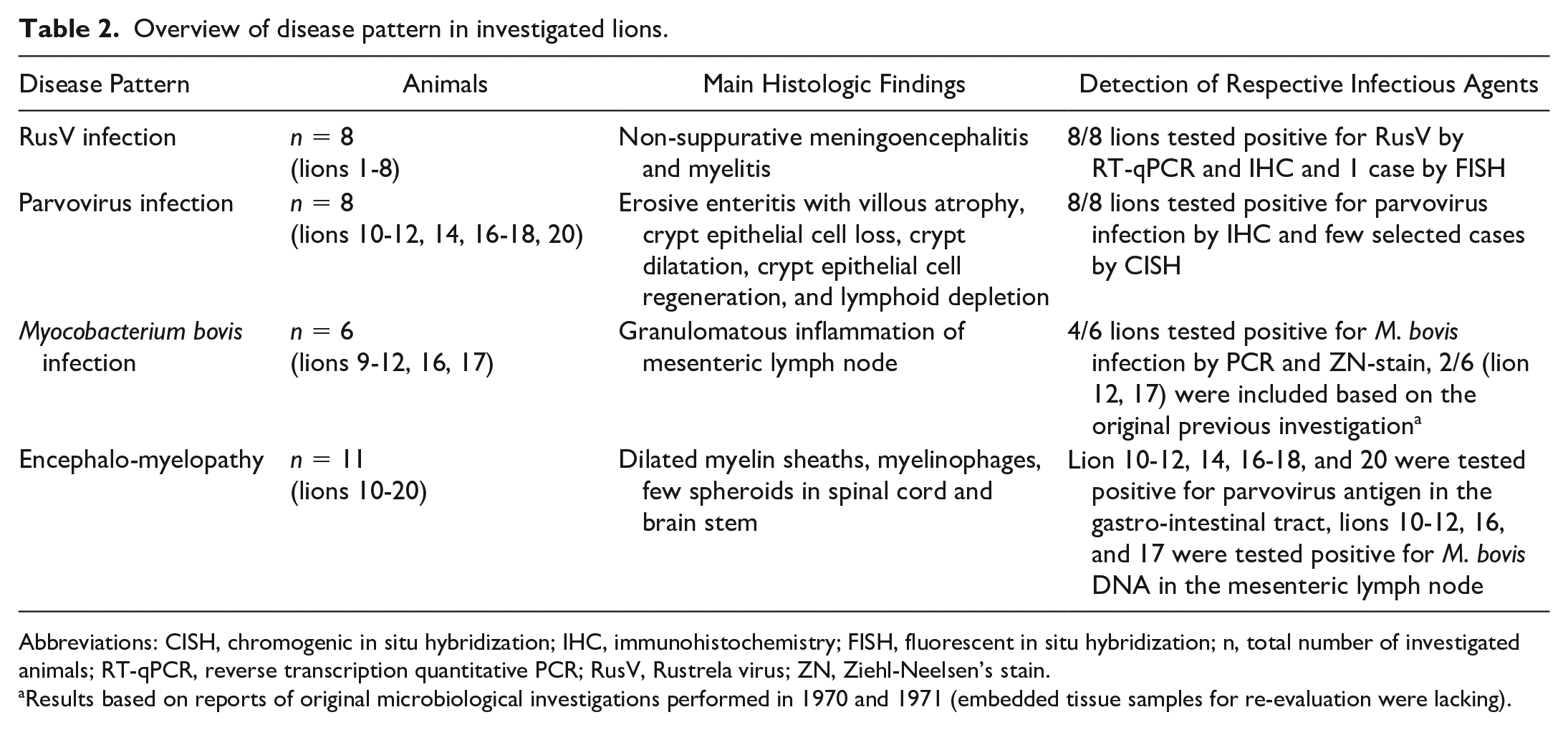
Abbreviations: CISH, chromogenic in situ hybridization; IHC, immunohistochemistry; FISH, fluorescent in situ hybridization; n, total number of investigated animals; RT-qPCR, reverse transcription quantitative PCR; RusV, Rustrela virus; ZN, Ziehl-Neelsen’s stain.
Results based on reports of original microbiological investigations performed in 1970 and 1971 (embedded tissue samples for re-evaluation were lacking).
Recorded Clinical Signs and Macroscopic Findings
According to the recorded medical history, the clinical signs of animals in the first group submitted in December 1970 (cases 1-8) included apathy, recumbency, epileptiform seizures, and spontaneous death, whereas lions in the second group submitted in April 1971 (cases 9-20) suffered from hind limb weakness. The reports mentioned no indications of enteritis or diarrhea.
According to the archived necropsy records, 1 lion (case 6) was in poor body condition. Four animals in the second group (cases 10 and 17-19) had enteritis with hyperemic intestinal mucosa, pasty to liquid dark brown to green contents, and swollen intestinal lymph nodes. Three lions (cases 10-12) had caseous lymphadenitis of mesenteric or ileocecal lymph nodes. Except for cases 1, 5-7, and 16, all animals had moderate to severe intestinal endoparasitosis with nematodes.
Histology
Histologic examination of the brain revealed multifocal to coalescing, mild to severe, lymphohistiocytic, predominantly perivascular, encephalitis in 8 lions (cases 1-6, 8, and 11). In addition, 4 of these animals also had meningitis (cases 1, 2, 5, and 8) and/or vasculitis (Fig. 1a), characterized by abundant mural accumulation of mononuclear cells (cases 2, 5, 6, and 8). Due to the heterogeneous spectrum of samples, a statement about a potentially different tropism of lesions in different brain areas was not possible. Except for 1 animal (case 11), glial nodules, microgliosis, and/or astrogliosis were observed in the brain of all animals that had encephalitis. Neuronal alterations, including marginalization/loss of Nissl’s substance (chromatolysis) and/or necrosis, were observed in 6 encephalitic animals (cases 1-6). One of these 6 animals (case 2) had severe, laminar, eosinophilic, neuronal necrosis within the cerebrum.
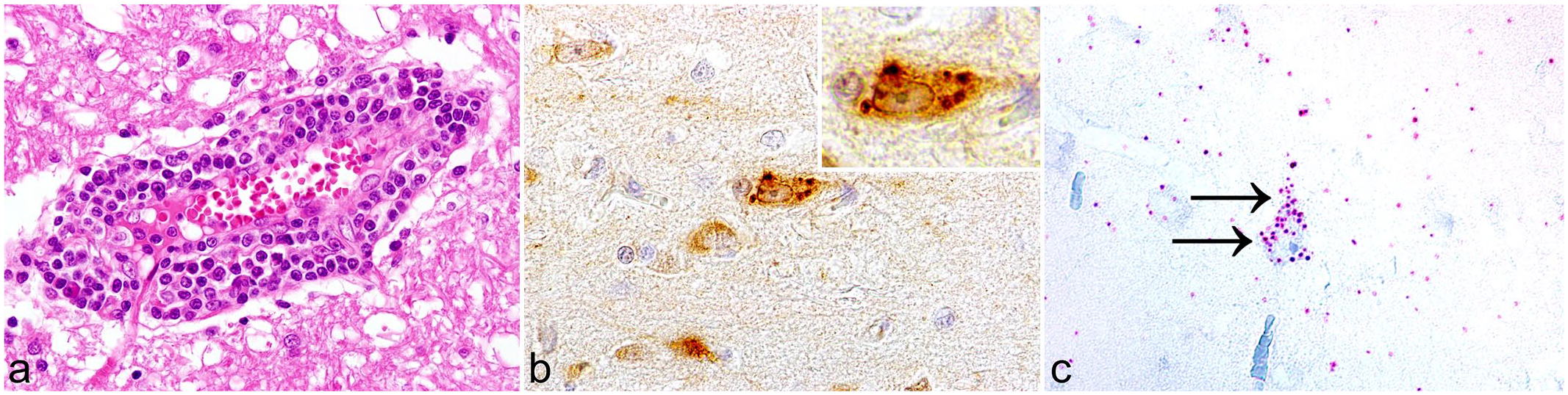
Rustrela virus (RusV) infection in a lion, cerebrum. Case 5. (a) Moderate, multifocal, lymphohistiocytic, perivascular encephalitis and vasculitis. Hematoxylin and eosin. (b) RusV capsid antigen is predominantly found within cerebral cortical neurons. Inset: Immunopositive reaction is characterized by a dark brown, cytoplasmic, granular signal within perikarya. Immunohistochemistry for RusV. (c) RusV RNA is detected within a cerebral cortical neuron (arrows). Chromogenic in situ hybridization for RusV.
The spinal cord of 5 animals (cases 1, 2, 4, 5, and 8) had mild, multifocal, lymphohistiocytic, perivascular poliomyelitis accompanied by formation of a few, subtle glial nodules and a few chromatolytic or necrotic neurons. Two lions (cases 1 and 8) had concurrent lymphohistiocytic meningitis, and 1 lion (case 5) had concurrent vasculitis. In 1 animal (case 20), additional mild, focal, lymphocytic, perivascular leukomyelitis was diagnosed. Occasionally, single-dilated myelin sheaths, sometimes admixed with myelinophages, were observed (cases 2, 3, 5, and 7). Three animals (cases 3, 7, and 8) had additional calcifications of the dura mater. The spinal cord was histologically unremarkable in 1 animal (case 9).
In 11 lions (cases 10-20), degenerative changes of a varying degree were found in the spinal cord and the caudal brain stem. In ascending funiculi, dilated myelin sheaths, myelinophages, and occasional spheroids were seen in the gracile and cuneate fasciculi as well as in the dorsal and ventral corticospinal tracts. Similar findings were also observed in descending funiculi, including the lateral and ventral corticospinal, the rubrospinal, the vestibulospinal, and tectospinal tracts (Fig. 2a). Occasionally, single chromatolytic or necrotic neurons were present in the ventral horns of the spinal cord or in the gracile or lateral cuneate nuclei of the medulla oblongata (cases 12-14, 17, and 20) (Fig. 2b).
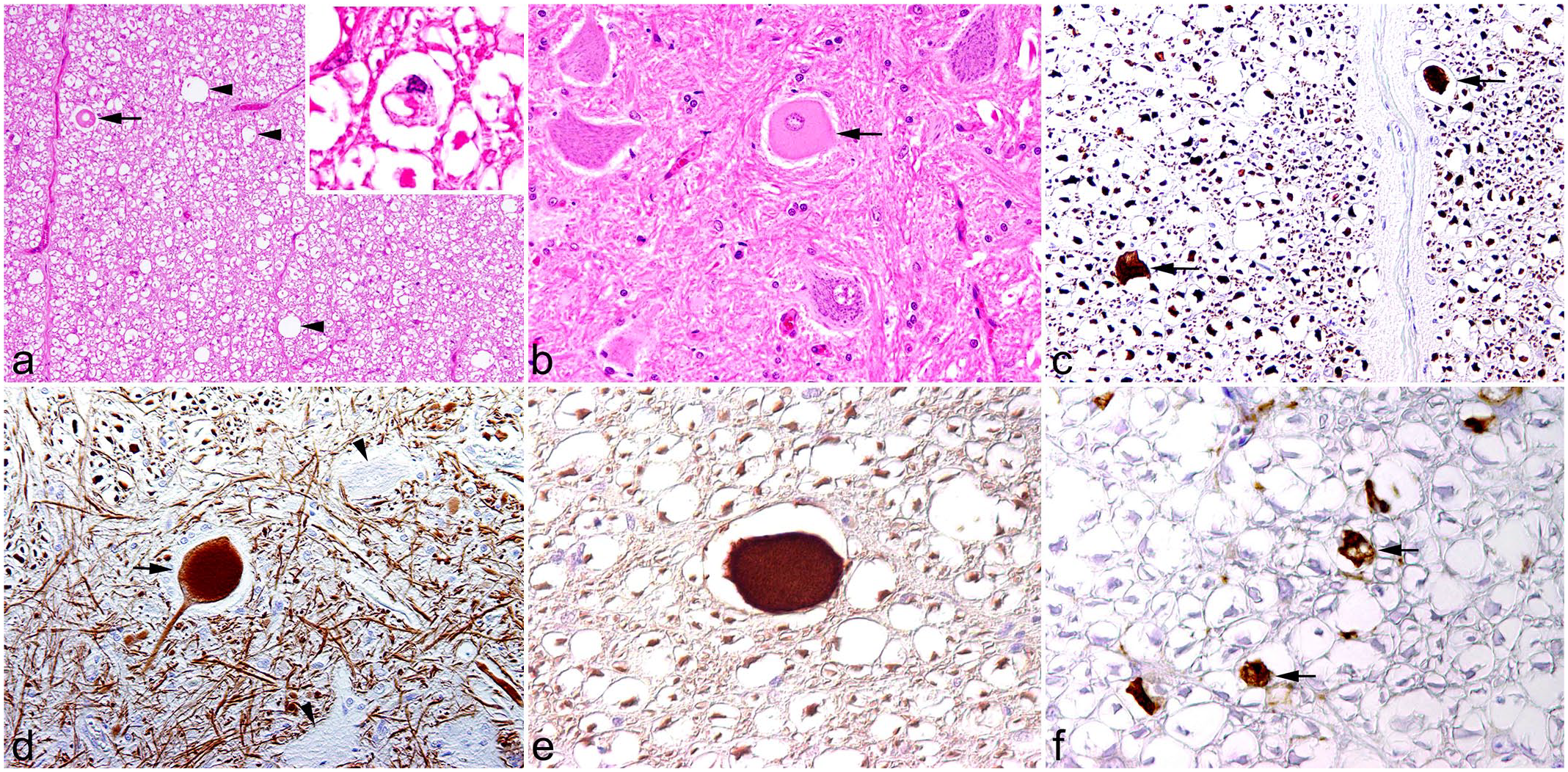
Degenerative changes in the spinal cord of 1 lion. Case 12. (a) Ventral descending funiculi contain dilated myelin sheaths (arrowheads) with occasional spheroids in the dilated myelin sheaths (arrow). Inset: Infiltration of myelinophages is observed. Hematoxylin and eosin (HE). (b) Ventral horn shows a single chromatolytic neuron (arrow) among normal neurons. HE. (c) Spheroids express phosphorylated neurofilament (arrows). Immunohistochemistry (IHC) for phosphorylated neurofilament. (d) In contrast to unaffected neurons (arrowheads), degenerated neurons have an accumulation of phosphorylated neurofilament (arrow). IHC for phosphorylated neurofilament. (e) Accumulation of non-phosphorylated neurofilament is also present in spheroids. IHC for non-phosphorylated neurofilament. (f) Myelinophages are identified in dilated myelin sheaths (arrows). IHC for CD204.
The small intestines of 9 animals in the second group (cases 10-12, 14, and 16-20) had mild to severe, diffuse, lymphohistiocytic, erosive enteritis accompanied by villous atrophy and crypt dilatation. In 7 of these animals (cases 10, 11, 14, 16-18, and 20), amphiphilic to basophilic intranuclear inclusion bodies were found within crypt epithelial cells (Fig. 3a). Seven animals (cases 10-12, 14, and 16-18) also had mild to moderate, multifocal enteritis with infiltration of eosinophils. The small intestine of 1 animal (case 2) was characterized by mild, diffuse, lymphoplasmacytic infiltration of the lamina propria. Furthermore, in 1 animal (case 3), moderate intestinal endoparasitosis with nematodes was observed. The small intestines of 3 animals (cases 1, 5, and 8) did not have histopathologic changes.
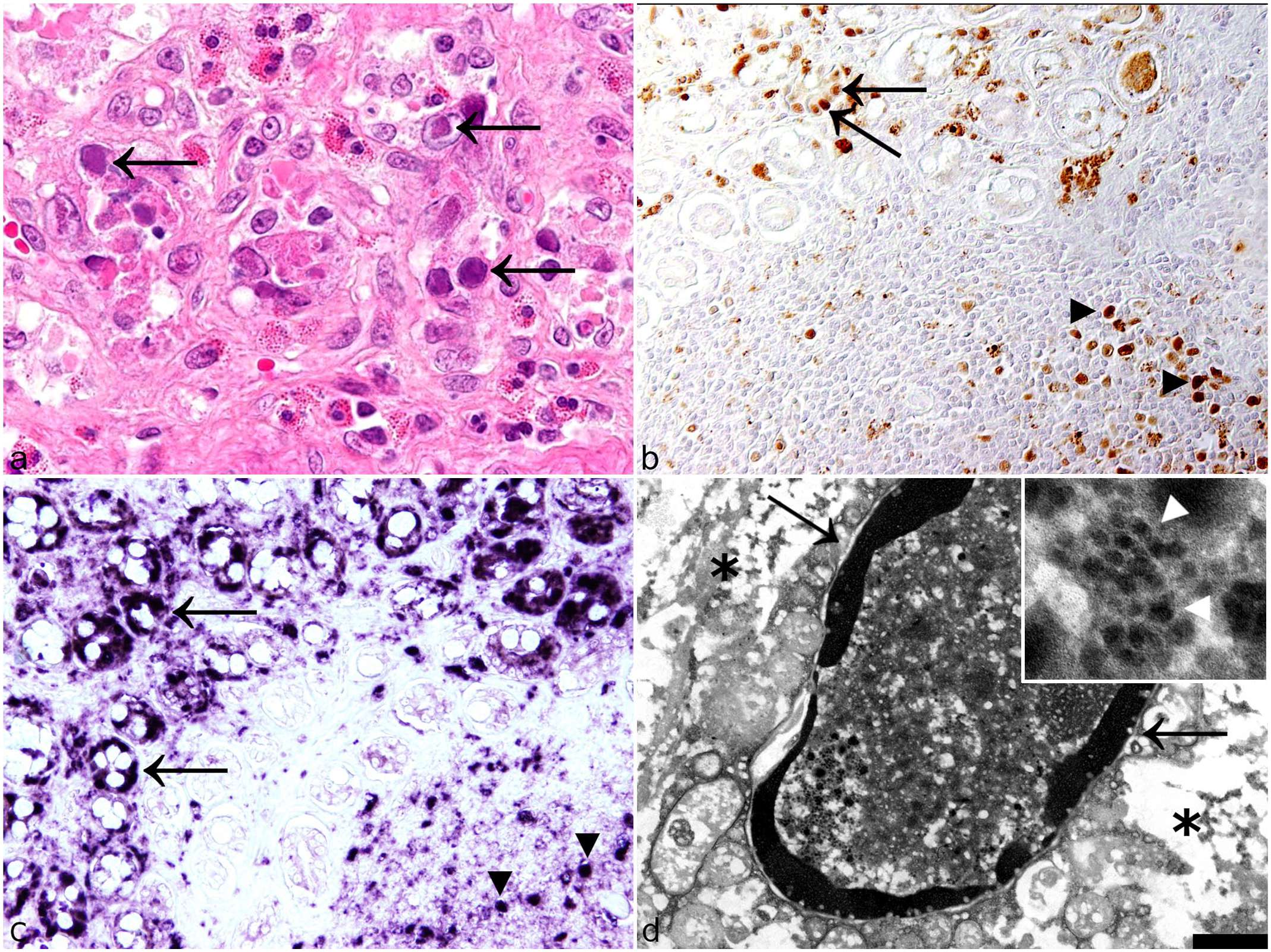
Parvovirus infection in lions, small intestine. (a) Crypt epithelial cells contain large amphophilic to basophilic inclusion bodies (arrows). Case 20. Hematoxylin and eosin. (b) Parvovirus antigen is found within crypt epithelial cells (arrows) and lymphoid cells of Peyer’s patches (arrowheads). Case 11. Immunohistochemistry for parvovirus capsid protein. (c) Parvovirus RNA is also detected in crypt epithelial cells (arrows) and lymphoid cells of Peyer’s patches (arrowheads) Case 11. Chromogenic in situ hybridization. (d) Intranuclear inclusion body within an enterocyte (asterisks) showing prominent accumulation of nuclear chromatin along the nuclear membrane (arrows). Inset: Nuclear inclusion bodies contain 20 to 30 nm diameter viral particles (arrowheads) consistent with members of the Parvoviridae family. Case 11. Transmission electron microscopy, scale bar = 1000 nm.
The main histological lesion of mesenteric lymph nodes was severe, multifocal to coalescing, chronic, granulomatous to pyogranulomatous, necrotizing lymphadenitis in 4 cases (cases 9-11 and 16) (Fig. 4a). Ziehl-Neelsen stain revealed slender intralesional acid-fast bacilli (Fig. 4b) in all 4 cases. In 2 of these animals (cases 9 and 11), inflammatory lesions were accompanied by multifocal mineralization. Minor changes included severe, follicular hyperplasia within the mesenteric lymph node of 2 lions (cases 15 and 16) and moderate, multifocal, follicular, and cortical lymphoid depletion of the mesenteric lymph node of another lion (case 20).
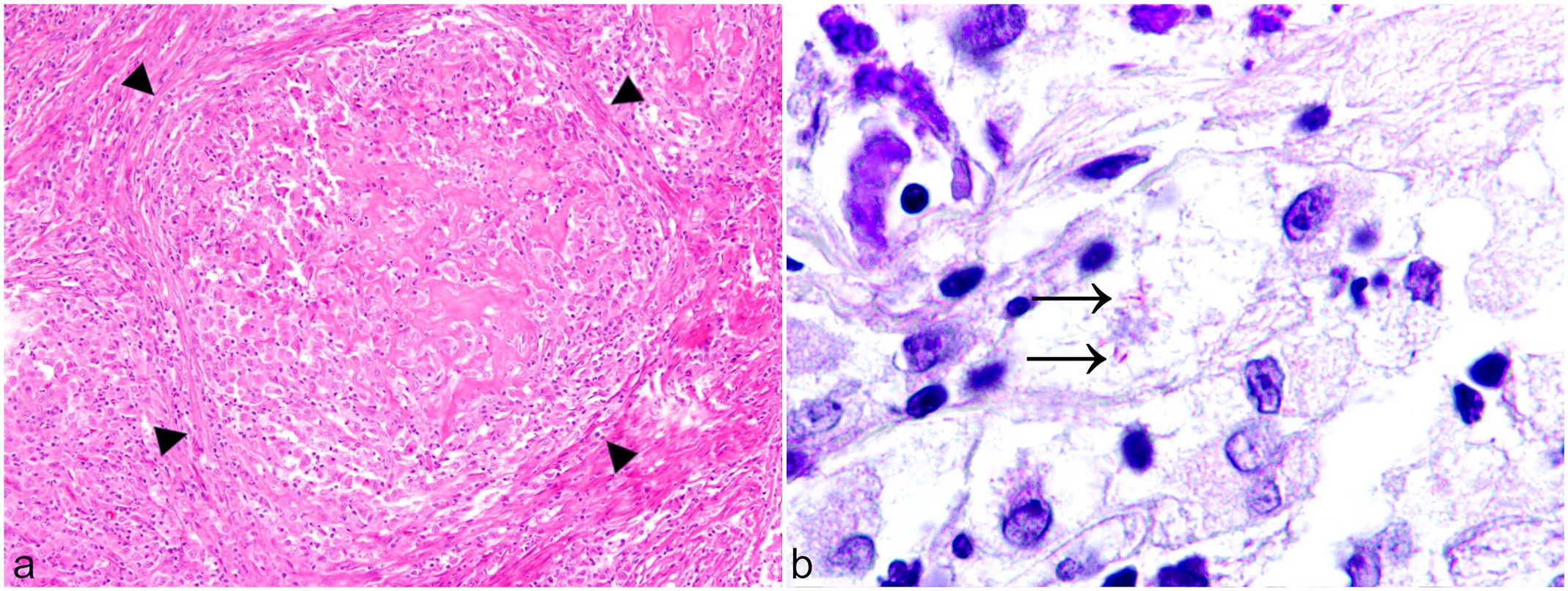
Mycobacterium bovis infection in lions, mesenteric lymph node. (a) Focal granulomatous inflammation (arrowheads), Case 10. Hematoxylin and eosin. (b) Rod-shaped acid-fast bacteria are detected within an ileocecal lymph node (arrows). Case 11. Ziehl-Neelsen stain.
The kidneys of 7 lions (cases 2-5, 7, 10, and 18) had a mild to moderate, focal to multifocal interstitial inflammation composed of lymphocytes, plasma cells, and macrophages. No significant histopathologic changes were found within the kidneys of the remaining animals (cases 1, 8, 9, 11, 12, 15-17, and 19).
The liver was characterized by mild, focal to oligofocal, lymphohistiocytic, and inconsistently plasmacytic, portal to randomly distributed inflammation in 5 animals (cases 4, 9, 11, 13, and 19). In addition, one of these animals had focal, moderate, pyogranulomatous inflammation (case 9) and another lion had mild neutrophilic inflammation with portal hemorrhages (case 4). Hepatic lesions of 2 animals (cases 1 and 17) included an increased number of arteriolar profiles in portal fields, lymphangiectasia, and centrilobular, perivenous fibrosis. Liver tissues of the remaining animals (cases 2, 3, 5, 7, 8, 10, 16, 18, and 20) revealed only minor or no histologic changes.
Inflammatory lesions of the lung were detected in 2 animals (cases 5 and 8) and were characterized by mild, multifocal, lymphoplasmacytic, and partially histiocytic interstitial pneumonia. Within the lung of 1 animal (case 3), moderate, focal, degenerative arteriopathy with segmental fibrosis of the intima and plaque-like calcifications was found. In the other animals (cases 1, 7, 12, and 16-20), only minor or no histologic lesions were observed.
The spleens of 3 animals (cases 5, 7, and 9) had mild to moderate follicular hyperplasia, and 1 animal (case 1) had multifocal follicular depletion. No other histologic lesions were found in the spleens of the remaining animals (cases 4 and 11). Multifocal, lymphoplasmacytic, perivascular, and adnexal dermatitis with intralesional conidia was found in 4 animals (cases 10, 12, 13, and 19).
The adrenal gland of 1 lion (case 19) had severe, acute cortical hemorrhages. Histologic examination of the adrenal gland from the other available lions (cases 9 and 20) revealed no histopathologic findings. Conjunctival tissue of 1 lion (case 4) had moderate, segmental, suppurative inflammation. No histologic lesions were seen in the heart, pancreas, nasal mucosa, thyroid gland, stomach, and large intestine.
Rustrela Virus Detection
All 8 lions in the first group (cases 1-8) tested positive for RusV capsid antigen in the CNS by IHC. Viral antigen was found in the cerebrum, cerebellum, brain stem, and spinal cord. Immunopositive reactions were characterized by dark brown, fine to coarse granular, cytoplasmic labeling in cortical neurons (Fig. 1b); axons; mid-sized elongated cells, most likely representing astrocytes; Purkinje cells; and in neurons of the spinal cord. Some RusV antigen-positive cells were present in areas of brain tissue lacking inflammation. Occasionally, immunopositive cortical neurons were present adjacent to perivascular cell infiltrates. The CNS tissues of the remaining 12 lions (cases 9-20) tested negative for RusV capsid antigen.
To further substantiate these immunohistochemical findings, CISH targeting RusV RNA was performed on CNS samples of 3 lions that tested positive and 3 that tested negative for RusV capsid antigen by IHC (cases 2, 5, and 6 and 13, 15, and 19, respectively). Positive results were obtained only for the RusV-positive lion 5, but not for the RusV-positive lions 2 and 6 or for the 3 RusV-negative animals (cases 13, 15, and 19). Examination of the CNS tissues of lion 5 yielded positive reactions in the cerebrum. RusV RNA was detected exclusively in cerebral cortical neurons in the form of multifocal, cytoplasmic, granular red dots (Fig. 1c). The distribution of RusV RNA was similar to the immunohistochemical expression of RusV capsid antigen and also exhibited an inconsistent co-localization with inflammatory lesions.
RusV-specific RT-qPCR was performed for all 20 lions included in this study. FFPE CNS tissues from lions 1 to 8 were positive with the cycle of quantification (Cq) values ranging from 25 to 39. In contrast, RusV RNA was not detected in the brain sections of the remaining 12 lions (cases 9 to 20). Partial RusV p150-encoding sequences of 409 base pairs were generated for cases 1 to 6 and 8, whereas sequencing attempts failed for case 7, presumably due to the low amount of detectable RNA in the only available FFPE samples from the spinal cord. The 7 RusV sequences were divided into 2 sequence types that shared 96.1% nt identity. Phylogenetically, both types belonged to the same clade as the RusV sequences from the previously published encephalitic lions from Western Germany during the 1980s (Fig. 5). 21
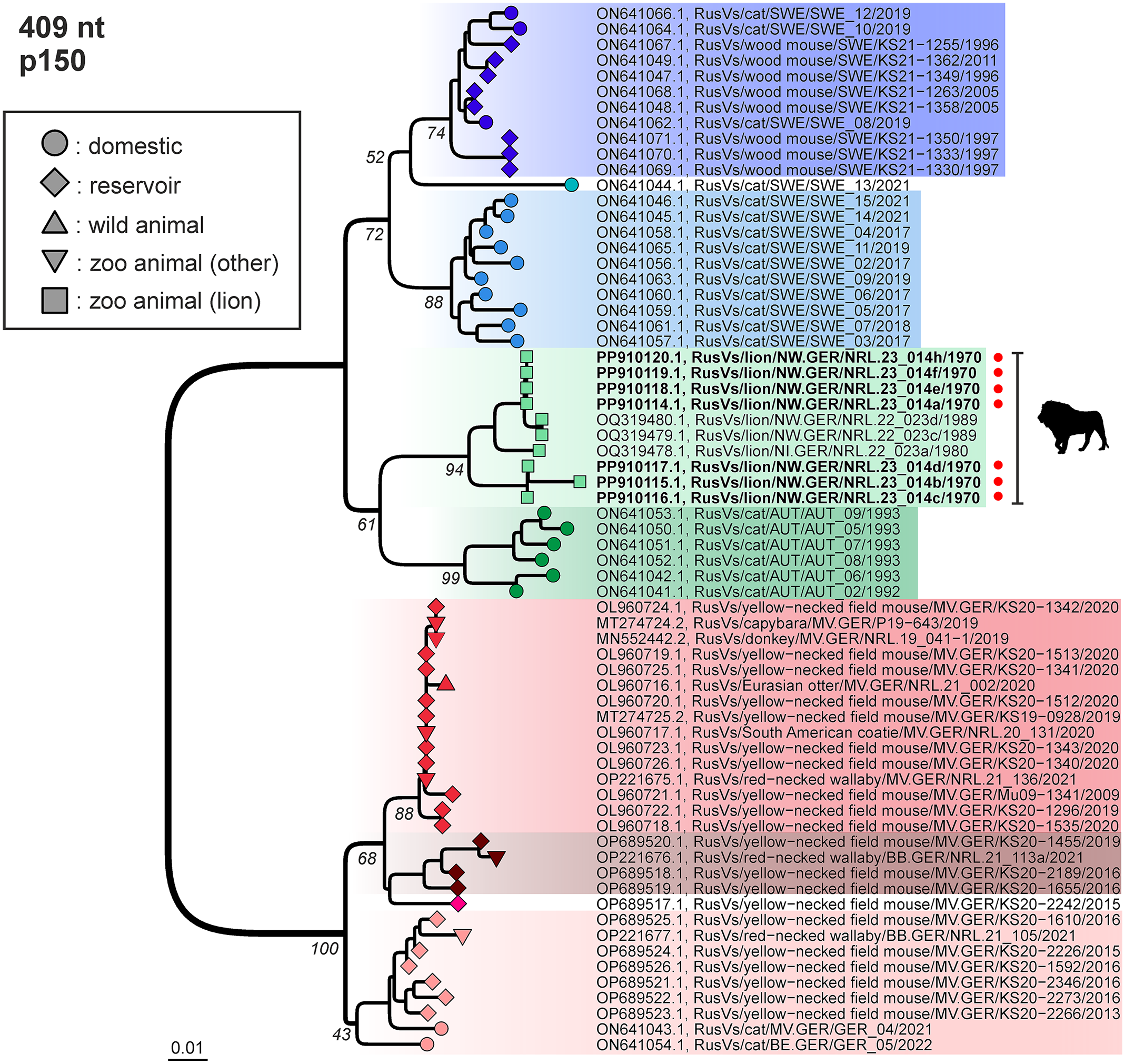
Phylogenetic analysis of rustrela virus (RusV) infections in Europe. Neighbor-joining phylogenetic tree of partial RusV p150 sequences (409 nt; MEGA version 11.0.13; TN93+G+I; 1,000 bootstrap replicates). Only bootstrap values at major branches are shown in the phylogenetic tree. RusV sequence names are shown in the format “host/ISO 1366 code of location (federal state.country)/animal ID/year.” Sequences generated during this study are depicted in bold and marked with a red dot. The respective hosts are indicated by circles (domestic cats), squares (lions), a triangle (wild animals), upside-down triangles (zoo animals), or diamonds (Apodemus spp.). AUT, Austria; GER, Germany; SWE, Sweden; BB, Brandenburg; BE, Berlin; MV, Mecklenburg-Western Pomerania; NI, Lower Saxony; NW, North Rhine-Westphalia.
Parvovirus Detection
Immunohistochemical analysis for parvovirus antigen revealed immunopositive reactions in 8 of 20 lions (cases 10-12, 14, 16-18, and 20), which all belonged to the second group. Immunopositive reactions were observed in the small intestines of all 8 lions and in lymph nodes and spleen. In the small intestine, variable numbers of immunopositive enterocytes, predominantly crypt epithelial cells (Fig. 3b), were observed. The intranuclear inclusion bodies were parvovirus antigen positive. Immunoreactive cells were limited to intestinal segments with mucosal lesions. In 3 lions (cases 10, 11, and 17), cells resembling macrophages located in the lamina propria adjacent to crypts were also positive for parvovirus antigen. The small intestine of 1 lion (case 11) contained Peyer’s patches in which moderate numbers of immunopositive cells, most likely lymphocytes and macrophages, were visible (Fig. 3b). The small intestines without histopathologic lesions of 3 control animals (cases 21-23) tested negative for parvovirus antigen.
Lymph nodes of 4 lions (cases 10, 11, 16, and 20) had mild to moderate numbers of cytoplasmic and intranuclear immunopositive cells, mainly within subcapsular sinuses. Based on their morphology, these were most likely macrophages. In addition, fewer immunopositive antigen-presenting cells and lymphoid cells were found in follicles.
In the spleen of 1 animal (case 11), parvovirus antigen was detected in the cytoplasm and nucleus of a few cells within lymphoid follicles, suggestive of antigen-presenting cells. No immunolabeling for parvovirus antigen was found in any other available tissue from the lions that were positive for parvovirus by IHC in the small intestines, lymph nodes, and spleen. Immunohistochemical examination of the remaining 12 animals yielded negative results (cases 1-9, 13, 15, and 19), including the CNS of the 8 RusV-positive lions in the first group (cases 1-8).
Parvovirus infection was confirmed by the demonstration of viral RNA in 2 parvovirus antigen-positive lions (cases 10 and 11). Viral RNA was detected in the small intestine (Fig. 3c), lymph nodes, and spleen, as already shown by IHC. Both the number and distribution pattern of positive cells were consistent with immunohistochemical results for the expression of parvovirus antigen. In addition, as already demonstrated by IHC, no positive signal was observed in the CNS of any of the 11 animals displaying encephalomyelopathy (cases 10-20).
Ultrastructurally, intranuclear inclusions of amorphous, electron-dense material were detected in several cells of the small intestine of 1 lion (case 11). Based on the morphology, these cells were identified as enterocytes. The nucleolus and the nuclear chromatin of these cells appeared marginalized. Randomly distributed within the inclusions, accumulations of virus particles were observed (Fig. 3d). These particles were polygonal, non-enveloped, and 20 to 30 nm in diameter.
Metagenomic analyses of the sequencing data sets generated from case 11 and case 14 unambiguously confirmed the presence of parvoviral reads (550 and 45 reads, respectively) and in addition revealed sequences presumably originating from members of the family Circoviridae (27 reads in case 11, 3 reads in case 14) and Totiviridae (1269 reads in case 11, 270 reads in case 14), respectively. The sequencing results are summarized in Supplemental Tables S3 and S4. The parvoviral sequences had the highest identities with various sequences belonging to the species Protoparvovirus carnivoran 1. The quality of the obtained sequence reads did not allow the assembly of reliable longer sequences due to the fragmentation of the nucleic acids and because the base modifications that were induced by formalin fixation led to a number of ambiguous positions in the sequences. Therefore, further detailed sequence analyses were not performed.
Testing for Other Potential Viral Etiologies
The investigation for the presence of CDV antigen was negative in the CNS of all 20 animals as well as in the lungs (n = 11), small intestines (n = 14), lymph nodes (n = 7), and spleens (n = 3), whenever available.
Investigation for FeLV antigen was negative in the CNS tissue of all 20 animals as well as in the lymph nodes (n = 7) and spleens (n = 3), whenever available.
Immunohistochemistry for Double-Stranded RNA
The diagnostic value of dsRNA immunolabeling using 3 different monoclonal antibodies was examined in tissues of all lions including the CNS tissues (n = 20), small intestines (n = 17), lymph nodes (n = 7), and spleens (n = 6). Overall, 19 of 20 animals tested positive with at least one of the 3 antibodies in the CNS tissues, although viral antigen was only detected in the CNS of the 8 RusV-infected lions (cases 1-8). In addition, CNS samples of all 8 parvovirus-infected lions (cases 10-12, 14, 16-18, and 20), of 2 animals with degenerative changes (cases 15 and 19), and of an M. bovis-infected animal (case 9) showed a positive signal, while the nervous tissue of 1 lion with degenerative changes (case 13) lacked a dsRNA signal.
In 15 of 17 lions, a positive signal was observed in the small intestine, including all 8 parvovirus-infected lions (cases 10-12, 14, 16-18, and 20), 4 RusV-positive lions without detectable parvovirus antigen in the intestines (cases 1, 2, 5, and 8), and 3 control animals from other zoos without intestinal lesions (cases 21-23). The intestines of 2 lions, 1 RusV-positive animal (case 3), and 1 lion with degenerative encephalomyelopathy (case 19) tested negative with the dsRNA antibodies.
The dsRNA labeling was also demonstrated in all investigated lymph nodes, including those from 4 parvovirus-infected animals immunopositive for parvovirus antigen (cases 10, 11, 16, and 20) as well as from 1 RusV-infected animal lacking RusV antigen in the lymph node (case 5), 1 M. bovis-infected lion (case 9), and 1 lion with encephalomyelopathy (case 15).
In splenic tissues, positive signals were detected in 5 of 6 lions, including a parvovirus-positive animal with splenic parvovirus antigen (case 11), 3 RusV-positive animals (cases 1, 4, and 7), and 1 M. bovis-infected animal (case 9). All examined samples tested positive with either monoclonal antibody J2 or K1 or both, while antibody 9D5 did not provide a signal for any of the samples. However, an immunopositive signal for 9D5 was observed in the positive control.
RNase III pretreatment to specifically degrade dsRNA was performed for the CNS of an RusV-positive lion with dsRNA signal (case 8) and an RusV-negative lion without dsRNA labeling (case 13). The RNase III pretreatment did not alter the signal for either of the animals.
Immunohistochemical Investigation of Degenerative Changes of Central Nervous System
Degenerative CNS lesions were observed in 11 RusV-negative animals (cases 10-20). Spheroids in the white matter (Fig. 2c) and degenerate and necrotic neurons had accumulated p-NF (Fig. 2d) and synaptophysin. Furthermore, spheroids occasionally had an accumulation of n-NF (Fig. 2e). Immunohistochemistry for CD204 antigen confirmed the infiltration of myelinophages (Fig. 2f). The anti-myelin basic protein antibody showed no reactivity in normal and altered lion CNS tissues. In contrast, the antibody generated an immunopositive signal in canine CNS tissue, which was used as a positive control.
Detection of Mycobacterium bovis DNA
Lymph node samples from 4 lions (cases 9-11 and 16) with visible tuberculous lesions and detection of acid-fast rods tested positive for the presence of the IS1081 DNA fragment, which is specific for members of the M. tuberculosis complex. By proof of the deletion of the RD4 region, the species within the M. tuberculosis complex was identified as M. bovis in all 4 samples. Further characterization by identification of the spoligotype of the M. bovis strains was not successful (data not shown). This was probably due to the low amount of target DNA, as shown by the cycle threshold values between 32.4 and 35.4 within the samples (Supplemental Table S2).
Discussion
The lions included in this study could be categorized into 2 distinct groups according to the occurrence of the disease and the cause. Animals from December 1970 had meningoencephalomyelitis due to an RusV infection. In contrast, 11 of the 12 lions from April 1971 had degenerative encephalomyelopathy, and all 12 animals were negative for RusV. The non-suppurative, perivascular meningoencephalomyelitis observed in the CNS of the 8 lions in the first group was highly indicative of a viral etiology. 80 Reported neurologic signs and histopathologic lesions of the brain and spinal cord were consistent with reports of recently confirmed RusV infections in zoo and wildlife animals, including domestic cats and lions.10,21,58,69,73,99,103 Suspected RusV infections in the present study were confirmed by RT-qPCR, IHC, and CISH. RusV RNA and capsid antigen were limited to the CNS. Other organs, only examined for viral antigen but not for viral RNA, were negative. As already described in previous studies, the present findings support the hypothesis of a strong neurotropism in RusV infections in encephalitic non-reservoir hosts.10,21,58,73,99 In the present study, RusV RNA and viral antigen were observed in proximity to inflammatory lesions and also in areas without histologic lesions. Therefore, we postulate that the vasculitis may have been caused by either a type III hypersensitivity reaction or by a virus-induced immune response.21,106 Using the newly established RusV-specific CISH assay, we were able to detect RusV RNA in only one of the 3 examined IHC- and RT-qPCR-positive lions, suggesting a lower sensitivity of the assay. 74 Another important factor to consider is formalin fixation and long-term storage conditions of the tissue samples, which may have led to more pronounced degradation of nucleic acids and consequently to false negative ISH results.14,85 As detailed clinical and anamnestic data, particularly concerning additional deaths at the same time, animal husbandry, feeding, and breeding, are lacking, further interpretations remain speculative. However, from October 1971 to April 1973, an additional 25 lions and 3 tigers (Panthera tigris) from the same zoological collection were submitted for necropsy to the State Veterinary Department in Münster, Germany, and were diagnosed with severe meningoencephalomyelitis, which appears to be suspicious for an RusV infection. At that time, however, the RusV was not yet known, so the cause of the inflammation could not be identified. 28 An additional study from 1990 on 3 lions with non-suppurative meningoencephalitis from the same zoo failed to detect the etiologic agent but assumed a viral infection. 95 A retrospective study from 2023 found RusV to be the cause of encephalitis in 2 lions submitted in 1989 by the same zoological institution. Phylogenetic analysis revealed that the RusV sequences detected in that study belonged to the same clade as those detected in lions in the 1980s. 21 These results confirm that RusV was already present in parts of Germany for more than 5 decades. The same RusV variant was also detected in a cat from Lower Saxony in 2017, suggesting that this variant is still circulating in areas of Western Germany. 58 Other RusV variants have been detected in contemporary or retrospectively analyzed samples from encephalitic domestic cats, zoo animals, and wild animals, as well as non-encephalitic yellow-necked field mice and wood mice in northeast Germany (Mecklenburg-Western Pomerania, Brandenburg, and Berlin) from 2009 onwards,10,58,73,69,99 in Sweden from the 1970s onwards, and in Austria predominantly in the early 1990s.58,90,103 These findings suggest that RusV was already widely distributed in Europe for a very long time, and it may be conceivable to assume that additional variants will be detected in further parts of Europe. Very recently, a more distant RusV variant was also detected in a mountain lion in the United States. 29 Although little is known about RusV transmission, as no viral shedding has been detected to date, the presence of RusV RNA in yellow-necked field mice and wood mice brains without histopathological lesions strongly supports the hypothesis that these mice represent the virus reservoir.10,58,73
A parvovirus infection was suspected in 8 lions in hematoxylin and eosin-stained sections of the intestinal tract and was subsequently confirmed by IHC, CISH, and TEM. 93 All of these also had signs of degenerative encephalomyelopathy, and according to the results of metagenomic analyses, the parvovirus present in the samples was most closely related to the species Protoparvovirus carnivoran 1. However, the poor quality of the RNA extracted from the archived FFPE material did not allow for more detailed analyses and a reliable determination of the viral species. The lesions observed are consistent with descriptions of parvovirosis caused by FPV in cats in the 1970s. 4 The route of infection and possible transmission between lions are uncertain, with domestic cats being the most likely carriers. Spread by personnel in contact with infected domestic cats is also conceivable. Further interpretations of this disease entity are limited by the inability to determine the exact species of detected parvovirus. No evidence was found for a connection between the parvovirus infection and the degenerative encephalomyelopathy.
The immunolabeling for dsRNA in the animals included in this study revealed positive signals in almost all investigated samples. Although dsRNA is expected exclusively in virus-infected organs with ongoing viral replication, positive immunolabeling was also detected in animals without evidence of virus infection, suggesting that the positive signals might result from endogenous dsRNA, a so-far undetected viral infection, or unspecific labeling, as suggested by positive signals even after dsRNA digestion by RNase III pretreatment.37,54 For this reason, the results should be interpreted with caution, as the dsRNA-positive reactions are most likely nonspecific signals. Potential effects based on the long-term FFPE storage, as well as the lack of clinical data that did not allow for any conclusions regarding the time point of infection represent other factors with an unknown impact on the results. 85 As recently published, the time point of infection appears to have a considerable impact on the results of dsRNA detection. It was demonstrated that particularly in the early stages of infection with a peak of viral replication, a high amount of dsRNA can be detected. 13 Due to the limited clinical data, it was not possible to establish any potential connection between clinical data and the dsRNA results, which could potentially have contributed to the interpretation of the present findings. Since endogenous retroviruses can also lead to the formation of dsRNA, their presence in the animals of this study cannot be ruled out and should be taken into consideration when interpreting the results.
Four animals had lesions in the mesenteric lymph nodes indicative of tuberculosis. M. bovis was subsequently confirmed by PCR in those 4 animals. Two more lions were considered M. bovis-positive due to the archived documentation, even though histologic evidence was lacking and more cases might have been missed due to incomplete records and available samples. Based on the consistent finding of granulomatous lymphadenitis in mesenteric lymph nodes and a lack of similar changes in the respiratory tract and associated lymph nodes, an alimentary infection is suspected to be the portal of entry. The origin and type of animal food, as well as the feeding practice of the zoo in the early 1970s are unknown, and therefore, the route of infection remains unclear. Other animals in the zoo, including native wildlife, might play a role in spreading M. bovis. 79 Clinical manifestation of tuberculosis in lions is associated with emaciation, respiratory signs, swollen lymph nodes, ataxia, hypermetria, and poorly healing wounds.19,40,41,92 Nevertheless, subclinical infections are thought to represent the majority of cases in free-ranging lions. 41 Concerning the animals in this study, the clinical relevance of tuberculosis is difficult to evaluate, especially as the available medical records of clinical signs are incomplete and nonspecific, including hind limb weakness. Five of the M. bovis-positive lions additionally had parvovirosis and degenerative encephalomyelopathy. As tuberculosis has been associated with ataxia before, a relationship to the previously mentioned hind limb weakness cannot be ruled out. Tuberculosis might have contributed to a generally poor health status and wasting of the animals. 19 However, tuberculosis was diagnosed exclusively in mesenteric lymph nodes, and no suggestive lesions were found in the CNS. Furthermore, 12 lions demonstrated similar clinical signs, but M. bovis was found in only 6 animals, so tuberculosis could also be a co-morbidity with rather low clinical impact. An etiologic connection to the encephalomyelopathy is not proven. The sixth lion did not have any other morphological changes apart from tuberculosis. Nonetheless, the quality and availability of the FFPE material must be critically considered in this case.
Degenerative encephalomyelopathy was found in 11 lions and was characterized by alterations in ascending and descending funiculi of the white matter of the caudal brain stem and the spinal cord, which are regarded as the cause of the locomotor disorder. Immunohistologic characterization of the spheroids in the white matter showed an accumulation of n-NF, p-NF, and synaptophysin. Under physiological conditions, n-NFs are synthesized in the neuronal cell body. Within the axon, n-NFs undergo a complex pattern of phosphorylation, resulting in p-NFs. 72 N-NFs are regarded as an early marker for axonal damage and may serve as morphologic evidence of neurodegenerative diseases.47,81 The degenerative and necrotic changes of neurons may result from axonal damage in the form of a dying back phenomenon. However, a separate mechanism causing direct neuronal damage cannot be excluded completely. The cause of the degenerative encephalomyelopathy in the investigated lions remains undetermined. The observed spectrum of neurodegenerative changes shares findings observed in a 1-year-old lion with confirmed vitamin A deficiency. 55 Vitamin A deficiency is associated with a variety of morphologic changes in various animal species. CNS manifestations are regarded to be associated with impaired bone remodeling and impaired absorption of cerebrospinal fluid.26,59,88 In young captive lions, suspected or confirmed vitamin A deficiency causes a neurologic disease due to hyperostosis of skull bones, notably the tentorium cerebelli osseum, with subsequent herniation of the cerebellar vermis.3,5,17,35,70,96 One lion from the same zoological institution as the above described cases had suffered from a confirmed vitamin A deficiency with hyperostosis and cerebellar herniation. 96 Histologically, Wallerian degeneration may occur in the cranial spinal cord and is probably associated with the direct pressure on the CNS tissues.17,70 In contrast, the pathogenesis of the diffuse degeneration of the white matter that has been described in 1 captive lion, and that shares similarities with the investigated 11 lions, is assumed to be caused by increased pressure of the cerebrospinal fluid. 55 However, leptomeningeal adhesions to the dura, thickening by fibrosis and osseous metaplasia, hydromyelia, and reactive astrocytosis, as described for that lion, were not observed in the investigated cases. 55 An additional differential diagnosis is leukoencephalomyelopathy of mature captive cheetahs and other large felids. 16 This neurologic disorder is characterized by bilateral degenerative lesions in the cerebral white matter and, to a lesser degree, in the white matter of the brain stem and spinal cord associated with astrocytosis and finally cavitation. Due to the lack of cerebral changes, this disorder of suspected neurotoxic origin can be excluded in the presented cases. In young cheetahs, an idiopathic acute onset of myelinopathy has been described with myelin sheath dilatation, occasional spheroids, astrogliosis evident in lateral and ventral funiculi and occasionally occurring in the dorsal columns, and scattered perivascular lymphohistiocytic cuffs. 100 The distribution of lesions and the inflammatory infiltrate in the cheetahs do not match the findings in the lions of the present study. In tigers, lions, and a leopard, cases of an axonal degeneration have recently been published. 61 Affected axons were randomly distributed throughout the white matter tracts without apparent predilection for a particular spinal segment or associated brain lesions. The distribution pattern in the reported large felids differs from the investigated lions, indicating a different disease entity.
Two of the lions showed histologic lesions in the liver suggestive of a portosystemic shunt. As these shunts can lead to neurologic signs, the clinical relevance of these findings has to be taken into consideration. 53 However, the lesions present in the CNS of the lions in this study lacked changes expected in hepatic encephalopathy, eg, spongy vacuolation of myelin and Alzheimer type II glial cells. 59
This retrospective analysis of a past increased incidence of morbidities illustrates how important archived FFPE material can be to resolve recent disease outbreaks. More importantly, FFPE is often the only material available and has been successfully used for virus detection.14,32,89 The size of the tissue samples, pre-fixation time, and time of fixation before embedding in paraffin, as well as time and conditions of storage in paraffin potentially interfere with the quality of FFPE material for further sophisticated analyses.9,15,20,44,85,98 FFPE samples from 1970 and 1971 used in this study were appropriate for various histochemical stains as well as for IHC, ISH, and PCR. According to the documents, the majority of lions were euthanized shortly before necropsy and none of the investigated tissues had changes of autolysis. Therefore, a short time span between the death of an animal, its pathologic examination, and subsequent further processing of tissue samples can be assumed. However, information on the duration of formalin fixation, the concentration, and pH value of the used formalin is lacking. Documenting further details on the sample processing could prove valuable for future research, improving the precision of retrospective studies that depend on FFPE material. After embedding in paraffin, the material was most likely stored at room temperature with no further control regarding other external factors like air humidity. Two limitations regarding the usability of the material were encountered in this study. First, further characterization of the strain of M. bovis was not possible, and second, the present parvovirus could not be determined precisely. Whether this restriction on the investigation was connected to the time or condition of storage or was linked to other factors regarding fixation or handling of samples remains unknown.
The re-evaluation of this past outbreak by histochemical stains, IHC, ISH, PCR, and TEM revealed 4 different patterns of the aforementioned diseases, which could explain the poor health status of the animals. While the RusV-positive lions had no other lesions, infections with parvovirus and M. bovis as well as the degenerative encephalomyelopathy appeared partly as co-morbidities. Overall, obtained findings show that FFPE material is and will be an extremely useful source for investigations of outbreaks using various diagnostic techniques.
Supplemental Material
sj-pdf-1-vet-10.1177_03009858251335280 – Supplemental material for Retrospective analysis of clustered neuroinflammatory and neurodegenerative diseases in captive lions in the early 1970s
Supplemental material, sj-pdf-1-vet-10.1177_03009858251335280 for Retrospective analysis of clustered neuroinflammatory and neurodegenerative diseases in captive lions in the early 1970s by Madeleine de le Roi, Inga Nägler, Dennis Rubbenstroth, Martin Beer, Dirk Höper, Stefanie A. Barth, Adnan Fayyad, Christina Puff, Wolfgang Baumgärtner and Peter Wohlsein in Veterinary Pathology
Footnotes
Acknowledgements
We wish to thank Julia Baskas, Petra Grünig, Jana-Svea Harre, Claudia Herrmann, Kerstin Rohn, Kerstin Schöne, Caroline Schütz, Kathrin Steffen, Danuta Waschke, Lukas Wessler, and Patrick Zitzow for their excellent technical support. We are also grateful to Andrea Aebischer for providing the RusV-specific monoclonal antibody 2H11B1 and to Florian Pfaff for support with visualizing the phylogenetic analysis. We would also like to thank Frances Sherwood-Brock for proofreading the manuscript to ensure correct English.
Author Contributions
The manuscript was written by MD, IN, WB, and PW with contributions from the other authors. MD and IN performed histologic and immunohistologic evaluations. DR, MB, DH, SB, and AF performed molecular analysis. CP contributed to the experimental design. WB designed the study.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was in part supported by the Deutsche Forschungsgemeinschaft (DFG; German Research Foundation; -398066876/GRK 2485/1-VIPER-GRK) and by the European Union Horizon 2020 grant agreement no. 874735 (“Versatile Emerging Infectious Disease Observatory”; doi: 10.3030/874735). AF was supported through the An-Najah National University (![]() ) sabbatical leave program.
) sabbatical leave program.
Supplemental Material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.