
Abstract
Epithelial cell clusters found ectopically in lymph nodes are generally considered metastatic lesions from malignant tumors; however, benign epithelial inclusions (BEIs) are a known exception, and distinguishing them from malignancy is often challenging in human medicine. We report the presence of BEIs in a cynomolgus monkey with severe renal damage but no evidence of neoplastic lesions. The inclusions were observed in the subcapsular sinuses of the lymph nodes and were positive for cytokeratin and aquaporin-1 by immunohistochemistry. Similar epithelial clusters were also found within the lymphatic vessels of the renal hilum. Furthermore, migration of epithelial cells into the lymphatic vessels within the renal parenchyma was observed, suggesting that the BEIs in this case originated from renal tubular epithelium. To our knowledge, this is the first report of BEIs of renal tubular origin in an animal, providing valuable insights into their histological and ultrastructural features, as well as their underlying pathogenesis.
Keywords
Ectopic cells in lymph nodes are generally considered metastatic tumor cells from malignant tumors; however, benign inclusions are a rare exception. These inclusions, defined as foci of non-neoplastic ectopic tissues, are classified as epithelial, nevomelanocytic, or decidual. 12 Benign epithelial inclusions (BEIs) are clusters of benign, well-differentiated epithelial cells in lymph nodes.3,9,11–13 In human pathology, BEIs are reported in various anatomical locations. Differentiating BEIs from metastatic lesions is crucial in preventing overdiagnosis of carcinomas.1,3,9,12,13,16,17,19
BEIs are often observed in sentinel axillary lymph nodes following diagnosis of breast tumors or other suspicious lesions.1,3,9,11,12 Other epithelial inclusions reported in humans include salivary and thyroid tissue in cervical lymph nodes, mesothelial cell inclusions in mediastinal lymph nodes, glandular epithelial inclusions in peritoneal lymph nodes, Müllerian duct inclusions in pelvic lymph nodes, Tamm-Horsfall protein (THP) and tubular epithelial inclusions in renal lymph nodes, and glandular and squamous epithelial inclusions in peripancreatic lymph nodes.1,3,9,11–13,16,17,19 The origin of these inclusions is attributed to various theories, including embryogenesis, iatrogenic implantation, and benign metastasis of detached cells, with pathogenesis and differential diagnoses varying by anatomical site.5,9
In veterinary pathology, BEIs have only been reported in 3 dogs, 8 a cow, 4 and a dolphin, 2 all involving mesothelial cell inclusions in mediastinal lymph nodes, with or without associated lesions in adjacent tissues. We encountered a 4-year-old male cynomolgus monkey (Macaca fascicularis) with BEIs in the renal hilar lymph node, likely originating from renal proximal tubules. To our knowledge, this is the first report of BEIs originating from renal tubules in a lymph node of animals. This report highlights the histological characteristics of these BEIs and discusses possible pathogenesis.
The monkey, imported from Cambodia (purchased from Hamri, Japan) at 2 years and 6 months of age, was housed in a metal cage (680 × 608 × 770 mm) in a conventional air-conditioned room maintained at 23°C to 29°C with 35% to 75% relative humidity and a 12-hour light/dark cycle. It was fed 100 g of commercially available food (CMK-2; CLEA Japan, Inc, Tokyo, Japan) daily with free access to drinking water. Care was provided in accordance with the guidelines for the care and use of laboratory animals established by the Japanese Association for Laboratory Animal Science and our institution. The animal was kept as a standby for toxicity studies and received a single dose of a test substance. After 15 months of untreated care, it died suddenly. Necropsy revealed enlarged kidneys without detectable mass lesions. Other findings included subcutaneous edema; effusion in the thoracic cavity, abdominal cavity, scrotum, and pericardial sac; enlargement of the kidneys, liver, heart, and spleen; and brown discoloration of the lungs.
Samples from all organs, including the kidneys and renal hilar lymph nodes, were fixed in 10% phosphate-buffered formalin, embedded in paraffin, and cut at 4 µm. Samples from the liver, gallbladder, spleen, thymus, pancreas, lung, heart, thyroid/parathyroid, adrenal gland, testis, epididymis, submandibular salivary gland, brain, mandibular lymph node, mesenteric lymph node, skin, urinary bladder, tongue, esophagus, trachea, aorta, femoral muscle, sciatic nerve, stomach, duodenum, jejunum, ileum, cecum, colon, and rectum were also collected and processed using the same procedure. Sections were stained with hematoxylin and eosin, periodic acid-Schiff, and Watanabe’s silver stain. Immunohistochemical examinations were performed on sections of the renal hilar lymph nodes using the primary antibodies listed in Table 1. Immunolabeling was detected using a peroxidase-labeled polymer system with 3,3’-diaminobenzidine as the chromogen. As renal tissue was present on the same sections as the renal hilar lymph nodes, renal tubular epithelial cells showing the expected immunoreactivity were used as internal positive controls for wide-spectrum cytokeratin, aquaporin-1, calbindin, THP, and CD10. Renal interstitial cells served as internal positive controls for vimentin, and renal lymphatic vessels served as internal positive controls for vascular endothelial growth factor receptor 3 (VEGFR-3). Lymphoid tissue within the lymph node served as an internal positive control for ionized calcium-binding adapter molecule 1 (IBA-1) and Ki-67. Negative controls were prepared by omitting the primary antibodies.
Primary antibodies used for immunohistochemical examination.
For electron microscopy, renal hilar nodes were post-fixed in 1% osmium tetroxide, processed, and embedded in resin. Ultrathin sections were stained with uranyl acetate and lead citrate and examined with a transmission electron microscope (H7600; Hitachi, Ltd, Tokyo, Japan).
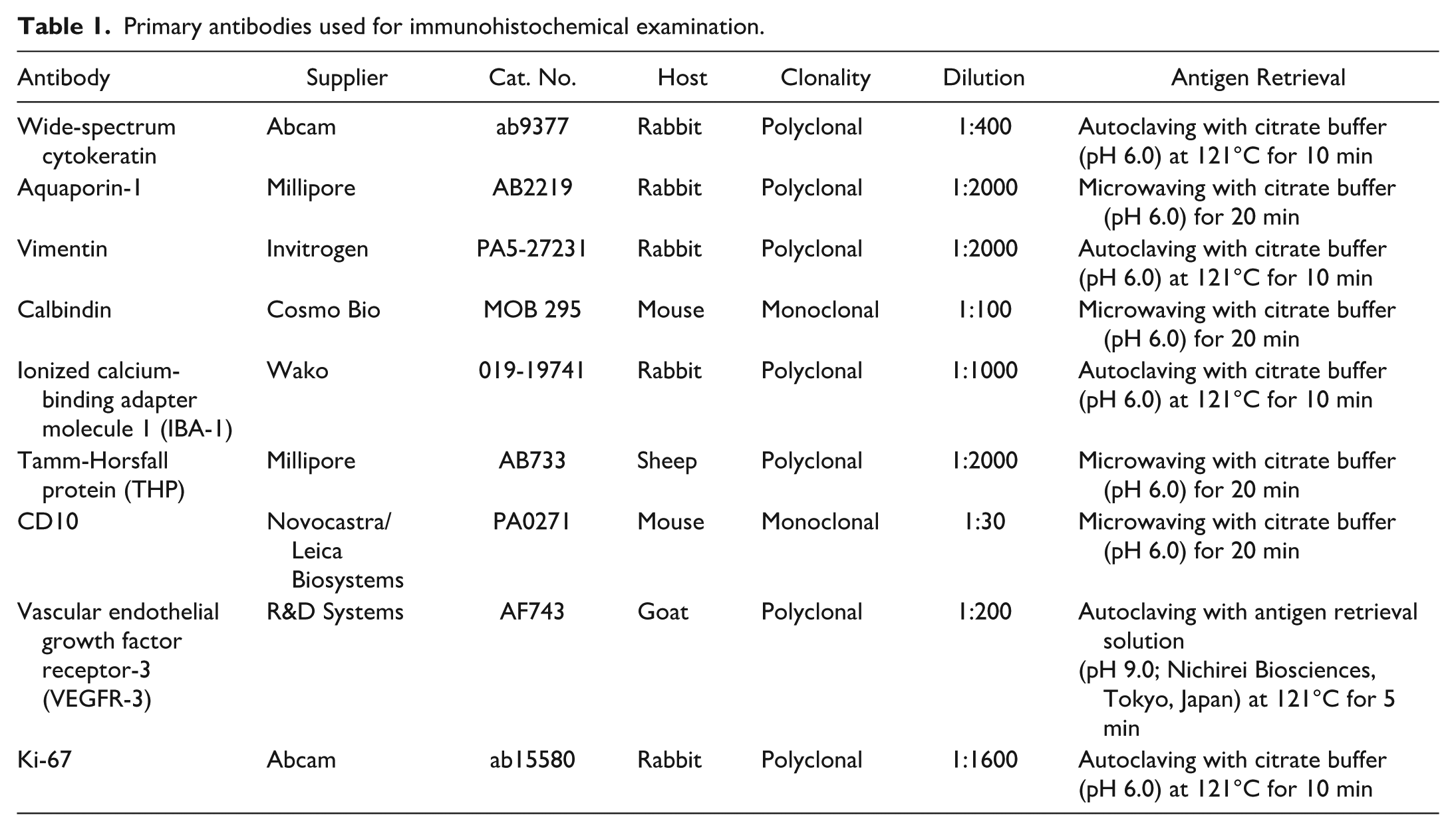
Microscopic examination revealed extensive renal damage. Severe glomerulonephritis with glomerulosclerosis was present, along with diffuse degeneration and necrosis of tubular epithelial cells involving proximal and distal tubules as well as collecting ducts, extending from the cortex to the medulla, accompanied by sloughing of epithelial cells into the tubular lumina. Some tubules were dilated, and hyaline casts were observed within the tubular lumina. Numerous regenerating tubules and interstitial fibrosis were also observed (Fig. 1a). In addition, predominantly at the corticomedullary junction, laminated concentric thickening of the arterial walls (onion-skin lesions) and necrosis of the surrounding interstitium with infiltration of neutrophils and macrophages were observed. The onion-skin lesions in arteries at the corticomedullary junction of the kidney were consistent with hypertensive vascular injury, and, together with the myocardial hypertrophy and the presence of hemosiderin-laden macrophages (heart failure cells) in the lungs, suggest that, in addition to the renal lesions, circulatory failure associated with sustained pressure overload and impaired left cardiac function may have contributed to death.
Histological features of a cynomolgus monkey with benign epithelial inclusions. Hematoxylin and eosin. (a) Kidney. Sclerotic glomeruli, interstitial fibrosis, numerous regenerating tubules (inset), tubular degeneration and necrosis, and dilated tubules containing hyaline casts are noted. (b and c) Renal hilar lymph node. Epithelial cell clusters are observed in the subcapsular sinus. They appear as small tubules (arrows), small aggregates (arrowheads), or petal-like clusters (double arrow). Calcification is present in a subset of clusters (asterisk). (d) Kidney. Epithelial cell clusters (arrowheads) in lymphatic vessels of the renal hilum.
In the renal hilar lymph node, epithelial cell clusters were observed, predominately in the subcapsular sinus. These clusters showed diverse morphologies including small tubules, aggregates, and petal-like formations (Fig. 1b, c). The petal-like clusters exhibited structures resembling inverted tubules, with the basal aspect of the cells oriented toward the center. Some clusters contained stromal-like components, occasionally with inflammatory cells at their cores. Clusters were attached to the lymphatic sinus wall, and others appeared to float freely within the sinus. The cells were homogeneous and cuboidal with pale eosinophilic cytoplasm. No nuclear atypia or mitotic figures were observed.
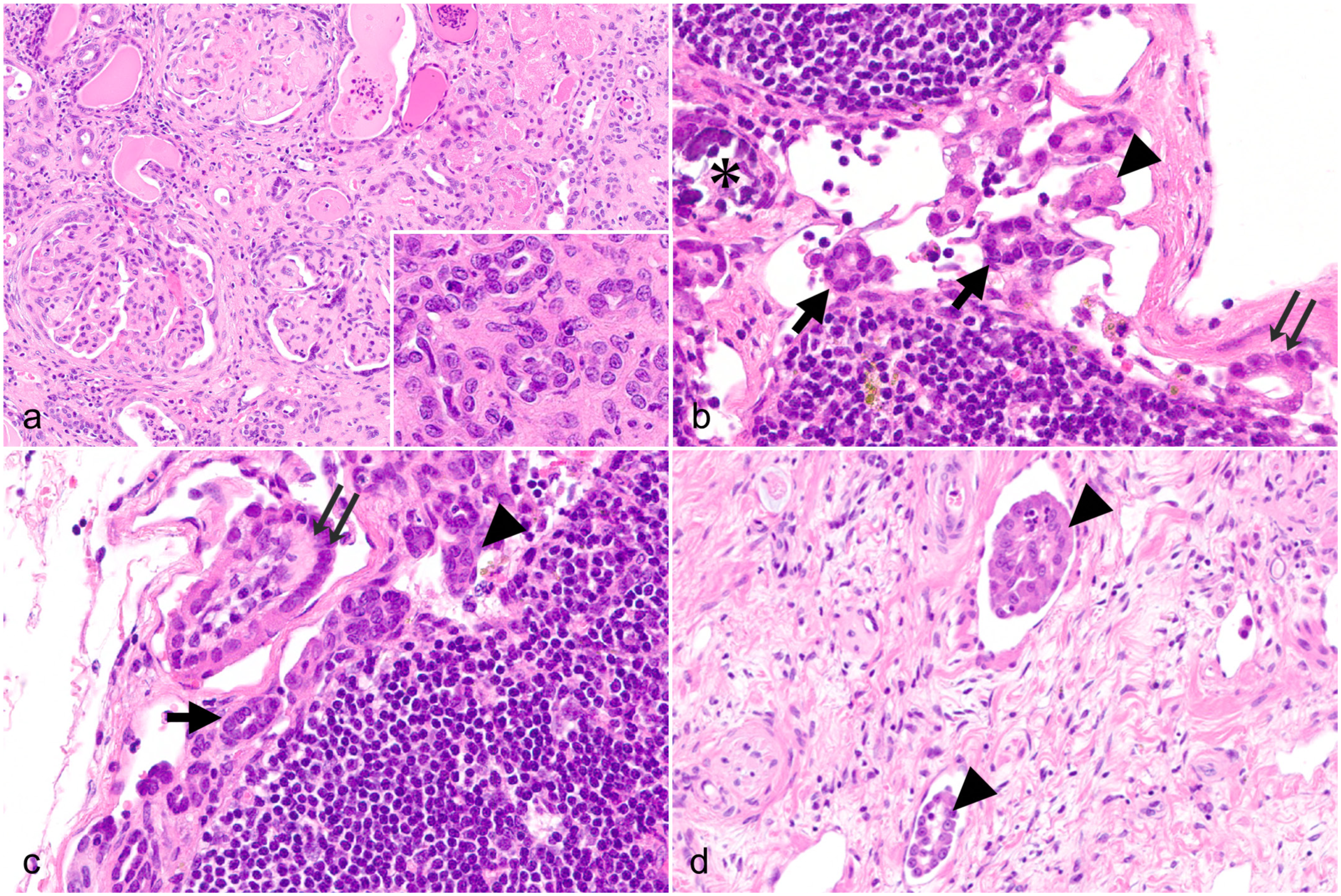
Calcification was noted in a subset of clusters. Similar clusters were also observed within the renal pelvic lymphatic vessels adjacent to the renal parenchyma (Fig. 1d). Rarely, epithelial cell clusters were seen protruding from the renal parenchyma across VEGFR-3-labeled lymphatic endothelium into the lumen, where similar epithelial clusters were present (Fig. 2a).
Histological, immunohistochemical, and ultrastructural features of the benign epithelial inclusions (BEIs). Cynomolgus monkey. (a) Kidney. Tubular epithelium migrating within lymphatic vessels; the endothelium is immunolabeled for vascular endothelial growth factor receptor 3 (VEGFR-3). VEGFR-3 immunohistochemistry (IHC). (b) Renal hilar lymph node. BEIs are immunolabeled for wide-spectrum cytokeratin. Wide-spectrum cytokeratin IHC. (c) Renal hilar lymph node. BEIs are immunolabeled for aquaporin-1. Aquaporin-1 IHC. (d) Renal hilar lymph node. Small tubular clusters are surrounded by a basement membrane-like structure (arrowheads) that is positive by Watanabe’s silver stain. (e and f) Renal hilar lymph node. Transmission electron microscopy reveals the presence of adhesive junctions (arrows) and short microvilli (arrowheads) on the surface of cell membranes at the periphery of BEIs. †, lumen; *, wall of the lymphatic sinus. Scale bar = 1 μm.
Immunohistochemically, the epithelial clusters were positive for wide-spectrum cytokeratin (Fig. 2b) and aquaporin-1 (Fig. 2c) but negative for vimentin, calbindin, IBA-1, THP, CD10, and VEGFR-3. No Ki-67-positive cells were observed in these epithelial cell clusters. Special staining revealed basement membranes surrounding the small tubular clusters, which were positive for Watanabe’s silver stain (Fig. 2d) and by periodic acid-Schiff reaction. In contrast, basement membranes were absent around the small aggregates and petal-like clusters.
Ultrastructural examination revealed adhesive junctions (Fig. 2e) between the cells of the petal-like structures floating within the lymphatic sinus without continuity with surrounding tissues. The cells had short microvilli present on the outer surfaces of the clusters (Fig. 2f). Examination of all other organs in this animal using the same procedures revealed no neoplastic lesions. Based on these findings, the cell clusters were diagnosed as BEIs. No similar lesions were observed in other lymph nodes examined, including the mesenteric and mandibular lymph nodes.
BEIs are ectopic clusters of benign epithelial cells observed in lymph nodes. Renal BEIs have previously been reported in association with pediatric tumors or obstructive renal disease in humans.16–19 In the present case, the cell clusters observed in the renal hilar lymph node were identified as epithelial cells, as they were immunohistochemically positive for wide-spectrum cytokeratin, and adhesive junctions were identified with transmission electron microscopy. Moreover, the absence of a primary tumor, along with homogenous nuclear morphology, lack of mitotic figures, and absence of Ki-67-positive cells, supports the exclusion of metastatic malignant lesions. Accordingly, these cell clusters are classified as BEIs.
Regarding the cellular origin, the presence of cell clusters within the renal pelvic lymphatic vessels, their immunopositivity for aquaporin-1, and the observation of microvilli on ultrastructural examination suggest that these clusters share features with proximal tubular epithelial cells. In contrast, the BEIs in the present case were negative for vimentin, which is not consistent with mesothelial cells, as mesothelial cells generally co-express cytokeratin and vimentin. 10 Considering the severe parenchymal damage observed in the kidney, we speculate that regenerating tubular epithelial cells entered the lymphatic lumen from sites of nephritic injury.
It should be noted that aquaporin-1 expression is not specific to renal tubular epithelium and has also been reported in other tissues; particularly in humans, high expression has been reported in the choroid plexus of the brain, pancreatic duct epithelium, and biliary epithelium. However, in the present case, considering the anatomical relationship between the kidney and the renal hilar lymph node, a renal tubular epithelial origin with lymphatic transport to the renal hilar lymph node appears most plausible. 6 In fact, the observation of regenerating tubular epithelial cells protruding into the lumen of lymphatic vessels, as well as the presence of epithelial clusters floating within the same lumen, supports this hypothesis.
The BEIs in our case were positive for aquaporin-1 but negative for CD10, a marker of proximal tubules. In the renal parenchyma, mature proximal tubules with normal architecture were positive for both aquaporin-1 and CD10, whereas most regenerating tubules, morphologically similar to the BEIs, were positive for aquaporin-1 but negative for CD10. This finding is considered to reflect differences in the degree of differentiation in regenerating tubules. In the present case, the BEIs were positive for aquaporin-1 but negative for CD10, as seen in regenerating tubules, suggesting that the BEIs originated from regenerating tubular epithelial cells.
In reports of kidney-derived BEIs in humans, BEIs have been described in association with THP depositions, suggesting that distal tubular epithelium migrates via lymphatic vessels to the lymph nodes along with THP.16,19 However, no such depositions were observed in the present case. In our case, the BEIs are considered more likely to have originated from proximal tubules, and differences in the cellular origin of BEIs might be contributing to the absence of THP depositions.
In the present case, some of the BEIs formed mature small tubular structures with basement membranes. Similar tubular structures have also been documented in a previous report. 19 The report suggests that the basement membrane forms as a result of reconstruction within the lymphatic sinus through interactions with stromal components that migrate alongside the epithelial clusters.
Petal-like clusters with a loss of polarity and an inverted appearance were also observed. However, the reason for such morphological differences remains unclear and can be regarded as an intriguing morphological feature. A plausible explanation is that regenerating tubules lose their basement membrane during detachment into the lymphatic vessels, fail to maintain their tubular structure, and are subsequently transported to the lymph nodes in an inverted state. Another possibility is that cell aggregates transported to the lymph nodes undergo reconstruction while retaining their loss of polarity. The formation of epithelial structures with reversed polarity (basal-in and apical-out) has been reported in proximal tubule organoid models, which may support the present hypothesis. 7 Generally, since extracellular matrix and renal stromal cells are considered crucial for tubulogenesis and differentiation,14,15 the environment within lymphatic vessels or lymphatic sinuses, which lacks extracellular matrix, may contribute to the loss of intrinsic architecture.
It is uncertain whether the BEIs in this case would engraft and survive over time. The calcification observed in some of the BEIs is consistent with dystrophic calcification arising secondary to prior cellular necrosis. This finding suggests that at least some BEIs may undergo degenerative change over time, rather than persist indefinitely within the lymphatic sinuses.
Reports of BEIs in animals are extremely limited. BEIs have been reported in dogs with pericardial effusion, 8 in normal cattle, 4 and in a bottlenose dolphin with localized pulmonary rupture, 2 all of which are considered to be of mesothelial cell origin. To date, no reports exist in animals describing BEIs suspected to originate from sources other than mesothelial cells.
To our knowledge, this case report is the first documented instance of kidney-derived BEIs in animals. Furthermore, while BEIs originating from the distal tubules during obstructive kidney disease caused by caliceal calculi or centrally located tumors have been reported in humans,16,17,19 no reports, including in humans, have documented renal tubule-derived BEIs arising from other etiologies.
Although various hypotheses exist regarding the pathogenesis, this case presents evidence of the tubules originating from the kidney and migrating lymphatically to the lymph nodes. Furthermore, it suggests that BEIs can develop in animal lymph nodes in association with inflammation and severe organ damage in the organ of origin. These findings are expected to help prevent overdiagnosis of BEIs as tumor metastasis in veterinary surgical pathology.
Footnotes
Acknowledgements
We thank Mr James Harada and Ms Kanae Tamatsukuri for their help with language editing.
Author Contributions
MU performed histologic evaluations and drafted the manuscript. YT conducted research on renal lesions. TD provided guidance on the interpretation of pathological lesions. NS and YN assisted with histologic evaluations. JS critically reviewed and edited the manuscript and contributed to histologic evaluations.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.