
Abstract
In March and April of 2023, an outbreak of highly pathogenic avian influenza virus, H5N1 strain Eurasian lineage goose/Guangdong H5 clade 2.3.4.4b, resulted in at least 17 mortalities of free-ranging California condors (Gymnogyps californianus) in Arizona. Condors presented dead or with neurologic signs and lethargy. Infection resulted in multisystemic inflammation and necrosis that most consistently and severely affected the brain, spleen, and adrenal glands. Immunohistochemistry performed on tissues from a subset of condors labeled cells in multiple organ systems for influenza A virus, most abundantly in neurons, epithelial cells, and mononuclear inflammatory cells and often colocalized with areas of inflammation. Acute blunt force trauma, presumably from ground collision after falling from a height, was a common finding and indicated a rapidly debilitating disease course and neurologic impairment. Hepatic lead concentrations were relatively low with no concurrent incidences of acute lead toxicosis, an otherwise common cause of death in free-ranging California condors. Bone lead reflected long-term lead accumulation in several condors. Assessment of ingesta in 8 condors via morphologic hair identification showed a mix of consumed taxa, most commonly Bovidae. In summary, California condors, like other New World vultures (family Cathartidae), are highly susceptible to this strain of H5N1, and this should be taken into consideration when planning release, feeding, and morbidity and mortality responses.
Keywords
The California condor (Gymnogyps californianus) is a New World vulture native to the western USA. They are a critically endangered species and have been protected under the federal Endangered Species Act since 1966. Historically, California condor ranges spanned the continent but dwindled to the western United States following extinctions of megafauna, which were an important food source. During the last century, anthropogenic stressors such as environmental lead contamination, indiscriminate shooting, and unintentional poisoning secondary to predator control efforts caused a precipitous drop in numbers. 38 To save the species from extinction, the California Condor Recovery Program was formed, and in the 1980s, the remaining 23 wild condors were taken into captivity for breeding and reintroduction. Currently, there are free-ranging populations in California, Arizona, and Utah collectively comprising approximately 350 individuals. 32 Lead poisoning is still the most common cause of mortality in wild adult condors.16,21,35,38,47
Condors are a long-lived species with a natural lifespan thought to be as long as 60 years 2 . These birds are slow to mature and have a low reproductive rate, typically laying a single egg every 2 years. Both the female and male are invested in the care of the egg and chick. Because of the ongoing risk of lead toxicosis as well as low reproductive rates, every California condor represents a valuable member of the population, and even a small disease outbreak could have devastating consequences.
The current clade of circulating highly pathogenic avian influenza virus (HPAIV), Eurasian H5N1 clade 2.3.4.4b, was first detected in North America in 2021 and has since spread throughout the continent, including all the US states in which California condors reside.6,44 This outbreak is notable for the effects on raptors, including vultures.13,19,30,33,49 There has also been spillover to mammals—including domestic cattle and domestic and wild Felidae species. This increases the risk of infection through scavenging, particularly on cattle—a food source where condor territory overlaps cattle ranches.10,26,34,42,43
In March and April of 2023, 17 adult condors from northern Arizona were confirmed dead from an outbreak of HPAIV. This represents a substantial portion of the condor population in Arizona, which numbered around 100 prior to the outbreak. Within this 1-month period, 4 more birds were listed as deceased based on telemetry data, but carcasses were unrecoverable. 46 In addition, 4 known infected condors were hospitalized, successfully treated, and later released, bringing the total number of known or suspected HPAIV infections to 25. 22 This postmortem study describes the gross and histologic findings in the 17 confirmed mortalities and viral antigen localization in a subset of cases taken from the beginning, middle, and end of the outbreak. Systemic lead concentrations and species of ingested carrion were also assessed to help determine risk factors for mortality.
Materials and Methods
Case Submissions
All condors were found dead in the field or were hospitalized and died before or during treatment. Histories were compiled for all birds that included age, location found, estimated death date, and any recent medical records. Age was determined from the California Condor Recovery Program studbook. Condors were assigned a number in chronological order of submission with case 1 being assigned to the earliest received mortality.
Necropsy and Imaging
All condor carcasses were sent to the U.S. Fish and Wildlife Service National Forensics Laboratory within 48 hours of retrieval. At the lab, carcasses were kept refrigerated from intake to necropsy. Necropsies were completed within 24 hours of receipt for all except 2 birds, which were necropsied after 4 days of refrigeration. Necropsies included full-body orthogonal radiographs and a complete external and internal examination by an American College of Veterinary Pathologists board-certified pathologist (TCV and RAK). All necropsies were conducted in a biosafety level 2 laboratory. Personal protective equipment included protective clothing, gloves, and National Institute for Occupational Safety and Health-approved N-95 respirators. Nutritional condition was subjectively assessed as good, fair, or poor based on the amount of subcutaneous and visceral adipose, skeletal muscling, and body weight.
Histology
Tissues for histology were fixed in 10% zinc formalin for at least 72 hours before further processing. Full tissue sets were collected except for cloaca, tongue, and pancreas, which were not consistently sampled. Tissues were routinely processed and paraffin-embedded, and 4-µm-thick tissue sections were stained with hematoxylin and eosin. Slides were evaluated by either TCV or RAK. Autolysis was subjectively assessed based on loss of cellular differential staining, postmortem lysis, and epithelial cell loss.
Inflammatory lesions, which included leukocyte infiltrates, necrosis, and hemorrhage, were scored by RAK using a subjective and semi-quantitative grade of absent (0), minimal (1), mild (2), moderate (3), or severe (4). Inflammation was considered minimal if there were only a few small areas of inflammation that did not always appear in a single 19.63 mm2 field. A score of mild was given to tissues with small areas of inflammation more consistently present but affecting less than 10% of the tissue in a 19.63 mm2 field. A score of moderate was given to tissues with larger individual, occasionally coalescing foci that affected approximately 10% to 50% of the tissue in a 19.63 mm2 field. Severe inflammation consisted of tissues with coalescing to diffuse lesions that affected more than 50% of the tissue in a 19.63 mm2 field.
Immunohistochemistry
A subset of 5 condors was chosen for immunohistochemical labeling with priority given to those with lowest levels of autolysis as assessed on hematoxylin and eosin-stained sections. Tissues were tested from condors that had died near the beginning (case 1), middle (case 6), and end (case 12) of the outbreak as well as 2 condors that had histologic evidence of a slightly more chronic course of disease (case 15 and case 17). Immunohistochemistry (IHC) was performed in the Histology Laboratory at the University of Georgia and followed the methods of Andreasen et al. 1 The positive control was lung from a ferret experimentally infected with HPAIV. A single isotype control slide was used for all condor slides.
Immunolabeling was scored by NMN using a subjective and semiquantitative grade of absent (0), minimal (1), mild (2), moderate (3), and severe (4) and followed the general scheme detailed in Lean et al. 24 The pattern of labeling was considered minimal if limited to single, isolated cells or a single, small cluster of cells. Labeling was mild if present in clusters of 2 to 5 cells, frequent individualized cells, or a single large focus of positive cells. The labeling pattern was moderate if present in multiple clusters of more than 6 cells or hundreds of widely scattered cells with strong labeling. The labeling pattern was severe if tissues had coalescing to diffuse, strong cellular labeling. In addition, weak, moderate, and strong labeling of individual cells in each tissue evaluated was subjectively determined based on the brightness of the labeled portion (nucleus or cytoplasm) of each cell.
Viral polymerase chain reaction
Spleen was the tissue of choice for viral testing because of its immune system function, the reliable presence of viral lesions in infected condors, and the relative persistence of splenic tissue in the face of decomposition. Because the carcasses were so rapidly retrieved and shipped for necropsy, field collection and testing of cloacal and choanal swabs was not done. Spleens from all condors were submitted frozen to the Oregon State University Veterinary Diagnostic Laboratory, a biosafety level 2 laboratory, for avian influenza real-time reverse transcription polymerase chain reaction (rRT-PCR) using the standardized protocols for the National Animal Health Laboratory Network for HPAI virus rRT-PCR (NVSL-SOP-0068).31,39 The aim of the analyses was to determine the presence or absence of H5 influenza.
A 0.2-g portion of each spleen was minced using a sterile scalpel and transferred to a 2.0-ml Safe-Lock (Eppendorf) tube containing a 5-mm stainless steel grinding ball and 1 ml of Dulbecco’s modified Eagle’s medium. The sample was homogenized using a TissueLyser (Qiagen) for 2 min at 18 Hz. A 200-µl aliquot of the tissue homogenate was transferred to a deep-well extraction plate and total nucleic acids were extracted using the MagMAX CORE Nucleic Acid Purification Kit (ThermoFisher) run on a KingFisher Flex magnetic particle processor according to the manufacturer’s protocol. Extracted nucleic acid samples were tested using a general influenza A virus (IAV) rRT-PCR targeting a conserved region of the IAV matrix gene (Spackman & Suarez 2008, NVSL-SOP-0068). 39 Samples testing positive for IAV matrix gene were further tested using rRT-PCR targeting the avian influenza H5 hemagglutinin gene for the North American, Eurasian, and Mexican lineages of IAV.39,45
Samples testing positive for IAV-H5 were forwarded to the National Veterinary Services Laboratory for confirmatory testing and lineage determination using the procedure as in Lamb et al 22 (in press); Nguyen et al 34 ; and Youk et al. 50
Tissue Minerals
Liver samples from all condors were taken at the time of necropsy and fixed in 10% zinc formalin for at least 72 hours before being sent for assessment of tissue mineral concentrations. Bone lead and other heavy metals were determined from sections taken from the mid to distal tibiotarsi. Bone samples were taken from 6 of the condors (cases 3, 8, 10, 13, 15, and 16). Bone was not available from all condors because the collection of this tissue was done retrospectively. Liver and bone were sent to the Michigan State University Veterinary Diagnostic Laboratory for sample processing and mineral analysis. Due to concerns with known or suspected HPAIV infection, testing for tissue minerals was done only after formalin fixation to inactivate any virus. Formalin-fixed tissues are considered adequate substitutes for fresh tissues for the analysis of trace elements in liver and bone as assessed through internal validation and in several published studies.7,28 Liver results were compared with reference ranges for bald eagles compiled by the Michigan State University Veterinary Diagnostic Laboratory. Bone mineral levels were compared with concentrations obtained by Martin et al. 27
For sample preparation, liver and bone were dried overnight in an oven with temperatures ranging 85°C to 90°C. They were removed from the oven, allowed to cool, and weighed. Tissue samples (<0.2 mg) were digested with 2 ml concentrated 67% to 70% nitric acid (Aristar Plus, VWR, Radnor, PA, USA) at 95°C overnight in 15 ml polypropylene digestion vessels. Bone samples (approx. 0.5 mg) were brought to a final weight of 25 g with the addition of concentrated 67% to 70% nitric acid (Aristar Plus, VWR, Radnor, PA, USA) and subsequently digested at 95°C overnight in 50 ml polypropylene digestion vessels. The digests were then diluted 1:10 and 1:100 in deionized water (ELGA Purelab Flex, Woodridge, IL, USA) prior to analysis.
For mineral analysis, sample digests were analyzed by inductively coupled plasma mass spectrometry (Agilent 7900 ICP-MS, Agilent Technologies, Santa Clara, CA, USA). Elemental concentrations were calibrated using a 6-point linear curve of the analyte–internal standard response ratio. Calibration standards were from Inorganic Ventures (Christiansburg, VA, USA.) The National Institute of Standards and Technology (NIST, Gaithersburg, MA, USA) Standard Reference Materials (Bovine Liver 1577c, Mussel 2976, and Bone Ash 1400b) were used as controls. A second source calibration verification (High Purity Standards, North Charleston, SC, USA) was also used for quality control on the inductively coupled plasma mass spectrometry.
The limits of quantitation for the elements of interest in fixed tissue were as follows: Cu 2.0 μg/g, Fe 5.0 μg/g, Pb 0.1 μg/g, Se 0.1 μg/g, and Zn 0.5 μg/g. The limits of quantitation for the elements of interest in bone were: Ca 2,500 μg/g, Mg 40 μg/g, Mn 2.5 μg/g, Pb 5 μg/g, and P 1000 μg/g.
Hepatic lead concentrations were considered to be toxic at >30 µg/g dry weight (dw), elevated at <30 µg/g dw and ≥6 µg/g dw, and consistent with background exposure at <6 µg/g dw. Bone lead was considered to be potentially toxic at >10 ppm. 27 A diagnosis of acute toxicity was made based on liver lead concentration.
Hair Identification
Hairs from the upper gastrointestinal tract lumen were retrieved from cases 3, 8, 9, 12, 13, 14, 15, and 16. For viral inactivation, all hairs were immersed in 10% zinc formalin for a minimum of 72 hours before further processing. Hairs were lightly washed with water, blotted dry, and mounted on slides for light microscopic examination. A subsample of black hairs from case 8 was treated with hair bleach, as per packaging instructions, for 45 minutes prior to being mounted on a slide.
Hairs were examined microscopically by RAK, and morphologic features of guard hairs were compared with descriptions in the literature and a known reference sample set to determine order or family of origin.9,29 In cases where guard hairs were not present and/or hairs were too degraded for definitive determination, hair origin was described as undetermined.
Results
The outbreak lasted approximately 1 month, with unusual behavior being observed in the field as early as 9 March and the first mortality being found on 20 March 2023. 46 The last known mortality occurred in the hospital on 12 April 2023. There were 9 males and 8 females. The age of the condors skewed toward adults with a mean age of 10.76 years and a median age of 9 years. The youngest birds were 2 years old, and the oldest bird was 28 years old. The most common age was 2 years (4/17). Signalment and dates of hospitalization (if applicable) and death are recorded in Table 1.
Signalment and histories from mortalities of California condors naturally infected with clade 2.3.4.4b highly pathogenic A/goose/Guangdong/1/1996 H5 influenza A virus.
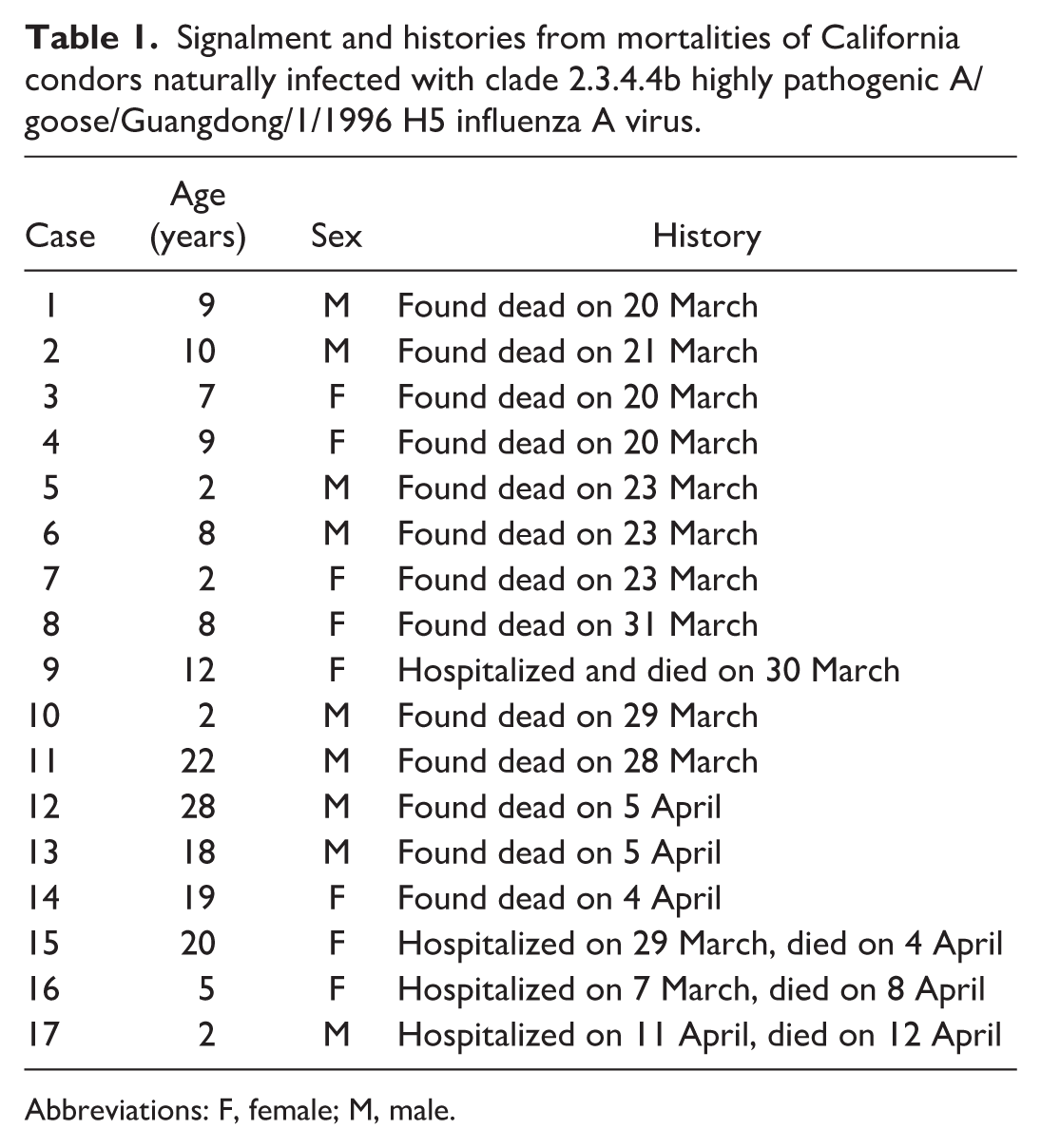
Abbreviations: F, female; M, male.
Based on telemetry data, 11 of 17 mortalities were confirmed to have occurred in 6 separate small clusters, all within an approximately 730 square mile area in a national monument. This area was remote from ranches or human habitation but close to and at established condor release sites. Five of the remaining 6 condors were found at or very near 1 of the 6 clusters, but precise coordinates were not provided. No location information was provided for the sixth bird. Three of the condors were retrieved alive and hospitalized. Two died the day after admission, and the third (case 15) died after 6 days.
Radiographic Findings
Metal particulates were present in the gastrointestinal content of 1 bird (case 1) and consisted of a few pinpoint fragments in the crop. There were radiographically visible acute bone fractures in 2 birds. Case 3 had multiple wing fractures and a femoral fracture. Case 5 had a left humeral fracture, a right ulnar fracture, and a right sternal fracture at the coracosternal joint.
Gross Findings
Lesions directly attributed to viral infection were present in the spleen, liver, and gastrointestinal tract. There was enlargement of the spleen and liver of 13 of 17 and 10 of 17 birds, respectively (Fig. 1a, b). Affected spleens were variably firm and discolored dark red to tan. Signs of hepatomegaly included displacement of adjacent viscera and rounded lobe margins (Fig. 1b, c). Nephromegaly was also noted in 1 condor. Recognition of organ enlargement was based on experience. To the authors’ knowledge, there are no published organ measurements for this species.
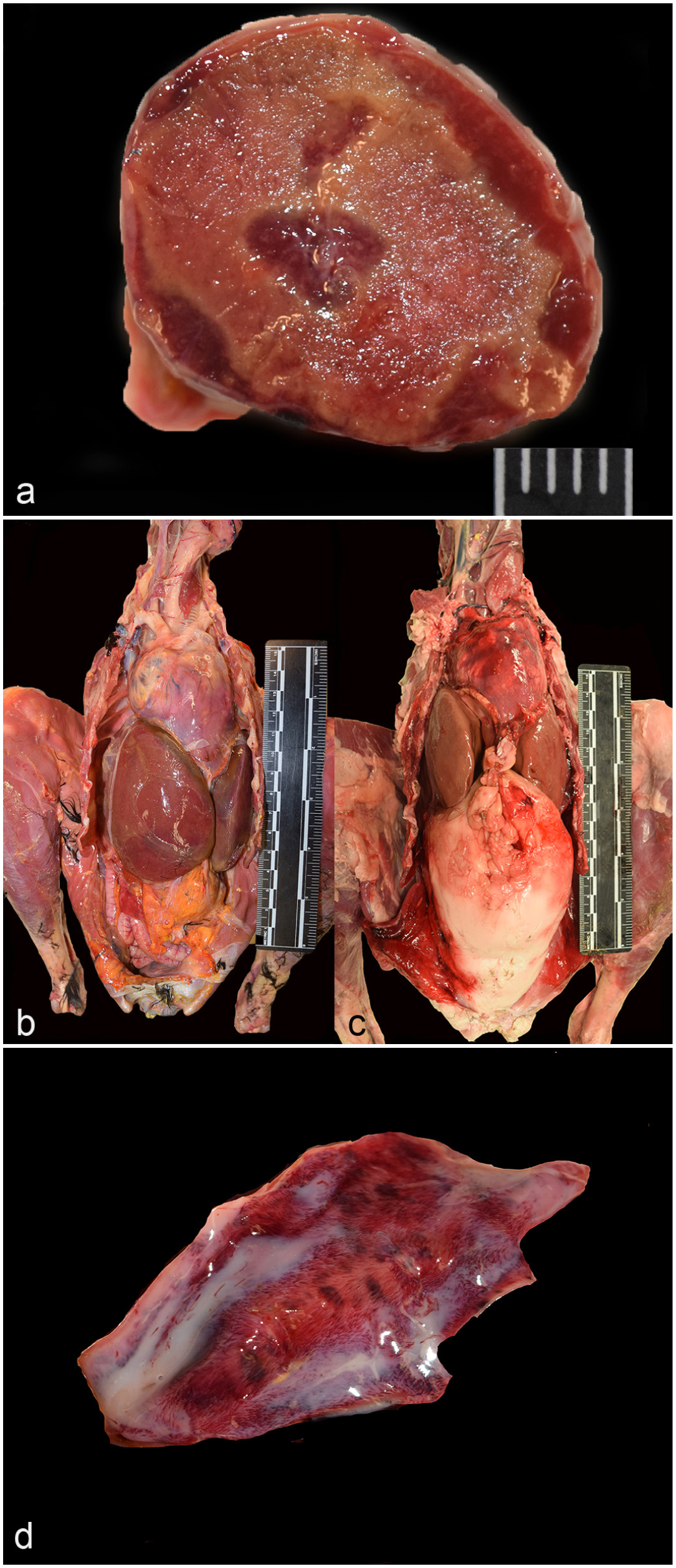
Highly pathogenic avian influenza, H5 clade 2.3.4.4b, California condors. (a) Spleen, cut section. The spleen is enlarged and firm with coalescing pale foci of necrosis. Scale = 5 mm. Case 15. (b) Liver enlargement. Case 15. (c) A grossly normal liver in an uninfected California condor showing the expected organ size. (d) Duodenum. The mucosa is mottled dark red. Case 11.
Multifocal to coalescing mucosal hemorrhages of the gastrointestinal mucosa were grossly visible in 2 of the birds (Fig. 1d). In case 11, the gastrointestinal mucosal hemorrhages were present throughout ventriculus, duodenum, and cloaca. In case 14, hemorrhages were in the small intestinal mucosa and the cloaca. Multifocal, pinpoint areas of pancreatic hemorrhage were present in cases 12 and 14.
There were lung lesions in 2 of the hospitalized condors suggestive of a concurrent bacterial and/or fungal infection. One consisted of masses suspected to be granulomas (case 15), and the other appeared grossly as regional tan discoloration (case 17).
There were signs of blunt force trauma in 12 of 17 birds. This included bone fractures in 6 birds, cutaneous bruising of 8 birds, and internal organ contusions and/or lacerations in 4 birds. Blunt force trauma in case 5 was severe enough to have been the immediate cause of death with skull, wing, pectoral girdle, spine, and rib fractures and fracture and hemorrhage of multiple internal organs. In 4 condors (cases 4, 11, 12, and 14), skin on the nonfeathered portions of the head and neck had small to moderate numbers of scattered, small, often scabbed abrasions consistent with superficial bite wounds and were attributed to conspecific interactions (grooming behavior).
Most (14/17) of the condors were in good nutritional condition. The remaining 3 were in fair (slightly thin) nutritional condition. Crop and/or gastric content was present in small amounts in 15 of 17 condors and largely consisted of hair or other fibers and a variable mixture of gritty fluid, small rocks, and microtrash. One condor had a small, ingested bone in the crop.
Histologic and IHC Findings
Combined histologic grades and associated IHC grades for each examined organ are in Table 2, and individual results are in Supplemental Table S1. All condors had histologic lesions in the spleen. Most (15/16; 94%) were severely affected having multifocal to coalescing large areas with necrosis, heterophilic inflammation, and hemorrhage (Fig. 2a), which colocalized with immunohistochemical labeling for IAV antigen in all 5 tested condors (Fig. 2b). Labeling occurred within areas of necrosis and was intracytoplasmic (IC) in mononuclear cells and intranuclear (IN) in presumptive macrophages. Labeling ranged from mild to severe with an average grade of severe. Splenic vascular changes varied widely from mild to moderate arteritis to diffuse vascular necrosis with strong IHC labeling in arteriolar smooth muscle cells. In the spleen of case 17 (chronologically the last mortality), there was no necrosis, but there were moderate numbers of cells with foamy cytoplasm suspected to be macrophages.
Histopathologic grades and associated viral immunohistochemical (IHC) labeling grades of lesions attributed to infection with clade 2.3.4.4b highly pathogenic A/Goose/Guangdong/1/1996 H5 influenza A virus in California condors.
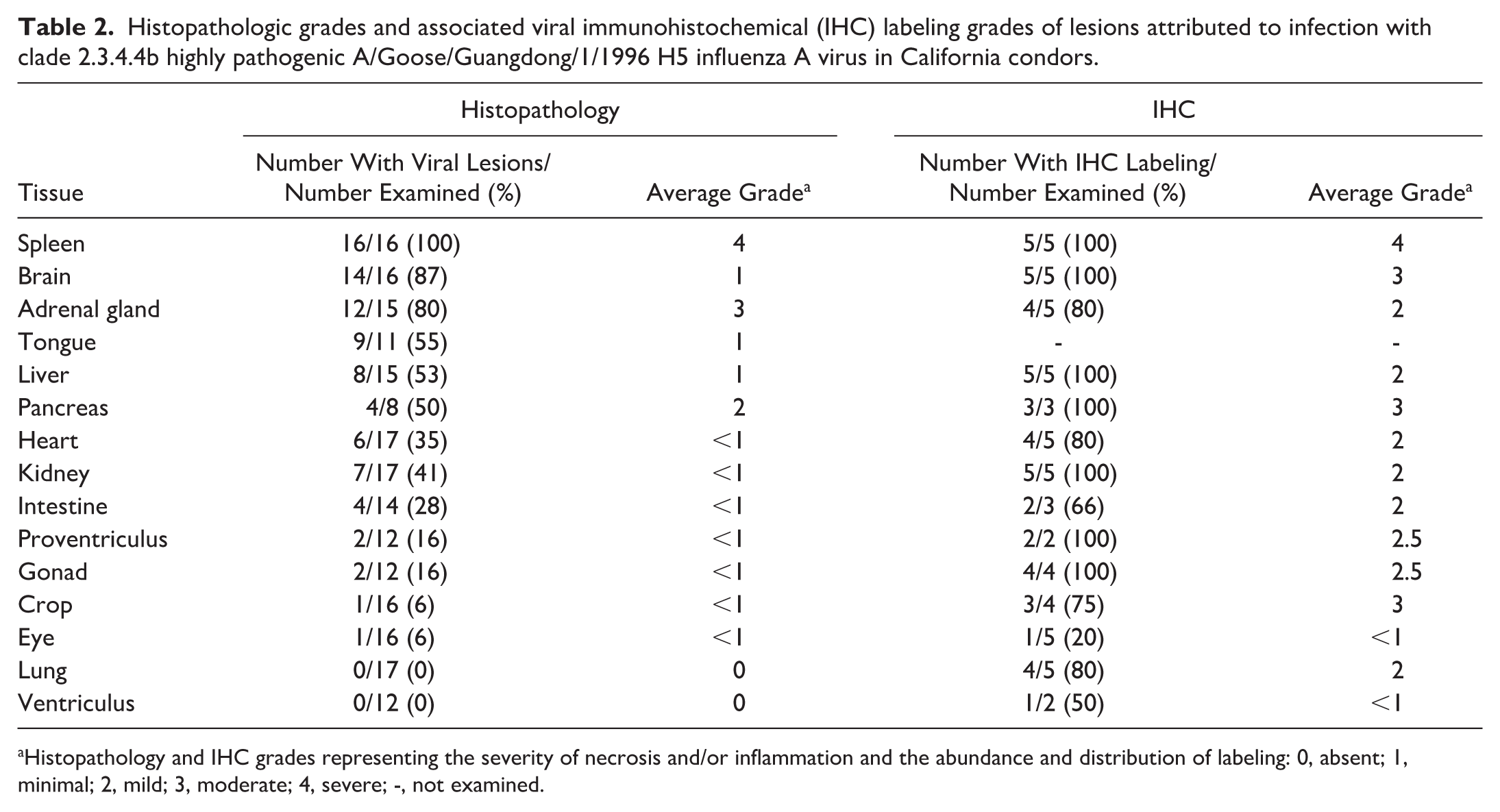
Histopathology and IHC grades representing the severity of necrosis and/or inflammation and the abundance and distribution of labeling: 0, absent; 1, minimal; 2, mild; 3, moderate; 4, severe; -, not examined.

Lesions in highly pathogenic avian influenza (HPAI), H5 clade 2.3.4.4b infected California condors. Hematoxylin and eosin (a, c, e) and immunohistochemistry for influenza A nucleoprotein (b, d, f, g, h). (a) Spleen. Multifocal to coalescent necrosis of lymphoid tissue. Case 15. (b) Spleen. Within this necrotic area, there is strong labeling of mononuclear cells and scattered labeling of debris and presumptive macrophages. Case 15. (c) Cerebrum. Encephalitis with a largely mononuclear cell population and occasional heterophils. Case 12. (d) Cerebrum. Neurons are strongly positive for influenza A antigen. Case 12. (e) Adrenal gland. There is multifocal to coalescent necrosis of the interrenal tissue with sparing of the chromaffin cells (asterisks). Case 6. (f) Adrenal gland. The cytoplasm of interrenal cells strongly labels for influenza A. There is no labeling of chromaffin cells (asterisks). Case 6. (g) Crop. Epithelial cells have strong intranuclear and weak intracytoplasmic immunopositivity. Case 12. (h) Lung. There are no pulmonary lesions associated with viral infection in HPAI-infected condors. There is rare influenza A immunolabeling of circulating mononuclear cells and background debris. Case 6.
Encephalitis was a feature of these infections (Fig. 2c) with moderate IHC labeling of neurons both within and remote from areas of inflammation (Fig. 2d). Most of the condors (14/16; 87%) had a minimal to mild encephalitis and/or meningitis. Lesions consisted of predominantly mononuclear inflammation and necrosis in the cerebral (6/16) and cerebellar (6/16) white and gray matter with occasional vasculitis. There was also rare heterophilic and lymphocytic inflammation in the choroid plexus, which was not present on all slides, and in the meninges. Areas of meningeal and perivascular inflammation were often accompanied by small numbers of erythrocytes. In case 16, there was multifocal cerebellar gliosis.
While hundreds of widespread neurons were estimated to be positive for IAV in the cerebrum, there was stronger individual labeling of neurons in the cerebellum, midbrain, and periventricular region. Labeling was often strong in nuclei and mild to moderate in the cytoplasm. Pyknotic debris in these areas presumed to be fragments of necrotic neurons is also labeled. In cases 15 and 17, there was IN and IC glial cell labeling.
The third most often affected organ was the adrenal gland. Adrenal glands were present in sections from 15 of 17 condors. Of these, there was multifocal necrosis in 12 of 15 (80%) of the condors. Necrotic areas were typically centered on adrenal cortical cords (interrenal cells) and were often accompanied by hemorrhage (Fig. 2e). Lesions ranged from mild to severe with an average grade of moderate. Viral antigen was present in 4 of 5 adrenal glands and was minimal to severe with an average grade of mild (Fig. 2f). Interrenal cell cytoplasm most consistently labeled, though medullary cell cytoplasm was minimally labeled in 2 of 5 adrenals.
Birds affected later in the outbreak (cases 14, 15, and 17) lacked acute adrenal necrosis and hemorrhage. Instead, interrenal areas were effaced or replaced by lymphoplasmacytic infiltrates and large numbers of vacuolated or foamy cells presumed to be macrophages. In case 17, there were also multifocal stellate colorless clefts within affected areas. Adrenal peripheral nerve ganglia in 1 of 5 condors had minimal neuronal labeling.
There was hepatic necrosis in 9 of 17 condors. Generally, lesions consisted of minimal, peracute, multifocal, random hepatocellular necrosis. There was often moderate to marked hepatocellular swelling and macrovesicular change. IHC labeling was mild and more prominent in hepatocytes in areas of necrosis. Hepatocellular labeling was both IC and IN, but more common in the former. There was also IN and IC labeling of bile duct epithelial cells.
The pancreas was examined histologically in 11 of 17 condors but was severely autolyzed in 3 of 11 condors. In the remaining 8 condors, there was minimal to severe inflammation and necrosis in 4 of 8 (50%) condors. Pancreas was available for IHC in 3 of 5 condors, and abundant labeling occurred in all 3, most consistently in the acinar epithelial cells (IC and rarely IN) as well as occasionally in mononuclear and ductal epithelial cells. Labeling did not necessarily colocalize with lesions, occurring in the absence of necrosis in 1 of the condors.
There was myocarditis in 6 of 17 (35%) of the condors. Most of those (4/6) were found in birds dying toward the end of the outbreak, on or after April 4. The first lesions to appear were in case 6 and consisted of mild endocardial lymphoplasmacytic inflammation. Later in the outbreak findings included mild, multifocal to coalescing myocardial degeneration and acute necrosis. Myocyte IHC labeling was present in 4 of 5 condors evaluated, with an average grade of mild. Antigenic presence was strongest in cardiomyocyte nuclei, often with concurrent weak, granular IC labeling. Small, distinct clusters of myocytes (from <10 to dozens) labeled, and these were widely scattered throughout the myocardium, involving left and right free walls and the septum. Strong labeling of case 12 was in the absence of histologic lesions.
Nephritis affected 7 of 17 (41%) of the condors ranging from tubular or tubulointerstitial necrosis of varying severity to mild lymphoplasmacytic interstitial inflammation. Kidneys in 5 of 5 individuals had minimal to confluent labeling for IAV antigen within tubular and/or glomerular epithelial cell nuclei regardless of whether there were histologic lesions.
There was mild to moderate intestinal mucosal necrosis in 4 of 14 (28%) of the condors. Tissues were available for IHC in 3 of 5 of the selected condors, and labeling was moderate in 2 and absent in 1. In case 12, moderate viral antigenic labeling was occasionally evident in pyknotic debris within mucosal necrotic foci, and there was weak to moderate labeling in necrotic glandular epithelial cells. In case 15, which had no histologic lesions, there was moderate labeling of the deep glandular epithelial cytoplasm. Proventricular histologic lesions were limited to the gastrointestinal-associated lymphoid tissue of cases 4 and 5 and consisted of lymphoid necrosis. Lymphoid tissues in both cases labeled mildly to moderately for viral antigen.
Reproductive tissues were rarely affected, with a lymphoplasmacytic endometrial infiltrate in case 15 and testicular interstitial lymphohistiocytic inflammation and Sertoli cell necrosis in case 2. None of the IHC-labeled condors had distinct histologic lesions in reproductive tracts, though case 12 had marked germ and Sertoli cell sloughing and tubular cellular debris accumulation that was difficult to interpret. Despite the lack of definitive histologic lesions, gonadal tissues consistently labeled for IAV antigen. In IHC-labeled tissues from the 3 male condors, seminiferous tubular epithelial cells labeled (IC in 2 and IN in 1), most often in sloughed cells. In the 1 set of female IHC-labeled tissues, occasional single-cell IC labeling was present in the stromal and ductal epithelia.
Crop lesions were limited to case 15 and consisted of acute necrosis in the deep mucosal layers. In contrast, labeling in the crop occurred not only in case 15, but also in 2 of 3 other condors that did not have histologic lesions. Labeling was moderate to severe, affecting large numbers and continuous segments of mucosal epithelial cells with strong IN labeling and weak to moderate IC labeling (Fig. 2g).
Ocular lesions were only present in case 14, manifesting as a histiocytic and heterophilic posterior uveitis with focal retinal necrosis. Weak IC labeling was present in the iridial epithelial cells of case 1 and was absent in the rest. Other tissues evaluated with IHC included bone marrow from 2 condors; 1 with no labeling and the other with IC and IN labeling of a few mononuclear cells. No IHC labeling was evident in opportunistically evaluated adipose tissue around the heart, intestinal serosa, kidneys, and cloaca. There were no histologic lesions in the ventriculus (12/17 examined). There was minimal labeling with IAV in 1 of 2 in the glandular epithelium.
There was glossitis in 9 of 11 (55%) of the condors. Glossitis was superficial and mild in all cases except in case 5, where the entire dorsal surface of the tongue was affected. Lesions included mucosal erosion and ulceration with heterophilic inflammation, generally at the base of the tongue. There was also multifocal swelling and congestion of the lingual glandular epithelium with rare acute epithelial necrosis. The tongue was not reliably sampled nor was it included in the immunohistochemical evaluation.
Lung inflammation was only present in 2 of 17 condors and consisted of granulomatous fungal pneumonia and an acute, necrotizing fungal and bacterial pneumonia. Fungi in both cases had features consistent with Aspergillus species. IHC for IAV did not label any of the examined lung parenchyma, although there was occasional labeling of circulating mononuclear cells (Fig. 2h).
Viral Characterization
All spleens tested positive by avian influenza rRT-PCR and were characterized as fully Eurasian HPAIV H5N1 clade 2.3.4.4b genotype A3 per GenoFlu (GitHub—USDA-VS/GenoFLU: Influenza data pipeline to automate genotyping assignment). Virus sequences were consistent with other wild bird sequences from Colorado and Wyoming. Ct values for Subtype H5 clade 2.3.4.4 were between 9.28 and 31.74 (Table 3).
California condor splenic rRT-PCR Ct values.
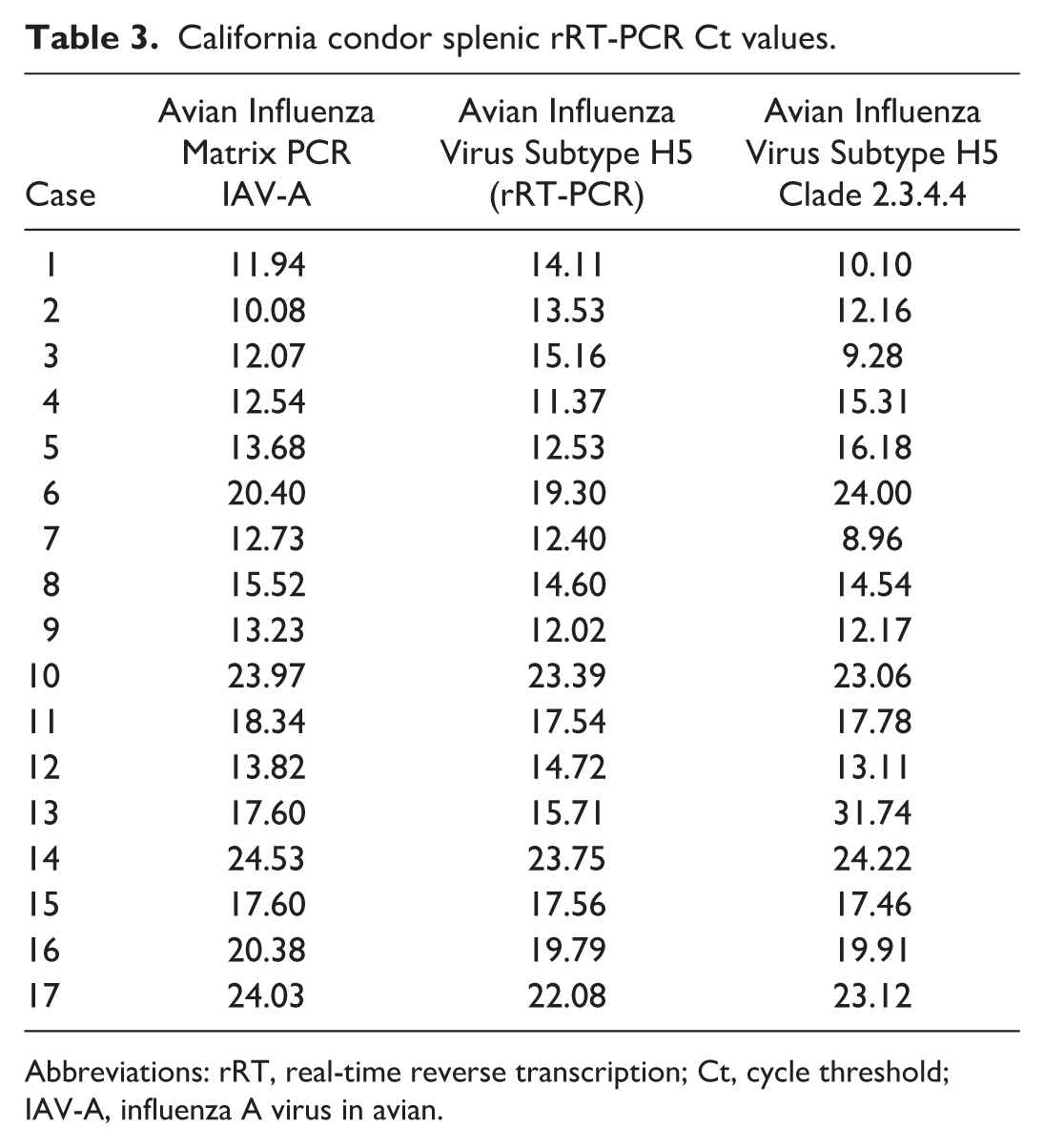
Abbreviations: rRT, real-time reverse transcription; Ct, cycle threshold; IAV-A, influenza A virus in avian.
Tissue Minerals
Hepatic and bone lead levels are listed in Table 4. The highest lead concentration was 11.99 µg/g dw in case 1. None of the condors had concentrations or postmortem findings indicative of clinical lead toxicosis. Lead concentrations in 3 of 17 condors were elevated but remained below toxic levels. Low levels of lead consistent with background exposure were present in the remaining 13 condors.
Lead concentrations from mortalities of California condors naturally infected with clade 2.3.4.4b highly pathogenic A/Goose/Guangdong/1/1996 H5 influenza A virus.
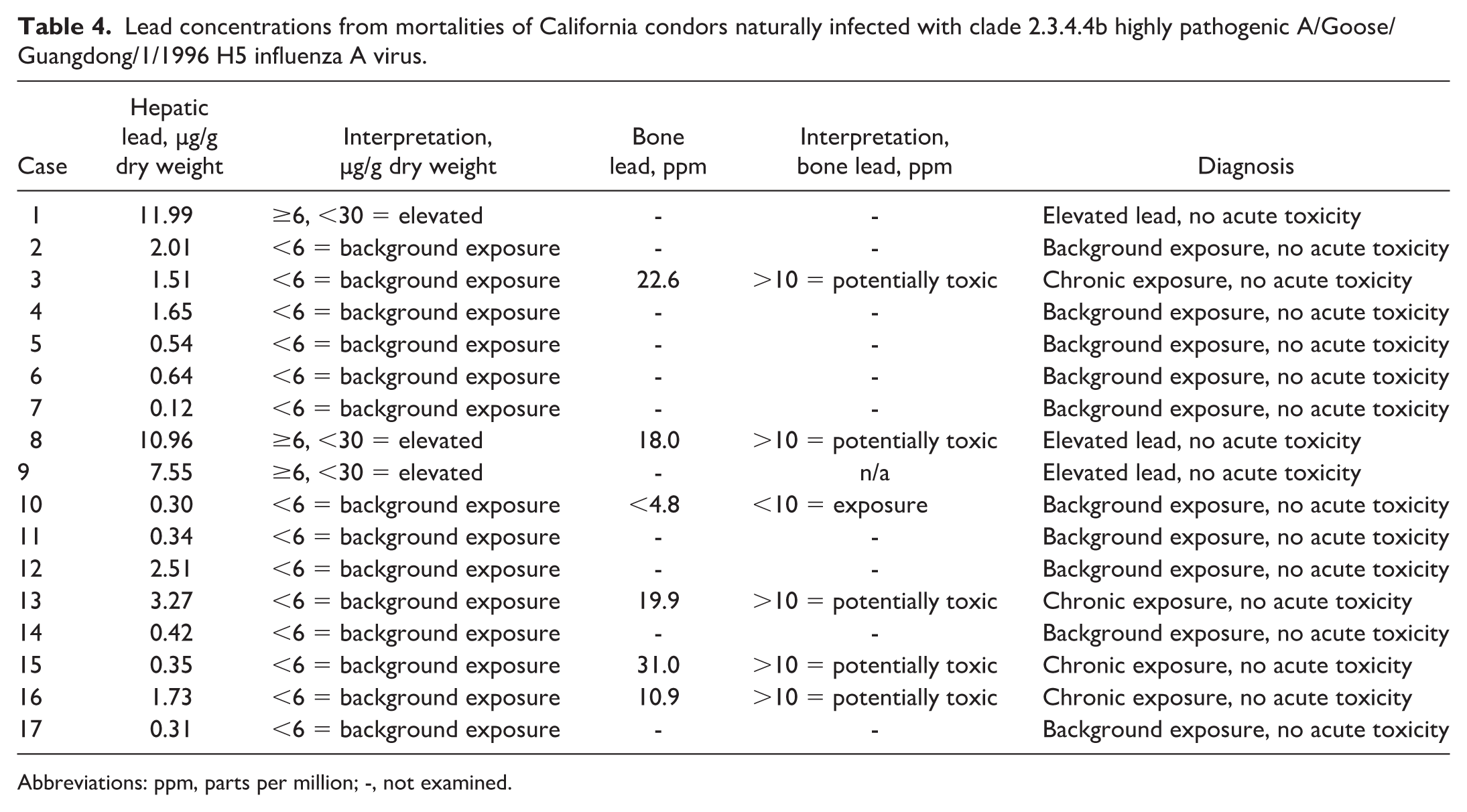
Abbreviations: ppm, parts per million; -, not examined.
Bone lead was analyzed in 6 of the condors. In all but 1, concentrations were above 10 ppm dw, indicative of potential poisoning. The highest bone lead concentration was 31 ppm dw (case 15). When assessed in conjunction with the liver lead, findings were consistent with exposure, but not acute toxicosis.
Liver copper concentrations in 13 of 17 condors were elevated above the bald eagle reference range of 4.0–50.0 µg/g dw and ranged from 10.68 to 518.84 µg/g dw. Other surveyed tissue minerals were not elevated.
Hair Identification
Microscopic analysis of ingested hairs indicated consumption of Cervidae (1/8), Bovidae (5/8), Lagomorpha (1/8), and Rodentia (1/8). Two of these condors had ingested hairs from 2 different taxa. Hairs in 2 condors were too degraded to determine their origin.
Discussion
This study describes the history, signalment, necropsy findings, IAV antigen distribution, and heavy metal concentrations in California condor mortalities caused by HPAIV infection. Overall, gross and histologic findings in the California condor are similar to other reports in raptors with the spleen and adrenal glands being most severely and consistently affected. Encephalitis was also common, though not as severe. Lesions in these 3 organs showed strong colocalization with IAV antigen, whereas other organs (heart, liver, pancreas, and gonad) had labeling most often in epithelial and mononuclear cells, but not necessarily in areas of inflammation. Splenitis was a striking feature of this outbreak and was often grossly appreciable as enlargement, firmness, and mottling. Pulmonary edema and congestion were reported in raptors affected by HPAI in 2014–2015, but viral pneumonia has not been a feature of this current outbreak and was not present in these condors.13,20,23,24,37,49 Lung lesions, when they did occur, were caused by secondary fungal infection in 1 condor and a secondary fungal and concurrent bacterial infection in another. Both affected condors had been hospitalized during their illness, allowing for a slightly protracted disease course and the opportunity for secondary infection.
Viral lesions manifested largely as acute necrosis. Inflammatory cells were less abundant and primarily mononuclear. Viral antigen was detected in multiple organ systems, most frequently and strongest in neurons. Labeling in the brain was overall much more pronounced than the accompanying histologic lesions, and IHC may be a more reliable way to identify infection in this organ. In other tissues, labeling was often present in epithelial cells and sometimes in circulating mononuclear cells. Although background labeling and severe necrosis hindered immunohistochemical evaluation in some birds, generally, labeling was identifiable and was often colocalized with areas of necrosis and inflammation. Immunolabeling of circulating mononuclear cells identified the virus in organs with no histologic lesions, such as the lungs.
In the reproductive tracts, cells in the testes, ovary, and oviduct were labeled for avian influenza in all 3 examined condors (2 males and 1 female). Histologic lesions were only present in the testicular tissues of 1 of 7 and in the oviduct of 1 of 8 condors; however, cellular sloughing and debris accumulation in seminiferous tubules could be interpreted as pathologic, particularly considering positive IHC labeling for avian influenza. Histologic lesions and IHC labeling for avian influenza in the reproductive tracts of wild birds during the current outbreak are poorly described but have been observed in both wild raptors and shorebirds.20,23 Consistently positive labeling of epithelial cells in both male and female California condor gonadal tissues raises the possibility of future reproductive impacts.
Although the timespan for this outbreak was relatively brief at 23 days, it appeared as if most of the condors were infected early on, with histologic lesions in birds dying later appearing more chronic. This was evidenced by cerebellar glial nodules and a shift in the splenic and adrenal lesions from widespread hemorrhage and necrosis to accumulation of lymphocytes and plasma cells in mortalities occurring in the last week of the outbreak. Myocardial lesions were also more prominent later in the outbreak. In contrast, black vulture (Coragyps atratus) HPAIV outbreaks lasted up to 270 days in Florida, and lesion patterns consistently were acute throughout numerous mortality events. 13 It could be that the rapid field and veterinary team response to the outbreak prevented new infections; however, the low number of individuals in our study and unknown infection time points across individuals make it difficult to draw any firm conclusions regarding the progression of lesions and disease duration.
Acute trauma was relatively common and sometimes of a severity to have significant impacts on health and survival. The most severe injuries were consistent with blunt force trauma, indicating falls from a height, either from a perch or during flight. In at least 1 case, traumatic injury included a fatal skull fracture. The presence of concurrent bone and internal organ fractures may have increased and likely accelerated mortalities. Trauma reflected the rapidly debilitating nature of the infection, with encephalitis as a probable contributor. Blunt force trauma is not otherwise a frequent cause of California condor mortality. For comparison, in 24 months following this outbreak, of the condors examined at the National Fish and Wildlife Service Forensics Laboratory, only 8% of the mortalities (3 deaths) were caused by blunt force trauma. All of those were young, otherwise healthy birds with contributing circumstances such as weather events or vehicle collision.
California condors have large ranges, so all birds in the Arizona population (about 100 birds) were considered at risk. Condors are intensively monitored both visually at known areas of congregation and through tracking using global positioning systems and very high frequency antennae. Mortality signals are emitted from the trackers after a designated period with no detected movement. This means that biologists generally can quickly identify and retrieve ill or dead condors, and all members of the population can be accounted for. Though not discussed in detail here, it should be noted that an additional 4 condors from the Arizona population were hospitalized within the same timeframe and were diagnosed via rRT-PCR of cloacal and choanal swabs with H5N1 clade 2.3.4.4b, genotype A3. These birds recovered and were released. Details can be found in Lamb et al. 22
In all mortalities, viruses were identified as H5N1 clade 2.3.4.4b genotype A3 with high identity indicating a common source exposure. Condors may have become infected through ingestion of infected carcasses, as has been suspected in outbreaks in other avian scavenger populations and has been experimentally induced in falcons.5,8,13,19,33 In the infected condors, the frequency of IAV viral antigenic labeling in both the upper and lower gastrointestinal tract epithelium, both colocalized and in the absence of histologic lesions, suggests the gastrointestinal tract as a route of infection in condors; however, studies linking histopathologic findings to a particular infection route in raptors are lacking.
There was no clear evidence of an HPAIV respiratory component in these condors. There were no lesions of viral pneumonia, and the only antigenic labeling evident in lungs was in circulating mononuclear cells. Given the affinity of the virus for epithelium in other locations, it seems less likely that inhalation is the source of infection in the condors. That said, nasal mucosa was not sampled in any of the birds, and lesions and/or antigenic labeling in the upper respiratory tract cannot be ruled out. An intranasal route of spread through the avian olfactory system in other species has been speculated. 49
If the route of infection is through ingestion, what are the possible sources? Condors are obligate scavengers that primarily feed on mammalian carcasses.12,14,38 In these cases, hairs in the ventriculus contents showed that mammals were among the most commonly consumed food items. Although the timing of this outbreak predates the earliest reported HPAI detection in any mammal in Arizona by 8 months, undetected or undocumented infections in mammals at this time cannot be ruled out. 43
Bovidae hairs were amongst the identified ingesta. This is not unexpected as proffered carcasses at the condor release sites are often cattle and because condors can fly long distances to acquire food outside of their home range. At the time, provided carcasses were not monitored for avian influenza because the outbreak predated identification of HPAIV in domestic cattle by over a year.10,15,18,42 There is no strong evidence that cattle were the source of infection, though it remains a possibility. Cats are also susceptible to infection, but there was no evidence of consumption of Felidae in the examined condors.
Rare feathers in the ventriculus had features consistent with raptor feathers and could not be distinguished from self (ie, condor) feathers. Although ingestion of infected avian species is a possibility, as with cattle, there is no strong evidence to suggest this occurred. None of the condor carcasses had been scavenged, and there were no other avian tissues in the ingesta. In the authors’ experience, examination of the upper gastrointestinal tract contents in deceased California condors does not indicate that avian carcasses are a regular part of their diet in the wild. 47 It is unlikely that any retained ingested feathers would have degraded substantially during the brief course of illness, but ingesta may have been lost through regurgitation.
Hairs and feathers were examined as the only available trace evidence; however, there are limitations to this analysis. Fibers may persist in the ventriculus, and there is no way to be sure how recently they were ingested. Also, hairs are selected for microscopic examination based on quality, with an emphasis on hairs that are intact and lack heavy bile staining. Consequently, if there are multiple families within the ingesta, some could be missed. Degradation of hair either prior to or after consumption can prevent identification. It is entirely possible that consumed sources of HPAIV were never identified, and the lack of hair should not be taken as proof that a particular carcass was not part of the diet.
If no definitive link can be made to infection through carcass consumption, indirect exposure via contamination with fluids from other infected avian scavengers should be considered. Arizona hosts birds migrating along both the Pacific and Central flyways, offering increased opportunities for the influx of disease in the spring. Migration in Arizona begins in March (the time of the outbreak), peaking at the end of April and May (https://dashboard.birdcast.info/region/US-AZ?night=2023-03-18 accessed 20 March 2025). In the days leading up to the outbreak, migration numbers across Arizona were low to medium as spring migration season accelerated (https://birdcast.info/migration-tools/live-migration-maps/ accessed 20 March 2025).
Condors are social feeders, often congregating around carcasses with conspecifics and other species, including turkey vultures (Cathartes aura) and corvids.14,38 Close association with infected birds provides a possible route of exposure. Four of the adult condors (2 males and 2 females) had superficial skin injuries of the head and neck consistent with mutual grooming, and at least 3 of these individuals were found at the same coordinates. Findings such as these, as well as clustering of at least 16 of the mortalities within only 6 geographic areas, support close association as a means of viral transmission. The outbreak overlapped with egg laying season (January to April), providing more opportunities for conspecific spread and possibly an additional stressor. It is possible that after initial exposure through feeding on an infected carcass or association with an infected bird, interactions of the condors with each other facilitated the spread of the disease.
Other than 2 cases of West Nile virus, communicable disease as a direct cause of morbidity and mortality in California condors has not previously been identified.35,47 At least 1 study has shown seropositivity of healthy California condors for several avian diseases including avian adenovirus, infectious bronchitis virus, Mycoplasma gallisepticum, and avian paramyxovirus-2. 41 These findings indicate that there is at least exposure to diseases that may be carried by other avian scavengers.
Unfortunately, the presence of HPAIV in other species in Arizona at the time of the outbreak could not be corroborated. Though HPAIV had already been detected in Arizona, the most recent report in a poultry flock was in December 2022 and in wild birds (Canada geese) was in January 2023. In mammals, the first report in Arizona was in an Abert’s squirrel in August 2023, 4 months after the last condor mortality (https://www.aphis.usda.gov).
Given the persistence of lead toxicity as a cause of illness and mortality in the California condor, lead concentrations were examined to assess whether lead toxicity could be associated with influenza mortalities. Liver lead concentrations reflect relatively recent exposures to lead whereas deposition of lead into bone occurs over the life of the bird.3,17,36 Liver lead concentrations were not at toxic levels in any of the condors with HPAIV, and there were no necropsy findings to suggest clinical lead toxicity. Signs of lead toxicity on necropsy may include emaciation, bile stasis, upper gastrointestinal tract erosions, visceral pallor, and/or ingested lead.25,48 Based on the low levels of lead present at the time of death and the lack of necropsy findings of lead toxicity, acute lead toxicosis did not predispose to mortality in this outbreak.
Low levels of lead in the liver and bone are of uncertain significance. Of the tested tissues, almost all bone and 3 liver samples had levels indicating lead exposure. The remaining samples contained lead concentrations considered negligible (“background exposure”). Bone tissues can fix sublethal lead concentrations, making bone lead measurements a useful tool to assess cumulative, chronic exposure, but do not necessarily reflect lead levels at the time of sampling. In previous studies, bone lead in condors determined at necropsy typically exceeded 15 µg/g, which was similar to the bone lead concentrations in this study. 40 Specific bone lead concentrations that may be associated with subclinical signs, including effects on reproductive performance, bone turnover, immune response, or nervous system, are not established. Any potential influence of low-level long-term accumulation of lead on HPAIV lesion severity and mortality was not statistically assessed in this study due to the low number of available bone samples.
As of the date of this publication, no additional known avian influenza mortalities have occurred in free-ranging California condors. While the effect of the rapid and continued response from the field and medical teams cannot be known for sure, it seems likely that removal of infected condors from the environment and alteration of field procedures helped to prevent spread and possibly contributed to the short-lived nature of the outbreak. At the start of the outbreak, immediate steps were taken to protect the remaining condors, including the removal of proffered food and water at condor release sites and increased biosecurity to the highest level for handling and intake of birds. Field monitoring was increased to rapidly identify and recover any birds suspected of being infected. This consisted of field observations for lethargy or other abnormal behavior and analysis of movement data to analyze overlap as an exposure predictor. Facilities for condor holding, release, and medical care were renovated to increase biosecurity and facilitate cleaning and sterilization (T Hauck, personal communication).
To date, HPAIV infection in California condors remains a concern. The virus is still circulating, and treatment options are limited and typically unsuccessful in fulminant cases. Following the HPAIV outbreak, vaccine development and a vaccination program were initiated. 4 Vaccination is often considered an impractical strategy in free-ranging wildlife; however, in California condors, there is a successful precedent in the ongoing vaccination program for West Nile virus. 11 To evaluate the success of the vaccine, effectiveness of management practices, and possible long-term effects of resolved infections, continued monitoring and routine HPAIV testing of all mortalities are important. To date, all condors continue to be opportunistically swabbed for HPAIV surveillance, and mortalities are routinely tested for HPAIV. Vaccination and monitoring of serum antibodies to identify past exposure and vaccination response are ongoing.
In conclusion, California condors are susceptible to HPAIV H5N1 clade 2.3.4.4.b. Encephalitis is consistently present, as well as splenitis and adrenalitis. Splenitis was an especially severe lesion and was often grossly appreciable. Infection is suspected to be spread through association with infected birds, ingestion of infected carcasses, or a combination of the 2. Acute lead toxicity, the leading cause of mortality in this species, was not a factor in any of the condor HPAIV mortalities. Ongoing monitoring of individual and population health is critical as there are seasonal to continual detections of HPAIV in wild birds in the USA, and repeat outbreaks in this endangered species could have devastating impacts on California condor population recovery.
Supplemental Material
sj-pdf-1-vet-10.1177_03009858261457964 – Supplemental material for An outbreak of H5N1 clade 2.3.4.4b highly pathogenic avian influenza in California condors (Gymnogyps californianus)
Supplemental material, sj-pdf-1-vet-10.1177_03009858261457964 for An outbreak of H5N1 clade 2.3.4.4b highly pathogenic avian influenza in California condors (Gymnogyps californianus) by Rebecca Kagan, Tabitha C. Viner, Nicole M. Nemeth, John Buchweitz, Birgit Puschner and Rachel Sheffler in Veterinary Pathology
Footnotes
Acknowledgements
For their laboratory work and technical expertise, we thank the scientists and support staff at the Oregon State University Oregon Veterinary Diagnostic Laboratory, Histology Consultation Services, National Fish and Wildlife Forensics Laboratory, Dr Elizabeth Howerth and her team at the University of Georgia Histology Laboratory, the USDA National Veterinary Services Laboratory, and the Michigan State University Veterinary Diagnostic Laboratory. We also thank the U.S. Fish and Wildlife Service Office of Law Enforcement, the U.S. Fish and Wildlife Service California Condor Recovery Program, and Liberty Wildlife for their case support, animal care, and continued dedication to California condor conservation.
Supplemental material for this article is available online.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Disclaimer
All findings and conclusions in this article are those of the authors and do not necessarily represent the views of the US Fish and Wildlife Service. The mention of trade names does not imply US government endorsement of commercial products.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.