
Abstract
The ingestion of the pollen beetle Astylus atromaculatus causes gastroenteric disease in ruminants and possibly in other species. No toxic principle has been identified in these insects. Two studies were conducted to evaluate the effects of oral administration of A. atromaculatus in guinea pigs using fresh insects (n = 3) and insects dried for different periods (2, 30, 60, 120, or 240 days) (n = 5). All animals became sick and died spontaneously between 10 and 96 hours after inoculation. Clinical signs included depression; reluctance to move, eat, and drink; kyphosis; and increased frequency of vocalization, with no differences observed between the 2 experiments. Gross and microscopic findings consisted of diffuse ulcerative, fibrinonecrotizing, and hemorrhagic enteritis. No differences were observed in the type or severity of lesions between the 2 experiments or in animals inoculated with beetles dried for different durations. No clinical signs or lesions were observed in guinea pigs from the control group (n = 2). This study adds to the knowledge of A. atromaculatus-associated fatal gastroenteric disease in non-ruminants and highlights the persistent harmful effects of dried pollen beetles.
The pollen beetle Astylus atromaculatus, also known as the spotted maize beetle, is a coleopteran of the family Melyridae that is native to South America and is commonly found in crops.1,8 Its presence was reported in South Africa after introduction from South America. 11 A. atromaculatus feeds on pollen and is a regular floral visitor, acting as an efficient pollinator. The insect can be found on flowers of milo, corn, sunflower, pigweed, and soybean, among others.2,3,9,14,15 These beetles are black and yellow and have characteristic black spots in the prothorax and elytra.1,7 The ingestion of adult A. atromaculatus results in highly lethal gastroenteric disease in sheep and cattle.4,13 The causative role of this insect in the disease has been demonstrated after experimental oral administration in both species.5,7,11
Spontaneous outbreaks of fatal gastrointestinal disease due to A. atromaculatus ingestion occurred in ruminants during the summer of 2022–2023 in Argentina and Uruguay, after a severe drought that affected blooming crops and resulted in an overpopulation of the beetle in forages used to feed ruminants.13,14 During that period, A. atromaculatus-associated disease was also suspected in horses, dogs, and chickens, although a causal association was not demonstrated. While a toxic agent is suspected, no toxic principle has been identified in these insects to date, and the precise cause and mechanism of disease is poorly understood. 7 In guinea pigs, there has been only one experimental reproduction, more than 50 years ago, of gastrointestinal disease due to A. atromaculatus ingestion. 11 However, that study provided limited clinical and pathological characterization and evaluated the effect using fresh blended insects, without assessing their effects after prolonged drying.
Although outbreaks have been associated with ingestion of fresh beetles in pasture,4,13 the potential persistence of toxicity after drying remains unknown. This is particularly relevant under field conditions, as heavily infested crops may be harvested and conserved as hay, potentially exposing animals to beetles long after collection. Therefore, this study aimed to reproduce the disease in guinea pigs as an experimental monogastric model and to evaluate persistence of toxic effects of A. atromaculatus after drying, detailing clinical signs and pathological findings and providing insight into potential risks associated with contaminated hay and other stored feeds.
Two experiments were conducted. Both experiments were performed following the guidelines of the Institutional Committee for the Care and Use of Experimental Animals of the National Institute of Agricultural Technology, Argentina (protocol 252/2023). Pollen beetles were collected from heavily infested alfalfa paddocks where gastrointestinal disease in ruminants had been previously recorded. 4 For inoculum preparation, recently killed, fresh beetles (fresh inoculum; experiment 1) or beetles that had been dried, before grinding, and kept at 30°C in an oven during 2, 30, 60, 120, or 240 days (dry inoculum; experiment 2) were chopped and blended in an aqueous suspension of 5 mL distilled water before oral administration by syringe, slowly dosing small drops into the guinea pigs mouth while maintaining the animal in dorsal recumbency. In experiment 1, 3 adult guinea pigs were each orally dosed with fresh inoculum prepared with 4 g/kg of A. atromaculatus (Table 1). In experiment 2, 5 adult guinea pigs were each orally dosed with dry inoculum prepared with 4 g/kg of A. atromaculatus dried for different periods of time (Table 1). Dosage was calculated on a wet basis prior to drying. Two adult guinea pigs (control group) were administered 5 mL of sterile water.
Signalment, drying period, inoculation–death interval, and form of death in guinea pigs experimentally inoculated (4 g/kg body weight) with fresh (n = 3, experiment 1) or dried (n = 5, experiment 2) adult Astylus atromaculatus.
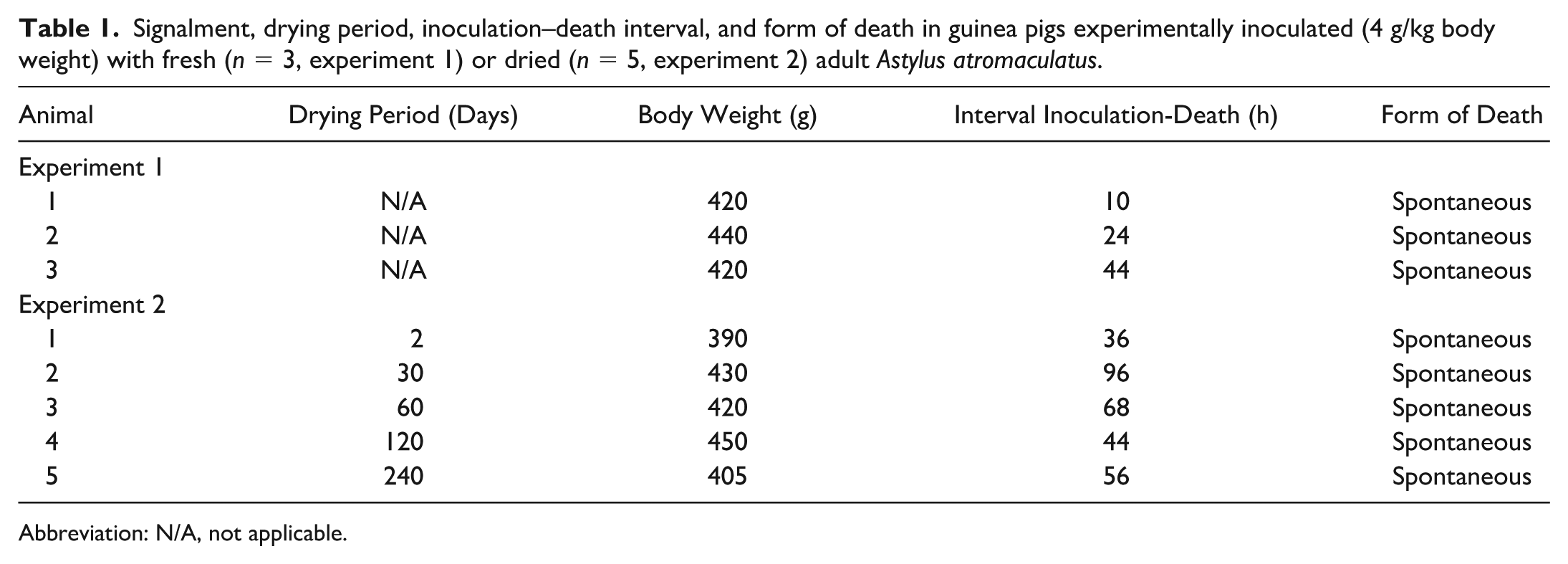
Abbreviation: N/A, not applicable.
All guinea pigs were kept in individual cages and were clinically evaluated before and every 4 hours after inoculation until spontaneous death or euthanasia. Clinical signs were recorded, including decreased motor activity, abnormal postures, decreased intake of food and water, and decreased grooming. 16 Necropsies of all animals were performed after death, and samples of stomach, small and large intestine, kidney, liver, heart, brain, lung, spleen, esophagus, and urinary bladder were collected and fixed by immersion in 10% buffered formalin for 72 hours. Tissues were processed using standard techniques to prepare 4-μm-thick hematoxylin and eosin-stained sections.
All 8 animals dosed with A. atromaculatus from both experiments showed similar clinical signs including depression; reluctance to move, eat, and drink; kyphosis; and increased frequency of vocalization. These clinical signs started between 4 and 8 hours and 24 and 48 hours after inoculation in experiments 1 and 2, respectively. These 8 guinea pigs died spontaneously 10 to 96 hours after inoculation (Table 1). Neither of the control guinea pigs show clinical signs and were euthanized at 72 hours after inoculation.
On necropsy, all animals inoculated with A. atromaculatus in both experiments showed diffuse transmural hemorrhage of the mucosa of the duodenum, jejunum, and ileum that was visible from the serosal side (Fig. 1a). Liquid and hemorrhagic small intestinal contents with fibrin strands attached to the mucosa were observed in 5 of these animals, including both experiments (Fig. 1b). Frequently (6/8) in the small intestine, there were multifocal to coalescent ulcers, occasionally covered with a fibrinous pseudomembrane, while nonulcerated areas were diffusely hyperemic, hemorrhagic, and thin. There were no differences in the type or severity of gross lesions between animals of both experiments or in animals from experiment 2 inoculated with beetles dried for different periods of time. No gross lesions were observed in other organs, including the esophagus, the stomach, and the large intestines. No gross abnormalities were observed in the 2 animals of the control group.
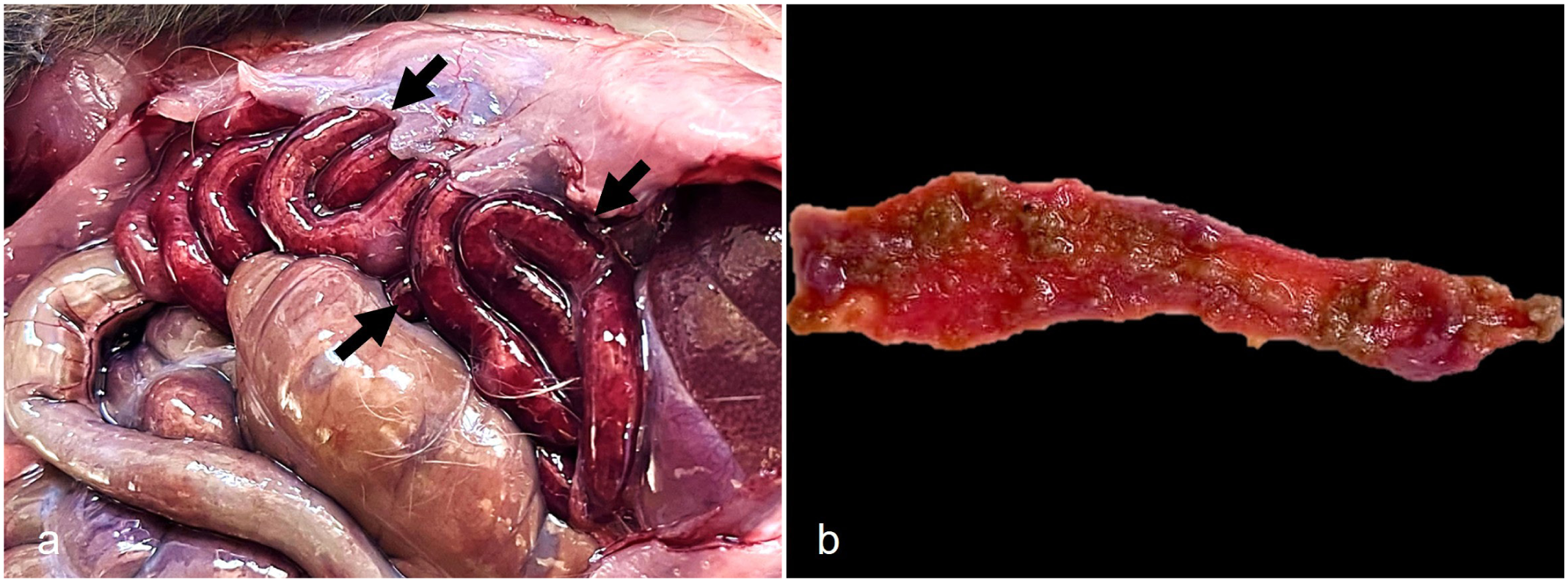
Gross findings in guinea pigs experimentally inoculated with Astylus atromaculatus. (a) Diffuse necrohemorrhagic enteritis affecting the duodenum, jejunum, and ileum (arrows), visible from the serosal side. (b) Jejunum. Diffuse necrohemorrhagic enteritis with multifocal attached fibrin strands.
Microscopically, in all A. atromaculatus-inoculated animals from both experiments, the lesions were primarily in the small intestine, with the jejunum being more severely affected, followed by the ileum and then the duodenum. The mucosa of these organs was lined by abundant fibrinous exudate mixed with necrotic debris, erythrocytes, and bacterial colonies. There was severe and diffuse villous atrophy and fusion, with marked congestion and apical hemorrhage (Fig. 2a, b). There were multifocal to coalescing areas of necrosis and ulceration, notably in the superficial mucosa, that occasionally extended deeply involving the whole thickness of this layer. These areas were covered by a pseudomembrane composed of cellular debris, erythrocytes, mixed bacterial colonies, sloughed epithelial cells, and fibrin (Fig. 2a, b). Deep in the mucosa, there were abundant dilated crypts with necrotic or attenuated epithelium, and karyorrhectic and pyknotic debris intermingled with viable and degenerate neutrophils (Fig. 2c). There were extensive areas of hemorrhage and infiltration of neutrophils and fewer lymphocytes in the submucosa. In 4 animals, 2 from each experiment, there was crypt herniation toward the Peyer patches in the ileum. The herniated crypts were dilated and contained necrotic debris and mucus. The large intestine of 2 guinea pigs from experiment 1 had erosive colitis/typhlitis with diffuse hemorrhage and lymphocytes infiltrating the mucosa and submucosa. In 3 animals, the liver had mild microvesicular vacuolation of centrilobular hepatocytes. Multifocal lymphocytolysis in the white pulp of the spleen was observed in 4 animals from both experiments (Fig. 2d). Thickening of the alveolar septa with hemorrhage and mixed infiltrates of monocytes and neutrophils was observed in the lungs of 2 animals from experiment 1. No microscopic abnormalities were observed in any of the other tissues examined. There were no differences in the type or severity of microscopic lesions between animals from experiment 2 inoculated with beetles dried for different periods of time. No microscopic abnormalities were observed in the 2 animals of the control group.

Microscopic findings in guinea pigs experimentally inoculated with Astylus atromaculatus. Hematoxylin and eosin. (a, b) Jejunum. Severe, diffuse fibrinonecrotizing enteritis with villous atrophy and fusion and marked mucosal hemorrhage. Abundant fibrinous exudate forming pseudomembranes is observed on the luminal surface. Inset: Higher magnification of the luminal surface with fibrinous exudate composed of sloughed epithelial cells, fibrin, necrotic debris, and erythrocytes. (c) Jejunum. Dilated crypts with necrotic or attenuated epithelium, and karyorrhectic and pyknotic debris intermingled with viable and degenerate neutrophils. Extensive hemorrhage is also observed. (d) Spleen. There is nuclear fragmentation (necrosis/lymphocytolysis) of lymphocytes in the white pulp.
The results of this study add knowledge by demonstrating that the toxic effects of A. atromaculatus persist after prolonged drying, which has direct implications for disease risk under field conditions. Inoculation of fresh or dried beetles resulted in lethal enteric disease. In experiment 2, the lethal effect of the beetles persisted in insects that had been dry for at least 8 months. This is an important finding considering that outbreaks, mainly associated with alfalfa pastures with high infestation of pollen beetles, were then processed for hay production. As our results indicate that toxicity may persist in dried beetles. This suggests a potential risk associated with hay or other conserved forages produced from heavily infested fields, even months after harvesting. It has been previously reported that an aqueous A. atromaculatus suspension stored for 5 weeks at 4°C induced only mild clinical signs in sheep, followed by full recovery. 11 Temperature or water dilution storage may therefore play a role in reducing toxic potential, in contrast to our study, in which adverse effects persisted after beetles were dried and stored at 30°C for several months. However, to the best of our knowledge, no suspected cases of A. atromaculatus intoxication have occurred in ruminants that consumed alfalfa hay from areas known to have been previously infested with this beetle.
Previous reports of spontaneous disease in cattle and sheep consuming alfalfa and mixed native pastures heavily contaminated with A. atromaculatus and experimentally in calves and sheep confirmed that this beetle causes gastrointestinal disease in ruminants.4,5,7,13 However, suspected disease in monogastric animals (horse, dog, and chicken) have been anecdotally and poorly documented, and no post-mortem examinations or laboratory tests have been reported in those cases. Our results provide evidence that the consumption of A. atromaculatus can also cause enteric disease in monogastric animals.
This study was designed as a proof-of-concept investigation; consequently, statistical analysis was not performed because of the limited number of animals available. The clinical signs and lesions in these 2 experiments are similar to those described in spontaneous and experimental cases in ruminants.4,7,11,13 In the experiment performed in guinea pigs by Kellerman et al, 11 the interval between administration and the occurrence of clinical signs and death was similar (24 hours). However, in our study, some variation was observed, which could be due to individual resistance. Although this interval variation was observed between experiments and between animals from experiment 2, no differences in the type or severity of lesions were seen. A longer interval between oral administration and occurrence of clinical signs and death was registered in animals inoculated with dry beetles (experiment 2). This could be consequence of a reduction of toxicity associated with the drying process.
We demonstrated that beetles kept dry at 30°C for up to 8 months remain toxic. Based on our results, spontaneous cases of enteric disease by A. atromaculatus may occur in animals consuming hay or other dry forage foodstuff contaminated with these beetles.7,11 This is similar to what occurs with blister beetles of the family Meloidae, whose toxic principle, cantharidin, persists in dead dried beetles during hay storage, resulting in spontaneous disease in horses that consume that hay.6,10,12 No cantharidin has been found in A. atromaculatus used to experimentally reproduce the disease.7,11
In our study, lesions were primarily seen in the small intestine, which is similar to what was previously described by Kellerman et al. 11 Forestomach and abomasal lesions were described in ruminants;4,7,13 however, no stomach lesions were seen in our guinea pigs or in those described by Kellerman et al. 11 This could be related to the retention of beetles in the ruminant forestomaches for longer periods, thereby increasing contact time and the likelihood of lesion development. Other possibilities include metabolic differences, whereby the eventual causative toxin may become more bioavailable during passage through the forestomaches and stomach, whose distinct environments could enhance its toxicity.
In contrast to the spontaneous cases in cattle described by Garcia et al, 4 we did not observe fibrinoid necrosis in the blood vessels of the intestinal submucosa or perivasculitis in the guinea pigs after oral administration of A. atromaculatus. In addition, we observed lymphoid cell necrosis in the white pulp of the spleen, a finding not described by Kellerman et al 11 in the guinea pigs in their experiment and not observed in spontaneous and experimental ruminant cases.4,5,7,13 Kellerman et al 11 described fatty degeneration and disseminated foci of necrosis in the liver of orally dosed guinea pigs. Similar to the findings reported by Santos et al 5 in sheep following experimental oral administration of A. atromaculatus, our study revealed mild, microvesicular hepatocellular vacuolation, probably due to reluctance to eat after beetle inoculation and enteric lesion development.
The pathogenesis of the intoxication by A. atromaculatus has not been determined, although a toxic principle is suspected. No intralesional chitinous insect fragments were found embedded in the mucosa of the intestine in any of the guinea pigs in this study, as has been described in some experimentally affected calves. 7 This suggests that the intestinal lesions are not due to mechanical trauma produced by insect parts.
Our findings highlight that A. atromaculatus intoxication should be considered a potential risk in stored forages. This expands the current understanding of the epidemiology of the disease and underscores the need for further studies to identify the toxic principle and its stability under different environmental conditions.
Footnotes
Acknowledgements
We thank Valeria Scioli for histopathological processing. FG and CM acknowledge support from the Uruguayan “Agencia Nacional de Investigación e Innovación” (ANII) through the “Sistema Nacional de Investigadores” (SNI) program.
Author Contributions
MFO, NC, JL, and JAG designed and performed the experiments; GJC and JP contributed to the experimental design; MFO, NC, and JL collected clinical data; MFO, JL, and JAG performed necropsies and histologic evaluation; FAU, FG, and CM helped with interpretation of the pathological findings and provided valuable insights based on their experience; FAU, GJC, FG, and CM contributed to conceptualization. The draft of the manuscript was written by MFO, JAG, and FAU. All authors reviewed, edited, and approved the final version.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by National Institute of Agricultural Technology (INTA) (RIST: I111) and “Asociación Cooperadora INTA Balcarce.”