
Abstract
The saccadic eye movements declining given the development of Parkinson’s disease (PD) still deserves thorough analysis. Recent studies confirmed that PD patients show poor saccadic control in visuomotor tasks. The purpose of this study was to investigate the dynamics of saccades parameters at the development of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) model of PD. The gradual decline of saccadic control was studied in two monkeys that executed the visuomotor task with low doses of MPTP being injected at a prolonged period of time. The experiment included investigating the horizontal, vertical, oblique visually guided saccades as well as the corrective saccades triggered by stimuli onset in various loci within a two-dimensional visual field in the Gap-Step-Overlap paradigm. Our study revealed that the execution of visually guided saccades with small amplitude and corrective saccades changed dramatically with MPTP-model progressing. These changes are also confirmed statistically at the presymptomatic stage of MPTP syndrome. Not only our study gives a robust report of PD dynamics development and saccadic control but the obtained data could also be helpful in developing methods for the early diagnosis of PD.
Introduction
Parkinson’s disease (PD) is a neurodegenerative disorder common for middle-aged and elderly people; it is characterized by the death of dopaminergic neurons in the substantia nigra pars compacta and results in profound functional changes that involve the whole fronto-thalamo-striatal circuit. PD manifests itself through severe motor impairments including disorders in eye movements (Baziyan, Chigaleichik, & Dmitriev, 1998; Kori et al., 1995; Schneider & Kovelowski, 1990; Shul’govskii, Latanov, & Petrov, 1994; Slovin et al., 1999). Consequently, studying eye movements during PD plays a pivotal role aimed at investigating the normal functioning of the human brain and its subsequent dysfunction.
Visually guided, or “reflexive,” saccades are most frequently considered to be a logical object for studying PD dysfunctions. In designing the outline of the experiment, the investigators often focus on one of easily manipulated experimental variables—the time between the fixation stimulus offset and the target stimulus onset. In this connection, three distinct experimental paradigms involving this variable could be outlined: the gap paradigm, in which the fixation stimulus is extinguished some time prior to the presentation of the target stimulus; the step paradigm, in which fixation offset and target onset are simultaneous; and the overlap paradigm, in which the fixation stimulus is presented continuously or extinguished some time afterwards. These frameworks are known to give rise to significant differences in saccadic reaction times (Kalesnykas & Hallett, 1987), and thus the effects of stimulus timing are the most studied (Chambers & Prescott, 2010; Chan, Armstrong, Pari, Riopelle, & Munoz, 2005; Mosimann et al., 2005; Ratmanova et al., 2006; Tereshchenko, Yudin, Kuznetsov, Latanov, & Shul’govskii, 2002).
The results presented in literature on the subject are rather contradictory despite the number of these studies and the varying methodology being applied in each case. Still prolonged saccadic latencies followed by the increased dispersion were reported in most studies (Armstrong, 1997, 2011; Nakamura et al., 1991; Ratmanova et al., 2006; Rottach, Riley, DiScenna, Zivotofsky, & Leigh, 1996; White, Saint-Cyr, Tomlinson, & Sharpe, 1983). Other studies showed no changes in saccadic latencies for PD patients (Jones & DeJong, 1971; MacAskill et al., 2012; Mosimann et al., 2005) and even the shorter ones (Briand, Hening, Poizner, & Sereno, 2001). Visually guided saccades were revealed to have smaller amplitude compared with the eccentricity of the target stimulus almost in all studies. In other words, they are characterized by an “undershoot” or hypometric saccades (Armstrong, 1997, 2011; Jones & DeJong, 1971; Nakamura et al., 1991; Rottach et al., 1996; White et al., 1983). Therefore, after the primary saccade patients execute a smaller secondary corrective saccade (Jones & DeJong, 1971; White et al., 1983) and even a few corrective saccades, this effect is also known as multisaccadic approaching of the visual target (Armstrong, 2011; Ratmanova et al., 2006).
Most data showed a decreased peak saccadic velocity in PD patients (Armstrong, 1997, 2011; Nakamura et al., 1991; White et al., 1983) except for one earlier study describing a weak increase in peak velocity which was not statistically confirmed (Jones & DeJong, 1971).
A comprehensive review of more than 40 different studies (Chambers & Prescott, 2010) argued that PD leads to prolonged latencies of reflexive saccades and summarized that this parameter accounts for varying experimental conditions.
The most relevant model to study the pathophysiology of PD was developed using 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) intoxication in monkeys. This model can reproduce not only the destruction of the dopamine system but also mirror the entire complex of PD symptoms (Brooks, Fuchs, & Finocchio, 1986; Benazzouz, Gross, Dupont, & Bioulac, 1992; Porras, Li, & Bezard, 2012; Schneider & Kovelowski, 1990).
Experiments with the MPTP model also demonstrated increased latencies of visually guided saccades and decreased amplitude and accuracy for these saccades as well as a notable decline in their peak velocity (Brooks et al., 1986; Kori et al., 1995; Latanov, Tereshchenko, Yudin, & Shul’govskii, 1998; Porras et al., 2012; Shcultz et al., 1989; Shul’govskii et al., 1994; Slovin et al., 1999; Tereshchenko et al., 2002).
Cognitive deficits were shown to precede motor deficits (Kori et al., 1995; Porras et al., 2012; Slovin et al., 1999) and to be independent from the motor deficits (Schneider & Kovelowski, 1990) in an MPTP model. The studies focused on memory-learned saccades, delayed saccades, antisaccades, and saccades with cued choice.
It should be noted that almost all previous experiments used a very narrow spatial range of target stimuli positions. Investigating the horizontal saccades prevailed and is represented in the majority of studies on the subject. In this case, target stimuli were positioned along the horizontal meridian at different distances from the central fixation point (FP). The number of targets was also small: 10 (Brooks et al., 1986; Nakamura et al., 1991; White et al., 1983), 8 (MacAskill et al., 2012; Mosimann et al., 2005), and even 2—one to the left and one to the right of FP (Briand et al., 2001; Chan et al., 2005; Jones & DeJong, 1971; Kingstone et al., 2002). In some experiments, the stimuli were positioned along the circle trajectory at equal angles and at a fixed distance from FP (Kori et al., 1995; Rottach et al., 1996).
The abovementioned stimuli set-up resulted in a simplified and predetermined patterns, guiding the execution of saccades. That differs significantly from oculomotor functioning in a natural environment where target stimuli may be spread along the whole visual space. Therefore, the influence of the spatial factor taken separately has not been tested thoroughly in the process of executing visually guided saccades in the MPTP model and in PD.
Thus, the aim of this study was to determine the contributions of PD development to saccadic eye movements studied in progress and their increasing manifestation through the MPTP model especially at the initial stage of the syndrome development.
Methods
Two adult Macaca mulatta monkeys (two males, 8 and 10 kg) were trained to perform a visuomotor instrumental task. The monkeys were trained to shift the gaze from the FP to the peripheral stimulus within the Gap-Step-Overlap paradigm. The gap delay was 300 ms. The positions of peripheral target stimuli were determined according to a (7 × 5) rectangular grid centered to FP with the size of cells of 6.5° along horizontal and vertical axes. Thus, the spatial range of the used two-dimensional visual field enclosed ±19.5° and ±13.0° on horizontal and vertical axis. One of three possible time onsets of stimuli appearance and one of 34 possible target stimuli positions altogether were chosen in a pseudorandom order for each trial. Animals successfully executed at about 1,000 trials per day.
Eye movements were recorded by electromagnetic search-coil method developed by D. Robinson (1963) and modified by our group (Tereshchenko, Kuznetsov, Latanov, & Shul’govskii, 2000). A search-coil was implanted into the monkey’s eye bulb. The head of the animal was fixed, and eye movements were recorded at the frequency rate of 500 Hz within the spatial accuracy of 0.1° concerning both coordinates.
After the control data from the animals in normal conditions was collected, the MPTP administration began. MPTP (as hydrochloride salt) was administered intramuscular once every other day in a dose of 0.2 mg/kg. The injections were given shortly after a testing session, and animals were returned to their home cages.
The animals were observed and rated on a daily basis after the beginning of the administration of MPTP both in their home cages and in the primate chair against the behavioral scale which was introduced by J. Schneyder and C. Kovelowski (1990). This scale included 10 indices of motility and reactivity, varying from 0 (normal condition) to 2 points (severe MPTP Parkinsonian-like disorder). Therefore, ranking against this scale could fluctuate from 0 point for animals in normal condition up to 20 points for severe MPTP-treated animals.
Care and treatment of monkeys were in accordance with the Directive 2010/63/EU of the European Parliament and of the Council.
Results
Figure 1 shows a typical record of eye movement trajectory at a single trial of a visual task.
Gaze trajectory in execution of visual spatial instrumental trial.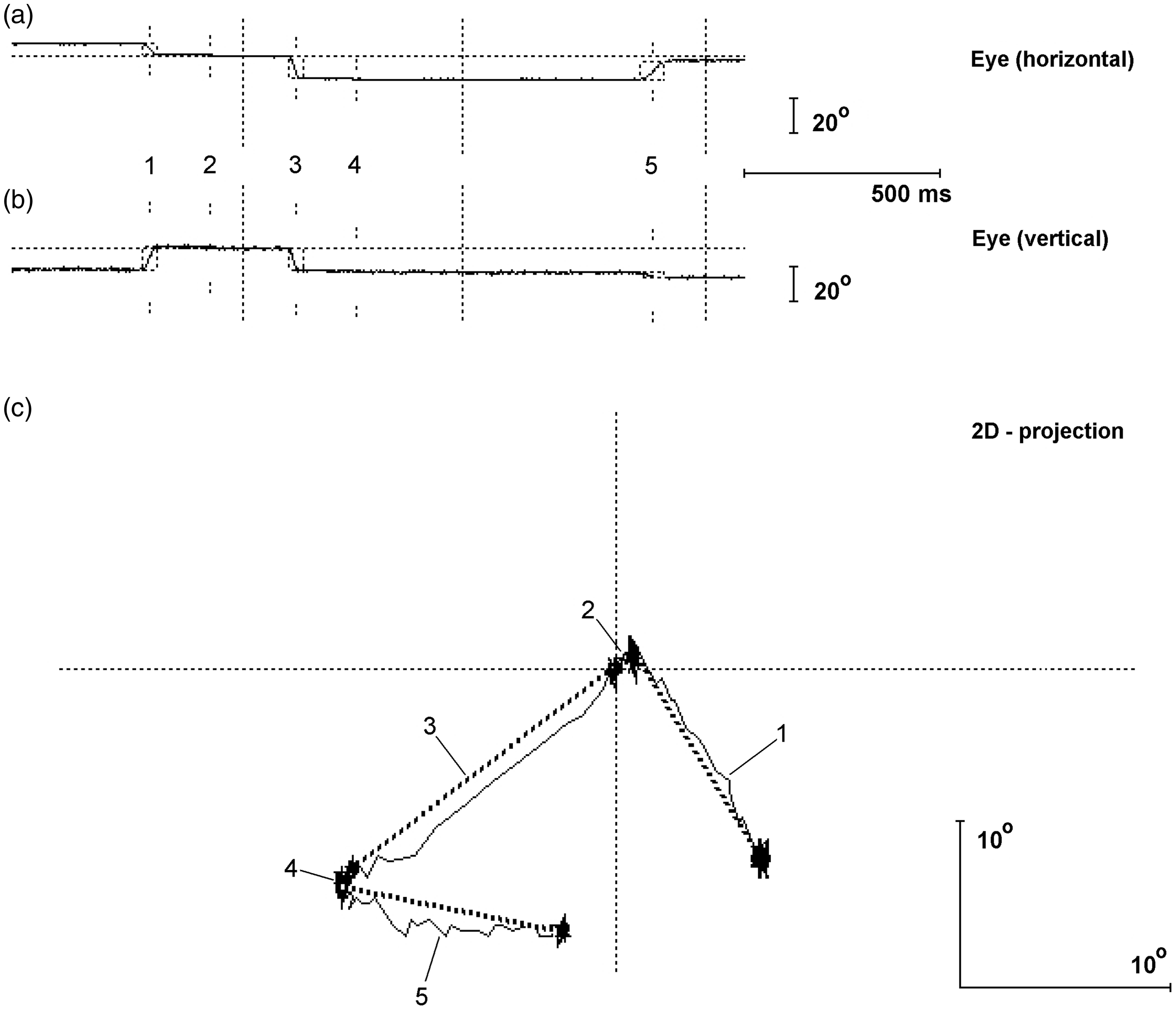
Following the first injections of MPTP, no behavioral alterations were observed. This stage of the syndrome development characterized by absence of any visual manifestation may be referred to as a presymptomatic stage of the MPTP syndrome.
After the presymptomatic stage of the syndrome developing, the monkeys’ condition started to decline conspicuously at the fourth (in relation to one monkey) and fifth injection of MPTP (in relation to the other monkey; the overall doze of 0.8–1.0 mg/kg). In addition to the overall decrease in activity, reduced appetite, the progressing pose disorder, separate cases of posturing were observed. The changes constituted 2 to 3 points against the behavioral scale, and since then the monkeys’ condition was going worse consistently. The monkeys tended to refuse executing the visual motor task and after the eighth injection of MPTP (the dose of 1.6 mg/kg), they stopped performing the tasks completely. Along the activity decline, some cases of disequilibrium were reported. The dynamics against the behavioral scale was evaluated as 5 to 6 points. Further on the tremor was visualized and the complex of parkinsonian syndromes manifested themselves to the full extent starting from values at the rate of 7 to 8. The behavioral changes in both monkeys were developing almost simultaneously: the symptoms came out with a difference of no more than 1 day of injections and the evaluation of the monkeys’ condition against the behavioral scale differed no more than by two points. Therefore, the parameters of eye movements were further on averaged in relation to both monkeys.
Latencies of visually guided saccades to the peripheral stimulus increased with the syndrome progressing. After the fourth injection (the dose of 0.8 mg/kg), the latencies increased by 10 to 20%%. The presaccadic intervals of corrective saccades increased significantly at the presymptomatic state of MPTP syndrome (Figure 2).
Dependence of visually guided saccades latencies and presaccadic intervals (ms, left) of corrective saccades at the MPTP-syndrome development.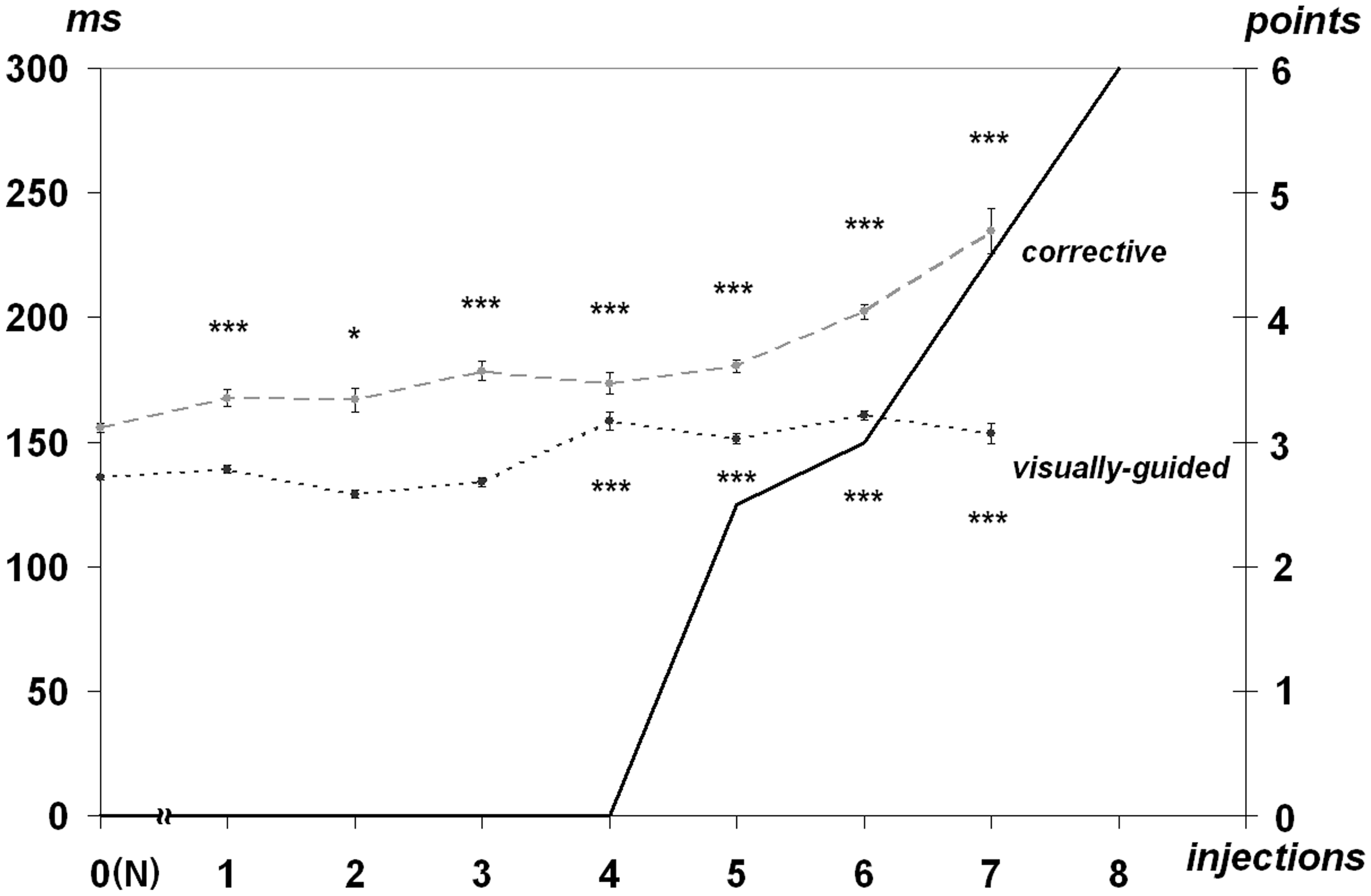
Following the dose of 1.0 mg/kg, the presaccadic intervals went up by 30 to 50%%. Latencies change, and the range of the target-oriented saccades exhibited no direct correlation. Latencies of low amplitude saccades increased by 20 to 44%% as compared with the normal rates at the presymptomatic state of syndrome development while latencies of average amplitude saccades increased at a lesser extent—by 5 to 17%%. However, latencies of greater amplitude saccades almost did not diverge from the norm even in case of a more pronounced demonstration of the MPTP syndrome (Figure 3).
Dependence of visually guided saccades latencies (ms, left) of three amplitudes—6.5°, 13°, and 19.5° at the MPTP-syndrome development. All references are the same as used for Figure 2.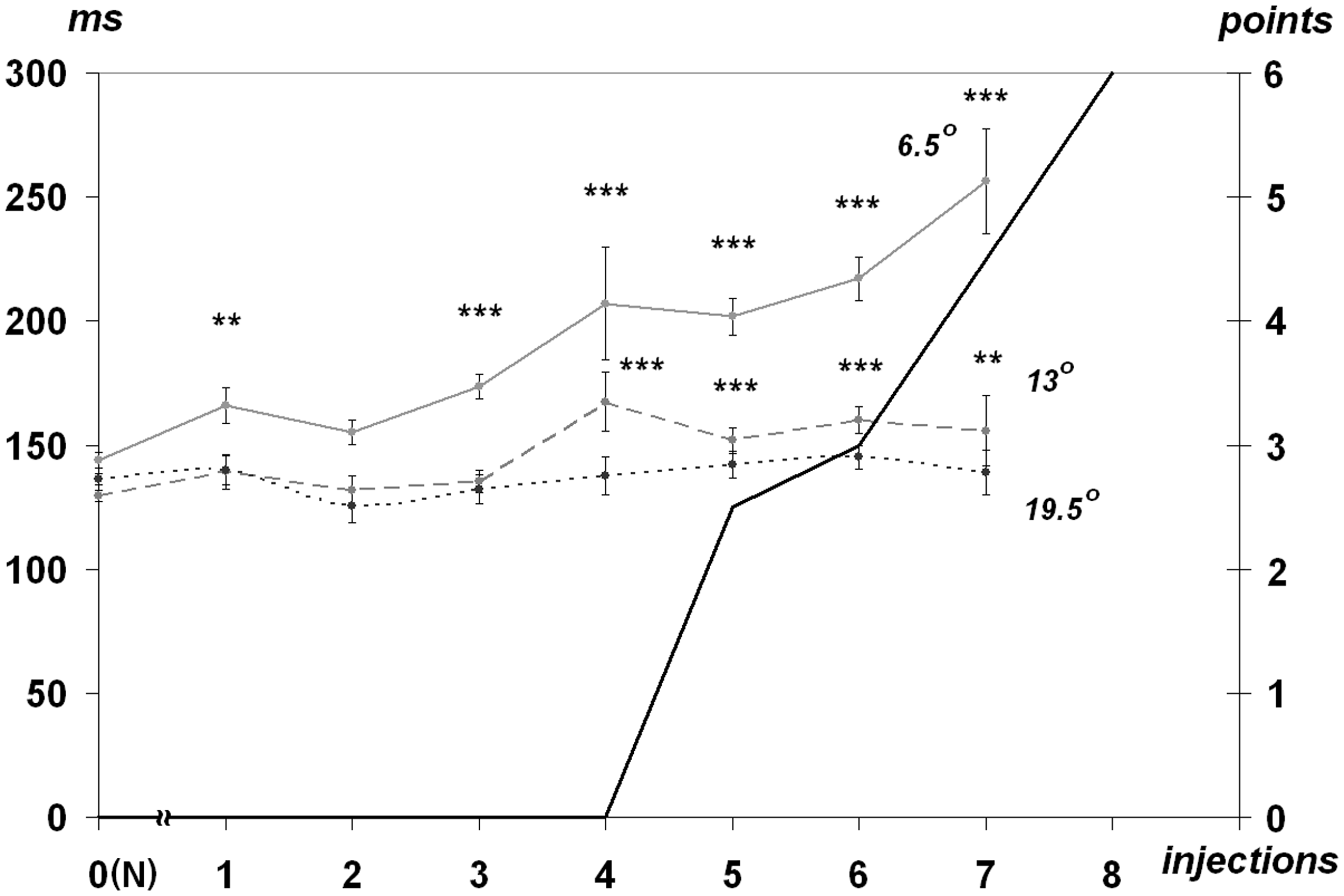
Visually guided saccades were followed by the subsequent corrective saccades within 55 to 60%% cases range. This value decreased to 48% only after the seventh injection prior to the stage animals stopped executing the visually guided task (Figure 4).
Dependence of execution rate of corrective and multisaccadic trials after visually guided saccades (%%, left) at the MPTP-syndrome development. Numbers of trials with execution of visually guided saccades are taken as 100%. All references are the same as used for Figure 2.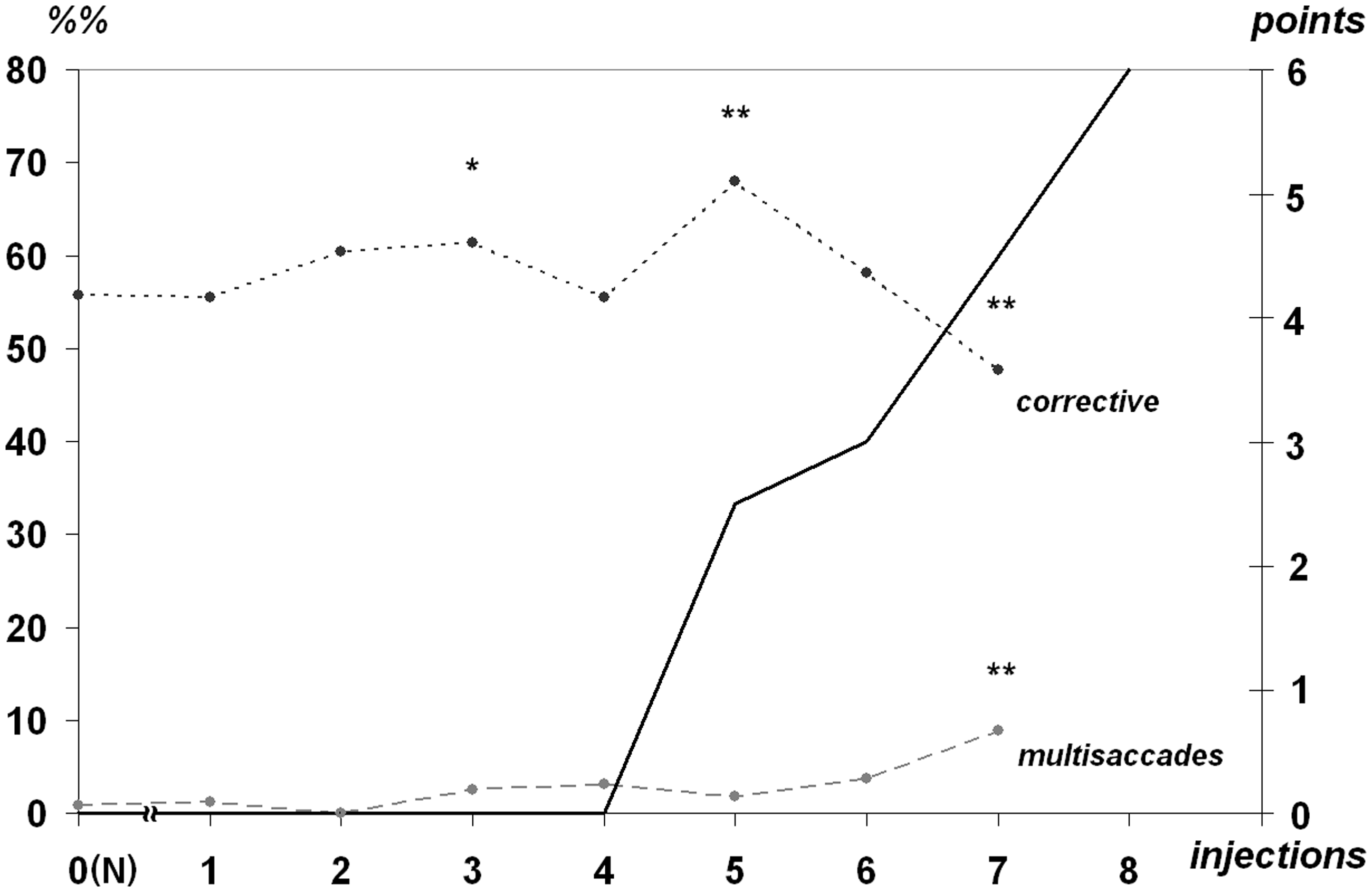
As the syndrome developed, the portion of additional corrective saccades (multisaccadic gaze transfer) was slowly increasing at the interval of 1 to 4%% and reached 9% after the same seventh injection (Figure 4).
An average peak velocity of saccades did not change significantly during the presymptomatic stage of syndrome. Visually guided saccades were characterized by peak velocity of 860 ± 230°/s while this parameter was 190 ± 80°/s for corrective saccades. Peak velocity increased significantly after the fifth and sixth injections (dose 1.0 to 1.2 mg/kg) both for visually guided and corrective saccades at 21% and 34%, respectively (Figure 5). Peak velocity values returned to previous values after the seventh injection.
The peak velocity (deg/s, left) dynamics for visually guided and corrective saccades at the MPTP-syndrome development. All references are the same as used for Figure 2.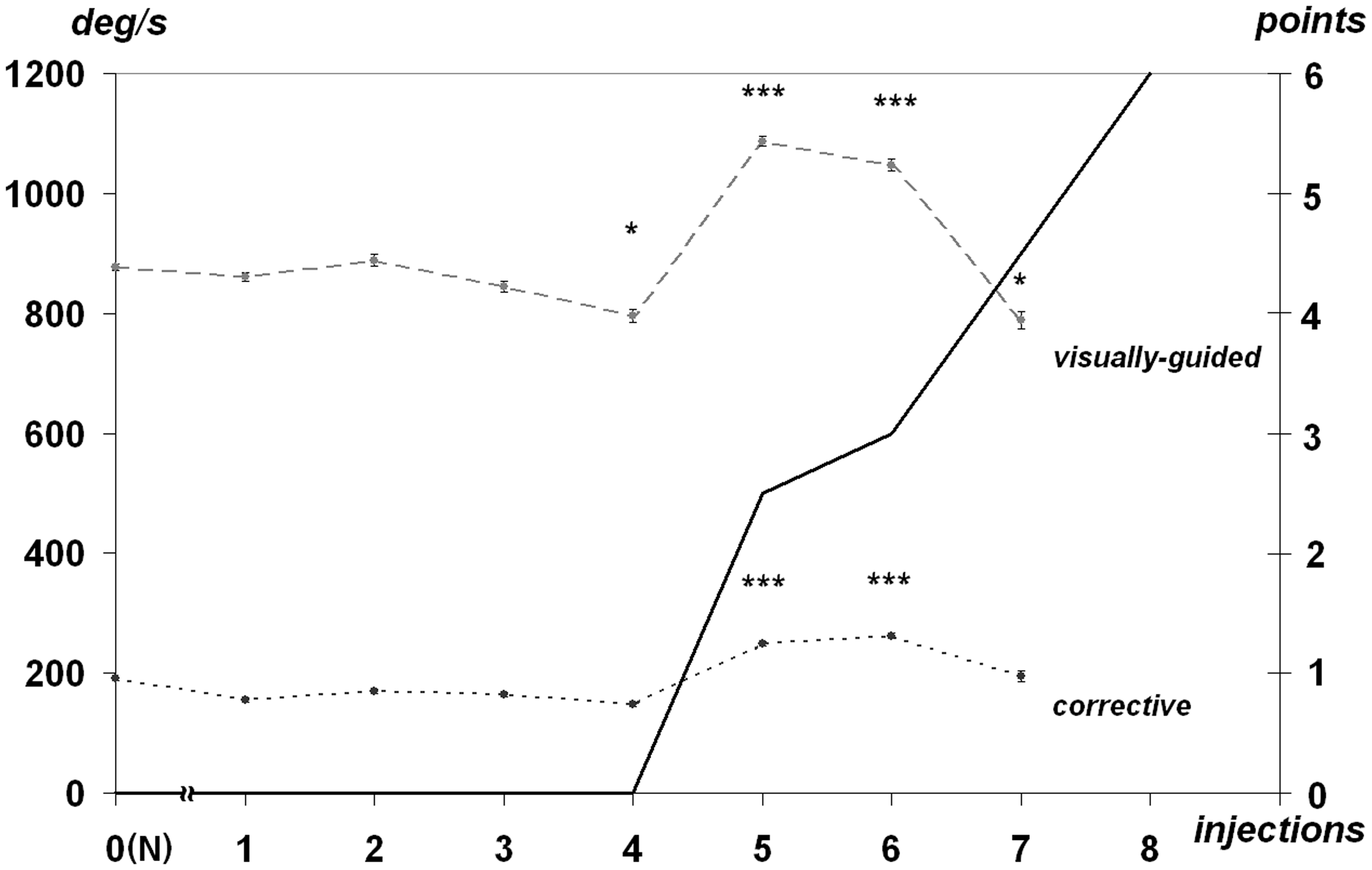
Discussion
It is crucial that behavioral changes in monkeys revealed as result of experiment described above become evident at the presymptomatic stage of the MPTP-syndrome development and precede the visible impairments in the condition and behavior of the monkeys. The visible demonstration of the MPTP syndrome starts only after the dose of 1.0 mg/kg.
Changes in the visually guided saccades latencies and the presaccadic intervals for corrective saccades after the first injection recover to a certain extent after the second injection (Figures 2 and 3). This retrace may be brought about by compensatory mechanisms of dopaminergic system which delays the motor symptoms at the early stage of PD (Bezard et al., 2001; Boulet et al., 2008; Mounayar et al., 2007). Irreversible dynamics of these parameters develop only after the third injection (the dose of 0.6 mg/kg).
We revealed an unexpected increase in peak velocity of visually guided and corrective saccades at the dose of 1.0 to 1.2 mg/kg after visible manifestations of motor disorders of MPTP syndrome. However, an insignificant increase in peak velocity is shown in relation to PD patients in the earlier studies (Jones & DeJong, 1971).
Our results pertaining to the execution of saccades with small amplitude (with the amplitude of 6.5° and corrective saccades) lead to the conclusion that foveation may already be damaged at the presymptomatic stage of MPTP syndrome: the gaze shift to target with small eccentricity is carried out with a considerable delay and in case of erroneous performance, its correction is executed with an additional delay. Thus, impairments of executing small saccades may cause deficits in focal mode during exploration of visual scenes in “ambient-focal” model of viewing (Unema, Pannasch, Joos, & Velichkovsky, 2005). Deficits in focal mode of viewing, in its turn, may cause cognitive disorders which are commonly described in PD and MPTP syndrome.
It should also be noted that the credible differences were observed in a number of parameters: (a) duration of presaccadic intervals of corrective saccades manifested themselves after the dose of 0.2 mg/kg; (b) saccadic latencies of low amplitudes (6.5°)—after the dose of 0.6 mg/kg; (c) saccadic latencies of average amplitudes (13°) after the dose of 0.8 mg/kg.
The obtained results could be of great importance for developing practical methods of early diagnostics of PD. It is recommended that visuomotor tests designated to explore execution of saccades of low amplitude and corrective saccades in visuomotor behavior are developed. The intensive development and expansion of computerized systems of video-oculography make clinical implementation of such research addressed to mass audience at risk much easier. Further comparison of the results among those of them who would face PD and unaffected individuals with consideration of the proposed factors would make it possible to prove the assumption that pathological changes at MPTP syndrome correlate with PD development and might make it possible to develop early PD diagnostics.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.