
Abstract
Tonality and harmony in music provide a structured framework to explore the neural mechanisms underlying uncertainty. The diminished seventh (dim7) chord, characterized by its multiple potential resolutions, serves as an ideal stimulus for examining the neural correlates of tonal uncertainty. This functional magnetic resonance imaging study investigated neural activity elicited by the dim7 chord in listeners with refined music listening skills, focusing on three harmonic progressions: CHANGE, modulating to a distantly related tonality; RETURN, resolving to the original tonality; and PERSIST, sustaining tonal ambiguity through repeated dim7 chords. A finite impulse response model was used to analyze responses in regions associated with uncertainty processing. The intraparietal sulcus (IPS) uniquely exhibited distinct temporal activation patterns across the three conditions. In CHANGE, IPS activity initially decreased, likely reflecting reduced confidence in the outdated tonal model’s predictions, followed by an increase as sensory evidence for a new tonality accumulated. In RETURN, IPS activity showed limited fluctuation. In PERSIST, a progressive increase in IPS activity may reflect heightened cognitive demands for maintaining and evaluating multiple tonalities. These findings deepen our understanding of predictive coding by highlighting the nuanced role of the IPS in processing uncertainty.
Uncertainty is an inherent part of everyday life, manifesting in diverse forms that require different approaches to resolve. Previous studies have identified several brain regions involved in processing uncertainty, with the anterior insula frequently activated across various tasks and contexts. A meta-analysis by Morriss et al. (2019) categorized uncertainty into three contexts: basic threat and reward uncertainty, decision-making, and associative learning. Their findings highlighted the anterior insula’s central role across all these contexts, suggesting it tags the salience of anticipated uncertainty in relation to internal bodily states. In addition, their study found that during associative learning under uncertain conditions, the dorsal anterior cingulate cortex (dACC) and amygdala are also activated, emphasizing these regions’ involvement in anxiety and stress responses.
A growing body of research over the past decade has underscored the role of the intraparietal sulcus (IPS) in processing uncertainty. As a node of the frontoparietal network (FPN), the IPS works closely with prefrontal regions and is consistently implicated in tasks requiring attention, quantification, calculation, and the maintenance or manipulation of information in working memory (WM) (Bray et al., 2015; Sokolowski et al., 2017). Feng et al. (2022) showed that the IPS, prefrontal cortex, and anterior insula are consistently co-activated across risk, ambiguity, and threat anticipation tasks, suggesting their shared role in processing uncertainty. Moreover, S. Wu et al. (2021) found that both risk and ambiguity activated the anterior insula, while ambiguity specifically engaged the dorsolateral prefrontal cortex (dlPFC) and IPS.
The IPS’s role in uncertainty processing reflects its broader involvement in prediction. Ficco et al. (2021) identified consistent IPS, anterior insula, and dACC activation across sensory modalities during predictive tasks. Similarly, Costa et al. (2024) found that prediction congruency and incongruency engaged a dynamic network—including the IPS, anterior insula, and inferior frontal gyrus—that supports context- and stimulus-dependent model updating.
This role in prediction is mirrored in the musical domain. For instance, research shows that medium-complexity rhythms evoke stronger IPS activity than high-complexity ones, suggesting it functions as an interface between motor-driven temporal predictions and auditory input (Matthews et al., 2020). Regarding tonal structure, the IPS directs attentional resources to maintain and manipulate musical pitch information in WM (Foster & Zatorre, 2010; Koelsch et al., 2009). This involvement becomes more pronounced when processing the fine-grained pitch structures of music as compared with the pitch structures in tonal languages (Tsai et al., 2018). Although this body of work establishes the IPS as a crucial node for holding, parsing, and integrating musical information, the precise mechanisms by which it coordinates these functions to resolve tonal uncertainty remain an open question.
Musical Context: The Diminished Seventh Chord
Music offers a compelling framework for exploring how the brain manages and reduces uncertainty, particularly through the lens of Bayesian predictive coding (Friston, 2010; Kilner et al., 2007). Within this framework, listeners construct internal models based on prior musical contexts, which are used to generate predictions about forthcoming musical elements. Deviations between predictions and actual outcomes generate prediction errors, prompting the brain to update internal models to reduce uncertainty (Koelsch et al., 2019). Tonality is fundamental to the internal pitch model, which may require revision—such as during modulation (key change)—when out-of-key notes introduce significant prediction errors (surprises). The Western musical tradition examined in this study showcases a rich array of techniques for achieving modulation.
Even when modulation does not occur, out-of-key notes can heighten tonal uncertainty. Li et al. (2021) demonstrated that chromatic music, which introduces tonal ambiguity with out-of-key notes, elicits greater FPN activation, compared with the tonally stable diatonic music. The authors proposed that during exposure to chromatic music, the IPS supports the representation of relevant harmonic progressions across multiple keys, a process that imposes significant demands on WM.
Among the chromatic elements that heighten tonal uncertainty, the diminished seventh (dim7) chord stands out as a quintessential example. The dim7 chord features a symmetrical pitch structure, characterized by equal intervals between its notes. This symmetry allows the dim7 chord to be interpreted in four distinct ways, each leading to six possible resolutions (Piston & DeVoto, 1987). Consequently, this chord offers 24 potential functional interpretations, which contribute to its pronounced tonal ambiguity. This high level of tonal ambiguity allows the dim7 chord to serve as a pivot chord between distantly related tonalities, enabling remote modulations where the new tonality is separated from the original by more than two steps along the circle of fifths.
Despite its theoretical importance and frequent use in Western music, the dim7 chord has been largely overlooked in neuroscience research, as the field has traditionally focused on violations of expectation (surprises) in chord progressions (Seger et al., 2013; Steinbeis et al., 2006; Tillmann et al., 2003). While recent predictive coding studies have started to distinguish between surprise and uncertainty (Cheung et al., 2019; Gold et al., 2023), this distinction underscores the need to examine tonal uncertainty—defined as ambiguity within the internal pitch models—on its own terms. This is particularly crucial given evidence that the FPN and cingulo-opercular network (CON; including the dACC and anterior insula) respond differentially to these two forms of information. T. Wu et al. (2021) found that while both networks are robustly activated by event-related surprise, the FPN shows greater and more selective sensitivity to contextual uncertainty. Their findings suggest that uncertainty processing is particularly central to the functional role of the FPN.
The Present Study
The present study leverages the unique ambiguity of the dim7 chord to investigate how the brain processes tonal uncertainty. We compared neural responses across three distinct harmonic contexts: RETURN, in which the chord resolves to the original key (low uncertainty); PERSIST, in which the chord is repeated without resolution (sustained uncertainty); and CHANGE, in which the chord resolves to a distantly related key (high transient uncertainty).
Given the IPS’s multifaceted role in WM, attention control, and probabilistic inference in uncertain contexts (Bray et al., 2015; Sokolowski et al., 2017; S. Wu et al., 2021), we formulated three specific hypotheses about the temporal dynamics of IPS activation. In RETURN, we predicted a modest IPS increase, reflecting brief surprise and quickly resolved uncertainty. In PERSIST, we expected sustained IPS activation due to the ongoing evaluation of multiple competing tonal interpretations. Although uncertainty remains elevated, surprise is comparable to the other two conditions, since the same dim7 chord is repeated rather than newly introduced. In CHANGE, we hypothesized that the IPS dynamically updates these representations as internal models undergo revision. The neural response in the IPS would unfold in two distinct stages. In the first stage, the introduction of out-of-key notes disrupts the previously established key. This disruption likely leads to a transient reduction in IPS activation, as the brain recalibrates its attentional resources by reducing reliance on the internal model associated with the original tonality. In the second stage, as sensory evidence supporting the new tonality accumulates through the sequence of chords, the IPS is engaged in updating the tonal model. This process may be marked by a corresponding rise in IPS activation. In such tasks, IPS activity has been shown to correlate with the progressive integration of sensory information to resolve uncertainty (de Lafuente et al., 2015; Hanks et al., 2015; Roitman & Shadlen, 2002).
Our hypothesis regarding the biphasic IPS response—characterized by suppression followed by rebound—drew inspiration from analogous patterns observed in the sensorimotor cortex. Sensorimotor beta rhythms are typically suppressed during movement initiation, reflecting the recalibration of neural activity to adapt to dynamic changes, followed by a rebound during post-movement adjustments that signifies the restoration of system stability (Engel & Fries, 2010; Kilavik et al., 2013). We proposed that a similar suppression-rebound dynamic in the IPS reflects broader predictive coding mechanisms that operate across both motor and cognitive domains. Supporting this interpretation, Darch et al. (2020) suggest that beta suppression reflects a preparatory disengagement from existing motor predictions, facilitating the formation of new ones—paralleling our interpretation that IPS suppression reflects reduced reliance on the outdated tonal model.
To investigate the complex temporal dynamics of neural activity, this study employed the Finite Impulse Response (FIR) model. Unlike conventional Hemodynamic Response Function (HRF) approaches that impose a fixed double-Gaussian shape on the BOLD response, the FIR model makes no a priori assumptions about response shape (Damascelli et al., 2022; Lindquist et al., 2009; Pomares et al., 2013; Reynolds et al., 2006). This data-driven flexibility is particularly well suited for modeling neural responses to complex stimuli, which may elicit non-canonical or multiphasic patterns. In our study, the FIR model proved critical for revealing the biphasic profile of IPS activity in CHANGE—an effect that may have been obscured using HRF-based convolution. By estimating each time bin independently, the FIR approach enabled us to track the unfolding of tonal uncertainty across time. We applied this approach to key regions implicated in uncertainty processing, including the anterior insula, dACC, amygdala, and IPS, to gain deeper insight into the temporal evolution of neural responses to tonal ambiguity.
Methods
Participants
Participants were recruited through an online public announcement specifying a requirement for advanced familiarity with Western classical music. The study did not restrict participation to individuals with formal musical training, in line with the view proposed by Tillmann et al. (2000): through extensive exposure to Western music, individuals can develop tonal perception without explicit instruction in music theory. This suggests that even those without formal training can acquire tonal perception and sensitivity to harmonic structure.
To ensure the recruitment of listeners with refined music perception, prospective participants completed an online pre-screening test designed to assess their ability to detect tonal modulations and mode shifts in musical excerpts. For the modulation task, participants listened to six short musical passages, each constructed using the same design principles as the stimuli in the present dim7 study. Each condition was represented by two stimuli, presented in a fixed order. Participants were asked to identify whether each passage (1) remained in the original key (RETURN), (2) modulated to a new key (CHANGE), or (3) left the key ambiguous (PERSIST).
For the mode-shift task, participants listened to another set of four passages and were asked to identify the mode trajectory of each excerpt, choosing among the following categories: (1) starting in major and ending in major, (2) starting in major and ending in minor, (3) starting in minor and ending in major, or (4) starting in minor and ending in minor. Each of the four categories was presented once, in a fixed order.
A total of 111 volunteers completed the online test, and 51 who achieved perfect scores (100% accuracy) were subsequently invited to participate in the functional magnetic resonance imaging (fMRI) experiment. Of the 22 healthy adults initially enrolled, two were excluded: one due to a dental retainer that could compromise fMRI data quality, and another who chose to withdraw due to discomfort during the scanning procedure. The final sample consisted of 20 participants (12 male, 8 female) aged 20 to 30 years. All participants reported no history of neurological or psychiatric disorders, and 10 reported formal musical training of 6 years or more.
Stimuli
Three types of stimuli were used in this study, as illustrated in Figure 1. These stimuli were meticulously crafted to elicit distinct types of tonal uncertainty by varying harmonic progressions. Each stimulus consisted of nine chords arranged for four voices, spanning five bars (8 s) in a 3/4 time signature. The first three bars were identical, establishing a stable tonal context. Tonal uncertainty was introduced at the end of bar 3 with a dim7 chord, which disrupted the diatonic framework and created ambiguity about the impending resolution.
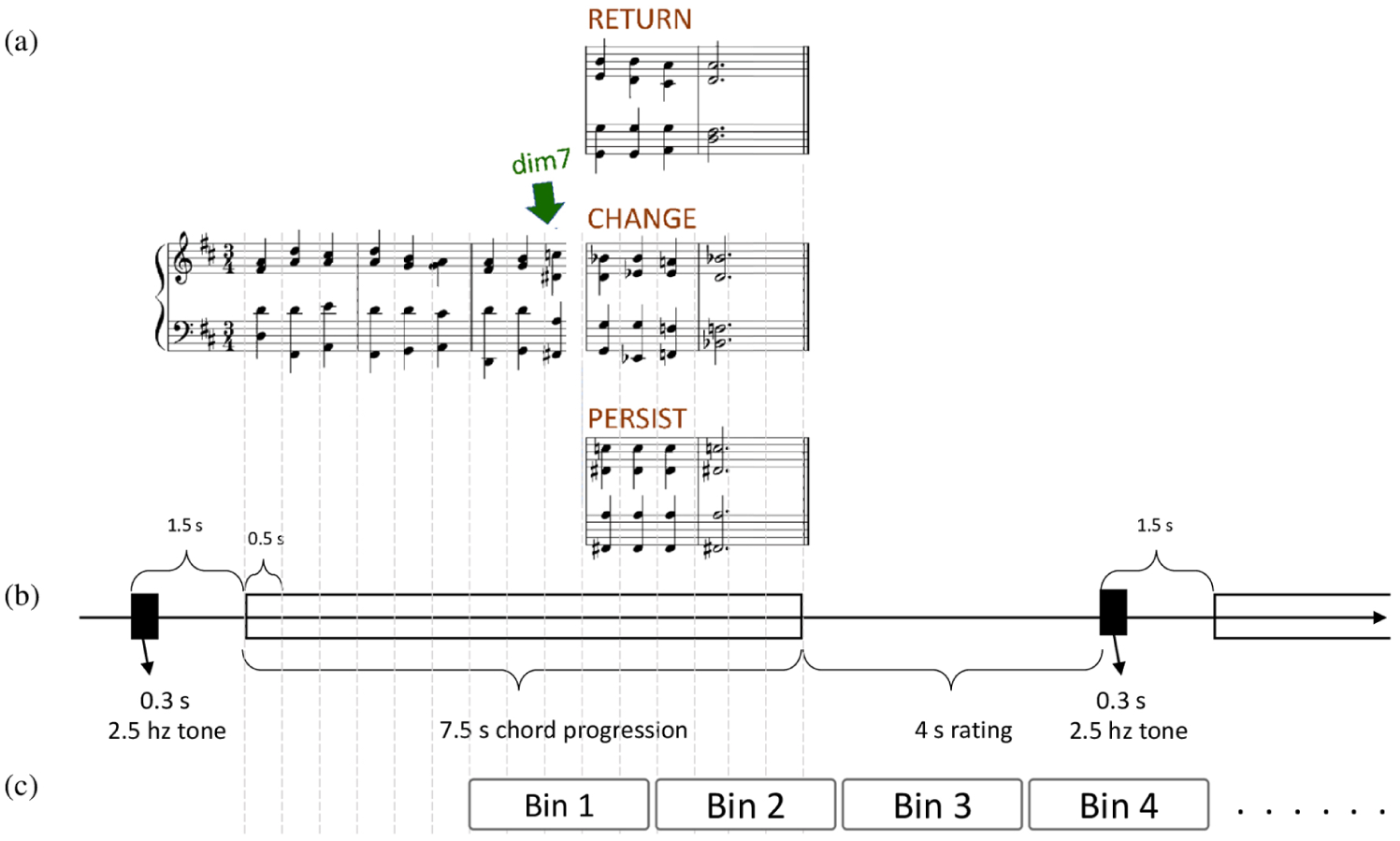
Experimental design and stimuli. (a) Musical stimuli illustrating the three harmonic conditions. Note. Tonal ambiguity is introduced at the end of the third bar with a dim7 chord. In the subsequent two bars, the stimuli diverge: the RETURN stimulus resolves back to the original tonality (D major); the CHANGE stimulus modulates to a remote tonality (B♭ major); and the PERSIST stimulus repeats the dim7 chord from the third bar, leaving the ambiguity unresolved. (b) Timeline of a single trial. (c) Time bins used in FIR analysis.
The final two bars of the stimuli diverged to represent three distinct conditions. In RETURN, diatonic chords from the original key were employed, reaffirming tonal stability. In CHANGE, the dim7 chord served as a pivot to modulate into a distantly related key. Both the RETURN and CHANGE stimuli concluded with an authentic cadence, resolving from the dominant to the tonic chord. By contrast, the PERSIST stimuli sustained tonal ambiguity by repeating the dim7 chord introduced in the third bar.
All stimuli were composed by the first author of this study. The notes in these stimuli spanned a range from F2 to C6. The tone sequences were encoded in MIDI format and converted to MP3 files using Reason 7.0 (Propellerhead Inc., Stockholm, Sweden) with the virtual instrument “BRIGHTPIANO.”
Procedure
On the day of neuroimaging, participants completed a pre-scan practice session outside the scanner to familiarize themselves with the procedure. During this 3-min session, they were presented with four sample stimuli in a pseudorandom order: two PERSIST, one CHANGE, and one RETURN stimuli. After each stimulus, participants rated their subjective emotional valence on a 4-point Likert-type scale using a button press (1 = mostly negative, 2 = somewhat negative, 3 = somewhat positive, 4 = mostly positive). Participants were instructed to interpret positive emotions as feelings of resolution or comfort, and negative emotions as feelings of tension or unease.
The 52-min fMRI session began with an anatomical scan, followed by five functional runs—three from a separate mode-shifting study alternating with two runs from the current study. This design minimized affective habituation to similar musical stimuli. The stimuli used in the mode-shifting study resembled those employed in the online pre-screening mode-shift task.
During the two fMRI runs specific to this study, each trial began with a 0.3-s warning tone at 2.5 kHz, followed by 1.2 s of silence to signal the onset of the stimulus. After each stimulus, participants had 4 s to rate their subjective emotional valence on a 4-point Likert-type scale using a button press. To minimize potential confounds from brain activation related to finger movement, the button press order was counterbalanced across participants. The study comprised three conditions, with each condition including 18 trials (each stimulus presented twice), resulting in a total of 54 trials. These trials, each lasting 13 s, were presented in a pseudorandom order across the two fMRI runs.
MRI Data Acquisition and Pre-Processing
Participants underwent scanning at the Imaging Center for Integrated Body, Mind, and Culture Research at National Taiwan University using a 3T MR system (MAGNETOM Prisma, Siemens, Erlangen, Germany) equipped with a 20-channel array head coil. Functional scans were acquired using gradient echo planar imaging (EPI) to capture axial slices of the entire brain, with a slice thickness of 2.5 mm. The imaging parameters were as follows: repetition time = 2,500 ms, echo time = 30 ms, flip angle = 87°, field of view = 192 × 192 mm2, and acquisition matrix = 78 × 78 × 45. For spatial normalization during preprocessing, a Magnetization Prepared Rapid Gradient Echo (MPRAGE) T1-weighted scan was acquired for each participant, providing isotropic 0.9 mm spatial resolution.
Image preprocessing and analysis were performed using SPM12 in MATLAB. The preprocessing pipeline included standard steps such as slice timing correction, realignment, co-registration, normalization to Montreal Neurological Institute (MNI) space, and spatial smoothing with an 8 mm full width at half maximum Gaussian kernel. The preprocessed data were subsequently used for statistical analysis, including the application of a FIR model.
Behavioral Analysis
Participants’ responses to the three conditions were analyzed using a within-subject analysis of variance (ANOVA) with Condition as a factor. Post hoc pairwise comparisons were corrected for multiple comparisons using the false discovery rate (FDR). Statistical significance was set at p < .05, and effect sizes were reported as partial eta-squared (η²p) for the ANOVA and Cohen’s d for post hoc tests.
FMRI Analysis
Functional MRI data preprocessing and analysis were conducted using SPM12. Preprocessing steps included co-registration of the functional images to the individual anatomical image, normalization to MNI template space with resampling to 2 × 2 × 2 mm voxels, and spatial smoothing with an 8 mm FWHM Gaussian kernel. All 20 participants demonstrated head motion within acceptable limits (<3.0 mm displacement and <1° rotation of the head in any direction within a session).
We employed an FIR model in SPM12 to capture the temporal dynamics of the BOLD response without assuming a fixed hemodynamic response function. The model encompassed 8 time bins of 2.5 s each, covering a 20-s window starting from the beginning of the third bar to capture neural activity following the onset of the musical stimuli coding the three conditions (Figure 1). This approach allowed for the estimation of condition-specific BOLD responses at each functional time point relative to the mean response across the two fMRI runs. The three conditions were modeled separately with six realignment-derived motion parameters included as nuisance regressors.
To ensure stable estimation despite multiple free parameters, the model maintained a balanced trial-to-regressor ratio. Each condition comprised 18 trials, yielding a trial-to-regressor ratio of 2.25:1, which is acceptable within a constrained region-of-interest (ROI) framework (Glover, 1999; Ollinger et al., 2001). The bin width was set equal to the repetition time (TR) (2.5 s) to minimize interpolation error and improve modeling precision. Visual inspection confirmed smooth and physiologically plausible BOLD response profiles, supporting stable FIR estimation despite the conservative trial number. This approach balances temporal resolution with statistical power in ROI-based FIR analysis.
We conducted a ROI analysis to examine the temporal dynamics of neural activity in key brain regions associated with uncertainty processing. The analyzed regions included the bilateral IPS, amygdala, anterior insula, and dlPFC. These regions were defined based on previous literature (Feng et al., 2022; Morriss et al., 2019; S. Wu et al., 2021), as summarized in Supplemental Table S1. Using the Marsbar toolbox, spherical volumes with a 6 mm radius were created for each ROI, and mean beta weights were extracted across all voxels within each ROI for each time bin and condition.
Although regions such as the inferior frontal gyrus, superior temporal gyrus, and basal ganglia are frequently implicated in processing harmonic expectancy violations (Bianco et al., 2016; Musso et al., 2015; Seger et al., 2013; Tillmann et al., 2006), we deliberately excluded them from the ROI analysis. This is because, in our stimuli, the initial dim7 chord was identical and time-locked across all three conditions, between-condition contrasts would largely subtract out the chord’s “surprise” itself. Consequently, we focused on regions implicated in resolving post-violation uncertainty rather than canonical violation detection.
The extracted contrast weights for each condition and time bin for each individual were then used to construct FIR time series plots, visualizing BOLD signal intensity changes across the 20-s time window from condition stimuli onset. Statistical analyses were conducted using R (version 4.1.0). A repeated-measures ANOVA was performed with two within-subject factors: Condition (RETURN, PERSIST, CHANGE) and Time Bin. The ANOVA model included Subject as a random effect to account for individual differences.
Post hoc analyses were conducted for significant main effects and interactions. Pairwise comparisons were performed using estimated marginal means for multiple comparisons. For significant Condition × Time Bin interactions, simple main effects analysis was conducted to understand how the effect of Condition varied across different Time Bins, or conversely, how the effect of Time Bin differed among Conditions. In addition, paired t-tests were conducted for each pair of conditions at each time bin. All post hoc comparisons, simple main effects analyses, and paired t-test employed FDR correction for multiple comparisons.
To explore potential associations between brain activity and subjective ratings, we conducted Pearson correlation analyses between the FIR amplitude in each ROI and the emotional valence ratings across all conditions and time bins. To control for the large number of comparisons, we applied an FDR correction with a threshold of q < 0.05.
Results
Behavioral Results
Repeated-measures ANOVA revealed a significant main effect of condition on participants’ ratings, F(2, 38) = 76.42, p < .001, partial η² = 0.80. Post hoc pairwise comparisons indicated that PERSIST (M = 1.94, SD = 0.60) was rated significantly lower than both CHANGE (M = 3.25, SD = 0.43; p < .001, Cohen’s d = 2.38) and RETURN (M = 3.39, SD = 0.36; p < .001, Cohen’s d = 2.79). However, no significant difference was found between CHANGE and RETURN (p = .109, Cohen’s d = −0.38). These results suggest that PERSIST elicited significantly more negative emotions than the other two conditions.
FMRI Results
The temporal dynamics of BOLD signal changes in the ROIs across conditions are shown in Figure 2. Across ROIs, activity rose to a local maximum between bins 3 and 6, though the peak timing differed by region. The anterior insula peaked at bin 3, whereas the dACC and dlPFC peaked at bins 4–5. By contrast, the IPS showed the most pronounced CHANGE-related suppression, yielding the only significant Condition × Bin interaction among the ROIs.
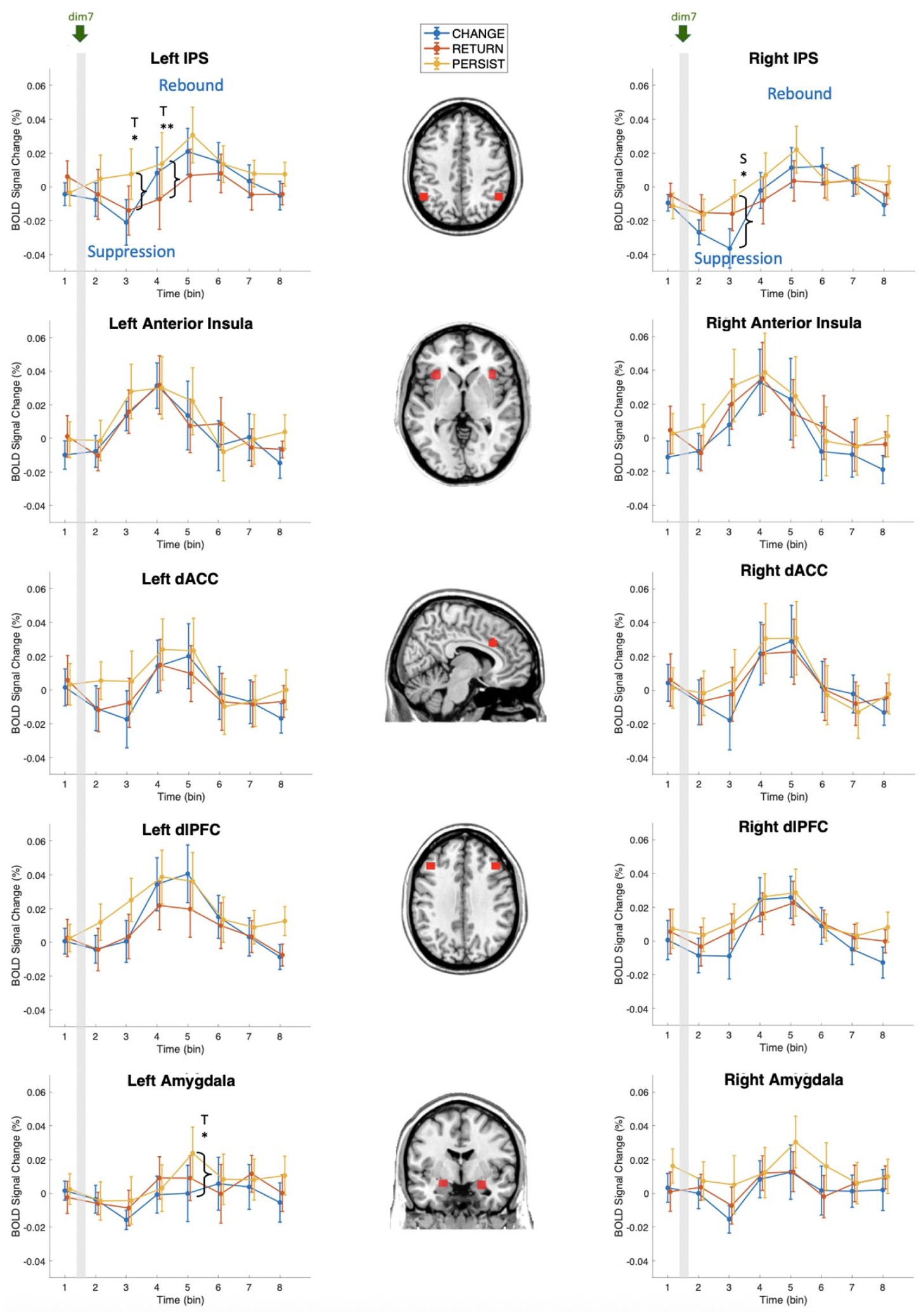
Temporal dynamics of BOLD signal changes in ROIs across conditions.
A repeated-measures ANOVA revealed significant main effects of Time Bin in several ROIs, including the right IPS, F(7, 133) = 3.719, p = .001, partial η² = 0.16, bilateral anterior insula, right: F(7, 133) = 3.428, p = .002, partial η² = 0.15; left: F(7, 133) = 4.56, p < .001, partial η² = 0.19, bilateral dlPFC, right: F(7, 133) = 2.641, p = .014, partial η² = 0.12; left: F(7, 133) = 4.369, p < .001, partial η² = 0.19, and right dACC, F(7, 133) = 2.574, p = .016, partial η² = 0.12. The temporal dynamics of BOLD signal changes in the bilateral IPS across conditions are shown in Figure 2. No significant main effects of Condition were observed in any ROI. Critically, significant interactions between Time Bin and Condition were observed in bilateral IPS, right: F(14,266) = 2.199, p = .008, partial η² = 0.10; left: F(14, 266) = 1.989, p = .019, partial η² = 0.09, indicating that the temporal dynamics of activation in these regions varied across harmonic progression types. Detailed results of the repeated-measures ANOVA are provided in Supplemental Table S2.
Detailed results of the post hoc analyses using estimated marginal means are provided in Supplemental Tables S3 and S4. The main effect of Time Bin (detailed in Supplemental Table S4) in the left anterior insula was driven by significant differences between bin 2 and bin 4, t(133) = −4.066, p_FDR = 0.037, Cohen’s d = −0.09, and between bin 4 and bin 8, t(133) = 3.996, p_FDR = 0.037, Cohen’s d = 0.89. Importantly, simple main effects analysis (detailed in Supplemental Table S5) of the interaction in the right IPS showed a significant difference between CHANGE and PERSIST at bin 3, t(209.98) = −3.796, p_FDR = 0.014, Cohen’s d = −0.85. Furthermore, within CHANGE, significant differences were observed between bin 3 and bin 5, t(257.95) = −4.682, p_FDR = 0.005, Cohen’s d = −1.05, and between bin 3 and bin 6, t(257.95) = −4.759, p_FDR = 0.005, Cohen’s d = −1.06.
Paired t-tests (detailed in Supplemental Table S6) revealed additional condition-specific differences. In the left IPS, significant differences between RETURN and PERSIST were observed at bin 3, t(19) = −3.42, p_FDR = 0.034, Cohen’s d = −0.32, and bin 4, t(19) = −4.64, p_FDR = 0.004, Cohen’s d = −0.26. The left amygdala showed a significantly higher BOLD response to the CHANGE condition than the PERSIST condition at time bin 5, t(19) = −3.75, p_FDR = 0.032, Cohen’s d = −0.33.
Correlation Between Brain Activity and Emotional Valence Ratings
Pearson correlation analyses revealed no significant associations between BOLD activity and emotional valence ratings after FDR correction (detailed in Supplemental Table S7).
Discussion
Musical tonal and harmonic structures introduce various forms of uncertainty, enriching the listening experience and engaging complex neural processes. In this study, we focused on the dim7 chord, a paradigmatic example of tonal ambiguity due to its multiple potential resolutions. To capture the temporal dynamics of neural responses to this ambiguity, we employed the FIR model, which offers the flexibility to model temporal patterns without assuming a fixed response shape. This approach was particularly suited to examining the hypothesized suppression–rebound pattern of IPS activity in CHANGE.
Our findings revealed significant main effects for the Time Bin factor in the bilateral anterior insula, right dACC, and bilateral dlPFC, with neural responses progressively increasing after the onset of the dim7 chord and peaking toward the end of the stimulus. However, these regions did not exhibit the same degree of sensitivity to dynamic differences across the three experimental conditions as observed in the bilateral IPS. The bilateral IPS demonstrated significant effects for the interaction between Time Bin and Condition. Notably, the hypothesized suppression–rebound pattern in CHANGE was observed, supporting the IPS’s role in recalibrating internal models during key changes. These findings align with and extend the conclusions of Costa et al. (2024), who identified the IPS as a component of a dynamic predictive network for integrating context- and stimulus-based predictions. The biphasic IPS responses observed in CHANGE highlight its specialized role in managing uncertainty, particularly in scenarios requiring rapid adaptation of predictive models.
In both CHANGE and RETURN, bilateral IPS activity exhibited an initial decline followed by a subsequent increase. The changes in IPS activity were more pronounced in CHANGE, while those in RETURN showed a more gradual and subdued pattern. To our best knowledge, this biphasic response has not been previously reported in predictive coding research. We interpret this dynamic response as reflecting the IPS’s roles in attention regulation and evidence accumulation. The initial decline in IPS activity, observed from bin 1 to bin 3, likely reflects a temporary disruption of attentional focus triggered by the tonal uncertainty introduced by the dim7 chord. This disruption may occur as the brain disengages from the outdated predictive model. Supporting this interpretation, previous studies on visual attention have documented decreases in IPS activity during transient disruptions, such as when irrelevant distractors enter the visual field (Gillebert et al., 2012) or when a tracked target unexpectedly disappears, causing a brief lapse in focus (Olson et al., 2004). The subsequent increase in IPS activity from bin 3 to bins 4 and 5 likely reflects the re-engagement of top-down attentional control (Corbetta et al., 2008; Spagna et al., 2023) to guide the accumulation of new sensory evidence needed to resolve uncertainty (de Lafuente et al., 2015; Hanks et al., 2015; Roitman & Shadlen, 2002). In CHANGE, this increase may reflect the updating of the tonal model to accommodate the modulation, while in RETURN, it may signify re-engagement with the original tonal model, resolving uncertainty by reinstating the prior tonality.
The biphasic IPS responses observed in CHANGE offer new insights into the predictive coding framework, where predicted precision reflects the expected confidence in hypotheses about future sensory events (Koelsch et al., 2019). The initial decrease in IPS activity likely reflects reduced precision assigned to the outdated tonal model, enabling attentional shift toward new input. The subsequent increase suggests elevated precision for predictions based on the new tonal model. This flexibility supports adaptive model updating under uncertainty. Notably, IPS activity during bin 3 of CHANGE was lower than in RETURN, possibly indicating lower precision assigned to outdated predictions when resolving to a remote key versus returning to the original tonality.
Our interpretation of this biphasic IPS response is conceptually aligned with findings from sensorimotor beta rhythm research. In motor domains, beta oscillations exhibit a suppression–rebound pattern during movement planning and execution (Kilavik et al., 2013; Pfurtscheller & Lopes da Silva, 1999), interpreted as a transition from stability to adaptability, where suppression reflects disruption and adaptation to novel demands, and rebound marks the return to a stable state. Tan et al. (2016) found that post-movement beta rebound correlates with confidence in motor-related predictions. Furthermore, Darch et al. (2020) proposed that sensorimotor beta rhythms reflect the process of motor adaptation, defined as the ability of an organism to modify its movement strategies to maintain accuracy in response to changes in the environment. In their experiments, both humans and cats showed reduced pre-movement beta oscillations during early adaptation phases. This reduction was interpreted as reflecting substantial internal model updating, providing a physiological substrate for behavioral flexibility. Critically, Spitzer and Haegens (2017) extended this framework to cognitive domains, proposing that beta oscillations support top-down control by flexibly reactivating task-relevant content in WM and decision-making. Taken together, the biphasic IPS response observed in our study mirrors the suppression-rebound dynamics of beta rhythms, suggesting a shared predictive coding mechanism across motor and cognitive systems.
In PERSIST, bilateral IPS activity exhibited a sustained upward trend beginning in bin 3, likely reflecting neural responses to the repeated presentation of the dim7 chord. As the chord repeated, the listener’s belief about the original tonality diminished, leading to increased tonal uncertainty. This growing uncertainty may necessitate the maintenance and evaluation of multiple potential tonalities, imposing heightened cognitive demands on predictive processing and WM. Such sustained uncertainty may explain why PERSIST elicited the most negative ratings of emotional valence. Consistent with this interpretation, a study on ambiguity avoidance demonstrated that when informational ambiguity persisted, individuals showed a greater tendency to avoid ambiguous options, and this behavioral pattern was associated with increased IPS activation as the level of ambiguity increased (Lawrance et al., 2022).
Previous research has shown that compared with diatonic music, chromatic music elicits greater IPS activity due to its higher tonal uncertainty and the increased cognitive demands of predicting multiple harmonic progressions (Li et al., 2021). These harmonic progressions, along with the hierarchical organization of tonal structures, can be conceptualized as hidden states—latent variables that the brain infers to interpret sensory input and predict future outcomes (Caucheteux et al., 2023; Friston & Kiebel, 2009). During PERSIST, the repeated presentation of the dim7 chord causes listeners to perceive increasing tonal ambiguity. This heightened ambiguity likely places demands on WM, requiring the brain to actively retain and evaluate multiple tonalities along with their associated hidden states to navigate the uncertainty.
The results of the two-way ANOVA revealed that activation in the bilateral anterior insula, right dACC, and bilateral dlPFC exhibited significant main effects for the Time Bin factor but not for the Condition factor or the interaction between Time Bin and Condition. This suggests that they are engaged in processing the overall aspects of uncertainty elicited by the dim7 chord.
In CHANGE and RETURN, activity in the anterior insula exhibited a sharp peak at bin 4, whereas activity in the dACC and dlPFC showed broader, flatter peaks spanning bins 4 and 5 (Supplemental Figure S1). The more transient activation of the anterior insula is consistent with its established role in detecting and tagging the salience of uncertainty, particularly in relation to internal bodily states (Morriss et al., 2019). The relatively sustained engagement of the dACC and dlPFC suggests their contributions to response processing once salient events have been identified. The dACC may support conflict monitoring and the modulation of cognitive control to adapt to shifting conditions (Dignath et al., 2020; Ham et al., 2013). The dlPFC appears to play a critical role in higher-order executive functions, such as the active maintenance and manipulation of information in WM (Barbey et al., 2013; Chai et al., 2018), as well as in regulating goal-directed and flexible behaviors to effectively manage uncertainty (Mushtaq et al., 2011; Schonberg et al., 2012).
This study highlights the role of the FPN in optimizing internal predictive models under tonal uncertainty, whereas the CON appears to support more general salience processing. Beyond its theoretical relevance, these findings also carry implications for music education. Given that the FPN is central to cognitive flexibility and WM, its engagement during the processing of tonal uncertainty suggests that listening to chromatic music may potentially strengthen these functions. Music provides what Koelsch et al. (2019) describe as epistemic affordance—a continuous opportunity to resolve uncertainty among competing hypotheses. Engaging with tonally ambiguous music may therefore serve as a form of cognitively enriching training, fostering the development of neural scaffolding for managing uncertainty and adapting to change in daily life. While this hypothesis requires further empirical validation, it aligns with the notion of cognitive transfer from music, whereby musical training benefits non-musical domains such as WM, mathematical reasoning, and spatial cognition (Wang, 2022).
This study has several limitations that warrant consideration. First, the study did not include a control condition consisting entirely of diatonic chords. Such a control condition would allow future research to better isolate neural responses specifically elicited by chromatic alterations, distinguishing these effects from those elicited by diatonic harmony.
Second, although the sample size was relatively small (n = 20), the observed effects were large (e.g., behavioral results: Cohen’s d > 2.0; fMRI results: partial η² up to 0.19), suggesting that the current findings are robust and unlikely to be driven by low statistical power. Nonetheless, future studies with larger and more diverse participant samples, including individuals with varying levels of musical expertise, are needed to validate and extend the generalizability of these results.
Finally, to avoid an excessively long experimental duration that might have reduced participants’ attention to the music, each trial in this study was limited to 13 s. While this design choice helped maintain engagement, it introduced potential confounds. Specifically, overlap may have occurred between neural activity related to button-press responses and activity elicited by the harmonic stimuli themselves. In addition, in CHANGE, overlapping BOLD signals for distinct sub-events (e.g., the two stages of modulation) may have reduced the temporal resolution of the findings. Future research should consider extending the duration of each trial and increasing the inter-stimulus interval to minimize these confounding effects. Employing EEG or MEG in future harmonic listening experiments offers a valuable complementary approach, leveraging their superior temporal resolution to reveal the rapid neural dynamics underlying tonal uncertainty.
The present study provides the first empirical evidence of condition-specific IPS dynamics in response to tonal uncertainty. The biphasic IPS response observed during remote modulation may reflect distinct phases of internal model updating. Moreover, our results highlight the pivotal role of the IPS in maintaining and evaluating hidden states associated with multiple tonalities under prolonged uncertainty. These findings shed light on how the IPS integrates attentional control, evidence accumulation, WM, and higher-order computation to support predictive coding in complex auditory contexts.
Supplemental Material
sj-docx-1-pom-10.1177_03057356261442576 – Supplemental material for Temporal Dynamics of Intraparietal Sulcus Activation in Response to Tonal Uncertainty: An fMRI Study of the Diminished Seventh Chord
Supplemental material, sj-docx-1-pom-10.1177_03057356261442576 for Temporal Dynamics of Intraparietal Sulcus Activation in Response to Tonal Uncertainty: An fMRI Study of the Diminished Seventh Chord by Chen-Gia Tsai, Ling-Yao Chien and Joshua Oon Soo Goh in Psychology of Music
Footnotes
Acknowledgements
We gratefully acknowledge the technical and facility support provided by the Imaging Center for Integrated Body, Mind, and Culture Research at National Taiwan University. Special thanks to Yi-Fan Fu for her assistance with data collection and to Jing-Yu Chuang for her contributions to data analysis. We also extend our gratitude to all the participants who took part in this study.
Ethical Considerations
The study was approved by the Institutional Review Board of National Taiwan University (202007HM031). Our research was performed in accordance with relevant guidelines/regulations and with the Declaration of Helsinki.
Consent to Participate
Written informed consent was obtained from all participants following a comprehensive explanation of the study. Participants were compensated approximately 16 USD upon completing the experiment.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by the grant from the Ministry of Science and Technology, Taiwan (MOST 108-2410-H-002-216).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available in the OpenNeuro repository, doi:10.18112/openneuro.ds005880.v1.0.1.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.