
Abstract
Which abiotic factors influence the number of native plant species on remote atolls is an important question to understand better the spatial pattern of the species observed on these low and vulnerable coral islands. However, this issue is still very poorly documented, often due to human degradation, partial botanical surveys or the difficult geographic access of remote atolls for researchers. The remote atolls of Eastern Polynesia, which are among the most isolated in the world, are of great interest for studies of native species’ distribution due to their isolation, low human density and urbanisation. In this study, we selected 49 remote atolls of Eastern Polynesia with complete botanical surveys to test the relative influence of eight abiotic factors on native plant species richness (i.e. indigenous and endemic species). Abiotic factors used as potential predictors included atoll area (km2), shoreline length (km), atoll elevation (m) and index of isolation (UNEP), but also the coastal index of the atoll (Ic), the distance to the nearest similar atoll (km), the distance to the nearest large volcanic island ≥ 1000 km2 (here, Tahiti as a potential stepping-stone island) and the distance to the nearest raised atoll ≥ 15 m a.s.l. (here, Makatea or Henderson as a potential refugium during sea-level highstands). Spearman’s rank correlation, linear regression analysis and frequency diagrams were used to assess the relative influence of these factors on native species richness. No relationship was found between the species richness and the index of isolation or the distance to the nearest similar atoll. Atoll area and distance to the nearest raised atoll of Makatea explained 47.1% and 40%, respectively, of the native species richness variation observed on the remote atolls. The distance to the volcanic island of Tahiti and the coastal index explained 36.9% and 27.3% of the variation, while elevation and shoreline length explained 23.3% and 18.4% of the variation, respectively. Native species richness on the atolls surveyed increased with the increasing atoll area, elevation and shoreline length, but decreased with the increasing distance to the nearest raised atoll of Makatea and the large volcanic island of Tahiti. This supports the view that the spatial pattern of native species richness observed on the remote atolls was strongly influenced by (i) atoll area but also by (ii) the distance to the raised atoll of Makatea, and (iii) the distance to the volcanic island of Tahiti. This finding suggests that the raised atoll may be viewed as a refugium during sea-level highstands while the large volcanic island played the role of stepping-stone island, both islands influencing the dispersal of native species on remote atolls and attenuating the isolation effect in the study area.
Keywords
I Introduction
The factors influencing native species richness (i.e. the total number of species naturally found on an island not including those introduced by humans) on remote oceanic islands are of particular interest in island biogeography. Native species richness on these islands is the result of species immigration (from the mainland and/or another island) and speciation leading to endemism (e.g. Gillespie, 2002; Rosindell and Phillimore, 2011). It is commonly accepted that speciation on the oceanic islands occurs in two geographic ways: sympatric speciation (i.e. one species splits into two or more island populations that become reproductively isolated from each other in situ due to genetic changes) and allopatric speciation (i.e. an island population becomes reproductively isolated from the mainland source population due to a geographic barrier such as the ocean) (e.g. Rosindell and Phillimore, 2011).
Several abiotic factors have previously been considered to explain the variation in species richness on remote islands including elevation, island area and isolation (e.g. Ackerman et al., 2007; Diamond, 1975; Paulay, 1994; Preston, 1962a, 1962b; Rosenzweig, 1995). For example, Hamilton et al. (1963) and Johnson and Raven (1973) reported that elevation was significant for native plant species richness and diversity in the Galapagos Archipelago and similar results were obtained on the tropical islands of the northeast coast of Australia (Buckley, 1985), West Indies (McMaster, 2005) and French Polynesia (Larrue, 2014). In addition, edaphic substrata, soil nutrients, climate and productivity (see species–energy theory) also influence species richness on the islands (Field et al., 2008; John et al., 2007; Kalmar and Currie, 2006; Wright, 1983). Consequently, coral atolls generally harbour a low species richness compared with high volcanic islands, possibly due to their flat topography, homogenous substrata, poor nutrients and dry climate (Stoddart, 1992).
Moreover, a large body of literature has highlighted the influence of island area on species richness showing that larger islands tend to harbour more species, thereby following the predictions of MacArthur and Wilson (1967). However, the reason for this species–area relationship (SAR) is still debated (Kreft et al., 2008; Turner and Tjørve, 2005). Many explanations have been advanced. Some island biogeography studies have pointed out a simple area effect per se by which the limited terrestrial space results in a limited number of species (Simberloff, 1976). Several workers have reported that large islands potentially contain more diverse habitats than smaller islands as the increasing area often leads to additional habitat diversity (e.g. Connor and McCoy, 1979; Triantis et al., 2003) and habitat diversity influences species richness on the islands (e.g. Deshaye and Morisset, 1988; Ricklefs and Lovette, 1999). The ‘equilibrium theory’, in which the immigration and extinction rates differ according to the island area and isolation (MacArthur and Wilson, 1967; Whittaker and Fernández-Palacios, 2007), has been most frequently used to explain species richness, but it does not include the speciation process (Rosindell and Phillimore, 2011). As pointed out by Whittaker et al. (2001) and Kalmar and Currie (2006), the literature addressing the factors that determine species richness on islands is heterogeneous, possibly because of the multiple scales of ecological processes, spatial and temporal analyses, and biogeographical studies (e.g. Triantis et al., 2012).
Some studies have reported a weak SAR on low coral islands owing to their homogeneous geomorphology with similar landforms and limestone substrata, low elevation and no orographic rainfall, thus providing poor habitat diversity (Manner, 1995; Mueller-Dombois and Fosberg, 1998; Stoddart, 1992). On the Kapingamarangi Atoll in Micronesia, Niering (1963) pointed out a small-island effect of 1.4 ha showing no SAR on the reef islands below this threshold, but a significant one above it, while some workers found a significant SAR even on the small Pacific atolls (e.g. Woodroffe, 1986). Using different geographical locations, taxons, numbers of islands, ranges of species richness, ranges of area and numbers of habitats, Triantis et al. (2006) have demonstrated that the small-island effect is an idiosyncratic phenomenon depending on the taxon studied and the characteristics of the islands such as geographical isolation, geological history, habitat diversity and degree of disturbance as well as stochastic events. In fact, there are conflicting opinions regarding the applicability of the small-island effect (Morrison 2014; Tjørve and Tjørve, 2011) and it is well known that multiple factors may influence SAR on the islands (e.g. Kalmar and Currie, 2006; Triantis et al., 2006).
The ocean is a barrier for numerous plant species that do not have the means to reach the remote islands from the mainland. Thus, the number of species reaching an island is assumed to decrease with increasing distance from the mainland source pool (Dahl, 1984; MacArthur and Wilson, 1967 ). The Pacific Islands are well known as the most isolated in the world. Accordingly, flora on the remote Pacific Islands tend to be derived from only a small pool of colonists leading to ‘taxonomic disharmony’ – i.e. taxa not dispersed by birds, ocean currents or wind are under-represented on the most isolated island (Florence, 1997; Gillespie et al., 2008a, 2008b; Meyer, 2004). Thus, isolation (sensu stricto the distance from the mainland) is often an important factor in predicting species richness on islands. However, isolation effects may be attenuated by many other factors such as the stepping-stone-island effect, which can increase the flow of seeds and propagules, for instance spores, roots and branches, to nearby islands despite their remoteness from the mainland (MacArthur and Wilson, 1967). For example, Weigelt and Kreft (2013) demonstrated that distances to large islands, stepping-stone distances and distances to climatically similar landmasses were more significant to predict species richness than distance to the nearest mainland.
The factors explaining species richness on the islands may also depend on the island type surveyed (e.g. high volcanic islands vs low coral atolls). In fact, the relationships between abiotic factors and species richness are well reported on high volcanic islands (e.g. Ackerman et al., 2007; Johnson and Raven, 1973; Larrue, 2014; McMaster, 2005; Triantis et al., 2003) but still poorly documented for the remote coral atolls.
On the coral atolls, glacial–interglacial cycles and sea-level variations during the Pleistocene and Holocene periods could be important to explain the species richness currently observed on large and small remote atolls in different contexts of isolation (Camoin et al., 2001). The flowing of islands during sea-level highstands impacted the native flora more on the small and low atolls than on the raised coral islands (Mueller-Dombois and Fosberg, 1998; Pirazzoli and Montaggioni, 1986) suggesting regression or extirpation of island biota on the low coral islands (Cibois et al., 2010). Raised atolls may act as ‘refugia’, i.e. a geographical area where native biota of geographically isolated organisms were protected as a result of relatively stable environmental conditions during times of great environmental change such as glacial–interglacial cycles and sea-level variations on the island. Organisms may then re-colonise a region when the environmental conditions return to within the organism’s tolerance levels (Bennett and Provan, 2008; Dahl, 1946). Thus, raised atolls may protect the specific biota of remote atolls during sea-level highstands and be a possible source of native pool species on the neighbouring atolls after the decline in sea level. In addition, cyclones, tsunamis and marine submersion can influence species richness on the low islands and it has been suggested that tidal waves or cyclonic swells could periodically devastate the smallest tropical islands, so that these are not maintained at ‘equilibrium’ (Fosberg, 1991; MacArthur and Wilson, 1967; Woodroffe and Stoddart, 1992 ). Consequently, the species richness of the remote atolls cannot be fully predicted by isolation alone and other factors, like the great susceptibility of these low coral islands to natural disturbances such as cyclones or sea-level variations and submersion during sea-level highstands in the Pleistocene and Holocene periods, might also influence their species number (Kreft et al., 2008).
In this study, several abiotic factors were tested to explain the spatial pattern of native vascular plant species richness on 49 remote atolls of Eastern Polynesia (east Pacific Ocean). We hypothesise that native plant species richness on the atolls is influenced by simple atoll characteristics such as atoll area, shoreline length, atoll elevation and isolation, but also by the coastal index, the distance to the nearest similar atoll, the distance to the nearest large volcanic island ≥ 1000 km2 (as a potential stepping-stone-island) and the distance to the nearest raised atoll ≥ 15 m a.s.l. (as a potential refugium during sea-level highstands).
II Study area and Data
1 Study area
Eastern Polynesia is a large phytogeographical sub-region of Polynesia including the Cook Islands, the five archipelagos of French Polynesia (i.e. from west to east: the Austral Islands, Society Islands, Tuamotu Islands, Gambier Islands and Marquesas Islands), Pitcairn Islands and Easter Island (Mueller-Dombois and Fosberg, 1998). The large volcanic island of Tahiti (Society Islands, French Polynesia) is the biggest and highest island of Eastern Polynesia, culminating at 2242 m above sea level (a.s.l.) with an island area of 1042 km2 (Dupon, 1993). It harbours the highest species richness and diversity in Eastern Polynesia with 460 native vascular plants, 107 of which are endemic (Butaud, 2013a; Florence, 1997).
The sub-region of Eastern Polynesia consists of 142 islands (Meyer, 2004), including 92 atolls scattered in the Pacific Ocean with high geographic isolation. In this article, 49 atolls with complete botanical surveys were included, mainly from the Tuamotu Islands (French Polynesia; Table 1).
Range of characteristics used in this study as predictors for native species richness and the number of coral islands surveyed in the Austral Islands, Society Islands, Tuamotu Islands, Cook Islands and Pitcairn Islands (Eastern Polynesia).
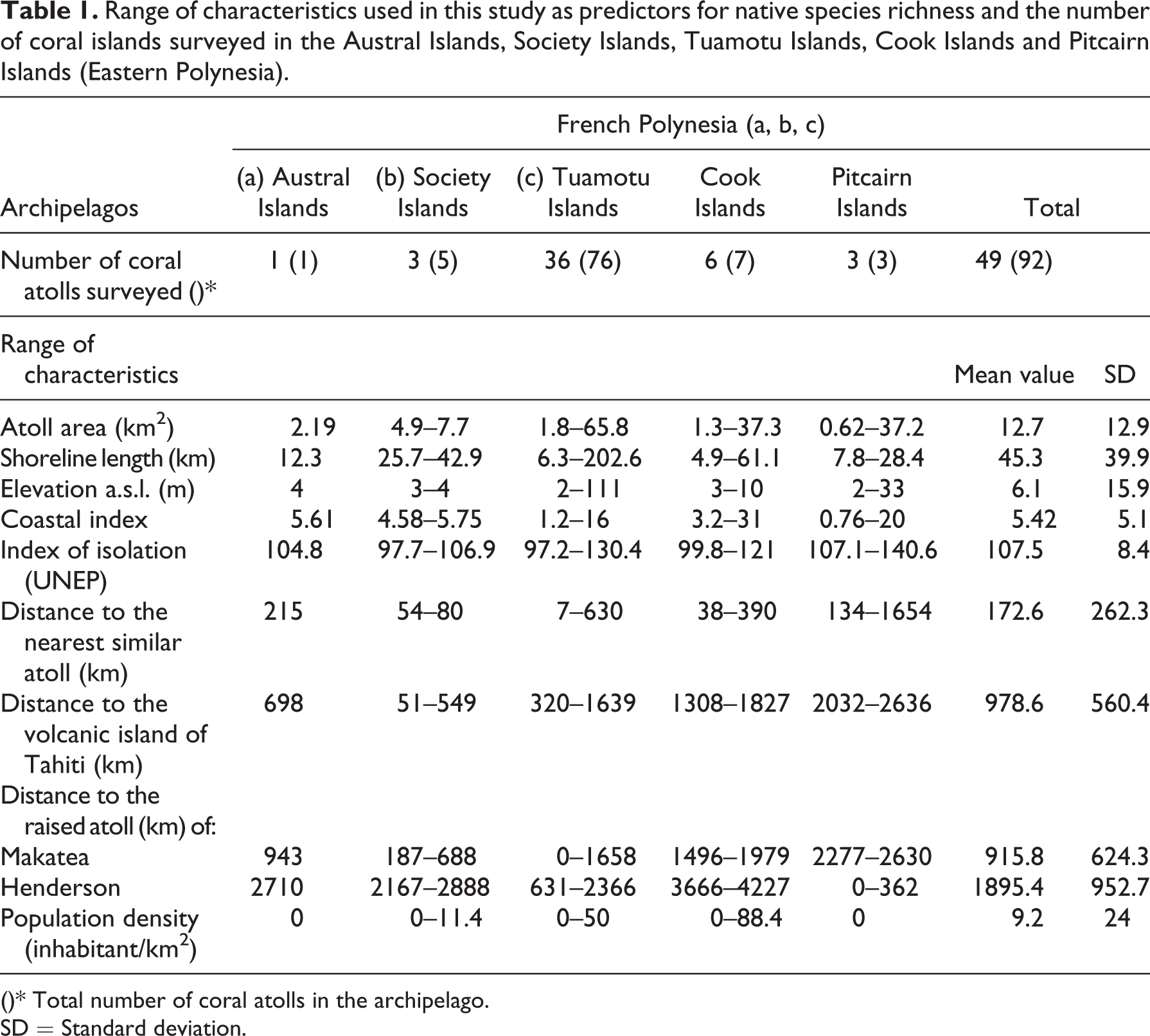
()* Total number of coral atolls in the archipelago.
SD = Standard deviation.
2 Atolls surveyed
The 49 atolls surveyed in this study are located between 8°59’57.6’S – 24°40’49.8’S and 165°50’20.5’W – 124°47’16.9’W. In French Polynesia 40 islands are found, six in the Cook Islands, and three in the Pitcairn Islands (Figure 1). These remote atolls are among the most isolated in the world and have never been connected to a mainland (Mueller-Dombois, 2002; Nunn, 1994). They are located in a tropical region influenced by south-easterly trade winds with a mean annual sea-level rainfall decreasing from west to east, from 2500 mm on the Cook Islands to 1750 mm on the Pitcairn Islands (Stoddart, 1992). The terrestrial areas of the 49 coral atolls range from 0.62 to 65.8 km2 (Table 1) and substrates correspond to sedimentary limestone composed of skeletons of marine organisms such as coral and foraminifera (Dahl, 1980; Dupon, 1993; Rougerie, 1995).
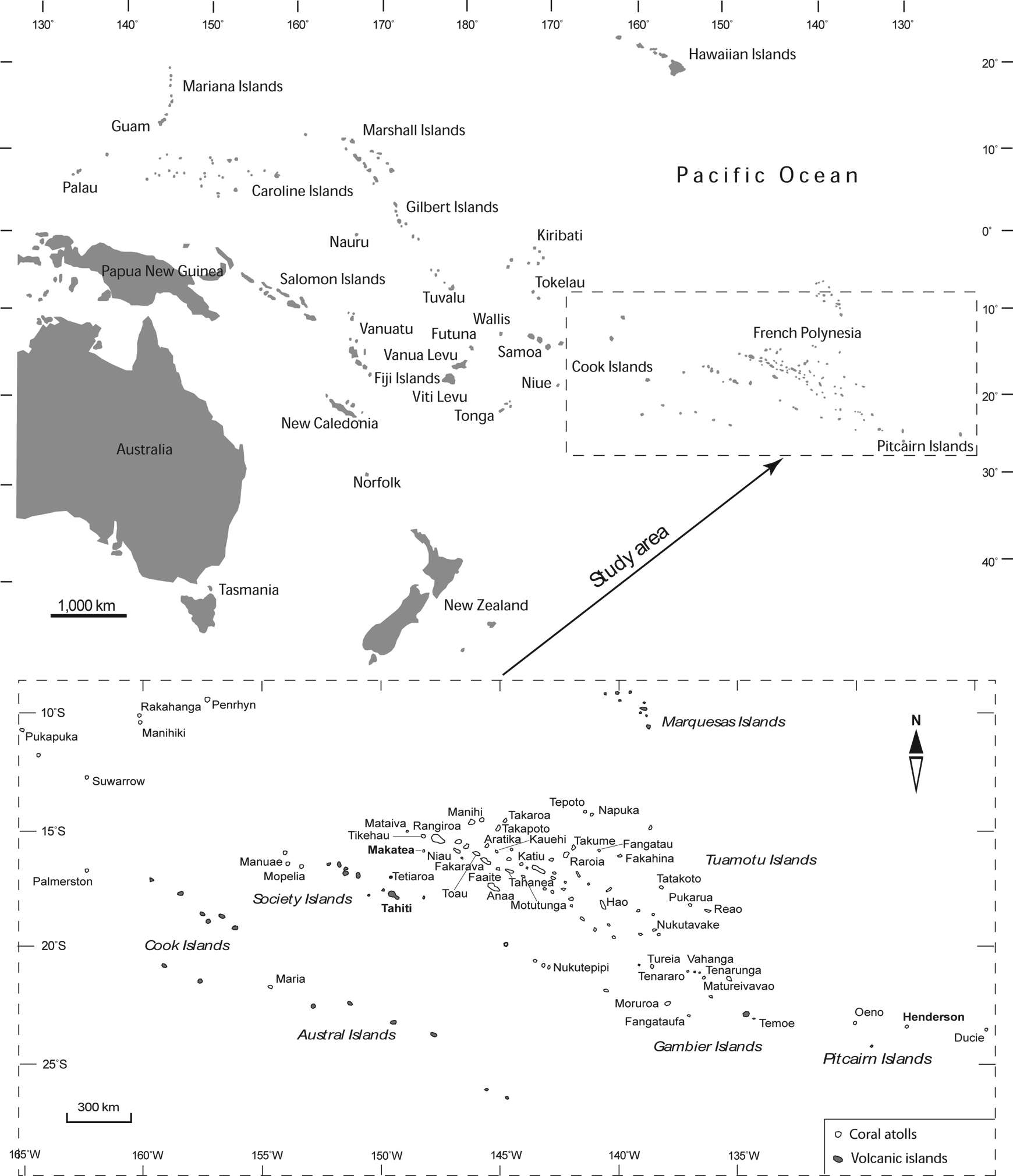
Study area and location of the 49 coral islands surveyed in Eastern Polynesia (Pacific Ocean) including 47 low atolls (regular character) and the two raised atolls of Makatea (111 m a.s.l., Tuamotu Islands) and Henderson (33 m a.s.l., Pitcairn Islands) (boldface). The large volcanic island of Tahiti (Society Islands, French Polynesia), 1042 km2 in area, has been added as a landmark.
The 49 atolls surveyed included 47 low atolls (i.e. low coral islands formed by islets called motu, with a ring-shaped coral reef and a coral rim surrounding a lagoon) and the two raised atolls of Makatea (111 m a.s.l., Tuamotu, French Polynesia) and Henderson (33 m a.s.l., Pitcairn Islands). The elevation of the 47 low atolls ranges between 2 and 6 m a.s.l. with a mean elevation of 3.2 m a.s.l. Anaa and Niau (Tuamotu Island) are the highest at 5 m and 6 m a.s.l., respectively (Dupon, 1993). The raised atolls of Makatea and Henderson are formed by an elevated terrestrial plateau (i.e. an old lagoon floor) surrounded by an eroded coral ring. Both were uplifted by the lithospheric flexure caused by the crustal loading of the volcanic islands of Tahiti and Pitcairn, respectively (Larnbeck, 1981; McNutt and Menard, 1978; Spencer, 1989). Finally, 70% of the islands surveyed are inhabited by Polynesians and the population density ranges from 0 to 88 inhabitants per km2 with a mean value of 9.2 inhabitants per km2 (Table 1).
III Methods
1 Characteristics of the coral islands
The following factors were selected as potential predictors of native plant species richness on the coral atolls surveyed: atoll area, shoreline length, atoll elevation (i.e. maximum substrata elevation), coastal index, index of isolation, the distance to the nearest similar atoll, the distance to the nearest large volcanic island ≥ 1000 km2 (here, the island of Tahiti), and the distance to the nearest raised atoll ≥ 15 m a.s.l. (here, Makatea or Henderson). Distances were measured with a Geographic Information System (GIS; Mapinfo® Professional version 10, WGS 1984 projection). Atoll area (i.e. surface of emerged lands), shoreline length and atoll elevation were obtained from the Atlas of Coral Reefs in French Polynesia (Andréfouët et al., 2005) and the United Nations Environment Programme (UNEP) island database (http://islands.unep.ch), although the elevations on the atolls were also obtained from various other sources.
There are several indices of isolation for islands (see Weigelt and Kreft, 2013). Here, we used the most commonly cited ‘index of isolation’ as defined by the UNEP (Daehler, 2006; Dahl, 1998) and calculated by
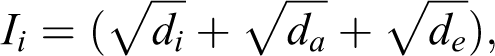
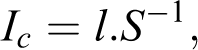
2 Native species richness on the atolls
The native species richness of vascular plants on the islands was recorded from different sources. On the islands of French Polynesia, the online database Nadeaud (Florence et al., 2007) and available botanical surveys (Butaud, 2009, 2013a,b; Butaud and Jacq, 2009) were used. These data were updated using the website of the National History Museum of Paris (2003–2014). Online biodiversity databases were used for the Cook Islands (McCormack, 2007) and Pitcairn Islands (Waldren and Kingston, 1998) together with reviewed literature (Florence et al., 1995; Kingston, 2001; Waldren et al., 1995).
According to their biogeographical status, native species observed on the atolls included (i) the indigenous species (i.e. native but not endemic) and (ii) the endemic species as mentioned by botanical databases, taxonomists and botanists working in Polynesia (e.g. Butaud, 2013b; Daehler, 2006; Florence, 1997; Florence et al., 1995, 2007; Imada, 2012; Kingston, 2001; Meyer et al., 2015; Wagner et al., 1990). Indigenous species means native to more than one archipelago while endemic species implies native to a single island/archipelago, but both terms are the result of only natural patterns and processes, i.e. with no species introduction. Here, native species richness included the indigenous and endemic species. Endemic plants compiled from the online Nadeaud database (Florence et al., 2007) identified four levels of endemism (as reported in Table 2). In this study, only strict endemic species observed on a single island or archipelago and called ‘insular endemic’ or ‘archipelagic endemic’ in Table 2 were counted as endemic.
List of the entire endemic species observed on the 49 atolls, focusing on endemic distribution on the coral atolls of Makatea, Niau, Anaa (Tuamotu Islands) and Henderson (Pitcairn Islands) where endemic species richness is the highest of the atolls surveyed (Butaud and Jacq, 2009; Florence et al., 1995; Florence et al., 2007; Waldren et al., 1995). Note that ‘French Polynesia endemic’ and ‘Eastern Polynesia endemic’, i.e. including the species that may be observed on more than one archipelago, were counted among the indigenous species in our analysis.

3 Statistical analysis
In order to test the independence between the factors, data were log10-transformed and multivariate analysis with Principal Component Analysis (PCA; XLStat® (version 2007.6) software) and Spearman’s rank correlation were used to present the inter-correlation of the eight factors used and the relationships between native species richness and factors. In addition, the influence of population density on the native species richness on the atolls was investigated as human impact may affect the analysis, even at low density. According to PCA results, linear regressions were then run to confirm which factor best explained the variation in native species richness (XLStat® (version 2007.6) software). Note that we also carried out stepwise multiple regressions in earlier work, found them lacking with regard to our botanical observations and, as an alternative, we used the results of the PCA and linear regressions, which appear more appropriate in the study area context.
IV Results
1 Native species richness on the atolls
On the atolls 120 native vascular plants were found, 21 of which were endemic. Indigenous and endemic species ranged from 3 to 71 and 0 to 9 species per island, respectively. Figure 2 illustrates the spatial pattern of native species richness found on the coral atolls surveyed. It shows a great native species richness on (i) the raised atolls of Makatea (77 native species) and Henderson (63 native species), and on (ii) the atolls close to the raised atoll of Makatea and the large volcanic island of Tahiti. Of the native plant species observed on the atolls surveyed, 58% were also found on the large volcanic island of Tahiti.
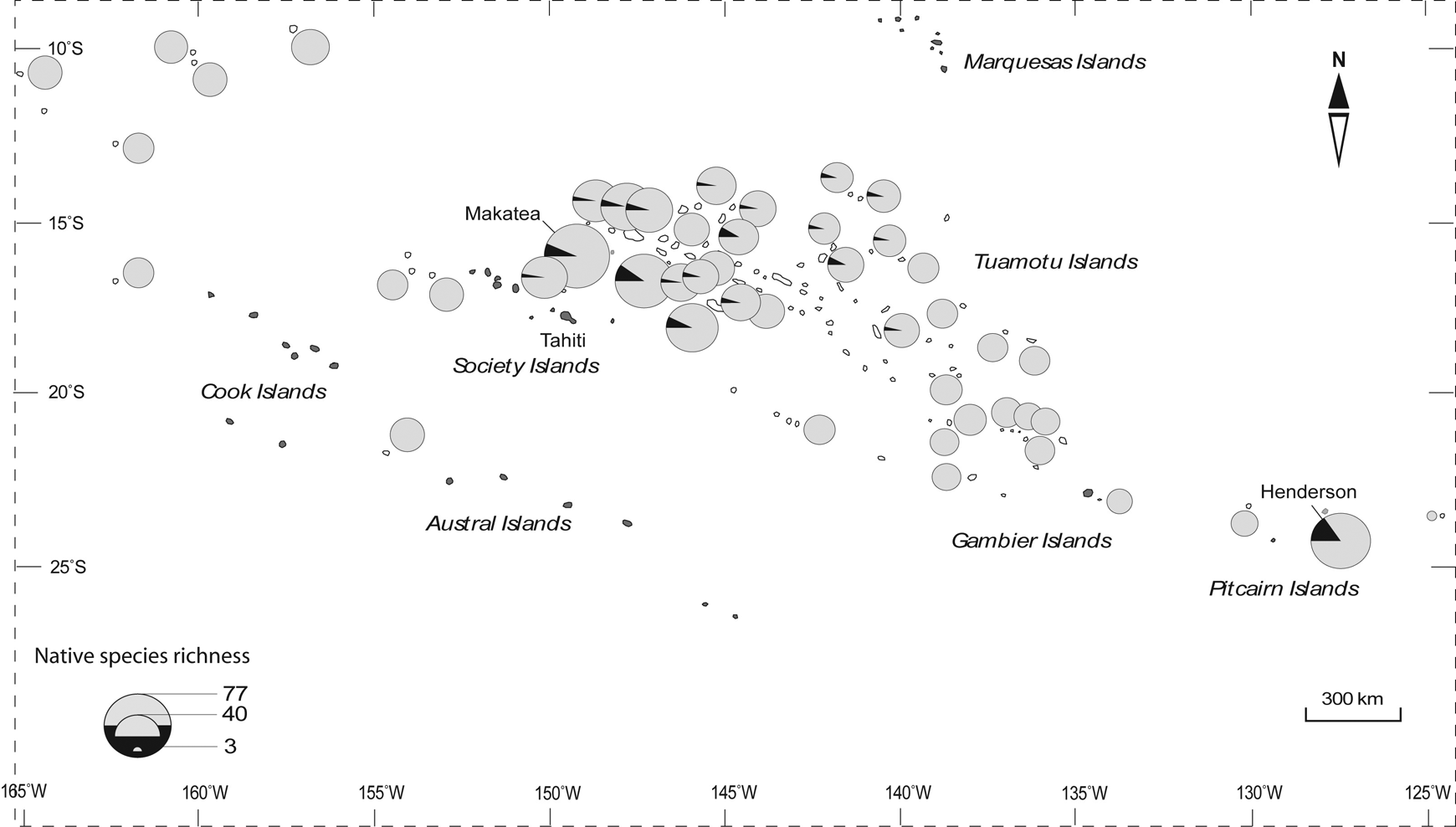
Spatial pattern of native species richness found on the 49 coral islands surveyed (Eastern Polynesia) with (i) indigenous species (gray part) and (ii) endemic species (black part).
The raised atoll of Makatea harbours 64% of native plant species observed on the atolls surveyed. Of the 10 endemic species observed in the Tuamotu Archipelago, five were listed on the raised atoll of Makatea (see Table 2). Five additional endemic species in the Tuamotu Archipelago were listed on the 5–6 m a.s.l. atolls of Anaa and Niau, respectively (see Table 2). With regard to the atolls in the Pitcairn Islands, 10 endemic species were only observed on the raised atoll of Henderson. No endemic species were listed on the atolls surveyed in the Cook Islands (see Figure 2).
2 Inter-correlation of the abiotic factors used
PCA provides a picture of the inter-correlation of the abiotic factors used and their correlation with native species (Figure 3). A relationship is observed between the distance to the raised atoll of Makatea and the distance to the volcanic island of Tahiti (r = 0.982, P < 0.01; Table 3) on account of the geographic proximity of these two islands in the study area (see Figure 1). Both distances to the raised atoll of Makatea and the volcanic island of Tahiti are negatively correlated with island area r = −0.615, P < 0.01 and r = −0.566, P < 0.01, respectively. A similar negative correlation is observed with the shoreline length (as atoll area and shoreline length are linked, r = 0.698, P < 0.01; Table 3). This shows that large atolls with a long shoreline tend to be close to the raised atoll of Makatea and the volcanic island of Tahiti.
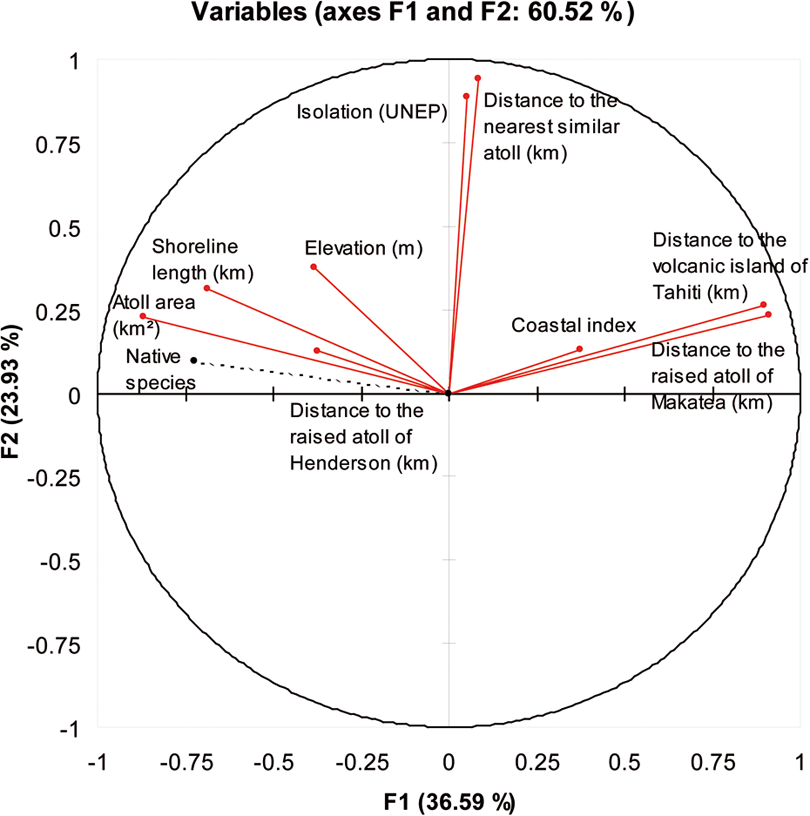
Visual result of inter-correlation of the abiotic factors used as predictors and their correlation with native species richness on the 49 atolls tested with Principal Component Analysis (PCA; XLStat® (version 2007.6) software). Native species richness (dashed line) has been added as a supplementary variable.
Inter-correlations of the abiotic factors used as predictors for native species richness (PCA, XLStat® (version 2007.6) software) on the 49 atolls surveyed in Eastern Polynesia (Pacific Ocean).
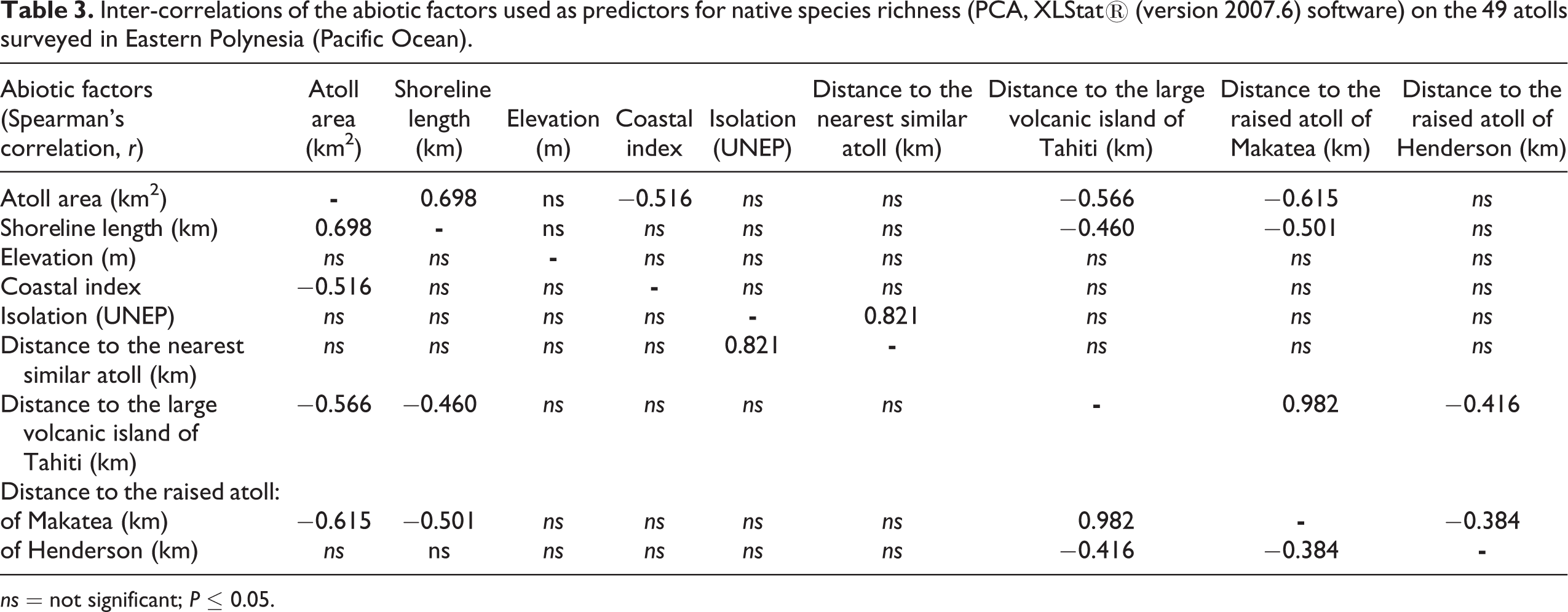
ns = not significant; P ≤ 0.05.
3 Relationships between abiotic factors and native species richness
Results with Spearman’s rank correlation and PCA
On the 49 atolls surveyed, native species richness was significantly related to atoll area (r = 0.732, P < 0.001), shoreline length (r = 0.557, P < 0.001), and elevation (r = 0.467, P < 0.001), respectively (Table 4). Native species richness was barely correlated with the coastal index (r = −0.289, P ≤ 0.05; Table 4). A strong negative correlation was observed between native species richness and distance both to the nearest raised atoll of Makatea and to the large volcanic island of Tahiti (r = −0.833, P < 0.001; r = −0.745, P < 0.001, respectively; Table 4). Indigenous species were strongly correlated with the distance to the raised atoll of Makatea (r = −0.833, P < 0.001; Table 4) and the distance to the volcanic island of Tahiti (rs = −0.742, P < 0.001; Table 4). Endemic species were significantly correlated with the distance to the raised atoll of Makatea (rs = −0.642, P < 0.001; Table 4) and the distance to the volcanic island of Tahiti (rs = −0.566, P < 0.001; Table 4). This confirms the hypothesis that the island of Tahiti and the raised atoll of Makatea strongly influence the spatial pattern of native species richness.
Correlations (Spearman’s rank correlation, XLStat® (version 2007.6) software) between factors and native species richness with endemic and indigenous species on the 49 atolls surveyed in Eastern Polynesia (Pacific Ocean).
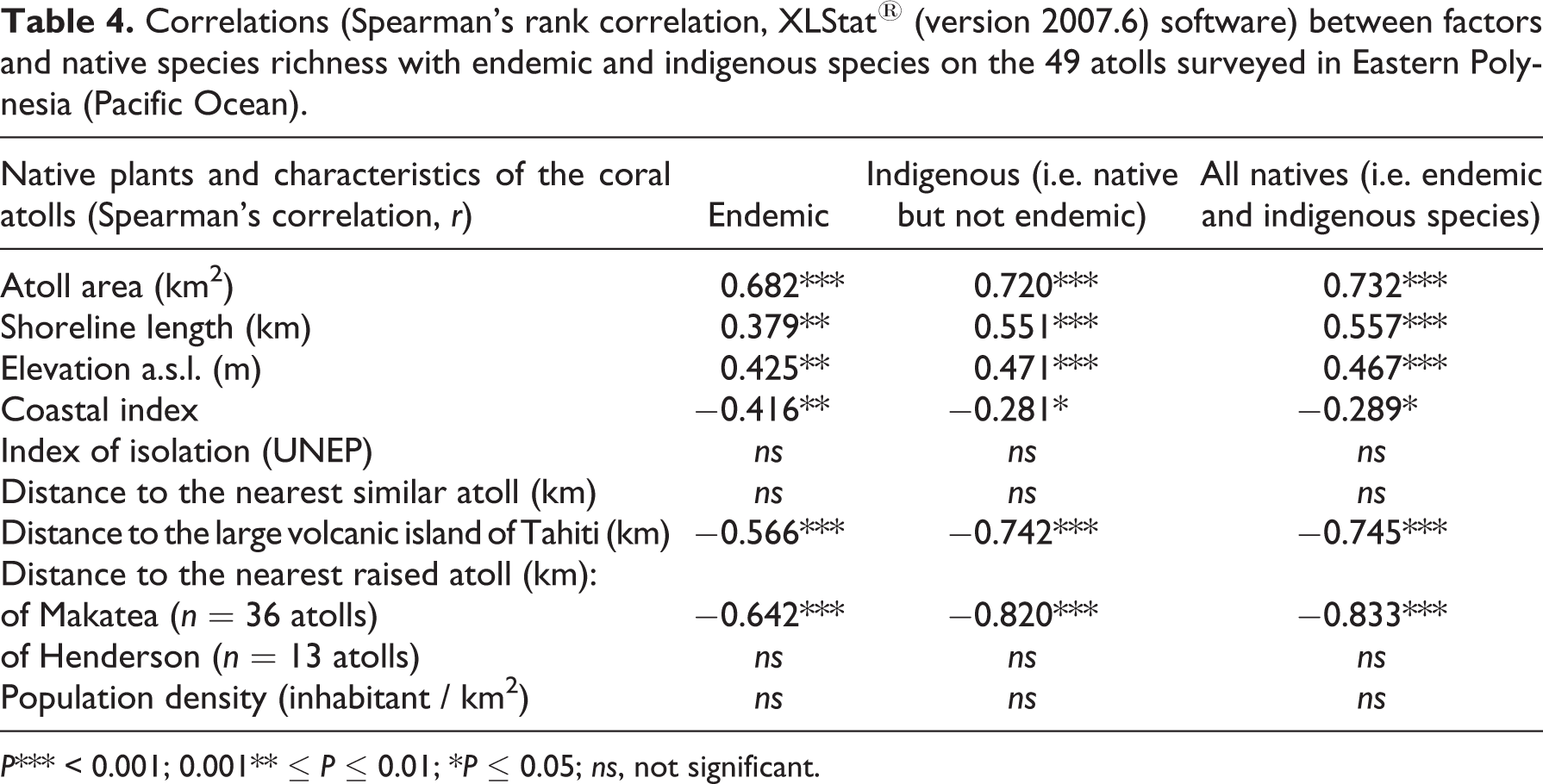
P*** < 0.001; 0.001** ≤ P ≤ 0.01; *P ≤ 0.05; ns, not significant.
No significant relationship was found between native species richness and distance to the nearest similar atoll, index of isolation, and distance to the nearest raised atoll of Henderson. No correlation was observed between native species richness and population density, suggesting only minor human impact on native species richness on the 49 atolls surveyed (Table 4).
The relationships between the native species richness and abiotic factors have been more closely examined by using a PCA. First, PCA showed that native species richness was strongly linked with the axis F1, F3 and F8 (Table 5). Second, the axis F1 explained a great part of the total inertia of the dataset (36.59%; see Figure 3) corresponding to the trace of the correlation matrix. This axis was led mostly by the distance to the raised atoll of Makatea (25.4%), the distance to the volcanic island of Tahiti (24.3%) and the atoll area (21%; Table 5). The axis F3 explained 14.8% of the total inertia; it was led by the coastal index (59%). The axis F8 (0.4% of the total inertia) was led by the atoll area (48%) and the shoreline length (27.6%). These findings confirm that the distance to the raised atoll of Makatea, the distance to the volcanic island of Tahiti, atoll area and coastal index were the most important explanatory variables. Shoreline length and elevation played a less important role. The axis F2 explained 23.93% (see Figure 3) of the total inertia but was poorly linked to native species richness (Table 5).
Relationships between the variables (here, the factors studied on the 49 atolls) and the decorrelated axes (F1 to F9) with PCA (XLStat® (version 2007.6) software). In order not to disrupt the dataset analysis, native species richness (i.e. the variable to explain) was added as a supplementary variable (for further information, see PCA with XLStat® software). Note that native species richness was strongly linked with the axis F1, F3 and F8.
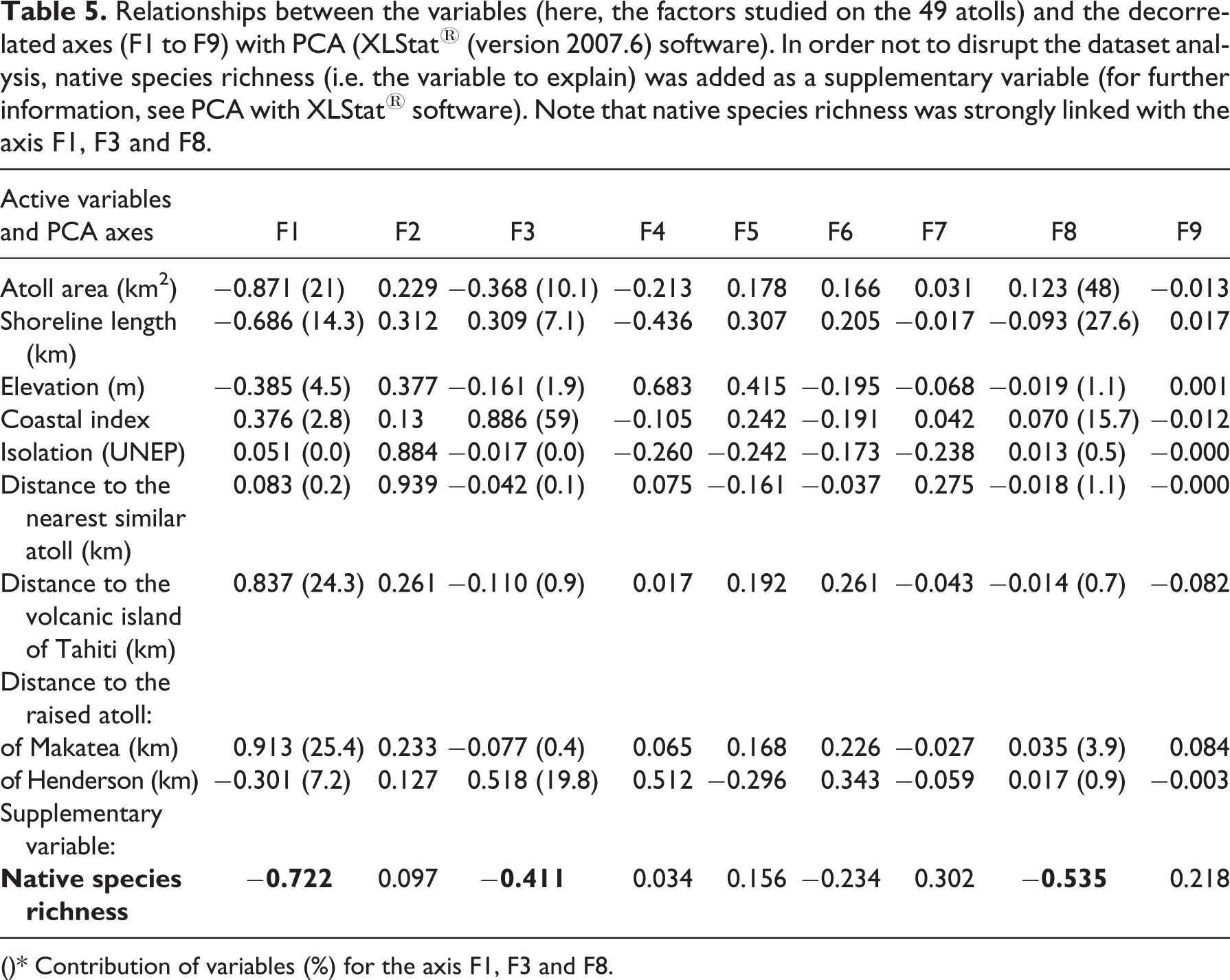
()* Contribution of variables (%) for the axis F1, F3 and F8.
Results with linear regressions
Linear regressions of log10-transformed species richness against factors showed that atoll area and distance to the raised atoll of Makatea explained 47.1% and 40%, respectively, of the species richness variation observed on the 49 atolls surveyed (Figure 4a–f). Of the variation 36.9% and 27.3% was explained by the distance to the volcanic island of Tahiti and the coastal index while elevation and shoreline length explained 23.3% and 18.4% of the variation, respectively (Figure 4a–f). The distance to the nearest similar atoll, isolation and the distance to the raised atoll of Henderson only explained 1.6%, 0.1% and 0.01%, respectively, of the native species richness variation (not shown in Figure 4a–f). These results were congruent with the PCA results.
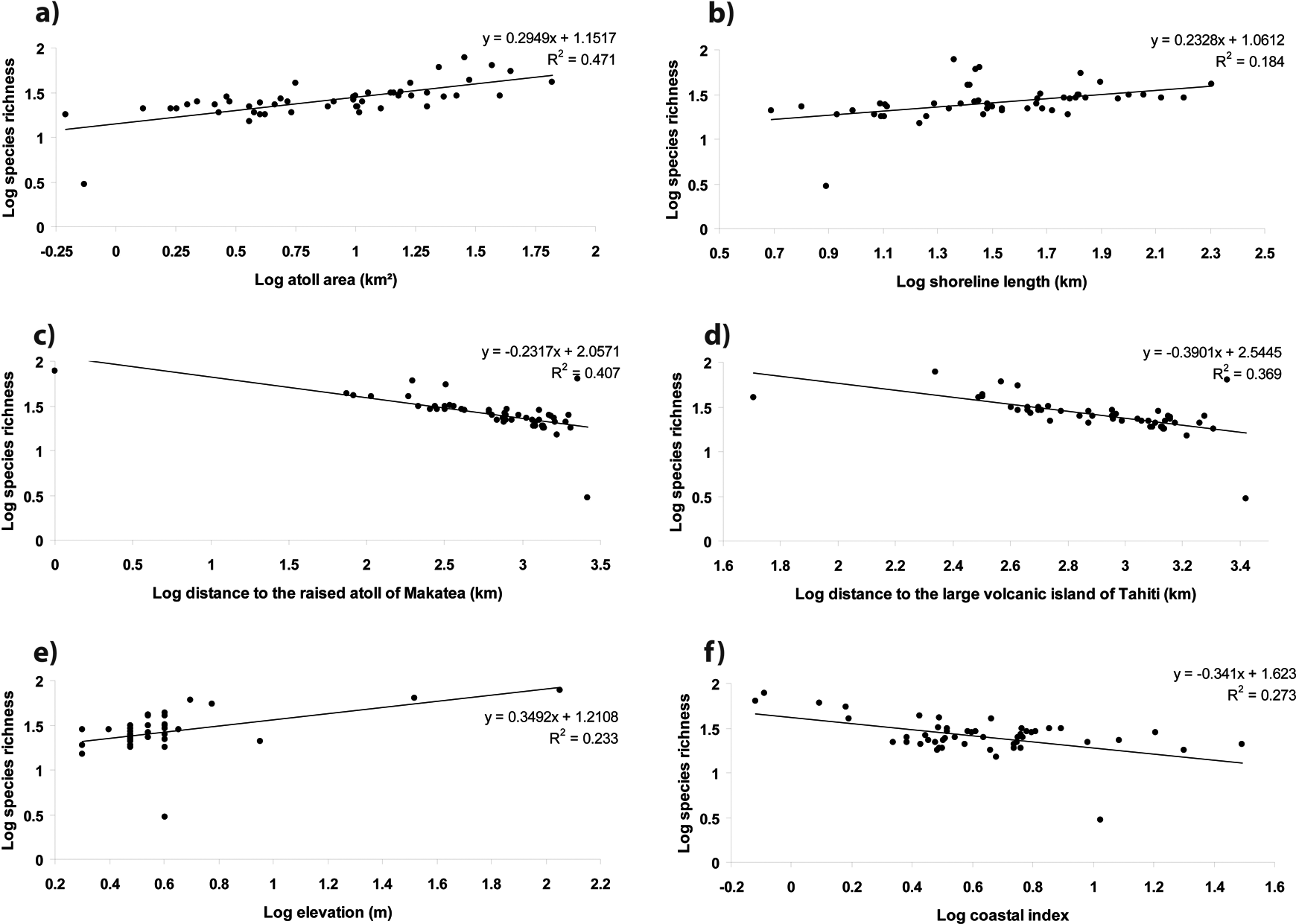
Linear regression of native species richness against significant factors on the 49 atolls surveyed in Eastern Polynesia with a) island area, b) shoreline length, c) distance to the raised atoll of Makatea, d) distance to the volcanic island of Tahiti, e) elevation and f) coastal index.
The inter-correlation between the distance to the raised atoll of Makatea and the volcanic island of Tahiti with the atoll area and shoreline length showed that the largest atolls tend to be close to Makatea and Tahiti, and conversely (see Figure 3 and Table 3). So, in order to confirm the influence of the raised atoll of Makatea and the volcanic island of Tahiti on species richness, a subsample of 13 pairs of atolls (i.e. 26 atolls among the 49 atolls surveyed) was extracted, individual pairs of atolls being similar in atoll area but presenting a maximum range of distance to Makatea and Tahiti. The distribution of native species richness according to similar atoll area at different mean distances to Tahiti and Makatea was compared (Figure 5). Despite the distribution in atoll area, Figure 5 supports that species richness on the neighbouring atolls of Makatea and Tahiti is greater than on those further away.
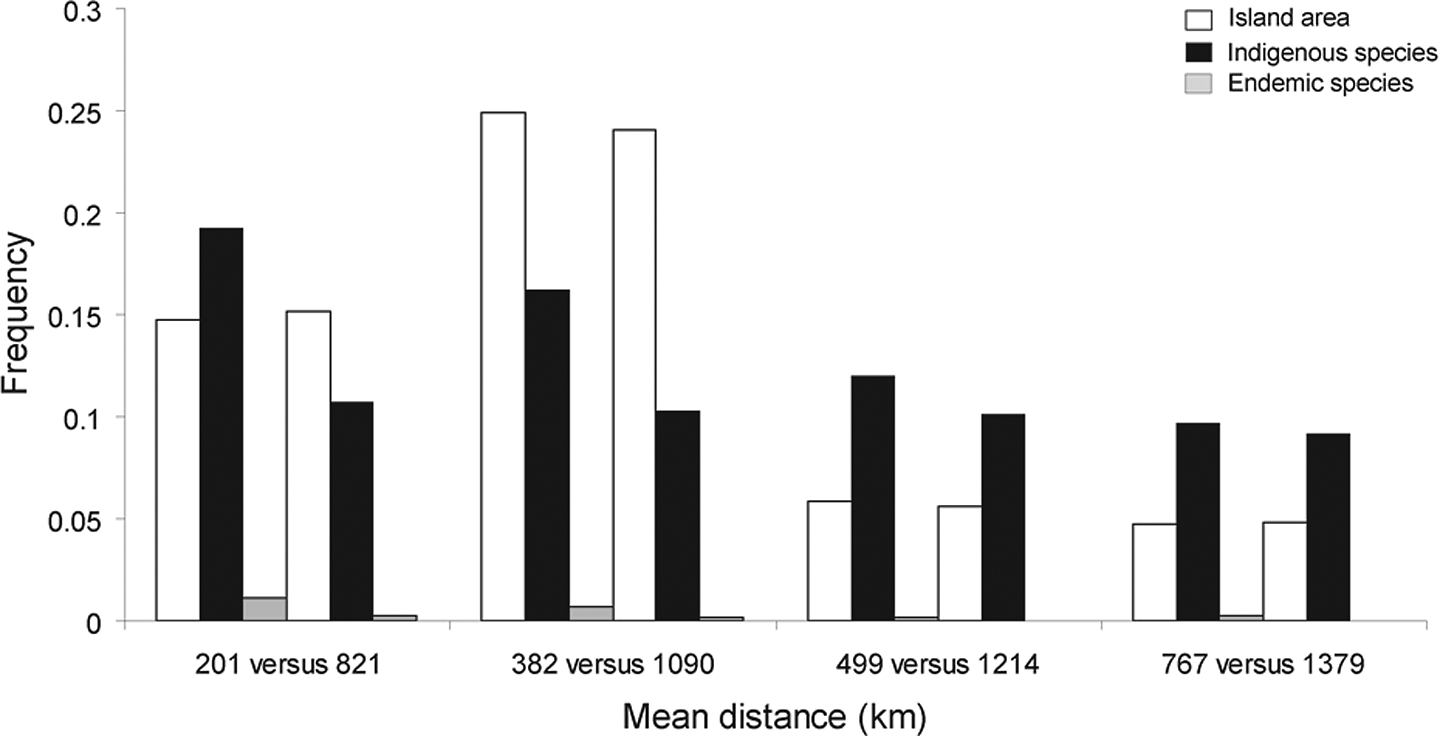
Distribution of similar island areas at different mean distances to the volcanic island of Tahiti and the raised atoll of Makatea compared with native species richness in the subsample of 26 atolls.
V Discussion
1 Native species richness and characteristics of atolls
In this study, native species richness was significantly related to atoll area and shoreline length (Table 4, Figure 4a and b). According to this result, the largest atolls with a long shoreline tend to contain more native species. First, this relationship could be explained by (i) the modality of native plant dispersion and (ii) the ‘island capacity’ to capture the species from oceanic dispersion. Seed dispersion from native plants mainly occurs via zoochory (birds) and hydrochory (ocean dispersal) in the Pacific coral islands (e.g. Florence et al., 1995; Fosberg, 1976; Mueller-Dombois, 2002). For example, native plant dispersion by bird zoochory and hydrochory represents 45% and 28%, respectively, on the atolls of Tuamotu (Florence, 2007). Thus, bird zoochory and hydrochory are important dispersal mechanisms for native plants on the atolls. There is no doubt that a large atoll area and a long shoreline (both are correlated, see Figure 3) harbour more chances to intercept species from ocean dispersal and provide more possibilities for the passage of birds because they are more likely to be seen than tiny and submersible atolls (see Lomolino, 1990, and the ‘target area’ hypothesis). In addition, the subsidized island biogeography hypothesis (SIB) suggests that spatial subsidies (i.e. material and resources on the island derived from outside the system) may increase the species richness of subsidized islands, depending on how the total productivity of the island is augmented by these subsidies (Anderson and Wait, 2001; Barrett et al., 2003). Here, large atolls with long shorelines might also capture more subsidies (e.g. allochthonous material from the mainland or another island) and this could influence the species richness observed on the atolls. Finally, large atolls have more habitats than smaller ones, with not only a coastal habitat but also a swampy area or ‘terrestrial’ habitat in the middle of the large islets where the influence of the salt and the wind is lower (Butaud, pers. obs.).
Second, extreme events, such as tsunamis and cyclones, are well documented in Eastern Polynesia (e.g. de Scally, 2008; Goff et al., 2011; Larrue and Chiron, 2010) and some authors have explained poor native plant richness on the small atolls of Suwarrow (1.7 km2, Cook Islands) and Ducie (0.74 km2, Pitcairn Islands) as being to the result of frequent marine submersion of these atolls during cyclones (Fosberg, 1991; Waldren et al., 1995; Woodroffe and Stoddart, 1992). Here, elevation was correlated with native species richness (see Table 4) supporting the idea that extreme events and their rise in the sea level can exert a long-term pressure on the native plant community established on small and low coral atolls. This result is in agreement with Fosberg (1976) who reported an increase in native species richness with the elevation, however small, of coral atolls of the Pacific Ocean. In the Cook Islands, cyclones have occurred on average every 3.8 to 8.8 years (de Scally, 2008) and every 13 to 39 years in the Tuamotu Islands (Larrue and Chiron, 2010). Thus, cyclonic swells could periodically devastate the smallest tropical atolls and this may control, at least partially, the number of native species established on these atolls. Additionally, native species richness was negatively correlated with coastal index (see Table 4). This supports the idea that large atolls offer more temporal plant stability than small atolls with a high coastal index and episodic instability, in which only a limited number of species can fit. We suggest that atoll vulnerability to marine submersion during cyclonic swells should be targeted as a hypothesis for future research.
Third, similar atoll areas were more frequently colonised by native species on the surrounding atolls of Makatea and Tahiti than on those further away (see Figure 5). This result supports that the SAR observed in this study was also influenced by the distance to the raised atoll of Makatea and the volcanic island of Tahiti.
2 Native species richness and index of isolation
Isolation often explains why remote islands support low native species richness, possibly owing to lower immigration rates and a poor rescue (MacArthur and Wilson, 1967 ; Whittaker and Fernández-Palacios, 2007). On the other hand, isolation effects (e.g. lower species immigration with increasing distance from the mainland) may be attenuated by many other factors such as a stepping-stone-island effect (MacArthur and Wilson, 1967) or inherent differences in plant dispersibility and abilities to colonise and maintain populations on islands (e.g. Daehler, 2006; Lomolino, 2000; Triantis et al., 2015).
Here, isolation was defined with the index of isolation (UNEP) and it was not significant to explain the native species richness observed on the 49 atolls surveyed, possibly because of the influence of different parameters. First, the scale of analysis focused on the atolls of Eastern Polynesia, which are all highly isolated. The mean value of the index of isolation was 107.5 with a standard deviation of 8.4 on the atolls surveyed (see Table 1). The range of isolation was not large and isolation remained high for each atoll surveyed, possibly making the sample less discriminating for this predictor. Second, the biota of more isolated islands tends to be derived from only small pools of colonists as a result of the ‘selecting effect’ imposed by distance (Herben, 2005). The native flora observed on the extreme remote islands of Eastern Polynesia is well known to be both impoverished and the easternmost limit of the range of a large number of genera in Pacific Ocean islands and Malaysia (Balgooy, 1993; Balgooy et al., 1996; Grant et al., 1974; Meyer, 2004; Stoddart, 1992). Thus, the pool of native species currently observed among the remote atolls of Eastern Polynesia may have been selected over a long period by plant dispersal capacity, remoteness and oceanic filter, resulting today in mostly widespread native plants in Eastern Polynesia despite its isolation. Finally, the geographic proximity of Tahiti and the raised atoll of Makatea may have tempered the isolation effect by increasing the flow of seeds and propagules in the study area and protecting native biota during sea-level highstands. In the study area, the distance to both these island types may be more relevant to characterise geographic isolation than the distance to the nearest mainland. This is congruent with Rosindell and Phillimore (2011) and Weigelt and Kreft (2013), who notably demonstrated that taking stepping stones into account was one of the important factors in increasing the explanatory power of isolation for endemism and species richness observed on the islands.
3 Native species richness and distance to the large volcanic island and raised atoll
The relationships observed between species richness and the distance to the volcanic island of Tahiti and the raised atoll of Makatea show that the number of native species decreases on the atolls with increasing distance to both these islands (see Table 4 and Figure 4c and d). This relationship may be explained by (1) a stepping-stone-island effect and (2) a refugium effect during the eustatic sea-level variations with highstands and lowstands in the Pleistocene and Holocene periods.
Stepping-stone-island effect
Native flora on the remote islands of Eastern Polynesia is relatively impoverished and mainly originates from the Indo-Malaysian Peninsula and Austro-Melanesian region located to the north-west (Balgooy et al., 1996; Fosberg, 1984; Meyer, 2004). It is commonly accepted that the decrease in native species richness from west to east on the Pacific Islands partly reflects the increasing distance of the remote islands from the mainland source pool of South-East Asia and the Western Pacific and the rainfall pattern across the Pacific Ocean from the wetter coral islands in the west to the drier ones in the east (Balgooy, 1993; Balgooy et al., 1996; Dahl, 1980, 1984; Manner, 1995; Mueller-Dombois, 2002; Stoddart, 1992). Here, we observed a negative correlation between the species richness on the atolls and the distance to Tahiti (see Table 4) and this factor explained 36.9% of the species richness variation (see Figure 4d). This suggests that the biggest and highest island of Tahiti influences the spatial pattern of native species on the atolls. The strongest correlation was observed with the indigenous species (rs = −0.742, P < 0.001; see Table 4), i.e. native plants with a wide distribution across the South Pacific Ocean. Additionally, 58% of the indigenous species observed on the atolls surveyed were also present on the volcanic island of Tahiti. The large volcanic island of Tahiti appears to be a source of indigenous colonists to the surrounding atolls and this result supports the ‘stepping-stone-island effect’ of Tahiti also observed on the remote volcanic islands of French Polynesia (Larrue, 2014).
Here, a correlation was also observed between the endemic species and the distance to Tahiti (rs = −0.566, P < 0.001; see Table 4). This suggests a possible allopatric speciation origin for the endemic species found on the atolls surveyed, with Tahiti as the island source of ancestor populations. Our finding is in agreement with Florence (2007) who reported that the Tuamotu Islands presented 78.5% of native floristic affinity with the Society Islands. Indeed, many endemic species found on the atolls surveyed belong to botanical genera also observed on the volcanic island of Tahiti (e.g. Meryta, Psychotria, Xylosma, Glochidion, Ixora, Pandanus, Geniostoma, Alyxia and Peperomia). This leads us to consider the volcanic island of Tahiti as a provider of colonists for immigration and speciation on the atolls, although only phylogeographic studies using molecular tools will clearly answer the question of the origin of these plant species.
Raised atoll as a refugium
The raised atoll of Makatea contains 64% of the native plants found on the atolls surveyed and the distance to this raised atoll explains 40% of the variation observed in species richness. Thus, the raised atoll of Makatea plays an important role in the spatial pattern of species richness in the study area, especially for the indigenous species (r = −0.833, P < 0.001; see Table 4). Endemic species were significantly linked with the distance to the raised atoll of Makatea (rs = −0.642, P < 0.001; see Table 4), but some genera found on the atolls of Henderson (33 m a.s.l.), Niau (6 m a.s.l.) and Anaa (5 m a.s.l.) (e.g. Meryta, Sesbania, Santalum and Psychotria; see Table 2) were not observed on Makatea (111 m a.s.l), despite similar habitats. This suggests that speciation originated from ancestors of islands other than Makatea and notably from the high volcanic island of Tahiti, especially for some genera with ‘no coastal’ affinity in Eastern Polynesia (e.g. Meryta, Santalum or Psychotria). Thus, a relationship was observed between endemic species and the raised atoll of Makatea, but the reason for this relationship may be explained differently from that of the volcanic island of Tahiti.
It is documented that during the Pleistocene and Holocene periods, glacial–interglacial cycles led to large eustatic sea-level variations with sea-level highstands and lowstands on the islands (e.g. Camoin et al., 2001; Dickinson, 2001; Hewitt, 2000; Kaufman, 1985). In circa 125 ka and 9 ka, the sea level was between 6 and 10 m and reached 23 m above the present sea level (a.p.s.l.), respectively (Camoin et al., 2001), while from 6000–5500 yr b.p. until at least 1200 yr b.p. the mean sea level was around 0.9 m a.p.s.l. (Pirazzoli and Montaggioni, 1986). During the highest sea levels, the raised atoll of Makatea was fully emerged (Cibois et al., 2010), while the raised atoll of Henderson may not have been completely submerged. Because of the long-term stability of emerged land, both raised atolls of Makatea and Henderson contain high native species richness (77 and 63 native species, respectively; see Figure 2) with five endemic species on Makatea, including the shrub Myrsine ovalis var. wilderi F.R. Fosberg & M.-H. Sachet [Primulaceae], the tree Homalium mouo H. St John [Salicaceae] and a new subshrub Scrophulariaceae sp. nov. (Table 2). After the decline in sea level, our findings suggest that indigenous and endemic plants were able to colonise the emerged atolls mainly from the raised atoll of Makatea (Table 4).
The raised atoll of Henderson has 63 native species of vascular plants, 10 of which are endemic (see Table 2), but the neighbouring small atolls of Ducie and Oeno (Pitcairn Islands) have only a few indigenous and no endemic species (see Figure 2) suggesting a low species colonisation from Henderson. However, St John and Philipson (1960) observed the endemic herb Bidens hendersonensis var. oenoensis Sherff [Asteraceae] on the atoll of Oeno, but this is now considered extinct (Waldren et al., 1995). In fact, the atolls of Oeno and Ducie are very small (0.62 and 0.74 km2, respectively) with a short shoreline and an elevation of 4 m a.s.l. Thus, no relationship was observed between species richness and distance to the raised atoll of Henderson, probably on account of its geographic context (see Figure 1), i.e. surrounded by only a few atolls providing poor habitat diversity, high vulnerability during cyclones and a weak capacity to rescue species from oceanic dispersion and maintain populations. Therefore, only the raised atoll of Makatea appears to be a source pool for plant dispersion to the surrounding atolls. This leads us to consider the raised atoll of Makatea a refugium, which is in agreement with Cibois et al. (2010) who used genetic and Bayesian tools to demonstrate that the atoll of Makatea played the role of an ‘ecological refugium’ for endemic land birds in the Tuamotu Islands during the last interglacial maximum and its highest sea level.
VI Conclusion
In this study, native species richness is strongly related to atoll area and this is congruent with many island biogeography studies (e.g. Ackerman et al., 2007; Chown et al., 1998; Diamond and Mayr, 1976; Price, 2004). Less important relationships were also observed between native species richness and coastal index, elevation and shoreline length. This suggests that atoll vulnerability to marine submersion during cyclonic swells should be targeted in future research as a hypothesis to explain variation in species richness.
Above all, the distance to the raised atoll of Makatea and the distance to the volcanic island of Tahiti were strongly significant in predicting native species richness. This confirms the hypothesis that both these island types are important factors to explain the spatial pattern of native species richness observed on the remote atolls. Our findings suggest that the volcanic island of Tahiti and the raised atoll of Makatea can be viewed as stepping-stone-island and refugium, respectively, for native species on the atolls. Furthermore, no correlation between native species richness and isolation was found in the study area, possibly as a result of both the stepping-stone-island effect of Tahiti and the proximity of the raised atoll of Makatea as a refugium.
Footnotes
Acknowledgements
We thank Roger Oyono (University of French Polynesia, Tahiti) for his logistic support during the mission 2013 and 2014 in French Polynesia and the two reviewers for their comments to improve the paper.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: We are grateful to the ‘Fonds Pacifique’ (Ministère des Affaires étrangères, Paris, France) and the ‘Ministère de l’écologie, du développement durable et de l’énergie’ (Paris, France) for their financial support for the missions in French Polynesia.