
Abstract
Background:
Rotator cuff tear is a prevalent musculoskeletal disorder with a high postoperative retear rate. Transcortical vessels (TCVs) are capillaries that cross cortical bone and represent key elements of bone microcirculation. However, the possible intervention and influence of TCVs for rotator cuff repair (RCR) have rarely been studied.
Purpose:
To investigate the effect of TCV-mediated revascularization in RCR and the mechanism of potential intervention.
Study Design:
Controlled laboratory study.
Methods:
Sprague-Dawley rats (N = 135) with supraspinatus tendon tear and delayed repair were randomly assigned into 3 groups based on footprint preparation: (1) cortical bone freshening up by debridement to expose cancellous bone, (2) cortical bone preservation as the control group, and (3) cortical bone preservation with oral D-sphingosine as a TCV-stimulating factor. Oxidative stress was measured using fluorescent probes; hypoxia and inflammation levels were measured via immunosorbent assay; and RCR and TCVs were evaluated with biomechanical and immunostaining assays.
Results:
Cortical bone preservation inhibited the revascularization of TCVs during RCR (P < .01), while freshening up or D-sphingosine treatment promoted TCV formation by reducing oxidative stress (P < .05 for all). Revascularization of TCVs alleviated hypoxia and inflammation-derived chronic tendinopathy after 6 weeks (P < .05 for all) and improved the biomechanical properties of rotator cuff at 12 weeks (P < .05).
Conclusion:
Footprint freshening up or D-sphingosine treatment improves tendon-to-bone healing, suggesting the importance of TCV-mediated revascularization for RCR.
Clinical Relevance:
Revascularization therapy such as cortical bone freshening up or D-sphingosine has the potential to improve RCR.
Rotator cuff tears are one of the most common musculoskeletal disorders. More than 270,000 rotator cuff repair (RCR) surgical procedures are performed each year in the United States, with the total annual costs exceeding $474 million.7,25,30 The healing rates after RCR remain lower than desired despite improved surgical techniques and implants. The uncertain success rate of RCR poses a great challenge and risk for surgeons, partially explaining the persistent shoulder pain and limited mobility for patients. 6 As a disease of the enthesis, rotator cuff injuries frequently occur at the insertion site, where the tendon anchors to the bone surface by parallel fibers and inserts into the tendon-to-bone interface (TBI).
Multiple factors could lead to the pathogenesis of TBI disorder, including intrinsic degeneration and genetic predisposition, extrinsic overhead exercise and subacromial impingement, acquired metabolic disorders, and chronic inflammation.3,5,10,12,19,29 Given that the regenerative ability of TBI is quite limited and the functional reconstruction of the multilayer TBI structure is hard to achieve, inferior postoperative healing of TBI is an important cause of rotator cuff retear.1,18 Indeed, most postoperative healing at the TBI site results in scar tissue, which hardly withstands the high mechanical loads and cycles of shoulder mobility.19,20,29,32 Therefore, it is essential to enhance the limited regenerative ability of TBI and improve its healing quality.
Transcortical vessels (TCVs) are blood capillaries that originate from the bone marrow and traverse the cortical bone toward the periosteal surface. 8 They extend longitudinally along the long bone diaphysis and cross the cortical bone perpendicularly, building a direct vascular bridge between the medullary circulation and the outer bone surface.14,26 There may be some functional vascular communication between the bone marrow vascular system and the external attachment circulation.8,9,11,24 We hypothesized that the main blood circulation at the distal end of the rotator cuff originated from TCVs on the TBI side and thus applied D-sphingosine as a positive rescuing factor to activate TCV formation while preserving the cortical bone of the footprint. 14
In this study, we employed a chronic rat RCR model and performed cortical bone debridement for footprint freshening to directly facilitate vascular communication between the bone marrow and the external tendon, as well as to test its healing quality on the TBI and rotator cuff. We aimed to investigate whether postoperative revascularization was related to the functional healing of the rotator cuff, whether the cortical bone preservation at the footprint would impair RCR by impeding TCV formation, and whether D-sphingosine had potential for use in RCR therapy by promoting TCVs.
Methods
Animal Model and Surgical Procedures
All of the operations on animals strictly conformed to the guidelines of the National Research Council's Guide for the Care and Use of Laboratory Animals. The animal experimental protocols were approved by the Animal Welfare Ethics Committee of Tongji University affiliated with the Shanghai Tongji Hospital (2024-DW-SB-036). A total of 135 Sprague-Dawley rats (8 weeks old, male, 280-320 g), which were purchased from the Shanghai Laboratory Animal Research Center, were included in the study for the RCR model. We randomly divided the rats into 3 groups based on the footprint preparation process: bone surface freshening (noncortical bone), cortical bone preservation without treatment (cortical bone), and cortical bone preservation with oral D-sphingosine. Briefly, we randomly chose a unilateral shoulder joint and made a skin incision 2 cm wide, then split the deltoid muscle longitudinally to expose the acromion; after which, we separated the underlying supraspinatus tendon from the subacromial bursa by blunt dissection and toward the distal end to visualize the whole rotator cuff and the greater tuberosity of the humerus. Then, we cut off the supraspinatus tendon at the bone insertion of the greater tuberosity and used 2 nonabsorbable 5-0 sutures to pass through the supraspinatus tendon by mattress suture. We cleaned the greater tuberosity surface and removed all soft tissues to expose the footprint area, then drilled a bone tunnel into the greater tuberosity by a surgical needle. For the cortical bone preservation control, we stitched the nonabsorbable sutures through the bone tunnel and fixed the distal end of the supraspinatus tendon onto the footprint area while preserving fibrocartilage and cortical bone. For the bone surface freshening, we debrided the bone surface using a commercial electrical dental drill with a spherical abrasion head (Saeshin) until bleeding from cancellous bone was observed (Figure 1A); we then preserved the local circulation and functional vascular tissue by intermittent abrasion and saline cooling. We subsequently fixed the supraspinatus tendon as previously described and performed an incision suture. D-sphingosine administration was performed by oral gavage (2 mg/kg) every day for 2 weeks in accordance with the methods of a previous study. 14
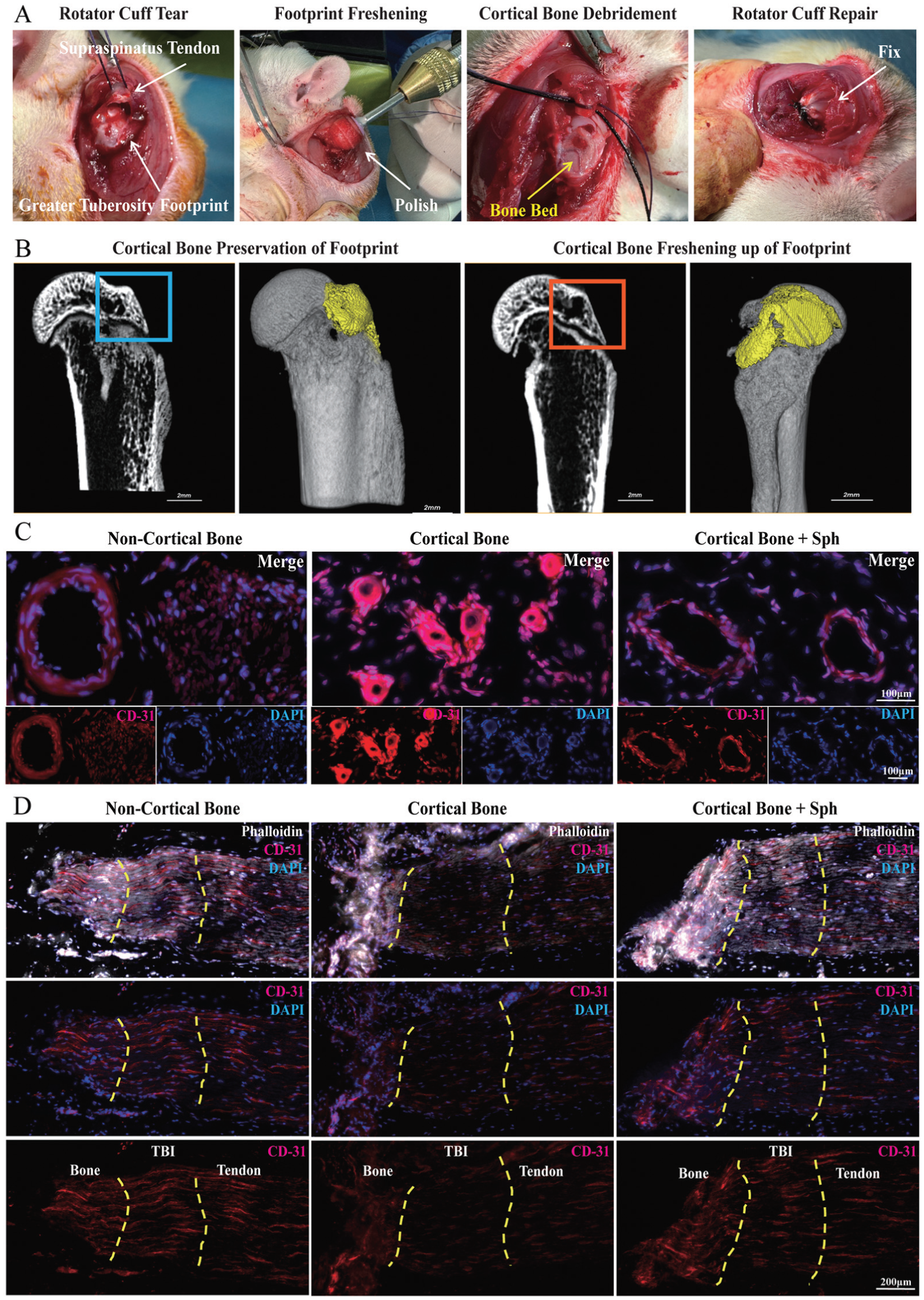
Operation at the footprint cortical bone during rotator cuff repair surgery and corresponding images of transcortical vessels at tendon-to-bone interface (TBI). (A) Intraoperative illustration of rotator cuff repair surgical procedures and the freshening up on the footprint cortical bone. (B) Postoperative micro–computed tomography images of the proximal humerus on a longitudinal section and 3-dimensional reconstruction. The attachment location (ie, the humeral greater tuberosity) is highlighted by line boxes, while the attachment surface (ie, the footprint area) is marked by a pseudo-color. Scale bar, 2 mm. (C) Morphology of transcortical vessels on a cross section of the TBI presented by CD31 immunofluorescence staining. Scale bar, 100 μm. (D) Distribution of transcortical vessels on a longitudinal section of the TBI, counterstained by phalloidin and 4',6-diamidino-2-phenylindole (DAPI). Yellow dashed lines indicate artificial boundaries of bone insertion and distal tendon for the TBI reference. Scale bar, 200 μm.
All of the included rats were housed in a specific pathogen–free animal facility with regular light-and-dark cycles, suitable humidity and temperature, and sufficient standard diet and water. During surgery, the rats were anesthetized with 2.5% inhalational isoflurane delivered via nose cone and received a subcutaneous injection of buprenorphine (0.05 mg/kg, 12-hour intervals for 3 days) to achieve postoperative analgesia. After surgery, they were given enough room without activity restriction and penicillin for infection prevention. Euthanasia was achieved through an excessive anesthesia method at 3, 6, and 12 weeks postoperatively to collect rotator cuff samples (Figure 4A).
Micro–Computed Tomography
We collected rat humeral samples and fully immersed them in 4% paraformaldehyde for >48 hours. Micro–computed tomography (micro-CT) scanning (NMC-200; NEMO) was performed along the long axis of bone with the following parameters: source voltage, 80 kV; source current, 0.06 mA; and resolution ratio, 35 μm. The region of interest (ROI) encompassed the footprint area of the greater tuberosity in the proximal humerus. Three-dimensional reconstructions were achieved by stacking original images of the ROI in Recon software. A square-shaped part of the ROI covering the footprint surface and TBI depth was selected to calculate bone volume/total volume fraction, trabecular bone computed tomography value, bone mineral density, and cortical thickness.
Histological and Reactive Oxygen Species Staining
Histological staining was performed to assess the tissue morphology and characteristics of the TBI and distal tendon at 3, 6, and 12 weeks postoperatively. Specifically, the supraspinatus-humerus samples were fixed with 4% paraformaldehyde, subjected to decalcification by 10% EDTA for >21 days, and embedded in paraffin wax. The samples were finally sectioned along the longitudinal axis of the bone and tendon into 5-μm-thick slices. Hematoxylin and eosin staining, toluidine blue staining, and Masson trichrome staining were conducted in line with the manufacturers’ instructions. Immunohistochemical staining for collagen I and alpha–smooth muscle actin (α-SMA) was performed as previously reported. Briefly, the sections were subjected to dewaxing in xylene, rehydration in gradient alcohols, and repeated washing. They were incubated with 3% hydrogen peroxide for 10 minutes. Next, they were immersed into 80°C sodium citrate buffer for 30 minutes away from light. After permeabilization by 0.1% Triton X-100, the sections were marked by the primary antibody, collagen I (ab254113; Abcam) and α-SMA (ab7817; Abcam); subsequently stained by horseradish peroxidase–labeled secondary antibody (ab6721 and ab97245; Abcam); and finally observed by a Slide Scanner (VS200; OLYMPUS). The semiquantitative analyses of histological staining and immunohistochemistry staining were performed in ImageJ software.
Reactive oxygen species (ROS) staining was performed in frozen tissue sections by an ROS fluorescence probe, 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA; D6883 [Sigma Aldrich]), in accordance with the manufacturer's instructions. Briefly, fresh supraspinatus-humerus samples were washed by cold saline, immersed in liquid nitrogen, and embedded with optimal cutting temperature compound. Next, they were cut into 5-μm-thick sections by freezing microtome. After drying, the sections were stained with DCFH-DA and DAPI (MBD0015; Sigma Aldrich) and visualized by a fluorescence microscope (Eclipse C1; Nikon). The semiquantitative analyses of ROS staining were normalized to sham operation by comparing the DCFH-DA fluorescence intensity of the unit area.
Immunofluorescence Staining
For bone and tendon immunofluorescence staining, supraspinatus-humerus samples were harvested. After removal of the external synovial membrane or periosteum by blunt dissection, they were fixed with 4% paraformaldehyde for 48 hours and subjected to decalcification by 10% EDTA for >21 days. Next, they were embedded in paraffin wax and cut into 5-μm-thick slices transversally or along the longitudinal axis. After dewaxing by absolute xylene and rehydration through gradient alcohols, the sections were permeabilized by 0.1% Triton X-100 buffer and blocked by 3% bovine serum albumin for 1 hour. For labeling capillary vessels crossing bone tissues, the sections were incubated with primary antibody, anti-CD31 (1:200, ab222783; Abcam), for 24 hours at 4°C. After washing, the sections were incubated with secondary antibody, namely goat anti-rabbit IgG H&L conjugated with Alexa Fluor 647 (1:200, ab150079; Abcam), and rhodamine phalloidin (1:200, R415; Thermo Fisher) for 12 hours without light. After washing, the sections were treated with an antifade mounting medium containing DAPI (P36966; Thermo Fisher). Staining images were acquired by a confocal laser scanning microscope (LSM 880; Zeiss). The immunofluorescence quantification of vessels, including cross-sectional area, longitudinal-sectional diameter, and branch number, was performed in ImageJ software.
Biomechanical Testing
We harvested the humerus–rotator cuff complex from the shoulder blade, carefully removed other irrelevant rotator cuff tendons and unnecessary joint capsule, and precisely preserved the supraspinatus tendon connected with corresponding muscle and greater tuberosity attachment. We kept the sample fresh and moist by saline solution while performing the following test. The cross-sectional area of the distal tendon was measured by a digital caliper from 3 sites. The elastic displacement of the supraspinatus tendon, maximal rupture load, and stress of the bone-tendon connection were measured by a biomechanical testing machine (Electroforce 3520-AT; BOSE). Briefly, the distal humerus was fixed in the clamp of the testing machine, and the proximal tendon-muscle was fixed in another clamp. The tensile force was applied along the longitudinal axis of tendon-bone, starting with a static preload (1 N, 3 minutes) and then steadily increasing to the ultimate failure load with the loading rate of 3 mm/min. The final rupture load and elastic displacement were recorded from the displacement diagram, and the maximal rupture stress was calculated from the final maximal load and cross-sectional area.
Enzyme-Linked Immunosorbent Assay
The sphingosine-1-phosphate (S1P) levels in rotator cuff samples were detected by a commercial enzyme-linked immunosorbent assay kit (EU2603; Fine Biotech). Briefly, the competitive microtiter plate was precoated with S1P; 50 μL of the supernatant from homogenized cell or tissue samples was added into the plate and mixed for 1 minute; and 50 μL of biotin-labeled S1P antibody solution was added in line with the manufacturer's protocol. After incubation for 1 hour and washing twice, 100 μL of horseradish peroxidase–streptavidin working solution was added, incubated for 30 minutes, and washed. Then, 90 μL of Tetramethylbenzidine substrate was added for 10 minutes. Finally, 100 μL of stop solution was added, and the absorbance at 450 nm was detected by the microplate reader. The hypoxia markers (HIF-1α, TGF-β1), inflammation markers (TNF-α, IL-1β), fibroblast activation markers (CD106, CD248), and fibrosis development markers (α-SMA, COL I/COL III ratio) of the distal supraspinatus tendon tissues were also detected by commercial enzyme-linked immunosorbent assay kits in accordance with the manufacturer's instructions (ER0191, ER1378, ER1393, ER1094, ER1358, ER0848, ER0853 [Fine Biotech]; MB7034B [Meibiao Biotech]; A108373 [Fusheng Industrial]).
Statistical Analysis
Each group required 3 types of tissue samples for different assays. The sample size was 5 rats per sample type at each of the 3 time points. Data were analyzed and presented as the mean with standard deviation (SD) through Prism software (Version 9.1.1; GraphPad). Since the rotator cuff disorders were small and discontinuous with a variable and patchy character, we assumed that the animal data did not comply with normal distribution. Thus, we used nonparametric tests for statistical analyses. The comparisons of multiple individual datasets were performed by the Kruskal-Wallis test with Dunn post hoc test. P < .05 was considered statistically significant.
Results
Direct and Micro-CT Observation
The original footprint was thick and compact, consisting of multiple calcified bone layers. After cortical bone debridement, the freshened footprint was thinner, with the underlying cancellous bone exposed (Figure 1B). Micro-CT analysis (Figure 3A) showed that footprint freshening markedly decreased the calcified bone mass and cortical bone thickness in the footprint area as compared with the cortical bone preservation control (P < .05). In contrast, D-sphingosine with footprint preservation did not significantly affect the cortical bone mass at the footprint (P > .05).
Histological and ROS Staining
For histological staining of the TBI, the Sharpey fibers and fibrocartilage were disrupted after freshening up and had not recovered at 3 weeks; however, at 12 weeks, fibrocartilage and Sharpey fibers increased notably (Figure 2A). In the preservation group, fibrocartilage decreased markedly at 12 weeks (Figure 2B), and Sharpey fibers became indistinguishable. D-sphingosine treatment with footprint preservation prevented this degeneration of the TBI, exhibiting enhanced Sharpey fibers and fibrocartilage at 12 weeks (Figure 3G). In this supraspinatus tendon, moderate differences in collagen fiber organization and collagen I content were observed between the footprint freshening and preservation groups at 3 weeks (Figure 2C). After 12 weeks, the collagen fiber ratio and collagen I content decreased significantly in the preservation group (Figure 2D), while D-sphingosine treatment and footprint freshening maintained a stable rate of increase in tendon collagen at 12 weeks (Figure 3H).
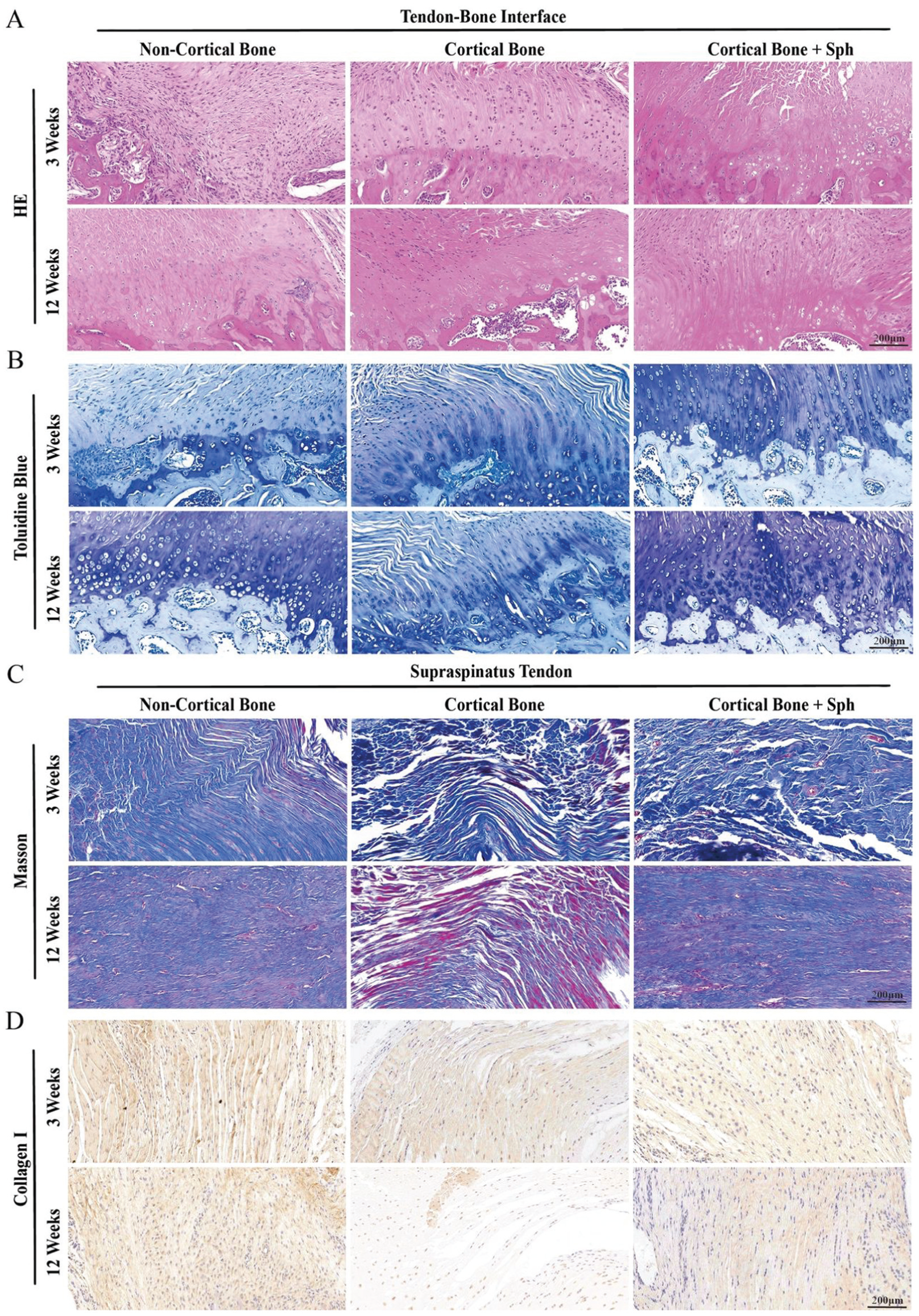
Representative postoperative images of tendon-to-bone interface (TBI) and distal supraspinatus tendon in a rat rotator cuff repair model. (A) Hematoxylin and eosin (HE) staining of TBI. (B) Toluidine blue staining of TBI. (C) Masson trichrome staining of the supraspinatus tendon. (D) Collagen I immunohistochemical staining of the supraspinatus tendon. Scale bar, 200 μm. Sph, D-sphingosine.
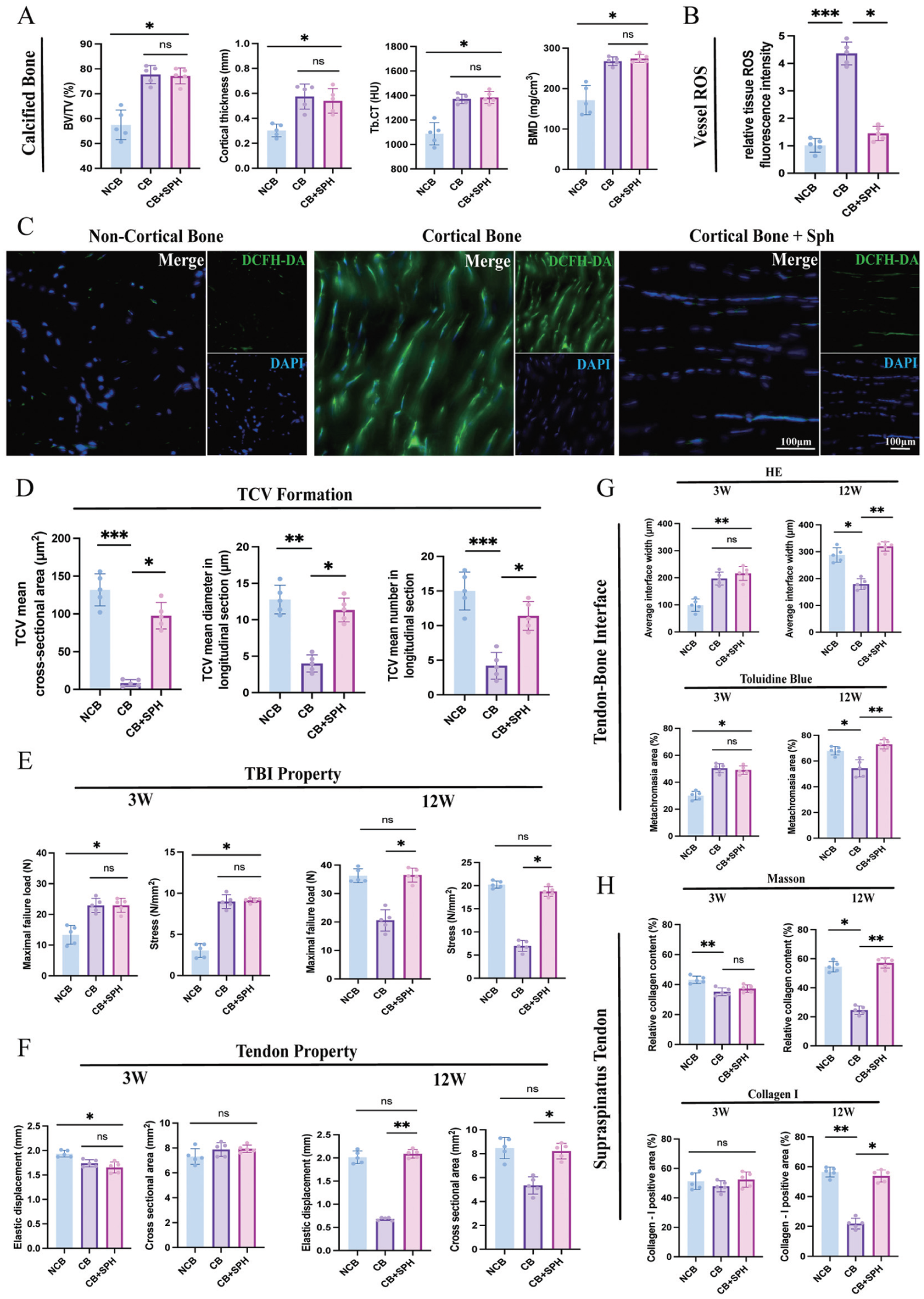
Quantitative analysis of cortical bone mass, oxidative stress level in vessels, transcortical vessel (TCV) formation, and biomechanical performance, as well as histological evaluation of tendon-to-bone interface (TBI) and tendon. (A) Quantitative micro–computed tomography analysis of calcified bone within TBI after the cortical bone freshening up. (B) Quantitative analysis of reactive oxygen species (ROS) normalized to the noncortical bone (NCB) group, determined by ROS fluorescence probe from 3 random fields in a frozen tissue section. (C) ROS accumulation within vessels at TBI; scale bar, 100 μm. (D) Quantitative analysis of TCV formation at the TBI based on immunofluorescence staining for TCVs. (E) Quantitative biomechanical testing of tendon-bone complex (maximum load and stress) at 3 and 12 weeks postoperatively. (F) Quantitative biomechanical testing of the supraspinatus tendon (elastic displacement and cross-sectional area) at 3 and 12 weeks postoperatively. (G) Analysis of the interface width and metachromasia ratio of TBI by hematoxylin and eosin (HE) and toluidine blue staining. (H) Analysis of total collagen content and type I collagen ratio of the supraspinatus tendon by Masson trichrome and immunohistochemical staining. Ns, not significant. *P < .05. **P < .01. ***P < .001. Kruskal-Wallis and Dunn multiple-comparison test. Data are presented as mean ± SD (n = 5). BMD, bone mineral density; BV/TV, bone volume/total volume fraction; CB, cortical bone; DAPI, 4',6-diamidino-2-phenylindole; DCFH-DA, 2′,7′-dichlorodihydrofluorescein diacetate; SPH, D-sphingosine; Tb.CT, trabecular bone computed tomography.
ROS fluorescence showed the revascularization of TCVs through cortical bone accompanied by ROS generation (Figure 3B). Footprint freshening and D-sphingosine treatment significantly alleviated ROS accumulation within vessels (P < .001 for noncortical bone, P < .05 for D-sphingosine) (Figure 3C).
Immunofluorescence Staining
The immunofluorescence staining of TBI cross sections showed that the blood vessels had a regular circular shape and capacious intraluminal space in the footprint freshening group (Figure 1C). In contrast, the blood vessels in the preservation group had irregular vascular walls with narrow and indistinct intraluminal spaces. In the D-sphingosine group, the vessel morphology was less regular and not fully circular, but the intraluminal space remained relatively wide. The cross-sectional vessel area in the D-sphingosine and footprint freshening groups was significantly greater than that in the preservation group (P < .05 for D-sphingosine; P < .01 for noncortical bone). The longitudinal sections showed the TCVs being distributed from bone to tendon across the TBI zone (Figure 1D). Fewer capillaries were observed in the preservation group, while footprint freshening brought abundant capillary vessels into the TBI zone and distal tendon. D-sphingosine stimulated vessel formation across the TBI into the distal supraspinatus tendon. Quantitative analysis of branch length and vessel number showed more new blood vessels forming in the D-sphingosine and footprint freshening groups (Figure 3D).
The absence of TCVs markedly enhanced fibroblast activation during tendon remodeling, while fewer fibroblasts were activated in the remodeling tendon alongside TCV formation (Figure 4B). Granulocyte infiltration during the tendon remodeling process was abnormal in the absence of TCVs (Figure 4C). Myeloperoxidase (MPO)–labeled granulocytes were abundant at 3 weeks in both groups; however, in the absence of TCVs, elevated MPO levels persisted within the tendon until 12 weeks, while MPO fluorescence diminished in the presence of TCVs. The absence of TCVs increased stromal fibrosis in the distal tendon, while TCV formation improved stromal fibrosis during tendon remodeling (Figure 4D). Remodeling of the proximal TBI was also affected by TCV formation (Figure 4E). Collagen II content within the TBI was reduced in the absence of TCVs, while the presence of TCVs promoted cartilaginous matrix integration by increasing the collagen II content.
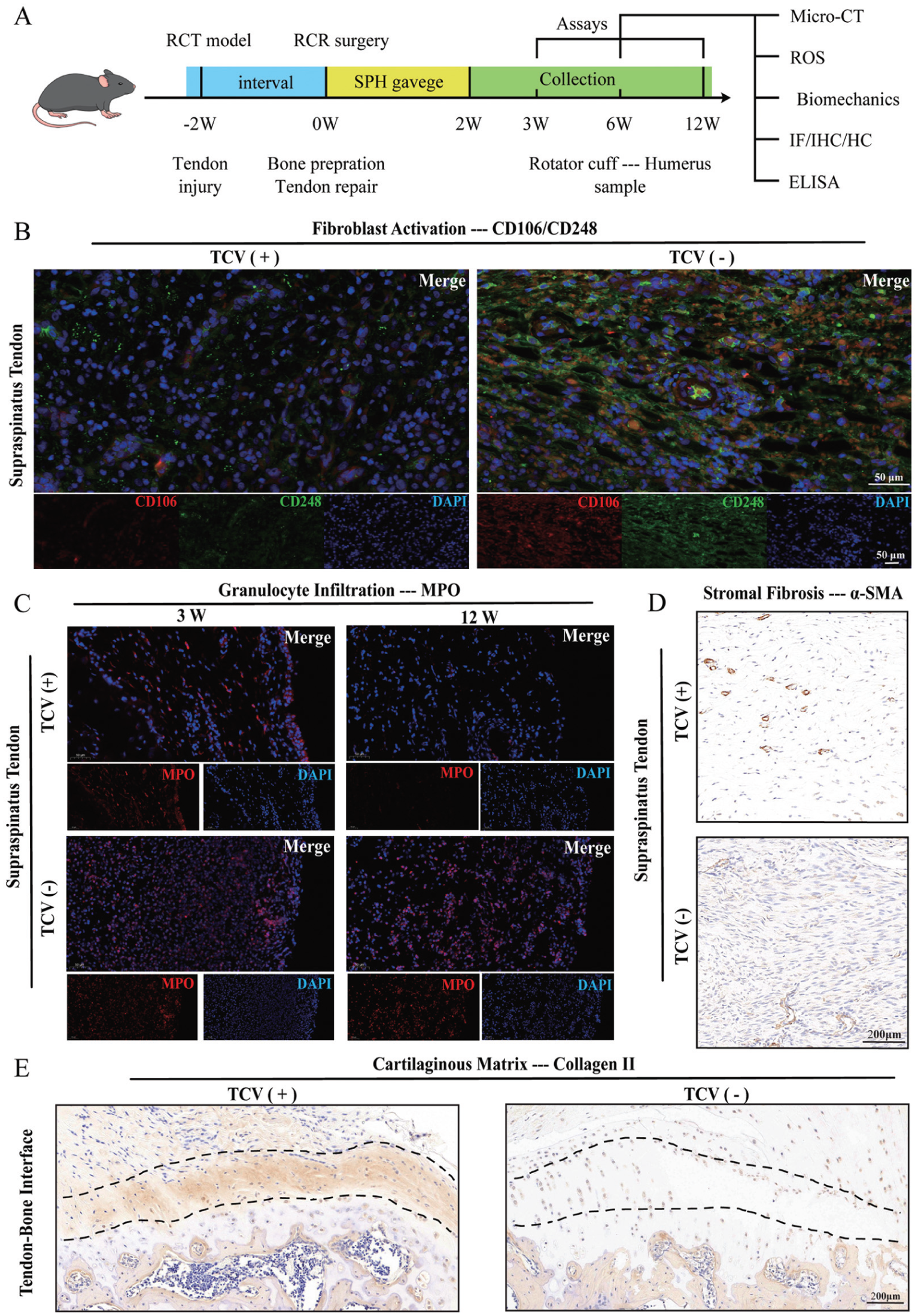
Absence of transcortical vessels (TCVs) impairs the healing quality of rotator cuff by an abnormal tendon-bone remodeling process. (A) Schematic illustration of the animal experimental procedures. Tissue samples are classified by TCV conditions. “TCV (+)” indicates the group with a significantly higher number of TCVs, as determined by quantitative analysis of TCV formation. (B) Immunofluorescence images of clusters of differentiation 106 and 248 (CD106 and CD248) depict fibroblast activation in the supraspinatus tendon. (C) Immunofluorescence images of myeloperoxidase (MPO) indicate granulocyte infiltration in the supraspinatus tendon. (D) Immunohistochemical images of alpha–smooth muscle actin (α-SMA) illustrate tendon stromal fibrosis of the supraspinatus tendon. (E) Immunohistochemical images of collagen II show the cartilaginous matrix content within the tendon-to-bone interface area (black dotted line). Scale bar for immunofluorescence, 50 μm. Scale bar for immunohistochemical staining, 200 μm. DAPI, 4',6-diamidino-2-phenylindole; ELISA, enzyme-linked immunosorbent assay; HC, histochemistry; IF, immunofluorescence; IHC, immunohistochemistry; micro-CT, micro–computed tomography; RCR, rotator cuff repair; RCT, rotator cuff tear; ROS, reactive oxygen species; SPH, D-sphingosine.
Biomechanical Analysis
The maximum failure load and stress of the TBI decreased after footprint freshening at 3 weeks (P < .05) (Figure 3E). In contrast, elastic displacement and stiffness of the supraspinatus tendon were minimally affected across all groups at 3 weeks (Figure 3F). At 12 weeks, the biomechanical properties of the TBI and supraspinatus tendon decreased significantly in the preservation group, while footprint freshening and D-sphingosine treatment enhanced these properties (P < .01 for elastic displacement; P < .05 for other parameters). No significant differences were observed between the footprint freshening and D-sphingosine groups (P > .05).
Enzyme-Linked Immunosorbent Assay
The S1P levels in the D-sphingosine and footprint freshening groups were significantly higher than those in the preservation group at 3 weeks (P < .05). These groups returned to baseline and showed no significant difference among groups at 6 or 12 weeks (Figure 5A). Granulocyte infiltration and inflammatory cytokines (TNF-α and IL-1β) increased rapidly in all 3 groups at 3 weeks. In contrast, levels of these markers in the footprint freshening and D-sphingosine groups returned to near baseline after 6 weeks. The expression of fibroblast activation markers CD106 and CD248 was significantly lower in the D-sphingosine and footprint freshening groups after 6 weeks (P < .05 for D-sphingosine; P < .01 for noncortical bone). In the preservation group, hypoxia-related factors and fibroblast activation markers remained elevated until 12 weeks (Figure 5, B-E).
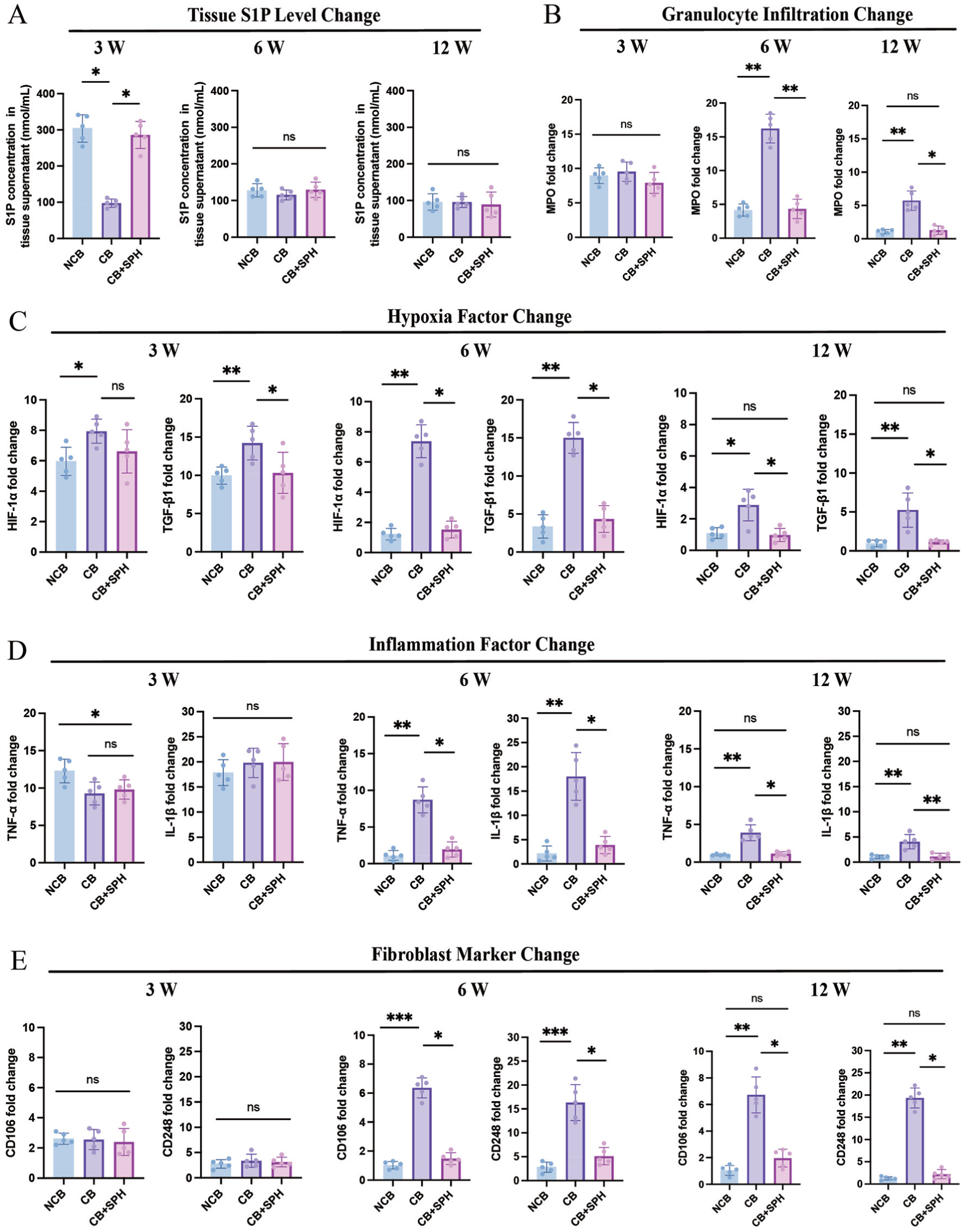
Quantitative analysis of sphingosine-1-phosphate (S1P) concentration, granulocyte infiltration, chronic hypoxia and inflammation, and fibroblast activation within tendon-bone tissue during postoperative healing. At 3, 6, and 12 weeks postoperatively, the fold change in tendon-to-bone interface tissue detected by enzyme-linked immunosorbent assay: (A) S1P concentration, (B) granulocyte infiltration marker (myeloperoxidase [MPO]), (C) hypoxia markers (hypoxia-inducible factor–alpha 1 [HIF-1α], transforming growth factor–beta 1 [TGF-β1]), (D) inflammation markers (tumor necrosis factor–alpha [TNF-α], interleukin–1 beta [IL-1β]), and (E) fibroblast activation markers (clusters of differentiation 106 and 248 [CD106 and CD248]). ns, not significant. *P < .05. **P < .01. Kruskal-Wallis and Dunn multiple-comparison test. Data presented as mean ± SD (n = 5). CB, cortical bone; NCB, noncortical bone; SPH, D-sphingosine.
Discussion
Blood circulation is necessary for bone development and tendon healing. Vascular networks are widely distributed in bone and play a critical role in maintaining microenvironment homeostasis for tendon tissue differentiation and maturation.8,14,28 TCVs provide >80% of arterial blood and 59% of venous blood across bone.8,26 Our results demonstrated that TCVs participated in the functional healing of the rotator cuff, while scar healing within the TBI and tendon was associated with the absence of TCVs. Current reconstructive surgery primarily restores the structured attachment of the ruptured tendon to the footprint bone but does not adequately address vascularization repair within the TBI transitional zone. Our study revealed a distinct reduction in vascular networks within the TBI area after RCR. Footprint freshening and D-sphingosine treatment enhanced postoperative revascularization of the rotator and improved the subsequent remodeling process during tendon-to-bone healing. Revascularization is mediated by endothelial cell migration and organization into vascular walls. 2 Previous studies have reported that D-sphingosine upregulates the angiogenic ability of endothelial cells and contributes to bone defect repair. D-sphingosine is a metabolite catalyzed by sphingosine kinase 1 to generate S1P, which is an important signaling molecule that stimulates the angiogenic activity in endothelial cells. 16 In the present study, local S1P levels in the rotator cuff increased markedly in the early vascularization phase in the D-sphingosine and footprint freshening groups and later stabilized to levels comparable to those in the preservation control during the tendon remodeling phase. These findings suggest that in the D-sphingosine treatment and footprint freshening groups, TCV-mediated revascularization serves as the key driver of the subsequent tendon-bone remodeling process (Figure 6).
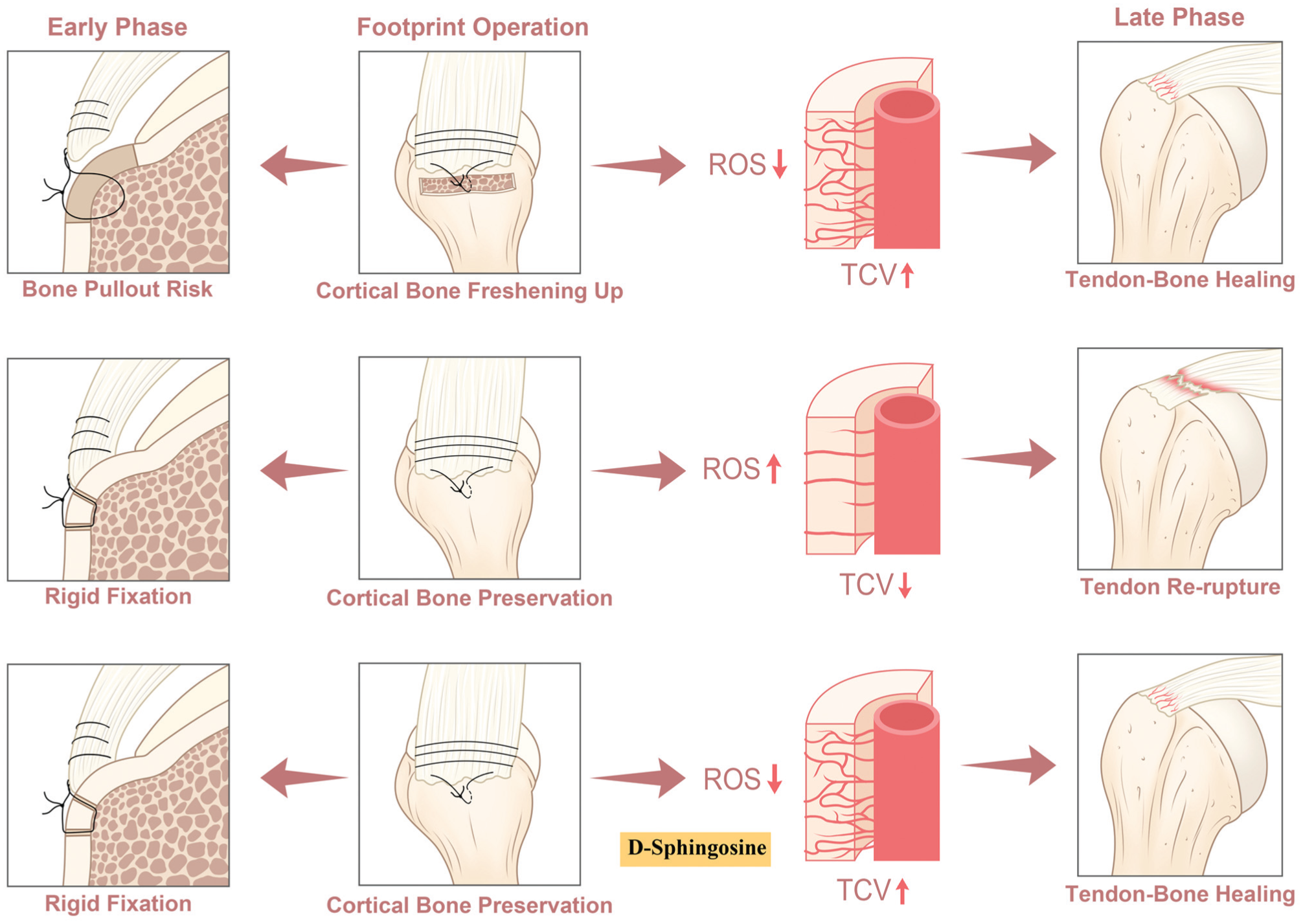
Schematic diagram illustrating the effect of transcortical vessels (TCVs) for rotator cuff repair and the underlying mechanism. Both freshening up and D-sphingosine therapy improve tendon-to-bone healing by TCV enhancement. ROS, reactive oxygen species.
Although tendon is frequently described as a hypovascular tissue and its metabolism relies on passive diffusion, many researchers propose that the functional collagen network is closely associated with the vascular network, especially for active metabolic requirements of the repair stage. 31 In ACL reconstruction, it has been demonstrated that bone insertion–preserved tendon grafts achieve more mature and stable postoperative imaging outcomes as compared with bone insertion–detached grafts. This advantage is attributed to enhanced graft vascularity, which facilitates the avascular necrosis and revascularization process during healing. 17 In this study, cortical bone preservation inhibited the TCV-mediated revascularization and resulted in inferior scar healing of the tendon and TBI. Scar tissue replacement alone is insufficient to meet the biomechanical demands of the rotator cuff, which may help explain the high incidence of rotator cuff reinjury. 13 Surgical techniques have been developed to improve the outcomes of RCR, such as footprint freshening and microfracture of the greater tuberosity, which aim to promote communication between the endosteal vascular system and external tendon attachment. However, these operations may lead to bone mass loss. In addition, secure fixation with a suture anchor requires adequate pullout strength, which relies on sufficient cortical bone preservation. The extent and area of footprint freshening are difficult to control intraoperatively because the transitional zone of the TBI is difficult to distinguish with the naked eye. According to anatomic measurements, the rotator cuff footprint on the greater tuberosity is approximately 23 × 16 mm, making it challenging to accommodate footprint preparation and suture anchor fixation within such a limited area. Considering the potential catastrophic consequences, such as suture anchor loosening or bone fracture, surgeons typically adopt a cautious and conservative approach to footprint preparation to preserve bone biomechanical integrity and avoid fixation failure.22,27 Our biomechanical tests show that footprint freshening weakens the early mechanical performance of RCR owing to cortical bone debridement. Therefore, strategies to enhance revascularization, while providing practical access to the adjacent bone marrow vascular network, are urgently needed for RCR.
Generally, the healing process of tendon reconstruction can be divided into 3 phases. 23 The early phase is characterized by an inflammatory response triggered by tenocyte necrosis and collagen fibril disintegration. The intermediate phase involves revascularization accompanied by tenocyte proliferation and collagen formation, which are crucial for tendon viability and remodeling. In the later maturation phase, these processes gradually stabilize through adaptive responses to mechanical stress and activity.4,28 In contrast, the Achilles tendon benefits from a relatively loose resting position at the ankle joint and a rupture site located near the tendon-muscle junction, both of which provide a rich blood supply, thereby contributing to abundant neovascularization from the muscle and achieving improved outcomes after repair surgery as compared with the rotator cuff. 15 In contrast, the feature of location-restricting tendons keeps the rotator cuff in a constant state of tension. The rupture occurs next to the bone enthesis but far away from the tendon-to-muscle transition zone, leading to less neovascularization from proximal muscle, which is accompanied by undesired scar healing. Thus, we turned our attention to revascularization and sought to improve the healing quality of the rotator cuff by promoting TCV-mediated angiogenesis.
By analyzing different samples and results under different TCV conditions, we found that regardless of whether the footprint was prepared by footprint freshening or D-sphingosine, the healing quality of the distal tendon was mainly associated with the revascularization of TCVs. In the absence of effective TCV-mediated vascular communication from bone to tendon, rotator cuff healing was impeded between the early inflammatory phase and the intermediate remodeling phase. Hypoxia and inflammation factors remained persistently elevated, and the resulting abnormal microenvironment induced sustained fibroblast activation and aberrant tendon remodeling. 21 This environment also promoted the repetitive formation of transitional granulation tissue. In the absence of an adequate vascular network, abnormal matrix deposition could not be effectively cleared, contributing to fibrotic tendinopathy and ultimately impairing healing at the TBI and the distal tendon. In contrast, the presence of TCVs gradually reduced granulocyte infiltration and inflammatory cytokine levels to near-normal levels, and fibroblast activity remained more regulated than when TCVs were lacking. These findings suggest a potential link between hypoxia-driven chronic inflammation and abnormal tendon remodeling, which may underlie scar-mediated healing in the rotator cuff and help explain the therapeutic benefits associated with TCV-mediated revascularization.
This study has some limitations. First, the animal model was limited to small rodents. It remains unclear whether the obtained results could be extrapolated to large primates, so additional work is needed to precisely simulate human rotator cuff disease. In future studies, a large-primate rotator cuff model should be adopted to further investigate the function and mechanism of TCV-mediated revascularization during tendon-to-bone healing for RCR, especially in the context of D-sphingosine combination therapy. For clinical research, retrospective analyses could be carried out to compare current bone surface preparation techniques at the footprint area, specifically cortical bone freshening versus cortical bone preservation, to determine whether significant differences exist in terms of clinical outcomes.
Conclusion
Preservation of the footprint cortical bone induces abnormal tendon remodeling by inhibiting postoperative revascularization. The absence of TCVs leads to hypoxia and chronic inflammation, which trigger fibroblast activation and fibrotic degeneration of the rotator cuff. Footprint freshening or D-sphingosine treatment promotes TCV formation by reducing ROS accumulation within vessels. D-sphingosine has the potential to improve tendon-to-bone healing of the rotator cuff through TCV-mediated revascularization.
Footnotes
Submitted October 14, 2025; accepted April 11, 2026.
One or more of the authors has declared the following potential conflict of interest or source of funding: This work and researchers were supported by the National Natural Science Foundation of China (82471607, 82472483, 82502878).
Data Sharing Statement
All data of this research are presented in the main article. Original data can be acquired from the corresponding authors through reasonable request.