
Abstract
Introduction:
Accumulating evidence demonstrates that subhaemolytic mechanical stresses, typical of circulatory support, induce physical and biochemical changes to red blood cells. It remains unclear, however, whether cell age affects susceptibility to these mechanical forces. This study thus examined the sensitivity of density-fractionated red blood cells to sublethal mechanical stresses.
Methods:
Red blood cells were isolated and washed twice, with the least and most dense fractions being obtained following centrifugation (1500g × 5 min). Red blood cell deformability was determined across an osmotic gradient and a range of shear stresses (0.3–50 Pa). Cell deformability was also quantified before and after 300 s exposure to shear stresses known to decrease (64 Pa) or increase (10 Pa) red blood cell deformability. The time course of accumulated sublethal damage that occurred during exposure to 64 Pa was also examined.
Results:
Dense red blood cells exhibited decreased capacity to deform when compared with less dense cells. Cellular response to mechanical stimuli was similar in trend for all red blood cells, independent of density; however, the magnitude of impairment in cell deformability was exacerbated in dense cells. Moreover, the rate of impairment in cellular deformability, induced by 64 Pa, was more rapid for dense cells. Relative improvement in red blood cell deformability, due to low-shear conditioning (10 Pa), was consistent for both cell populations.
Conclusion:
Red blood cell populations respond differently to mechanical stimuli: older (more dense) cells are highly susceptible to sublethal mechanical trauma, while cell age (density) does not appear to alter the magnitude of improved cell deformability following low-shear conditioning.
Introduction
Human red blood cells (RBC) constitute the functional element of blood that facilitates oxygen and nutrient delivery to tissues, while removing the waste products of metabolism. To perform this action, RBC (with a diameter of ~8 µm) must be able to traverse the vasculature, including the narrow apertures that constitute the smallest regions of the microcirculation (<3 µm diameter), requiring the cell to be highly deformable. 1 In the absence of disease, RBC are very well equipped to perform this action, continually traversing the circulation for up to ~120 days, maintaining membrane integrity while surviving a variety of osmotic, chemical and physical assaults. 2 Given RBC are anuclear, without the ability to resynthesise proteins, physiological ageing has been identified to progressively alter the physical and functional properties of these cells. 3
Investigations into physiological ageing of RBC have identified that senescence progressively induces metabolic depletion and decreased enzymatic activity (including antioxidant defence),3–5 altered cell-surface glycoproteins (particularly decreased membrane-bound sialic acid),6–8 increased spectrin–haemoglobin crosslinking (decreasing membrane elasticity), 9 and decreased mean cell volume (thereby increasing cytosolic haemoglobin concentration and intracellular viscosity). 10 The cumulative effects of physiological ageing significantly decrease RBC function and deformability;6,11,12 yet, before rheological complications are propagated in vivo, the reticuloendothelial system removes cells identified as ‘too old’.7,13,14 Intriguingly, while studies have characterised the biochemical and biophysical alterations of aged cell populations under static conditions (e.g. following collection into a tube at rest), their responses to mechanical stimuli and shear assaults remain largely undescribed.
Recent studies of RBC responses to shear stimuli (within physiological, subhaemolytic and haemolytic domains) have predominantly utilised whole blood that contains cell mixtures of all physiological ages.15–21 While being highly relevant to in vivo blood composition, elucidation of the differential cell response to mechanical stresses of aged and young cells thus would be of value, particularly in the case of transfusion products; blood products may age ex vivo without replenishment from cell turnover, which is likely to skew these products towards older cell types that may be more susceptible to haemolyse in high shear environments (e.g. rotary blood pumps). 22 Given the reported differences in biochemical and biophysical composition of subpopulations of RBC, this study was designed to determine whether density-separated RBC subpopulations (indicating the youngest and oldest cells within a sample) exhibit different responses to mechanical stimuli.
Methods
Subjects and sampling
A total of 10 healthy men (28 ± 10 years) were recruited to participate after providing informed consent. Participants were briefly interviewed to determine recent blood donation history and current health status, ensuring participants were free from cardiovascular, metabolic and haematological disorders, and had not used medications within 90 days. To minimise extraneous shear forces throughout the phlebotomy procedure, 10 mL of blood was collected from a prominent vein in the antecubital region with a 21-G needle and syringe. Blood was collected within 90 s of tourniquet application and immediately transferred into tubes containing ethylenediaminetetraacetic acid (EDTA; 1.8 mg mL−1). All protocols were completed within 4 h of initial collection. The experimental protocols were approved by the Griffith University Human Research Ethics Committee, which conforms with the Declaration of Helsinki.
Cell preparation and density fractionation
Following blood collection, whole blood samples were centrifuged at 1500g for 10 min. To isolate packed RBC, plasma and the buffy coat were aspirated and discarded. To further isolate RBC, packed RBCs were washed twice with isotonic phosphate-buffered saline (0.1 M PBS; pH = 7.40, 290 mOsmol kg−1), centrifuging at 1500g for 5 min each time. Following the final wash, without disturbing the RBC pellet, the supernatant was discarded and the isolated RBCs were aspirated according to density fractions, collecting the least and most dense 10% of the sample. Density fractions have previously been identified to be associated with cell age, with the least dense cells providing a proxy for young RBC and the most dense cells indicating aged RBC.23,24
Osmotic gradient ektacytometry
To assess the baseline deformability capacity of the least dense and most dense RBC suspensions, osmotic gradient ektacytometry was conducted in an annular Couette shearing system and ektacytometer (Laser-assisted Optical Rotational Cell Analyser; LORCA, Mechatronics, Hoorn, The Netherlands) operating at 37°C ± 0.2°C. For this measurement, 200 µL of isolated RBCs were suspended in 5 mL of a polyvinylpyrrolidone (PVP) solution (containing 5% 360 kDa PVP in 0.1 M PBS; pH = 7.40, 290 mOsmol kg−1, 31 cP) and placed in the aspiration chamber. As previously described, 25 during the measurement, the shearing system was configured to generate a constant shear of 30 Pa, while the RBC suspension was continuously aspirated and cycled through the Couette shearing system. Using hypotonic and hypertonic PVP solutions with the same pH and viscosity, the osmolality of the sample being sheared was progressively increased from 65 to 500 mOsmol kg−1. In conjunction with the increases in osmolality, a low-powered laser (655 nm, 3 mW) was passed through the suspension and diffraction patterns of the deforming RBC were captured. Diffraction patterns are typically circular for cells at rest and ellipsoidal for deforming RBC, thus an ‘elongation index’ (EI) was determined throughout the test by fitting an ellipse to the diffraction patterns, using the equation: EI = (A − B)/(A + B), where A is the long axis, and B is the short axis, of the discrete ellipse. The EI-osmolality data were analysed at the following points: (1) the nadir of EI detected within the hypotonic region (i.e. <200 mOsmol kg−1), (2) the maximum EI which typically occurs at physiological osmolarity and (3) the half-maximal EI occurring in the hypertonic region (EIhyper; typically >300 mOsmol kg−1). The ΔEI was determined by calculating the difference between the maximal and minimal EI. The EI-osmolality curves were integrated to obtain the area under the curve.
Shear conditioning of cells
Fresh isolated RBC of each density-fractionated subgroup were resuspended in isotonic PVP at a haematocrit of 0.5% and injected into the 300 µm gap of the Couette shearing system and ektacytometer. RBC suspensions were subsequently exposed, for 300 s, to discrete magnitudes of shear stress (SS) to induce either increased (10 Pa) or decreased (64 Pa) cell deformability; these SS-exposure duration combinations (i.e. ‘shear conditioning’) were based on previous studies15–17 and were confirmed during pilot testing. Shearing was carried out at 37°C in a Couette-type shearing system (gap: 300 µm) which was subsequently used to provide ektacytometry data (in the following). For both shear conditioning protocols (i.e. 10 and 64 Pa), cell suspensions (prepared fresh for every condition) were exposed to the discrete SS for 300 s, after which cellular deformability was determined immediately. To determine the influence of the shearing condition on RBC deformability, baseline (i.e. unsheared) deformability was assessed for a fresh sample prior to the shearing protocol. This process was performed for both the least and most dense cell subpopulations.
Cell deformability
Through manipulation of the rotational velocity of the outer cup of the Couette flow shearing system, the fluid SS were manipulated, and RBC deformability was determined across a range of SS (0.3–50 Pa). As described above, laser diffraction patterns of deforming RBC were analysed for an EI for each discrete level of SS. The collected EI-SS curves were parameterised with a nonlinear fit of the Lineweaver–Burk equation to determine the maximal theoretical EI that can be achieved at an infinite SS (EImax) and the magnitude of SS required to induce half of EImax (i.e. SS1/2). As previously described, 26 the ratio of SS1/2:EImax provides a more robust measure of comparing deformability between RBC populations (as in this study) as this quotient accounts for variance in the maximal deformability. Impairments of RBC deformability are associated with decreased EImax and increased SS1/2 and thus also increased SS1/2:EImax.
Functional tolerance assessment
A subset of the study cohort (n = 6) were invited to return to the laboratory and provide blood samples to facilitate investigation of the time course changes in cell deformability following exposure to sublethal levels of shear exposure (i.e. 64 Pa) for both the least and most dense RBCs. Fresh RBC suspensions were sheared at 64 Pa for 1, 4, 16, 32 and 64 s, prior to immediate assessment of RBC deformability. Cell deformability indices (e.g. SS1/2:EImax) were subsequently extracted from the raw EI-SS curves, and a monoexponential curve was fit to the data to obtain kinetic parameters of the deformability response to sublethal shear exposure.
Statistical analyses
Osmotic gradient ektacytometry data were grouped, following interpolation with a cubic spline, and analysed for a local minimum (within the hypotonic region), local maximum and area under the curve. All data from the osmotic gradient ektacytometry, RBC deformability (i.e. EI-SS curves) and the RBC deformability parameters of shear conditioned samples were investigated using a two-way analysis of variance (ANOVA) with repeated measures, applying the Bonferroni correction. The parameters from the osmotic gradient ektacytometry data and the RBC deformability curve fit of unsheared samples were compared between the least dense and most dense RBC with a paired samples t-test. All statistical analyses and non-linear curve fitting were conducted using commercial software (Prism 7; Graphpad Software Inc., La Jolla, CA, USA). Significance was determined at an alpha of 0.05. Unless otherwise stated, data are presented as mean ± standard error.
Results
Osmotic gradient ektacytometry
The EI of RBC measured across a range of osmotic gradients (mOsmol kg−1) is displayed in Figure 1 for the least, and most, dense RBC populations. The EI-osmolality curves for the least and most dense RBC were significantly different (p = 0.01). Comparative parameters were extracted from the EI-osmolality curve and are summarised in Table 1. Cell deformability parameters were consistently impaired in the most dense cells: (1) dense cells were not able to achieve the same minima (i.e. worse result) in hypotonic solution, representing an impaired tolerance to osmotic stress prior to haemolysis; (2) the dense cells were unable to achieve the same maximal EI in normal osmolality conditions, representing poorer cellular deformability at a constant SS of 30 Pa and (3) thus, the range (Δ EI) between the maxima and minima of EI was decreased in the most dense cells, indicating impaired functional capacity of the RBC across physiological osmotic gradients. The maximal EI in the hypertonic region (i.e. EIhyper), and the osmolality at which this point occurred, was significantly increased for the least dense RBC; these data indicate improved capacity to tolerate large osmolality gradients across the cell membrane.
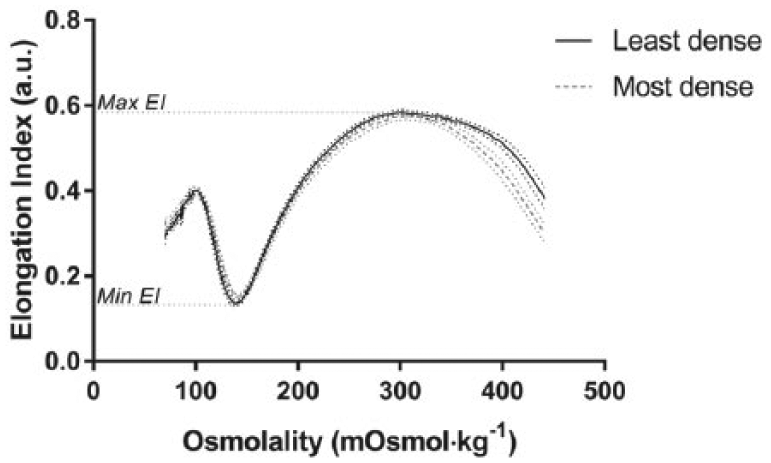
Elongation index (EI) of red blood cells at a constant shear stress (30 Pa) measured against varied osmolalities for the least and most dense cells.
Parameters determined from osmotic gradient ektacytometry.
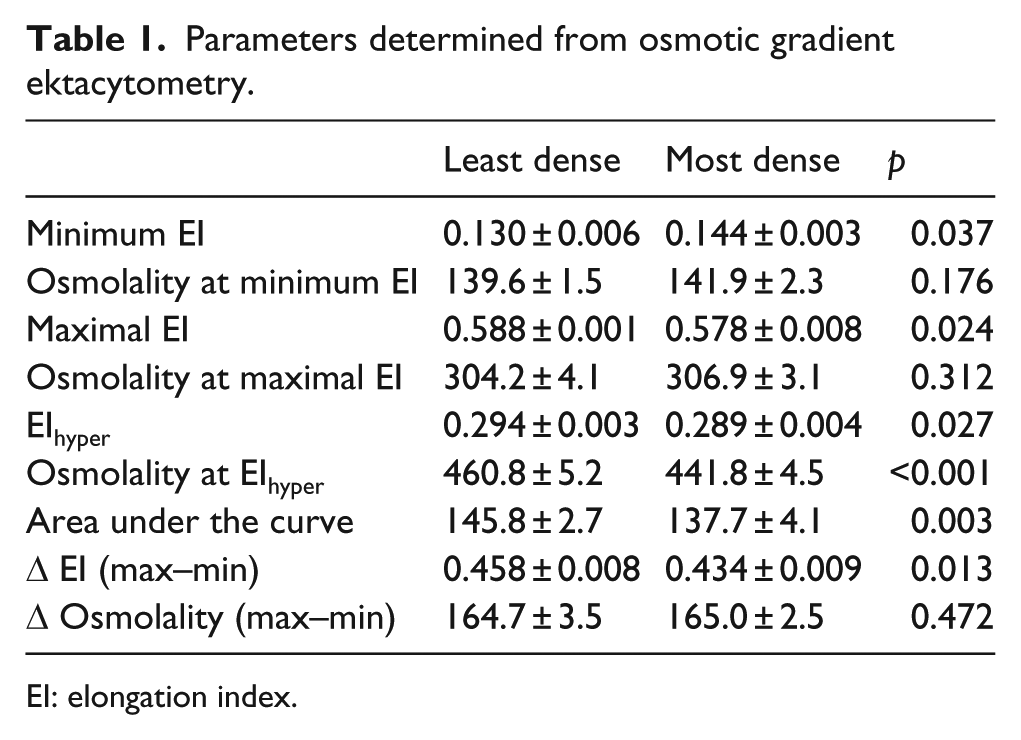
EI: elongation index.
RBC deformability
The dynamic deformability of RBC is presented in Figure 2(a) for the least and most dense cell subpopulations. RBC deformability was significantly decreased for the most dense RBC (p < 0.01), with the greatest differences being detected at the lower shear regions (0.53–9.09 Pa). The maximal theoretical elongation index (EImax) was 0.648 ± 0.004 (least dense) and 0.650 ± 0.004 (most dense), which did not differ. The shear required to induce half of EImax (i.e. SS1/2) was significantly lower (i.e. better) for the least dense RBC (2.54 ± 0.06 Pa) compared to the most dense RBC (2.82 ± 0.07 Pa). The ratio of these parameters (indicating RBC deformability) is displayed in Figure 2(b), where greater values indicate less deformable RBC. The most dense RBC had significantly increased SS1/2:EImax compared to the least dense RBC.
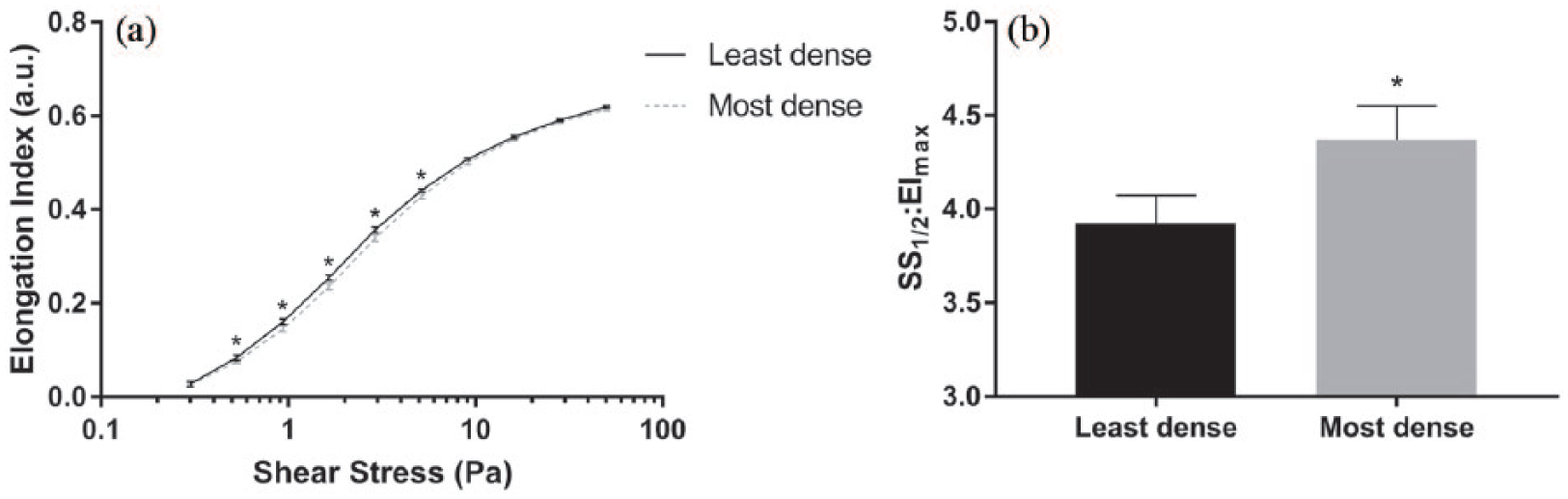
Deformability of the least, and most, dense red blood cell subpopulations. (a) The elongation indexes (i.e. deformability) of RBC suspensions were increased in the least dense cells across a discrete range of shear stresses and (b) an important parameter of RBC deformability (SS1/2:EImax) was significantly increased in the least dense cells, indicating decreased deformability (*significant difference between subpopulations, p < 0.05).
Shear conditioned RBC deformability
Prior exposure to 10 Pa for 300 s resulted in a significant shift of the EI-SS curve (i.e. improvement) for both the least and most dense RBC (Figure 3(a) and (b); p < 0.001). Conversely, exposure to 64 Pa for 300 s resulted in a significant shift of the EI-SS curve to the right (for all SS between 0.94 and 50 Pa; Figure 3(a) and (b)), indicating less deformable RBC. A paradoxical increase in the low-shear EI can be observed for both groups following shear to 64 Pa for 300 s. Parameterisation of the EI-SS curve supported that significant improvements in RBC deformability were observed for both subpopulations following exposure to 10 Pa for 300 s, while significant decrements occurred following the 64 Pa condition (Figure 3(c); all p < 0.001). Regarding group differences, SS1/2:EImax was significantly increased (i.e. decreased deformability) for unsheared condition and 64 Pa conditions for most dense cell populations when compared with least dense (p < 0.05).
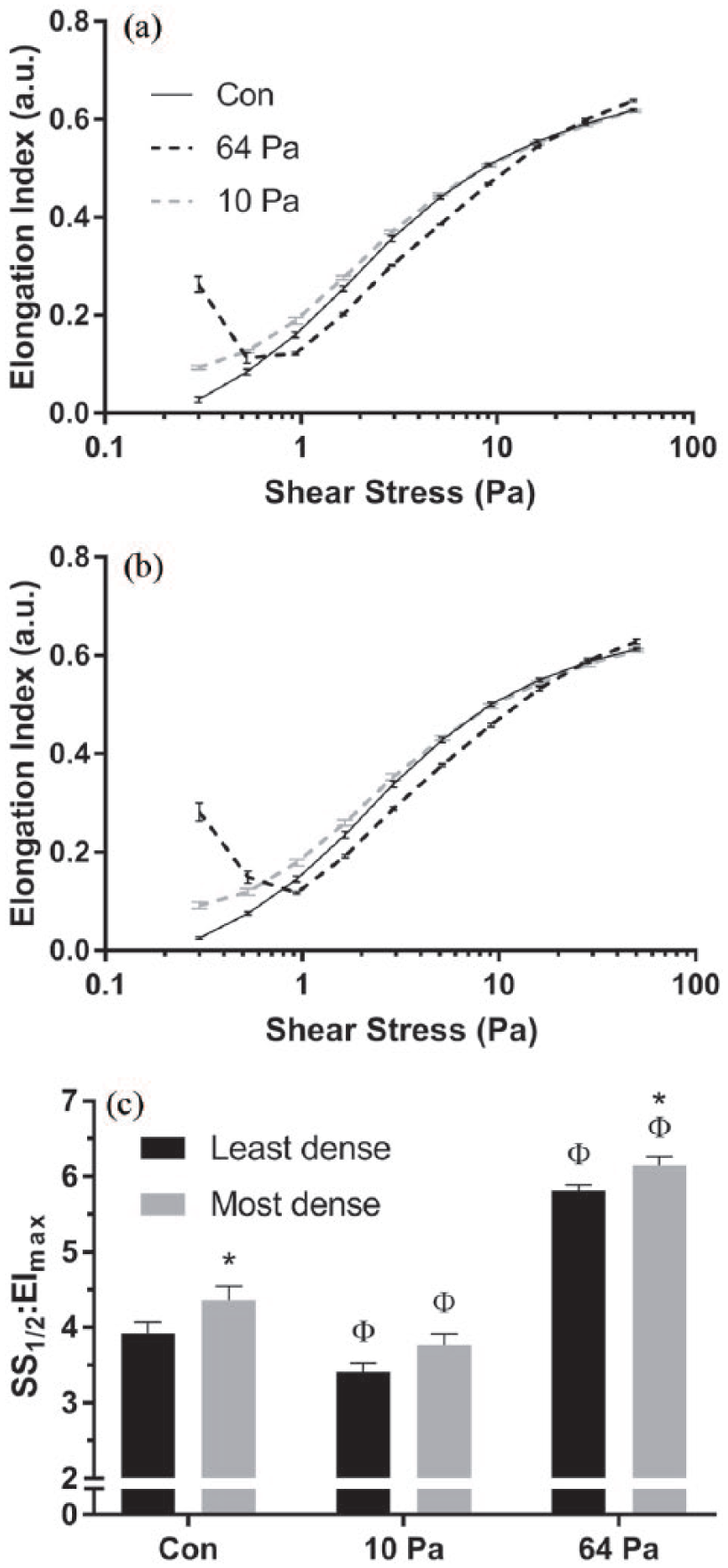
RBC deformability for unsheared samples (Con) and for samples following 300 s exposure to 64 or 10 Pa, for (a) least dense and (b) most dense RBC subpopulations. (c) The combined parameterised data, displaying an important index of RBC deformability; increases in this parameter indicate decreased cell deformability (Φsignificantly different relative to unsheared condition, p < 0.05; *significantly different to least dense RBC within shearing condition, p < 0.05).
The changes in SS1/2:EImax following exposure to 64 Pa for up to 64 s in the least- and most dense RBC subpopulations are illustrated in Figure 4. Following exposure to 64 Pa, measures of RBC deformability projected to an asymptotic SS1/2:EImax ratio of 4.79 ± 0.47 (least dense) and 5.03 ± 0.24 (most dense), which were not significantly different. However, considerable differences in the time constant of the response were observed, where the least dense cells required 42.6 s to achieve 63% of the rate-rise response, while the most dense cells achieved the same rise in 18.4 s.
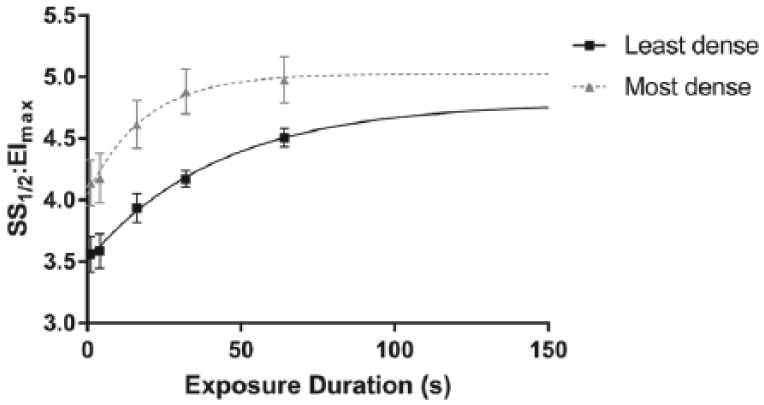
The cellular deformability (SS1/2:EImax) response following 64 Pa shear exposure for varied durations in the least and most dense red blood cell subpopulations. (Higher values represent worse deformability.)
Discussion
The primary findings of this study suggest that subpopulations of RBC separated by density gradients exhibit different capacities to deform under osmotic gradients and following mechanical stimuli. The salient findings include (1) the most dense RBC fraction (i.e. older cell population) demonstrated less functional tolerance to osmotic gradients in constant shear flow, when compared with the least dense RBC subpopulation; (2) the most dense RBC were significantly less deformable prior to conditioning SS (i.e. the ‘unsheared’ cells), when compared with less dense cells; (3) RBC uniformly responded (i.e. improved) to conditioning with shears reflective of the upper limit of the physiological range (10 Pa) and (4) following exposure to supraphysiological shears (64 Pa), cell deformability of the most dense RBC was significantly worse compared to the least dense cells. These findings collectively suggest that RBC subpopulations respond differently to osmotic and mechanical stimuli, with older fractions of RBC being more susceptible to sublethal trauma.
Following separation of RBC into least and most dense subpopulations, osmotic gradient ektacytometry indicated that the most dense RBC were functionally impaired under osmotic challenge compared with the less dense cells. The significantly decreased EIhyper, maximum EI, minimum EI and area under the curve are coherent with the observations of Lutz et al., 27 albeit not as extreme (likely due to the different centrifugation in this study resulting in potentially less separation). These findings support that more dense, possibly older, RBC have decreased ability to deform (supported by the lower range of ∆EI). Specifically, the significantly decreased EIhyper of the most dense population identifies a lower tolerance to a hyperosmotic environment (i.e. a diminished ability to shift morphology), possibly due to a decreased cell-surface area and volume relative to their less dense (likely younger) counterparts. 28 Prior investigations of density-fractionated RBC by micropipette aspiration identified that while older RBC had a smaller diameter and volume, the surface-area-to-volume ratio remained constant, with no evidence of increased membrane shear modulus, thereby not explaining differences in cell deformability.29,30 Older RBCs have been identified, however, to have increased anion exchange (explaining decreased cell water volume), 31 and as cell volume decreases, the relative concentration of intracellular haemoglobin (the primary determinant of cytosolic viscosity) increases. 32 Pfafferott et al. 33 supported these findings, observing that when young cells were osmotically induced to shrink and thus also adopt the same internal viscosity as older RBC, differences in deformability were no longer detectable. These studies, in conjunction with the present investigation, support that the most dense RBC have a diminished ability to deform, likely due to the increased intracellular viscosity secondary to shifts in cell hydration.
The decreased deformability of the most dense fraction of RBC observed in osmotic gradient ektacytometry was maintained when assessed across a varied range of SS in isotonic solution. The baseline deformability data of this study (i.e. Figure 2(a) and (b)) indicated that the most dense RBC had significantly decreased EI, particularly at levels of shear in the physiological range (0–10 Pa). This decrease in deformability may foreshadow impending cell removal from in vivo circulation via increased residence time within the spleen.
Following exposure to sublethal SS (i.e. 64 Pa) for 300 s, it was found that all cells, irrespective of their density, had decreased RBC deformability. Closer inspection indicated that the most dense cells were more affected by sublethal conditioning when compared with less dense cells; that is, impairments in RBC deformability for more dense cells reached near asymptotic levels more rapidly (Figure 4). To our knowledge, these data are the first to be reported, which definitively indicates that the most dense cells (i.e. likely older) are more sensitive to shear damage. It has been previously hypothesised, however, that shear conditioning may preferentially affect older and more dense cell populations.18,22 Simmonds and Meiselman 17 predicted that older cells may lack the functional reserve to accommodate such large mechanical stresses, theorising that the occasionally reported paradoxical ‘improvement’ in cell deformability18,19 may simply reflect that older cells are preferentially haemolysed, leaving behind a younger population with superior mechanical properties. This study provides evidence to support this thesis by demonstrating that more dense cells (reflective of potentially older cells) are rapidly impacted by sublethal trauma, which may reflect the first stage in the haemolytic process. 19 It must be emphasised, however, that while less dense cells were negatively impacted by sublethal SS exposure, the rate of this response was considerably slower, and cell deformability was significantly better when compared to most dense cells at all exposure times <64 s (Figure 4).
A noteworthy observation of this study was the paradoxical EI increase in the low-shear ektacytometry data (0–1 Pa) following exposure to the 64 Pa shear condition. Given the EI is determined via laser diffraction patterns of deforming cell populations, the low-shear increase in EI is likely caused by distortion of the diffraction patterns due to increased heterogeneity of cell populations subsequent to sublethal damage. This phenomenon has been recently reported,16–20 yet remains to be explained despite ongoing investigations. It has been speculated that this observation may be the result of increased tendency for RBC to tumble in shear flow, possibly due to an increased extensional recovery time or increased transitional unstable cell movements (i.e. damaged RBC are unable to align with low-shear flow).16–18 Ongoing studies are investigating the mechanisms underlying this response.
Unlike the 64 Pa condition, the observed improvements following the 10 Pa shear conditioning did not differ between the density-fractionated populations, despite the differences in baseline deformability (Figure 2) and osmotic ektacytometry (Figure 1). Cell deformability improvement following conditioning with physiological magnitudes of shear has been reported in recent studies;15–17,20 consequently, investigations of deformability have increasingly emphasised the involvement of biochemical processes involved in an ‘active’ regulation of cell deformability. Nitric oxide, for example, has been identified to be released by RBC-NOS in the presence of shear stimuli, resulting in S-nitrosylation of the membrane, subsequently increasing membrane elasticity and deformability (for review, see Simmonds et al. 34 ). Recently, older RBCs were identified to have a greater RBC-NOS activation and NO production than their younger counterparts, 35 possibly indicating a heightened active regulation to compensate for a less deformable cell. A compensatory active regulation of the denser RBC subpopulations may explain the equivalent improvement to less dense RBC following physiological shear conditioning.
While the difference in deformability of the density-fractionated subpopulations of RBC is likely a result of variances in cytosolic viscosity, the biochemical involvement in the active regulation of RBC deformability remains yet to be fully described. The present findings support recent hypotheses that sublethal mechanical stresses may ‘filter’ older and less deformable RBC from the blood, resulting in a potential ‘pseudo’ improvement in deformability of the remaining cells. Caution must therefore be taken when interpreting the mechanical properties of blood cells previously exposed to the high shear conditions, typical of mechanical circulatory support, to adequately determine whether the cell populations before and after shear exposure are reflective of the entire population or may be the result of preferential haemolysis.
Footnotes
Acknowledgements
The authors would like to acknowledge Mr Kai Robertson for his assistance with data collection.
Declaration of conflicting interest
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship and/or publication of this article.