
Abstract
In this article, the cytotoxicity of the venom from the nematocysts of jellyfish Cyanea nozakii Kishinouye on human hepatoma cells (Bel-7402, SMMC-7721) and human colon cancer cells (H630) was investigated first. Of the three kinds of cells, the venom had the strongest cytotoxicity on H630 cells with the 50% lethal concentration (IC50) of 5.1 μg/ml. However, the IC50 on Bel-7402 and SMMC-7721 was approximate 17.9 and 24.3 μg/ml, respectively. The cytotoxicity of the venom was affected by pH, temperature and storage conditions. At the pH 4.5–8.5, the venom displayed obvious cytotoxicity and the percentage of survival was about 50%. When pre-incubated at temperatures over 60°C for as short as 10 min, the percentage of survival sharply improved from 4.6% up to 80%. The venom was stored in a more stable condition at −80°C and in lyophilized state compared to other storage conditions used in this study. Lactate dehydrogenase release assay performed on H630 cells indicated that exposure to the venom could result in damage to the cell membrane.
Introduction
The jellyfish, a cnidarian, is known to produce venoms for food predation and self-defence against predator. Jellyfish venoms have many bioactive compounds with enzymatic activity, cardiac toxicity, netrotoxicity, haemolysis, hepatocyte toxicity and myotoxicity (Burnett and Calton, 1974; Gusmani et al., 1997; Nagai et al., 2000; Nevalainen et al., 2004; Radwan et al., 2000; Ramasamy et al., 2005). Severe stings of human beings by jellyfish are frequently reported, resulting in burning pain, local oedema, tingle tight breath, blood pressure depression, and even death. Such effects arise from the complex mixture of biologically active molecules that make up jellyfish venoms (Chung et al., 2001). Therefore, the species and its venom have become a focus of research for medical purposes. The biochemistry, pharmacology and toxicology of jellyfish venoms have been studied since 1960s, being searched for active components as a new source of medicine. It has been reported that the venoms are promising in curing cardiovascular diseases and widely used in nerve molecular biology (Wang et al., 2002). On the other hand, the investigation of the bioactivity of jellyfish venoms could be useful to prevent or decrease syndromes from envenomation of jellyfish. So, the study on jellyfish venoms will be beneficial for human health.
Cyanea nozakii Kishinouye (C. nozakii), a cnidarian of the class Scyphomedusae, the order Semaeostomeae and the family Cyaneidae, is a large-sized plankton. C. nozakii distributes widely in the East Sea, the Yellow Sea and the Bohai Sea of China and flourishes in late summer to early autumn. C. nozakii can seriously damage the fisher environment as its venom causes the death of halobios (Dong et al., 2005; Zhong et al., 2004). And it has garnered attention because of its stinging capacity and the resulting public health hazard. Recently, the haemolytic activity and lethal activity of the venom of C. nozakii (Feng et al., 2010) have been studied, but its cytotoxicity has never been reported.
In recent years, the cytotoxicity assays have been used to assess the toxicity of jellyfish venoms on different cells. Carli et al. (1996) reported that Aequorea aequorea venom greatly affected the growth rate of V79 cells during long-term experiments. And Rhizostoma pulmi venom showed remarkable cytotoxicity on V79 cells and killed all treated cells at the concentration of 15,000 nematocysts/ml within 2 and 3 h. Radwan et al. (2000) reported that the fishing tentacle plus lappets nematocyst venom of Chrysaora achlyos showed minimal cytologic damage to human liver cells (STCC CCL-13) after 30 s exposure to 10 μg protein/ml. On the other hand, the syndrome, skin necrosis, induced by envenomation of jellyfish venom was probably in connection with the cytotoxicity. The present work is the first to report on the cytotoxicity of the venom from jellyfish C. nozakii and the effect of some factors such as pH, temperature and storage conditions on the cytotoxicity. The aim of this study was to verify and characterize the in vitro cytotoxicity of the venom from the nematocysts of C. nozakii. This will provide references for finding new anti-tumour bioactive components from marine animals.
Materials and methods
Nematocyst preparation
The jellyfish C. nozakii was collected at the First Bathing Beach in Qingdao, Shandong Province, China, in August 2007. Tentacles were excised immediately and stored at −20°C until use. The preparation of nematocysts was according to the method of Bloom et al. (1998). Briefly, the refrigerated tentacles were dipped in distilled sea water (v/v, 1:1) for 1 to 5 days at 4°C. Each day the tentacles were stirred softly and the suspension was filtered through a fine kitchen sieve. The filtered fluid was settled for 24 h at 4°C. Then, the final sediments were washed with sea water by centrifugation at 15,000g for 20 min.
Venom preparation
Nematocysts were resuspended in cold Tris-HCl buffer (pH 7.8, 10 mM) and sonicated on ice for 20-s periods at 50 MHz. Then, the suspension was clarified by centrifugation at 15,000g for 20 min at 4°C and used as crude extract from nematocysts of jellyfish C. nozakii (CNN). Sample concentrations were determined by the method of Bradford (1976), using bovine serum albumin (BSA) as a standard.
Cytotoxicity assay
Bel-7402, SMMC-7721 and H630 cells were used to evaluate the cytotoxicity of CNN in the present article. Cytotoxicity was carried out by the method of Chong et al. (2000) with a slight modification. Cells were cultured in RPMI 1640 supplemented with 10% foetal bovine serum, 2 mM
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
Assessment of cell viability was according to the method of Mosmann (1983). At the end of each incubation period, 20 μl of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT, 5 mg/ml in 10 mM, pH 7.4 phosphate buffer saline) were added into each well and the cultures were further incubated at 37°C with 5% CO2 atmosphere. After 4 h, 100 μl of dimethyl sulfoxide (DMSO) was added into the wells to solubilize the formazan crystal products. The microtitre plates were incubated at 37°C for 10 h to make the formazan crystals solubilizable. The absorbance was measured at 490 nm using a microtitre plate reader. The percentage of survival was estimated as the percentage of control absorbance of reduced dye at 490 nm.
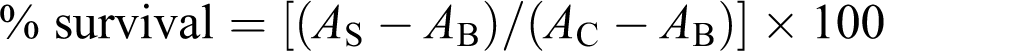
Assay of cytotoxicity affected by pH, temperature and storage conditions
Experiments were carried to investigate the effects of pH, temperature and storage conditions on the cytotoxicity of CNN. The CNN was diluted 10 times with different buffers (acetate 10 mM of pH 4.5 and 5.5, phosphate 10 mM of pH 6.5, Tris-HCl 10 mM of pH 7.8 and 8.5, and biocarbonate 10 mM of pH 9.5 and 10.5). After incubation with different buffers for 3 h at 4°C, the cytotoxicity was determined after incubation with CNN for 12 h according to the method mentioned above.
CNN was pre-incubated at 20, 40, 60, 80°C for different time (10, 30, 50 min), respectively. After cooling at 4°C and incubation with CNN for 12 h, the cytotoxicity was determined according to the method mentioned above.
The venom was stored for various days (2, 4, 6, 9 days) in liquid state at 4, −20, −80°C and in the lyophilized state at −80°C. Then, the cytotoxicity was determined after incubation with CNN for 12 h according to the method mentioned above.
Lactate dehydrogenase assay
Cell membrane damage was carried out by the method of Chong et al. (2000). After incubation with CNN for a period of time, 100 μl of the media overlaying the cells was removed to assay for lactate dehydrogenase (LDH) released in the medium. Then, 100 μl of 1% (v/v) Triton X-100 was added into these wells to lyse the cells for 10 min. Contents in the wells were suspended by repeated piping and removed from wells and assayed for cellular LDH. To determine the quantity of LDH in a specific sample, the sample was centrifuged (at 4°C at 7000 × g for 5 min) to remove cells or debris. Into each well of a UV-transparent microtitre plates, 180 μl of 0.1 M phosphate butter at 37°C, 30 μl of samples, 10 μl of NADH (nicotinamide adenine dinucleotide-reduced) solution (2.5 mg/ml) and 30 μl of pyruvic acid solution (1 mg/ml) were added in sequence. The decrease in absorbance at 340 nm was measured for 3 min. The above procedures were used to assay both released and cellular LDH. The data were expressed as the percentage of LDH released.

Results
Cytotoxicity of CNN
Cytotoxicity of CNN was studied on three different cell lines including Bel-7402, SMMC-7721 and H630. There were no significant differences in the percentage of survival of all cell lines at high concentrations (37–73 μg/ml). However, of the three kinds of cells, H630 cells were most sensitive to CNN at low concentrations (2.5–19 μg/ml; Figure 1a), with the approximate IC50 of 5.1 μg/ml. However, the IC50 on Bel-7402 and SMMC-7721 was approximately 17.9 and 24.3 μg/ml, respectively. Therefore, H630 cells were selected to evaluate the cytotoxicity of CNN.
(a) Effects of CNN on the production of formazan in three cell lines after incubation with different concentrations of venom for 48 or 64 h. (b) Effects of CNN on the production of formazan in H630 cell lines after incubation with different concentrations of venom for 12, 24 and 48 h. Data were presented as means ± SD of three parallel measurements.
The responses of H630 cells incubated with different concentrations (2.5, 5, 10, 19, 37, 73 μg/ml) of CNN for various periods of time, namely 12, 24 and 48 h, are shown in Figure 1b. The cytotoxicity of CNN was time and dose-dependent. The cytotoxicity improved with the increase of incubated time. At the concentrations below 37 μg/ml, the obvious differences in percentage of survival in three treated groups were found. At the concentration of 10 μg/ml, the percentage of survival was 65.6%, 43.2% and 35.9% after incubation for 12, 24 and 48 h, respectively. However, from 37 to 73 μg/ml the percentage of survival was slightly different in the three treated groups. The cytotoxicity also improved with the increase of the concentration of CNN. For the three incubated periods, at the concentrations from 2.5 to 19 μg/ml, the percentage of survival was markedly decreased from about 70% to 30%. The approximate IC50 of CNN on H630 cells was 15.9, 8.8 and 5.1 μg/ml after incubation for 12, 24 and 48 h, respectively.
Effects of pH and temperature on the cytotoxicity of CNN
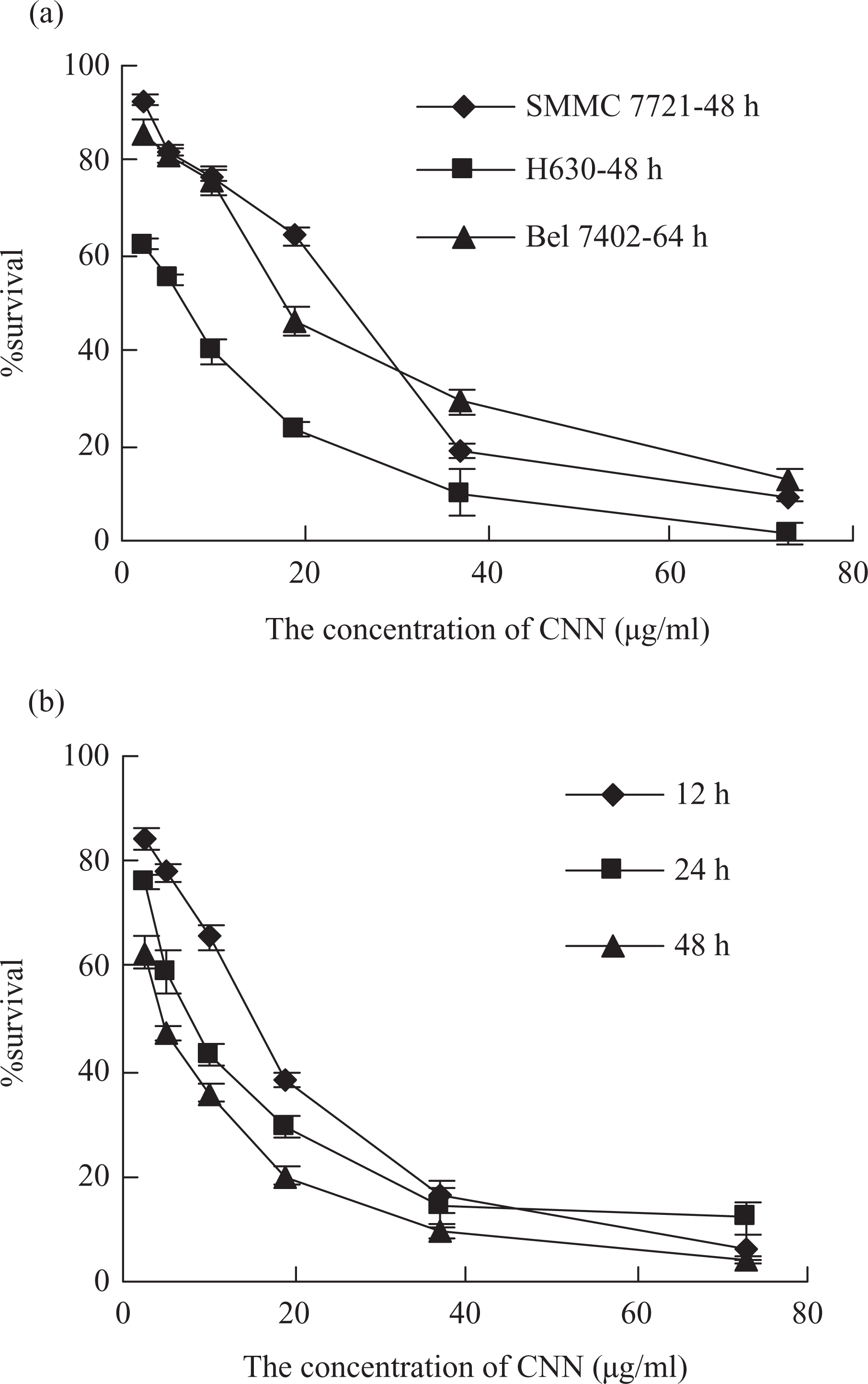
The effects of pH on the cytotoxicity of CNN were studied. As shown in Figure 2a, CNN had obvious cytotoxicity at the pH 4.5–8.5 and the percentage of survival was approximately 50%. However, the cytotoxicity was significantly inhibited at pH 3.5 and the percentage of survival was about 93.6%.
Effects of pH (a) and temperature (b) on the cytotoxicity of CNN. Data were presented as means ± SD of three parallel measurements.
The effects of temperature on CNN are shown in Figure 2b. In general, the cytotoxicity of CNN was temperature-dependent and it decreased with the increasing temperature and prolonging reaction time. No significant change in the cytotoxicity of the venom was observed when CNN was incubated at 20°C for 50 min; incubation at 40°C for 50 min, the cytotoxicity of CNN significantly decreased and the percentage of survival was from 17.7 to 43.6%; at high temperatures (60 and 80°C), a great loss of cytotoxicity was observed. The cytotoxicity was entirely lost when CNN was kept at 60°C for 50 min and at 80°C for 10 min.
The cytotoxicity of CNN after storage on different conditions
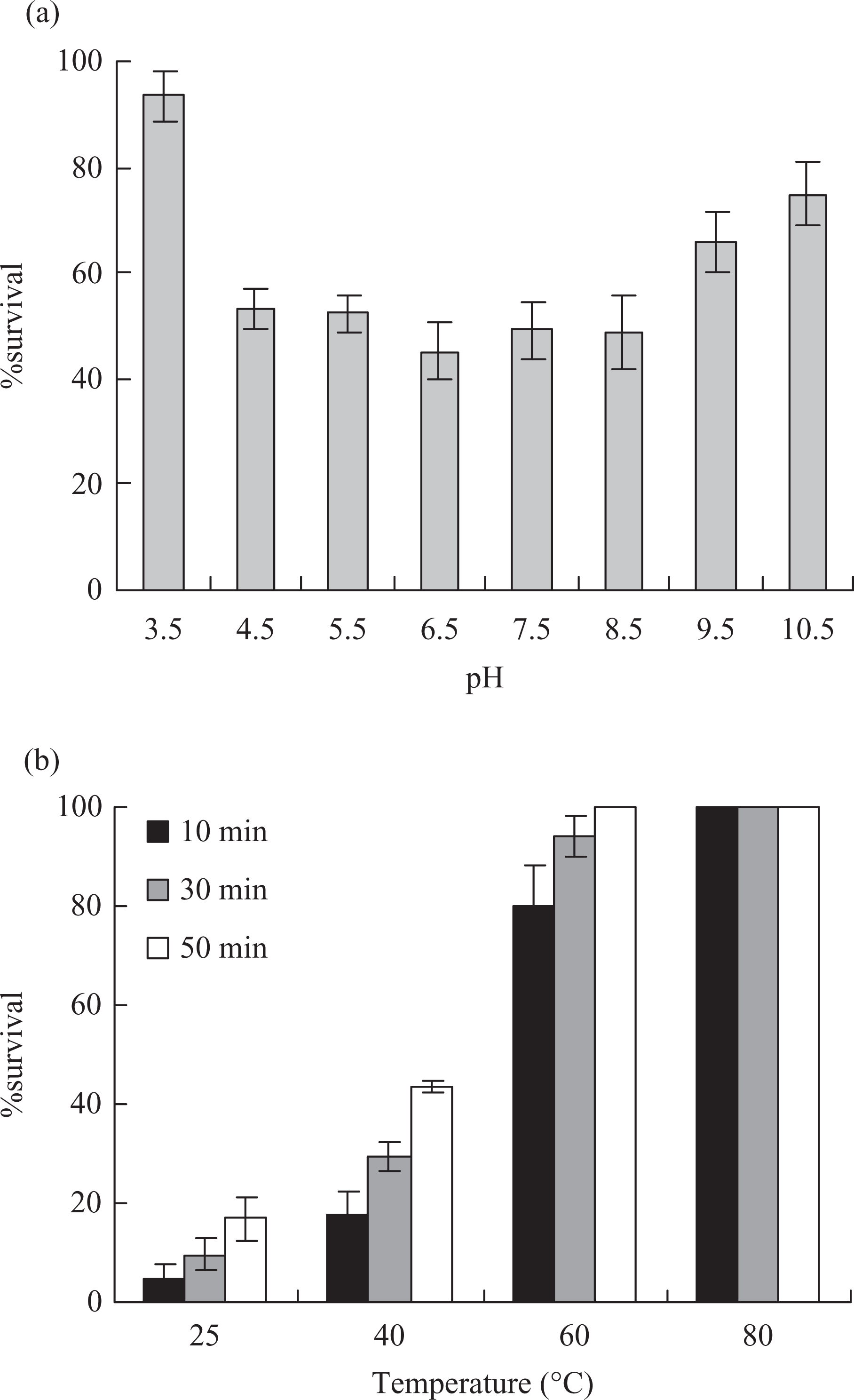
In order to assess the stability to storage of CNN, the cytotoxicity of CNN was investigated just after extraction and after storage for different days at 4, −20, −80°C and in the lyophilized state storage at −20°C. As shown in Figure 3, the cytotoxicity decreased with time. Clearly, storage at −80°C and in lyophilized state the cytotoxicity reduced slightly in 9 days; storage at 4°C for 6 days, the cytotoxicity did not impair remarkably. Nevertheless, storage at 4°C for 9 days, the cytotoxicity decreased sharply and the percentage of survival was enhanced to 96.9%; storage at −20°C for 6 days, the cytotoxicity was lower than that of the venom stored at 4°C. But for 9 days, the cytotoxicity was higher than that of the venom stored at 4°C.
Effects of different storage conditions on the cytotoxicity of CNN. The percentage of survival of the venom just after extraction was 39.7%. Data were presented as means ± SD of three parallel measurements.
LDH assay
Cytotoxicity of CNN on H630 cells was further assessed using the LDH release assay. The results are shown in Figure 4. LDH assay showed marked LDH leakage. The percentage of LDH released was also dose- and time-dependent. The percentage of LDH released improved with the increase in concentration and incubation time. At the concentrations from 2.5 to 19 μg/ml, the percentage of LDH released significantly increased at three incubation time, but it improved slightly at the concentrations from 37 to 73 μg/ml.
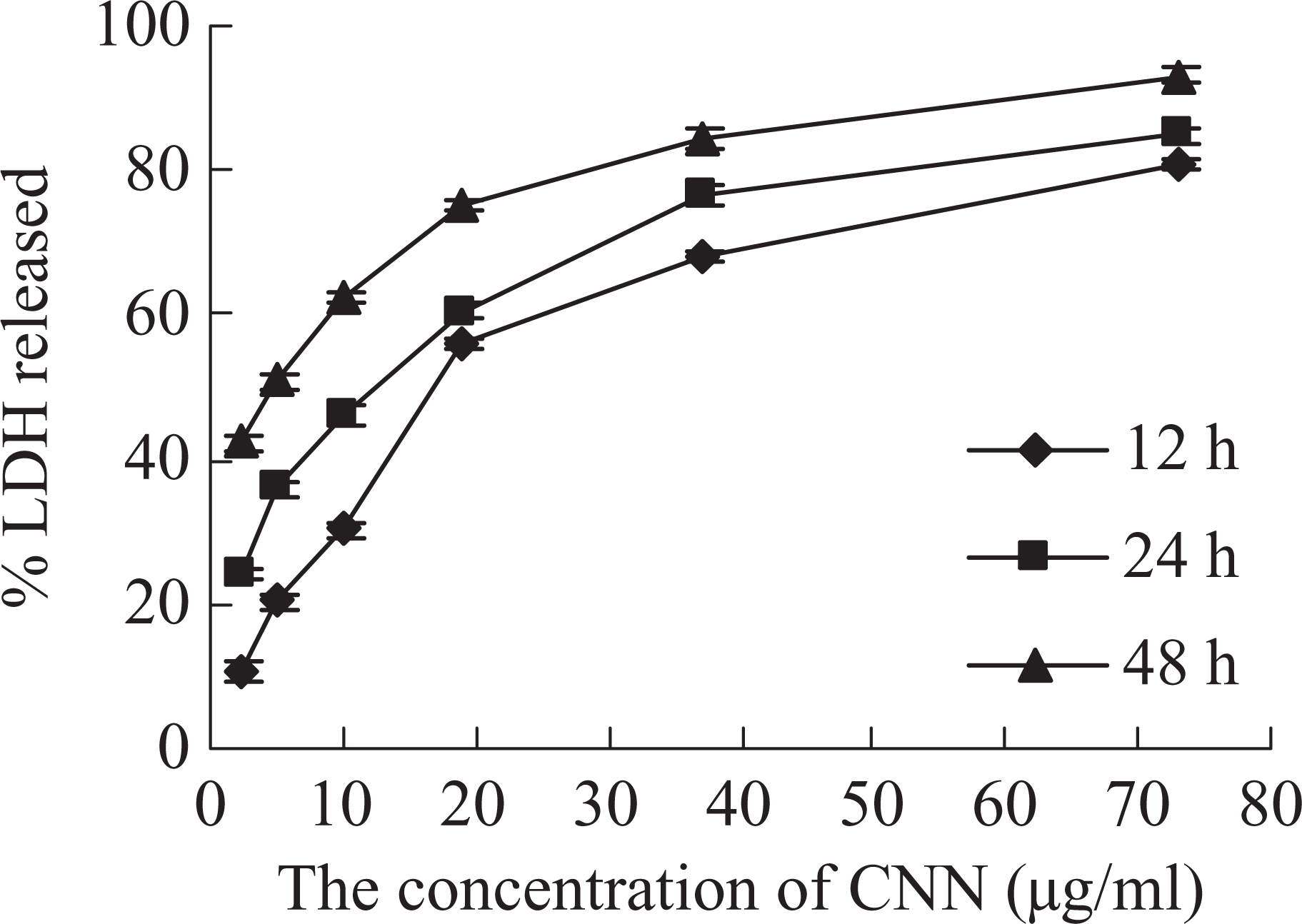
Effects of CNN on the percentage of LDH released in H630 cell lines after incubation with different concentrations of toxin for 12, 24 and 48 h. Data were presented as means ± SD of three parallel measurements.
Discussion
Cytotoxic tests are usually employed to evaluate toxicity of venoms, such as sea anemone, dinoflagellate and alga (Carli et al., 1996; Li et al., 2007; Naves et al., 2006). This method is easy, ethical and uses small amounts of active venom. Jellyfish venoms clearly vary in activity and in composition (Bailey et al., 2005). The results presented in this study showed that the nematocysts of C. nozakii contained powerful toxins with strong cytotoxicity on H630 cells. The approximate IC50 of CNN on H630 cells was 5.1 μg/ml after incubation for 48 h, while the IC50 of the Chiropsalmus quadrigatus venom on human malignant glioma cells (U251 cells) was about 80.3 μg/ml after incubation for 48 h (Sun et al., 2002).
pH had marked effects on the bioactivities of the venom from cnidarians. In previous studies, Marino et al. (2004) reported that the Aiptasia mutabilis venom displayed strong cytotoxicity at a pH of around 7.5. However, at pH 4.5 and 9.5, the cytotoxicity was nearly lost entirely. The cardiovascular activity of the venom from Chironex fleckeri (C. fleckeri) was active at a pH rang of 5–9 and lost at pH 3 (Winter et al., 2007). A 50% loss of haemolytic activity of the venom from Carybdea marsupialis was found in a few minutes after extraction by sonication in Krebs Ringer phosphate buffer (pH 7.4), but the stability was increased by lowering the pH values of the extract medium (Rottini et al., 1995). In this article, CNN had high cytotoxicity at a pH rang of 4.5–8.5, while at pH 3.5, the cytotoxicity decreased greatly. One possible reason for this result might be that pH mainly affected the charges of CNN, and the charges could influence the venom binding to the target membrane which was the first step to cell damage (Edwards et al., 2002). On the other hand, pH could influence the spatial structure of bioactive proteins with changes in the proteinaceous conformation.
The present study revealed that the cytotoxicity of CNN was temperature-dependent. When pre-incubated at 60°C, the cytotoxicity sharply reduced. Moreover, at 80°C, even for as short as 10 min, the cytotoxicity was destroyed entirely. It was indicated that the venom was most likely of a protein nature as the cytotoxicity of CNN was inactivated by excessive heat. Loss of cytotoxicity of CNN might be attributed to change in molecular structure, heat denaturation of the protein and presence of proteases in the venom. Some bioactivities of jellyfish venoms were significantly affected by temperature. The myotoxicity of the C. fleckeri venom was lost within 24–46 h at room temperature and inactivated by heating at 42°C (Endean et al., 1993). Heat treatment at 60°C for 1 h inactivated the cytotoxicity of Chrysaora venom (Ordóez et al., 1990).
The cytotoxicity of CNN was affected by storage conditions. Storage at −80°C and in lyophilized state did not affect the cytotoxicity greatly. And they were the best methods to store CNN for long time. As the cytotoxicity was relatively stable after storage at 4°C for 4 days, CNN could be stored at 4°C for short time. Storage at −20°C for 6 days, the cytotoxicity decreased greatly, and it was lower than that of the venom stored at 4°C. May be there were one or more components that could degrade the protein with cytotoxicity in CNN, which were more active at −20°C, so the cytotoxicity of CNN stored at −20°C was lower than that of CNN stored at 4°C.
Cytolytic toxins were known to operate by either of two general mechanisms: enzymatic and stoichiometric, with the stoichiometric lysins including pore formers (Hessinger and Lenhoff, 1988). LDH release assay performed on H630 cells indicated that exposure to CNN could result in damage to the cell membrane (Chong et al., 2000), but its detailed underlying mechanism was still unclear. In other studies, Man-of-war venom produced pore-like structures in the membranes of target cells, which lead to colloid osmotic swelling with subsequent release of intracellular proteins and cell lysis (Edwards et al., 2002). Li et al. (2006) reported that carp hepatocytes died in apoptosis at low concentrations of microcystins, while at high concentrations, they died in the form of necrosis. The activation of calpain and Ca2+/calmodulin-dependent protein kinase was believed to be critical in the microcystin-induced apoptolic process (Ding and Ong, 2003). Houck et al. (1996) reported that the sea nettle venom did not form large membrane channels similar to complement, nor did calcium appear to play a major role in the mechanism of toxicity in hepatocytes. Intracellular accumulation of toxic oxidative products of sea nettle jellyfish venom metabolism may play an important role in the cytotoxicity of the venom (Cao et al., 1998).
Conclusion
The cytotoxicity of CNN on Bel-7402, SMMC-7721 and H630 cells was investigated for the first time in this article. CNN could result in damage to the cell membrane because of the release of LDH, and it had the strongest cytotoxicity on H630 than on Bel-7402 and SMMC-7721. The cytotoxicity was affected by incubation time, temperature and pH. The cytotoxicity of CNN was stable when stored at −80°C and in lyophilized state, and they could be used as the storage conditions of CNN.
Footnotes
This work was financially supported by the National Natural Science Foundation of China (Grant No.41006095), Qingdao Science and Technology Project (08-1-3-51-JCH), Guangdong Province and Chinese Academy of Sciences overall strategy Cooperative Project (2010B080703027) and the Knowledge Innovation Program of the Chinese Academy of Sciences.