
Abstract
The aim of this study was to evaluate the morphological changes, liver function test (LFT), and oxidative stress damage caused by thiobarbituric acid reactive substances (TBARS), in mice exposed to vanadium via inhalation. Male CD-1 mice were exposed to vanadium pentoxide (V2O5) via inhalation (0.02 M), 1 hour twice a week for 6 weeks. At the end of the protocol, controls and exposed mice were killed to evaluate the changes. Histological analysis and LFT were performed to detect the damage. TBARS detection was assessed for oxidative stress. Inflammatory infiltration, binucleation, and meganucleus were detected in the liver of V2O5-exposed mice (p < 0.05). Alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels were also significantly increased (p < 0.05). Lipid peroxidation was significantly higher in V2O5-exposed animals compared to controls (p < 0.05). V2O5 exposure induced inflammation and cell damage detected by the increase in ALT and AST levels, as well as histological changes that suggest regenerative changes, such as binucleation and meganucleus.
Introduction
The liver as the major site for metabolism, biotransformation and detoxification of drugs and foreign compounds is constantly exposed to reactive oxygen species (ROS) resulting in oxidative stress and frequently, permanent and irreversible tissue damage (Bataller and Brener, 2005). In normal conditions, the basal production of ROS is inactivated by effective endogenous antioxidant systems; when an imbalance between both occurs, noxious conditions are established. During oxidative stress, ROS can activate Kupffer cells and redox-sensitive transcription factors, such as the nuclear factor κB (NF-κB) and activator protein 1 (AP-1), which in turn initiate the synthesis of proinflammatory cytokines and fibrogenic mediators (Bataller and Brenner, 2005). ROS can also covalently modify proteins, lipids and DNA with the consequent loss of biological functions, cell viability and ultimately degenerative events in the liver (McCord, 2000; Valko et al., 2005). In the liver, lipid peroxidation is responsible for necrosis and death of hepatocytes (Webb and Twedt, 2008), through elevated reactive lipid aldehyde production.
Environmental pollution caused by heavy metals is more often the result of combustion of oil derivatives (Figueroa et al., 2006; Huang and Ghio, 2006). Vanadium is a major pollutant found in the air of highly contaminated urban localities. Its increased concentration in autopsies of Mexico city residents, especially in the lung tissues, has been well documented (Fortoul et al., 1996). Vanadium accumulates in the lungs, bone tissue and liver (Roschin et al., 1980). Vanadium inhibits several key enzymes of cell metabolism, including adenylcyclases, kinases, ribonucleases, phosphatases and ATPases (Mujherje et al., 2004), in so doing it is logical to assume that the damage it causes at the cellular level is partially attributable to metabolic disruption and oxidative stress (Dafnis and Sabatini, 1994); oxidative stress could also act as a cofactor in pro-oxidant reactions by replacing the iron ions in Fenton’s reaction (Fickl et al., 2006; Galaris et al., 2008).
The aim of our work was to determine, in a more integrative manner, histological, biochemical and ROS-mediated protein modifications in the liver of mice exposed to vanadium pentoxide (V2O5) via inhalation, in conditions that mimic those faced by human beings living in highly polluted environments.
Methods
Animals
Eight-week-old healthy male CD-1 mice (bred in the vivarium of our medical school) weighing 35 ± 2 g were housed in hanging plastic cages in an animal facility (at an average temperature of 21°C, 57% humidity, controlled lighting [12:12 h light/dark regime]) and fed Purina rat chow and water ad libitum. The experimental protocol was in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals and the local guidelines on the ethical use of animals from Mexico’s Health Ministry (NOM-062-Z00-1999). For all experimental groups and procedures, animals were randomly selected.
Reagents
All reagents were purchased from Sigma-Aldrich Chemical Co (St Louis, MO, USA) or Merck (Mexico). Deionized water (Milli R/Q System, Millipore) was used for the preparation of all solutions.
Exposure regimen
Inhalation exposures were performed as described by Piñon-Zarate et al. (2008). Briefly, 0.02 M V2O5 (99.99% purity, Sigma-Aldrich) was added to deionized water containing 100 μl Tween-20/100 ml suspension. The aerosol inhalation chamber was an acrylic box measuring 45 cm × 21 cm × 35 cm that could house 25 mice per session and had a total volume of 3.3 L. The ultra-nebulization to generate the atmosphere was carried out using a DeVilbiss Ultra Neb 99 (Somerset, PA, USA) system that maintained a constant flow of 10 L/min. According to the manufacturer, about 80% of the aerosolized particles inhaled by the mice would be expected to have a mass median aerodynamic diameter of 0.5–5.0 μm.
A 0.22-μm filter was positioned at the external outlet of the ultra-nebulizer during each exposure period. At the end of each exposure the filter was removed and weighed. Vanadium concentration in each filter was analysed using a graphite furnace atomic absorption spectrophotometer (Perkin Elmer Mod. 2380, Shelton, CT, USA), at a wavelength of 318.4 nm and a slit of 0.7 nm.
In these studies, a set of 10 mice were placed in the acrylic box to inhale V2O5 for 1 h/d, twice a week, for 6 weeks. Control mice (10) inhaled only the vehicle deionized water/Tween during each exposure.
Evaluation of serum liver function tests (LFTs)
Blood samples were obtained by intracardiac puncture prior to dissection. Samples were centrifuged at 3000 rpm for 8 min at room temperature. Serum was separated and analysed in the automated system LX20 Mark (Automate Sample processing Systems, Beckman Coulter Inc, Fullerton, CA, USA), by following the manufacturer’s instructions. Concentrations of albumin (g/dl), alkaline phosphatase (IU/L), alanine aminotransferase (ALT; IU/L), aspartate aminotransferase (AST; IU/L), direct bilirubin (DB; IU/L), and total bilirubin (TB; IU/L) were determined.
Tissue sampling and preparation
At the end of the inhalation protocol, mice from each group (n = 10/cohort) or vehicle exposure were killed. The animals were anesthetized with sodium pentobarbital and perfused, via the aorta, with saline solution containing 10% formaldehyde in phosphate buffer. The liver was then removed and placed on ice. One fraction was processed for histological examinations (i.e. paraffin embedding and staining with haematoxylin-eosin for subsequent light microscopy evaluation). Sections were evaluated by two independent observers unaware of the tissue experimental procedure. The other fraction was homogenized within 10 min in 10 volume of ice-cold saline using a Dupont’s Sorvall RC-5B centrifuge (refrigerated superspeed centrifuge, LV Scientific Ltd, UK); the homogenates were then centrifuged at 4000 rpm for 10 min at 4°C to yield a low-speed supernatant fraction that was used immediately for thiobarbituric acid reactive substances (TBARS) assay.
Assay of lipid peroxidation
An accurate technique to evaluate lipid peroxidation is by estimating malondialdehyde (MDA) a by-product of peroxidated lipids using TBARS assay (Santamaria and Rios, 1993). In brief, for TBARS assay, the liver tissue was homogenized in 1 ml of saline solution (0.9% sodium chloride [NaC1]). An aliquot of 0.25 ml of this homogenate was added to 0.5 ml of the thiobarbituric acid ([TBA] 0.5% weight/volume [w/v] of TBA + 16% w/v of trichloroacetic acid + 2.5% volume/volume [v/v] of hydrochloric acid). To this mixture, 10/11 of aqueous desferrioxamine (1.5 mM) and 10 pl of butylated hydroxytoluene (3.75% w/v) in methanol–water (50% v/v) were added in order to prevent artifactual increase in lipid peroxidation due to the presence of ferrous ions and the in situ overproduction of TBARS as a result of the boiling process in the technique, respectively. The mixture was submerged in a boiling water bath for 20 min and centrifuged (5000 rpm for 15 min). The absorbance of the supernatant was measured at 532 nm in a Beckman DU-6 UV visible spectrophotometer (GMI Inc., USA). A standard curve for TBARS formation was constructed using a periodic acid oxidation rate of 2-deoxy-
Statistical analysis
Results were expressed as mean ± SD. Comparisons among the groups were carried out by one-way analysis of variance (ANOVA) followed by Tukey’s post test. Differences are considered to be statistically significant at p < 0.05. The complete analysis was processed with SPSS 11.0 (Statistical Package for the Social Sciences, Chicago, IL, USA).
Results
Histological alterations
Mice liver tissue was evaluated for the presence of histological alterations, such as steatosis, swelling, necrosis, degeneration and fibrosis. Control group (Figure 1 ) presented a normal morphology; with hepatocytes measuring between 20 and 30 μm in diameter and nuclei between 7 and 10 μm in diameter. No signs of inflammatory response, degeneration, necrosis or liver fibrosis were found. Figure 2 depicts the appearance of liver tissue from the V2O5-exposed group. Aberrant foci inflammatory were evident (eosinophils, neutrophils and mononuclear cells). Additional changes in this group comprised the presence of meganucleus. The number of the meganucleus in this group was significantly higher (p < 0.05) than that of the controls (Figures 3 and 4 ). With regard to the number of binucleated hepatocytes (another marker of morphologic hepatic alteration), these were significantly increased in the V2O5-exposed group when compared to the controls (p < 0.05; Figures 4 and 5 ).
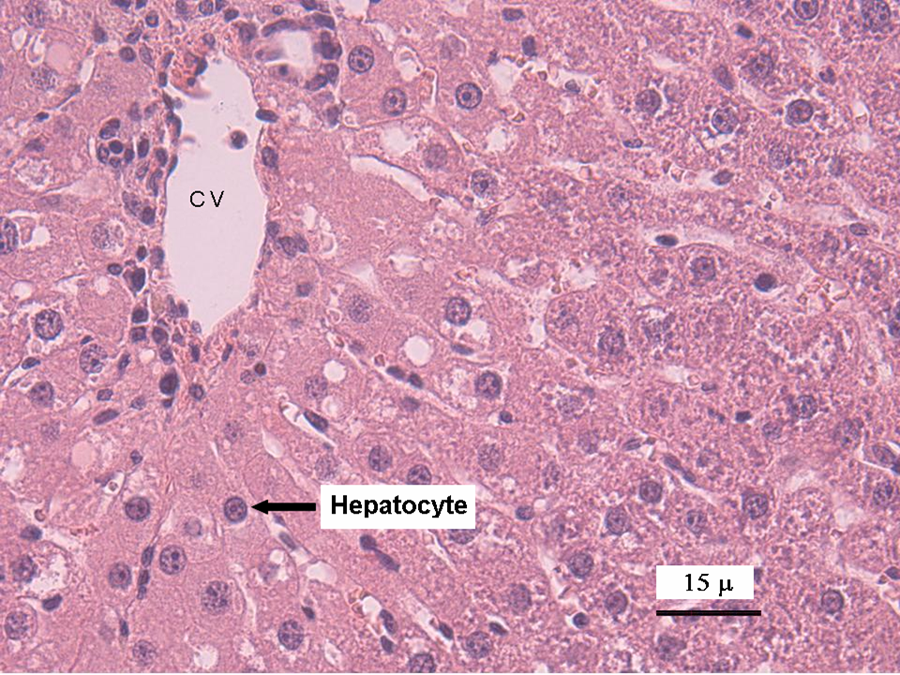
Photomicrograph of the liver of control mouse. A normal distribution of parenchymal tissue can be observed. Hepatocytes present a normal appearance. The central vein (CV) is shown, and the arrow indicates a hepatocyte nucleus. Haematoxylin and eosin staining (×40).
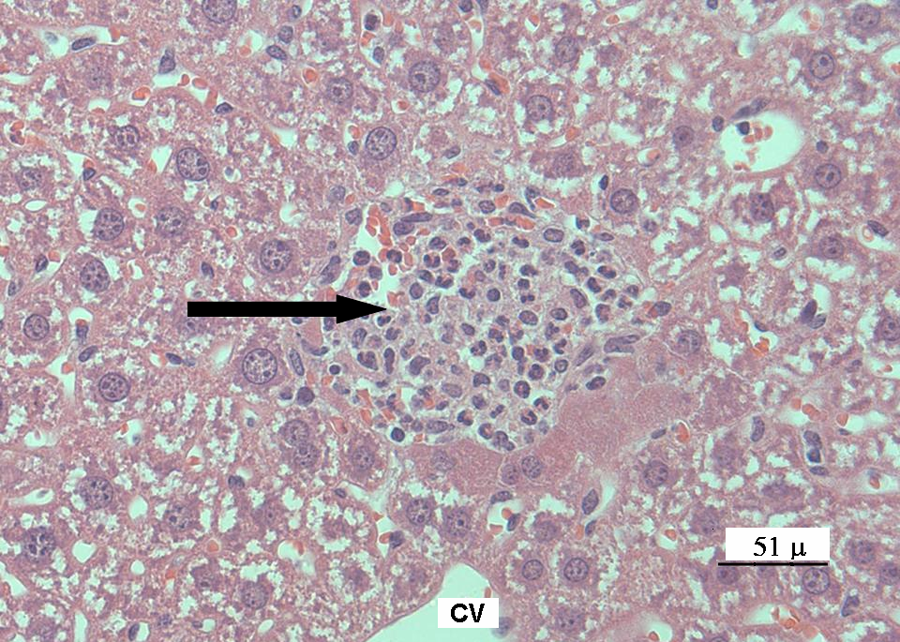
Photomicrograph of an infiltrated inflammatory periportal area with eosinophils, neutrophils and mononuclear cells from the liver of vanadium pentoxide-exposed mice. Black arrow indicates the inflammatory focus. Haematoxylin and eosin staining (×40).
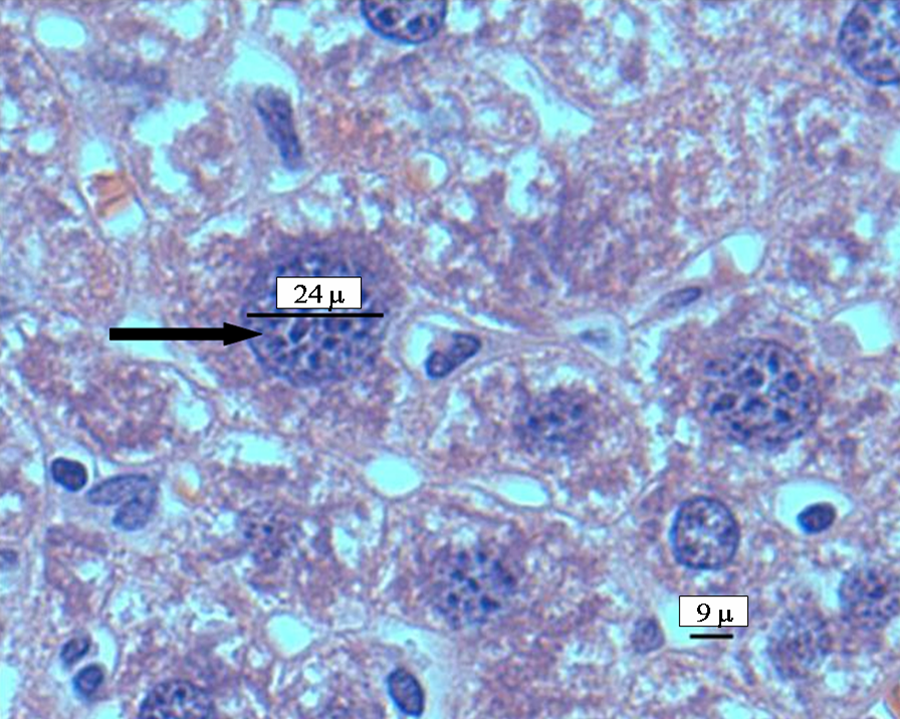
Photomicrograph of a hepatocyte with a meganucleus (indicated by the black arrow) of vanadium pentoxide-exposed group. Haematoxylin and eosin staining (×40).
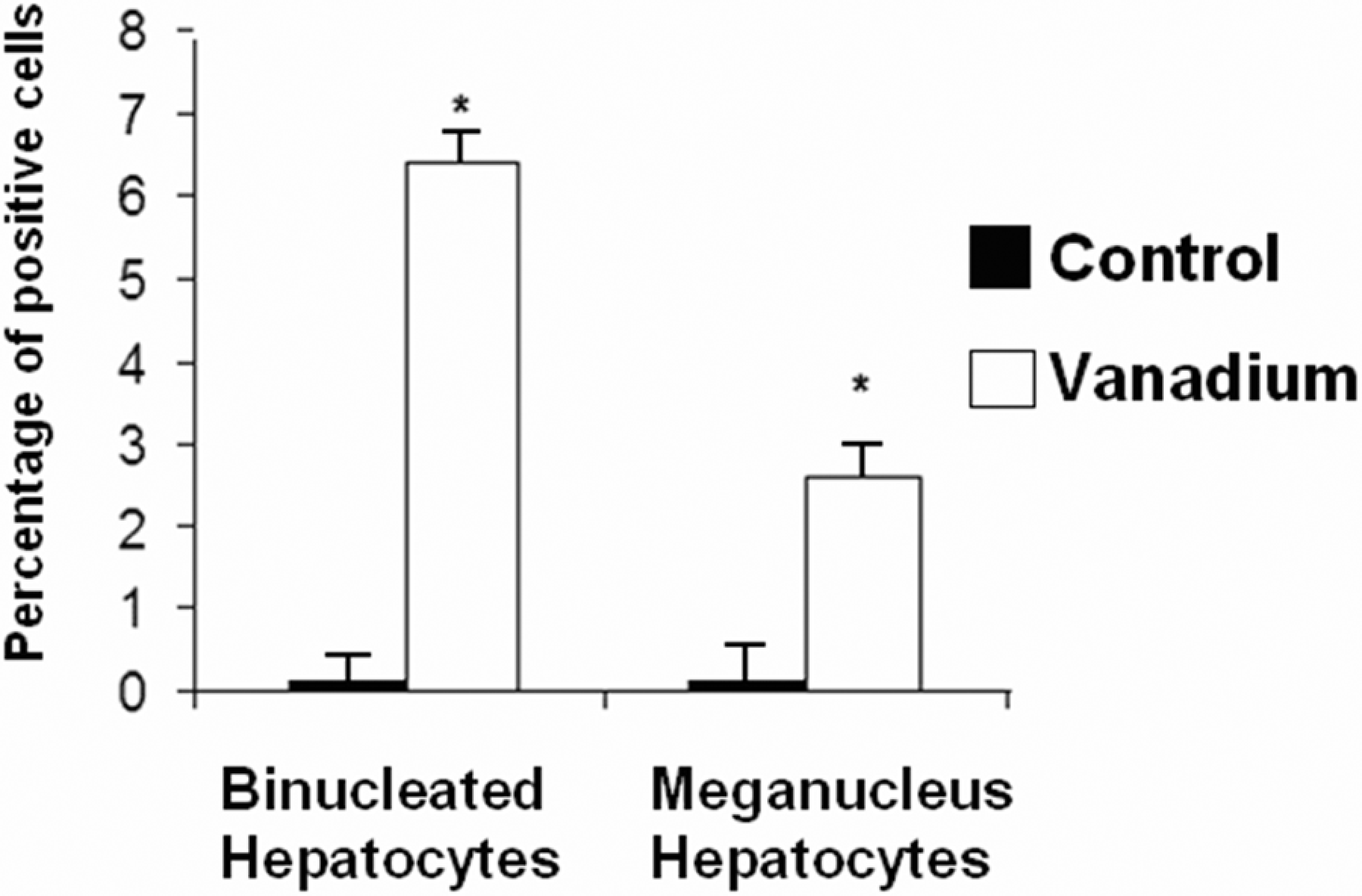
Percentage of binucleated hepatocytes and hepatocytes with meganucleus in the liver tissue from the control and vanadium pentoxide-exposed mice. Data from n = 10 mice per group. *p < 0.05 denotes differences when compared with the control group. One-way analysis of variance (ANOVA) followed by Tukey’s post test.
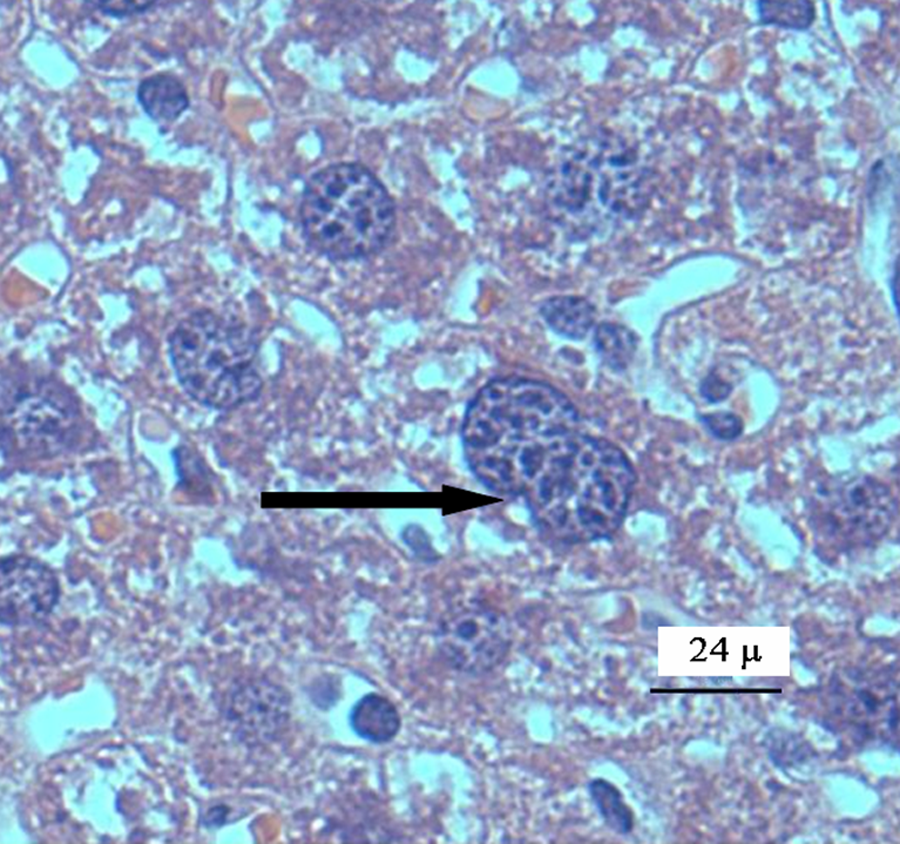
Photomicrograph of binucleated hepatocytes (pointed by black arrows) from vanadium pentoxide-exposed mice. Haematoxylin and eosin staining (×40).
Oxidative damage to membranes
The levels of LP in the liver tissues of V2O5-exposed mice and controls are shown in Figure 6 . TBARS formation was significantly increased in the V2O5-exposed group when compared with the control group (p < 0.05).
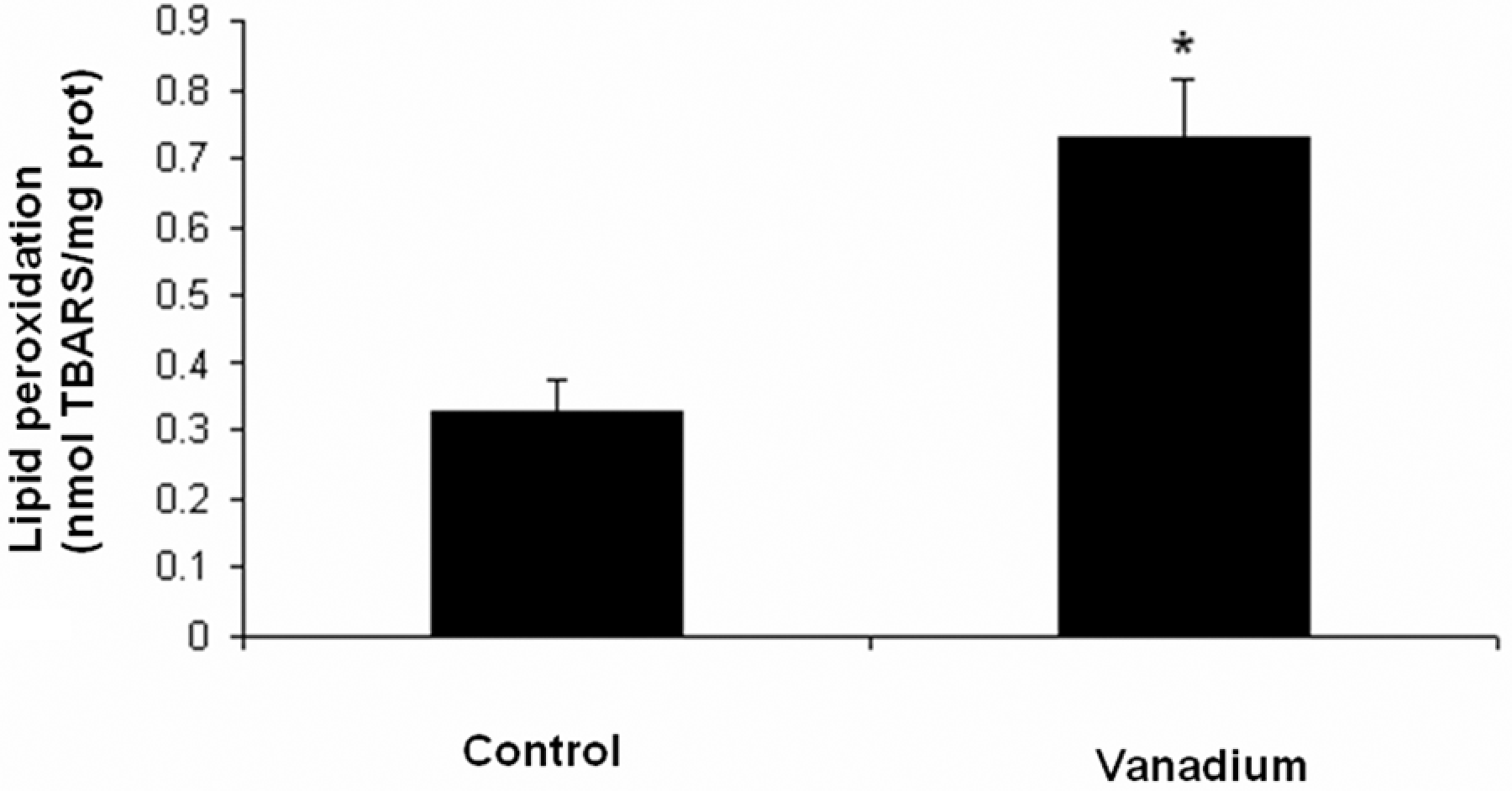
Quantification of lipid peroxidation in liver tissue from the control and vanadium pentoxide-exposed mice. Data from n = 10 mice per group. One-way analysis of variance (ANOVA) followed by Tukey’s post test *p < 0.05 denotes differences when compared with the control group.
Liver function
In regard to the results of those markers of liver function, there were no significant changes between the control and the V2O5-exposed groups in DB, or albumin. In contrast, ALT and AST increased twofold with respect to the control group (p < 0.05; Figure 7 ).
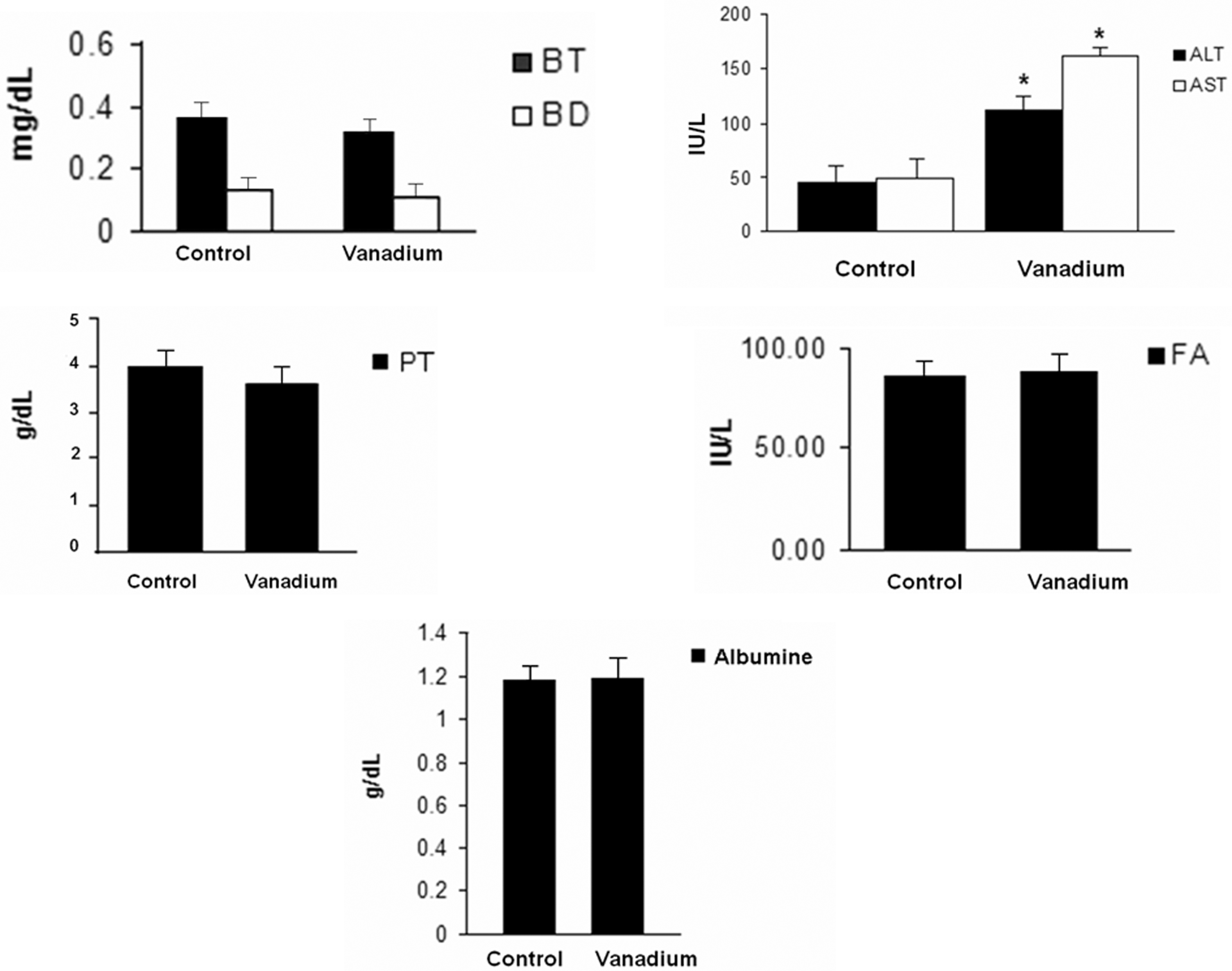
Liver function tests (LFTs) of the serum from the control and vanadium pentoxide-exposed mice. Data from n = 10 mice per group. One-way analysis of variance (ANOVA) followed by Tukey’s post test. *p < 0.05 denotes differences when compared with the control group. Total protein (TP; g/dl), albumin (g/dl), direct bilirubin and total bilirubin (IU/L), alkaline phosphatase (IU/L), were not statistically significant, when compared to alanine aminotransferase (ALT; IU/L) and aspartate aminotransferase (AST; IU/L). There was a statistically significant difference between the vanadium-exposed group and the control group.
Discussion
Regardless of the primary cause of degeneration, liver damage can be associated with specific patterns of biochemical abnormalities detectable in blood, and these alterations are grouped as general markers of LFT, commonly used in clinical trials for the estimation of the extent of liver damage (Chopra and Griffin, 1985; Reichling and Kaplan, 1988) and also useful in assessing pharmacological hepatotoxicity (Sheig, 1996). Furthermore, hepatocellular necrosis in blood is confirmed by means of aminotransferases, including AST and ALT. These enzymes catalyse the transfer of aspartate or alanine groups to ketoglutaric acid to produce oxaloacetate and pyruvate, respectively (Giannini et al., 2005). ALT is a cytoplasmic enzyme, while AST is present in cytoplasmic and mitochondrial domains. It is assumed that the increase in the activity of these enzymes in serum is the result of their leakage from injured cells, thereby reflecting the extent of injury to hepatocytes. Therefore, high levels of these enzymes are seen in several liver diseases, especially in those associated with a significant degree of hepatocyte necrosis, such as acute viral hepatitis and ischemia. It is also important to mention that ALT is relatively specific to liver, whereas AST is located in heart muscle, kidney, brain, pancreas and blood cells, in addition to hepatocytes (Sheig, 1996). In general terms, liver damage can be identified by TB and DB, as well as alkaline phosphatase, ALT, AST and albumin (Giannini et al., 2005). Its toxicity depends on its valence and its dual property to behave as cation and anion (Barceloux, 1999; Chakraborty et al., 2005).
Our results clearly show general alterations produced in the liver from mice exposed to inhaled vanadium, although in contrast to previous reports, there was no evidence of necrosis or degeneration (Roschin et al., 1980). Nonetheless, our results support the hypothesis that inhalation is an important route for liver poisoning caused by vanadium, and thus, this toxicant might considerably contribute to liver damage by altering its structure and function (Barceloux, 1999). As judged by our findings, vanadium is able to create pro-oxidant conditions directly in hepatocytes, and this has been supported by previously reported evidence (Dafnis and Sabatini, 1994).
Although our model explored only acute and subacute in a high-dose exposure to V2O5, as a preliminary approach this was sufficient to evoke hepatic alterations mediated by this metal in the liver, in our model the vanadium concentration in the inhalation chamber, the average concentration in the 18 filters was 1436 ± 225 μg/m3 during the whole experiment. This simple condition has not been reported not far from cases of human exposure, because to our knowledge it is simulating situations such as exposures during intense fires, burning oil wells, during burner cleaning or other working exposures scenarios. In this regard, ‘burner’ syndrome, in which one of the symptoms is greenish tongue, has been characterized as typical of vanadium poisoning (Dafnis and Sabatini, 1994; Roschin et al., 1980; Woodin et al., 2000).
Going through the assessment of liver function, we were able to collect only limited evidence of alterations induced by the toxicant. Therefore, various conditions of vanadium exposure should be explored in further studies in order to clarify any possible alteration induced by this metal, simulating the human exposure condition. In the meantime, in order to validate our model, any inference in relation to vanadium exposure in humans should be made on the basis of all those factors accounting for liver damage (Giannini et al., 2005). Despite the limitations in our model, the extent of damage found in liver correlated with the markers of oxidative stress (OS) explored as well as with the degree of morphologic alterations in hepatocytes.
The changes found in the liver from vanadium-exposed mice also mimicked those alterations described for poisoning with alkaloid pyrrolizidine (Jago, 1969), in which a pathognomonic finding includes increase in the size of the nuclei of hepatocytes. The size of the nuclei of hepatocytes increased to about 30 μm which is a criteria currently employed to define meganucleus (normal sizes range from 7 to 10 μm); in our model, this is suggestive of hypertrophy and liver regeneration due to liver damage (Jago, 1969; Svoboda et al., 1971).
The increase in the number of binucleated hepatocytes might be related to the extent of damage produced by vanadium to the cytoskeleton at the nuclear level, a condition that may be involved in pausing mitosis (Domingo, 1996). The polyploidy of the liver is also generally considered to show terminal differentiation and senescence (Toyoda et al., 2005). It was revealed that the increase in the oxidative stress can result in significant oxidative DNA damages in the liver of mouse. It was also reported that hepatic polyploidy is induced by oxidative injury and attenuates the proliferative potential of cells (Gorla et al., 2001). Our findings regarding a stimulated LP in liver caused by vanadium exposure could be the result of major alteration in the antioxidant defense systems (Halliwell and Chirico, 1993). This may be suggested on the basis of previous reports showing lower activity of superoxide dismutase (SOD), decreased levels of the endogenous antioxidant, and reduced glutathione (GSH), in other toxic models with oxidative alterations such as LP (Webb and Twedt, 2008). Increased LP in liver is the unequivocal result of the deleterious actions of ROS and shows decline in hepatocyte membrane functions, further leading to altered fluidity, reduction in the electrochemical cell potential, and increased permeability across the membrane and cell damage (Webb and Twedt, 2008).
In regard to the alterations in ALT and AST levels found in our study, it is known that the increase in these enzymes in serum is the result of their leakage from injured cells, and therefore, it reflects a certain degree of injury to hepatocytes, especially to their membranes (Reichling and Kaplan, 1988). These enzymes are high in many forms of liver disease, but particularly in those hepatocytes resulting in necrosis, as it has been observed in acute viral hepatitis and ischemia (Chopra and Griffin, 1985; Giannini et al., 2005). Taking into consideration that ALT is specifically located only in the liver, while AST is widely located, besides hepatocytes (Giannini et al., 2005), it can be inferred that vanadium is capable of causing damage in central nervous system and lung (Fortoul et al., 1996; Valko et al., 2005), as judged by the elevated levels of AST. Concerning to the raised levels of ALT, we could suggest a smolder inflammatory process as a consequence of the exposure, since the half-life of vanadium is about 50 days, and its presence could maintain the oxidative stress in the liver (Ramaiah, 2007).
In summary, these events may be playing a role in the further development of liver pathology, and more research should lead us to consider the possible damage of the liver in areas with high atmospheric pollution.
Footnotes
Acknowledgements
The first author thanks the support given by Posgrado en Ciencias Biológicas, UNAM. G Cano-Gutiérrez received a scholarship from CONACyT, México. The authors thank Veronica Rodríguez-Mata for her technical assistance.
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.