
Abstract
The aim of this study was to explore the effectiveness of
Introduction
Dioxins and dioxin-like polyhalogenated aromatic hydrocarbons are toxicants detrimental to biological systems that are produced as unwanted by-products in industrial processes (Kulkarni, 2008). 2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD) is a highly toxic persistent organic pollutant among these hydrocarbons (Lee and Jacobs, 2009). TCDD displays a wide spectrum of toxic effects, including dermal toxicity, reproductive and developmental toxicity, immunotoxicity, hepatotoxicity, carcinogenicity, teratogenicity, neurobehavioral, endocrine, and metabolic alterations (Chopra et al., 2010; Gan et al., 2009; Li et al., 2010; Xu et al., 2008; Yin et al., 2011). Moreover, the increased levels of multisite cancers are established in TCDD-treated animals (Bock, 1994; Grassman et al., 1998; Hung et al., 2006). Dioxins accumulate along the food chain particularly in liver and adipose tissue (Scrogi, 2008). Dioxin exposure in mammals leads to a broad spectrum of adverse affects, including hepatotoxicity of varying severity (Boutros et al., 2008). Industrial or military exposures to dioxin and related compounds have been linked to increased prevalence of fatty liver in human beings (Lee et al., 2010). The marked change induced by TCDD is degeneration of the hepatocytes (Czepiel et al., 2010). Also, TCDD is a potent promoter of liver cancer (Huff et al., 1994). Oxidative stress is involved in the onset of several disorders after TCDD exposure (Ciftci et al., 2011; Lee et al., 2002). Antioxidants play an important role in inhibiting and scavenging free radicals, thus providing protection to humans against infectious and degenerative diseases (Nader et al., 2010). Hence there is an increasing interest to find the appropriate agents with antioxidative protection for hepatocytes. On the other hand, there are equivocal findings of chromosomal aberrations in humans exposed in vivo to TCDD (IARC, 1997) and the increases in production of 8-oxo-2-deoxyguanosine (8-oxo-dG) in the liver of mice (Hung et al., 2006). However, there are equivocal findings of chromosomal aberrations in humans exposed to TCDD in vivo (IARC, 1997). Today, it is reported that dioxin-like chemicals alter the expression of numerous genes in liver, but the pathways remain unknown, leading to major toxicities such as hepatotoxicity, wasting, and lethality (Forgacs et al., 2010; Moffat et al., 2010).
In certain situations, people require the reasonable substitution of different dietary components. These include, among other things, the amino acid glutamine (Gln), which plays a central role in this study. Gln is the most common free amino acid found in the human body (Rohde et al., 1996). The nonessential amino acid Gln has recently been the focus of extensive scientific interest because of its importance in cell and tissue cultures and its physiologic role in animals and humans. Abundant evidence suggests that Gln may become a ‘conditionally essential’ amino acid in the critically ill patients (Lacey and Wilmore, 1990). Gln has been shown to restore muscle mass in cachexia due to cancer, rheumatoid arthritis, AIDS, and in critically ill trauma patients (Berk et al., 2008; Clark et al., 2000; Kuhls et al., 2007; May et al., 2002; Marcora et al., 2005). Depletion of Gln stores can lead to severe complications, such as infection, poor wound healing, impaired immunity, increased intestinal permeability, and finally multiple organ failure (Fenoglio et al., 2008; Lin et al., 2002; Müller et al., 2010; Palmer et al., 1996). Gln appears to be a unique amino acid, serving as a preferred respiratory fuel for rapidly proliferating cells; a regulator of acid–base balance through the production of urinary ammonia; a carrier of nitrogen between tissues; and an important precursor of nucleic acids, nucleotides, amino sugars, and proteins (Bort et al., 2010; Colombo et al., 2010; Gao et al., 2009). Gln is a key amino acid used to produce urea in liver (Darmaun, 2000). Studies have demonstrated a remarkable dependence of the hepatocyte function by Gln (Yang et al., 2010). These researchers have indicated that Gln via increasing activity of antioxidant enzymes and/or glutathione (GSH) level decrease oxidative stress (Babu et al., 2001; Mates et al., 2002). Further investigations to determine the mechanisms underlying this protective effect in chemical-induced toxicity are required (Kawada et al., 2009). However, no attention was paid to the effects of Gln in hepatoprotection against TCDD; and also the information regarding Gln upon micronucleus (MN) in hepatocyte cells remains unknown. In our present study, we examined the effects on histological structure of Gln in TCDD-induced liver injury. We also evaluated the role of Gln on antioxidant activities (enzymatic and nonenzymatic) and MN rates after TCDD treatment.
Materials and methods
Animals
Experiments were performed on male Sprague-Dawley rats, 8 weeks old, weighing 165–190 g. The animals were kept on a 12-h light–dark cycle and allowed free access to food and water. All experiments were performed in accordance with the Guide for the Care and Use of Laboratory Animals (National Research Council,1996). The study was approved by the local ethics committee.
Experimental design
The animals were randomly divided into four equal groups (1) controls (only dimethyl sulfoxide (DMSO)), (2) rats that received 8 μg/kg body weight (b.w.) TCDD (CAS No. 1746-01-6, Sigma-Aldrich®, St Louis, MO, USA) intraperitoneally (i.p.) for 21 days; (3) rats that received 15 mg/kg b.w. Gln (CAS No. 56-85-9, Sigma-Aldrich®) i.p. for 21 days; (4) rats that were pretreated with Gln 30 min prior to i.p. administration of TCDD for 21 days. The doses were selected according to the literature data (Bergamini et al., 1995; Czepiel et al., 2010; Pereira et al., 2007). After the following treatments of all chemicals, the animals were anesthetized with ether and the liver samples were obtained. Liver tissues were separated into three portions. The first part was used for the determination of antioxidant enzyme activities and GSH level, the second part was used for MN analysis, and the third part was used for histological examinations.
Biochemical studies
The tissue preparation was performed according to the procedure previously described by Skrzydlewska and Farbiszewski (1997). The liver samples placed in iced sodium chloride (NaCl) (0.15 M), perfused with the same solution in order to remove the blood cells, then blotted on filter paper, weighed and homogenized in 9 volume (vol) of ice-cold 0.25 M sucrose and 0.15 M NaCl with addition of 6 pl of 250 mM butylated hydroxytoluene (BHT) in ethanol to prevent the formation of peroxides during the assay. The homogenization procedure was performed under standardized conditions. Homogenates (10%) were centrifuged at 10,000g for 15 min at 4°C, and the supernatant was kept on ice until assayed. The protein content in the liver was measured by following the method of Lowry et al. (1951). Superoxide dismutase (SOD) activity in supernatant was measured according to the method of Misra and Fridovich (1972). The catalase (CAT) activities of liver tissues were measured according to the method of Aebi (1984). Liver glutathione peroxidase (GSH-Px) activity was determined according to the method of Lawrence and Burk (1976). The measurement of GSH in liver tissue was performed as described by Akerboom and Sies (1981).
Liver micronucleus assay
Liver MN assay was done using the method of Suzuki et al. (2009). Isolated hepatocytes from rats were prepared by the collagenase perfusion technique (Wang et al., 2002). Liver was perfused through the hepatic portal vein with calcium-free Hank’s balanced salt solution to remove blood for about 10 min at a flow rate of 2.5 ml/min. As soon as the liver becomes grayish brown in color, a second buffer solution containing collagenase (Hank’s balanced salt supplemented with 4 mM calcium chloride and 0.5 mg collagenase/ml) was perfused at the same rate until the liver appeared to have broken. Then the liver was minced into 3- to 4-mm pieces with a sterile scalpel. Following mechanical dissociation, the cells were filtered through gauze and centrifuged at 1350 rpm for 5 min. The hepatocyte pellets were suspended in 10% neutral-buffered formalin and stored under refrigeration. Immediately prior to evaluation, 10–20 μl of hepatocyte suspension was mixed with an equal volume of acridine orange (AO)–DAPI (4′,6-diamidino-2-phenylindole dihydrochloride) stain solution (AO, 0.5 mg/ml; DAPI, 10 μg/ml) for fluorescent staining. Approximately 10–20 μl of the mixture was dropped onto a glass slide and covered with a cover glass. Samples of well-isolated hepatocytes were evaluated with the aid of a fluorescence microscope for counting the number of mononucleated hepatocytes (MNHEPs) in 2000 hepatocytes for each animal. MNHEPs were defined as hepatocytes with round or distinct MNs that stained like the nucleus, with a diameter 1/4 or less than that of the nucleus, and confirmed by focusing up and down, taking into account hepatocyte thickness.
Histological studies
Rats were killed by ether and immediately dissected. Livers were used for slide preparations. The liver samples were fixed in 10% formaldehyde and were embedded in paraffin for microscopic examination. Then the sections obtained from paraffin blocks (5 μm) were stained with hematoxylin–eosin (HE).
Statistics
The experimental data were analyzed using one-way analysis of variance (ANOVA) and Fischer’s least significant difference (LSD) tests to determine whether any treatment significantly differed from the controls or each others. Results presented as mean ± SD values and the level of 0.05 were regarded statistically significant.
Results
Table 1 shows the effects of Gln on biochemical parameters in the tissue homogenates of all experimental animal groups. The hepatic antioxidant activity of SOD, CAT, GSH-Px, and GSH content significantly decreased (p < 0.05) in the group administered with TCDD (8 μg/kg b.w.). The animals treated with Gln (15 mg/kg) showed significant increases in the levels of GSH and also in the activity of GSH-Px (p < 0.05). However, administration of Gln could not restore the reduced activities of SOD and CAT by TCDD.
Changes in the activities of superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GSH-Px), and the level of reduced glutathione (GSH) in liver of rats treated with Gln, TCDD, and their combination a
Gln:
aEach value represents mean ± SD of five animals; means in the same column followed by the different letters present significant differences at the p < 0.05 level.
Table 2 shows the results of the liver MN assay in Sprague-Dawley rats. The tested dose of TCDD (8 μg/kg b.w.) induced a statistically significant increase in the formation of MNHEPs although Gln did not change the MNHEPs number as compared to the control group. Moreover, Gln minimized the increased MNHEPs rates by TCDD.
Results of liver MN assay in rats treated with Gln, TCDD, and their combination a
Gln:
aEach value represents mean ± SD of five animals; means in the same column followed by the different letters present significant differences at the p < 0.05 level.
In controls, microscopic examination showed liver parenchyma consisting of normal hepatocytes. Sinusoids and central vein were also clearly visible (Figure 1). The livers in TCDD-treated group revealed severe pathological damages such as parenchymal degenerations, sinusoidal dilatation, and lipid accumulation (Figure 2a and b). When compared to controls group, the Gln did not lead to a histopathologic finding in liver samples (data not shown). Also, the presence of Gln with TCDD prevented hepatic lesions. Only, the nucleus of hepatocytes became dark stained. And Gln displayed a strong protective effect against TCDD, and the livers of rats were similar to controls (Figure 3).
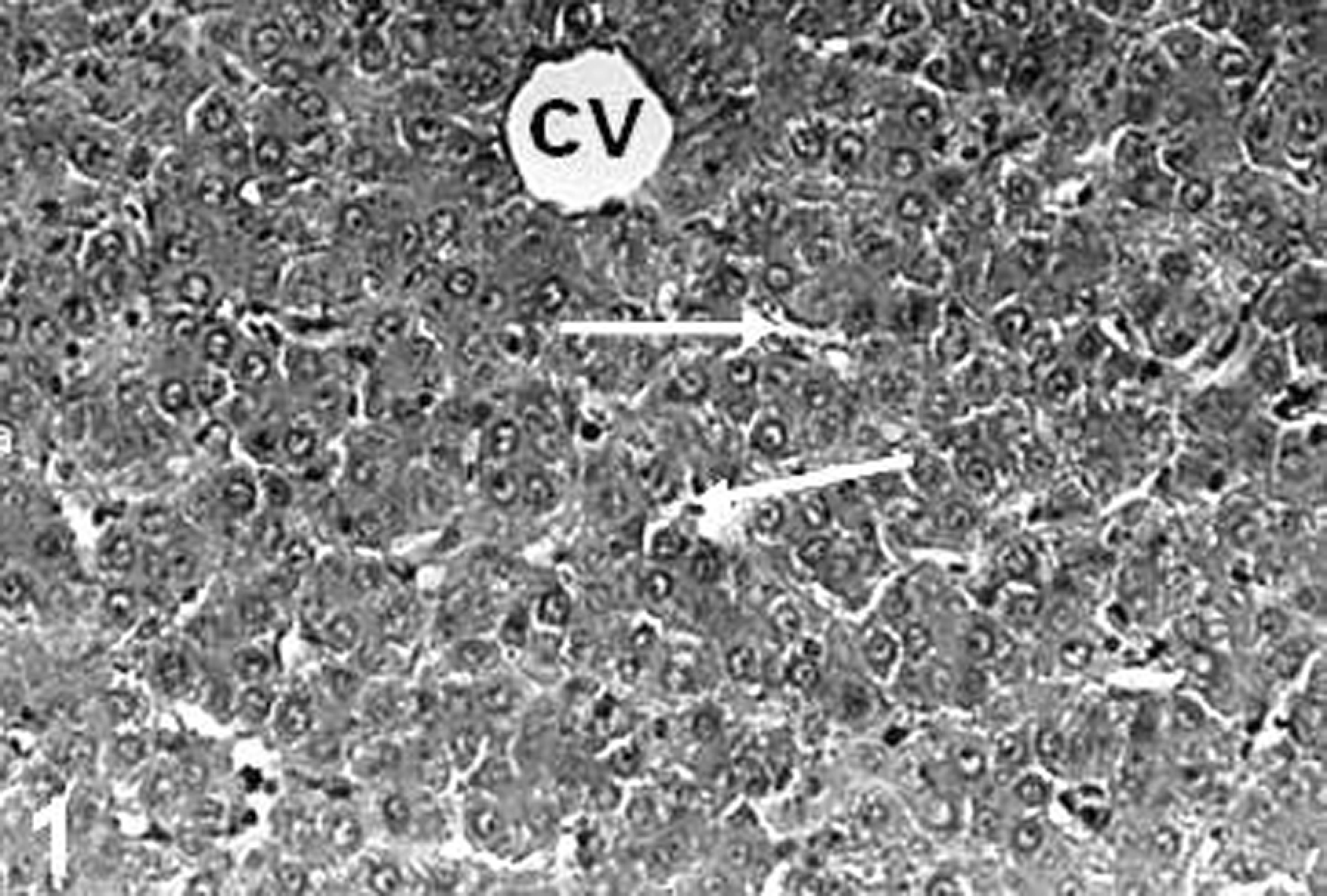
The liver of control rats. Sinusoid (arrows), central vein ((CV) hematoxylin–eosin (HE); ×68).
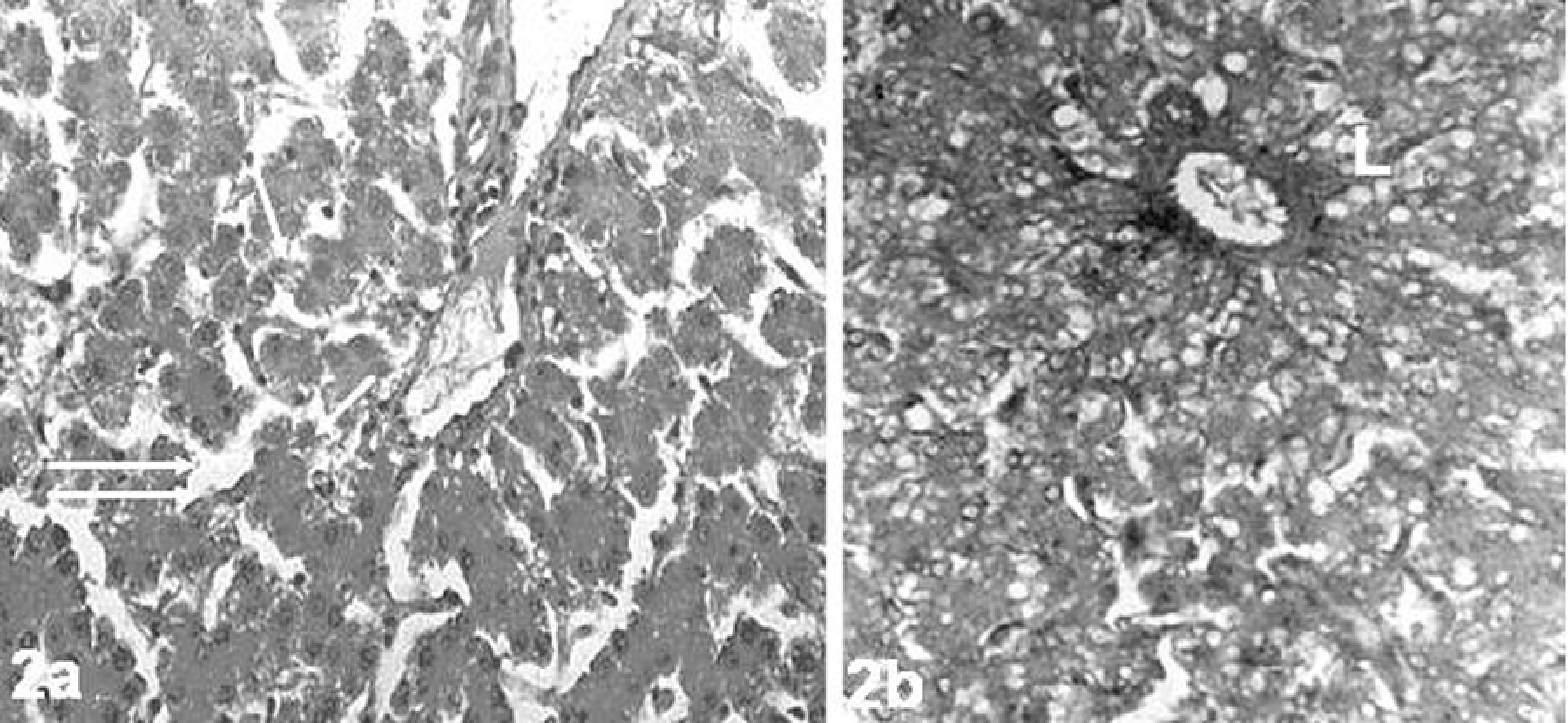
The liver of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)-treated rats. (a) Sinusoidal dilatation (double arrow), parenchymal degenerations (arrows) and (b) lipid accumulations ((L) hematoxylin–eosin (HE); ×68).
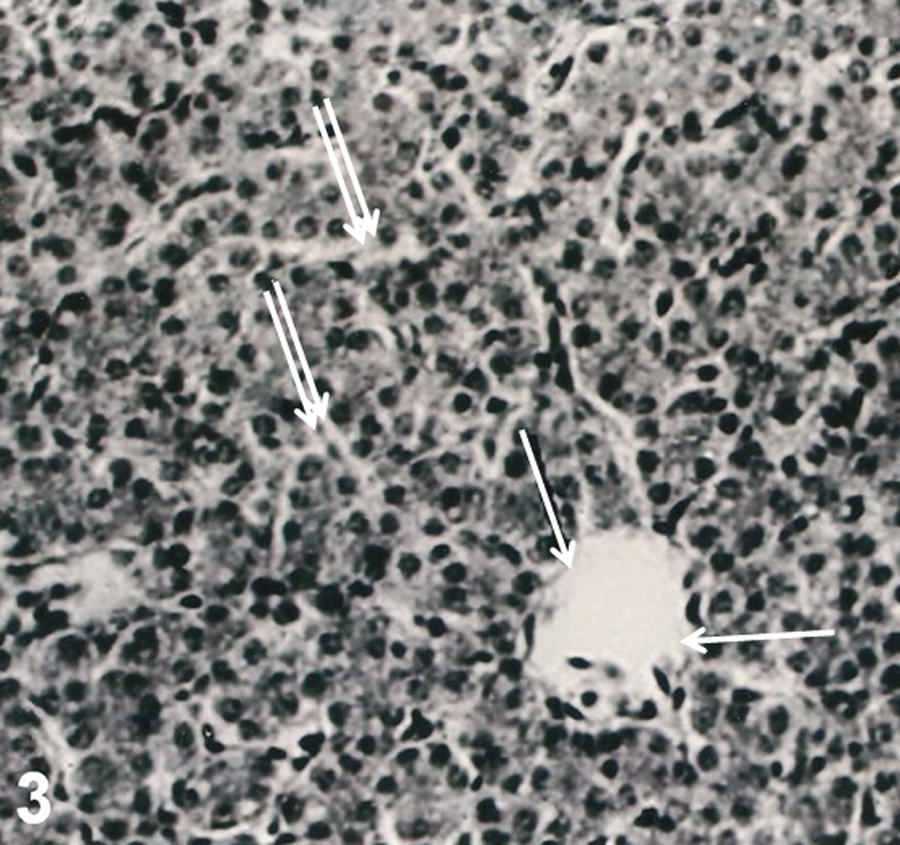
The liver of rats after combined treatments with 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) and
Discussion
The present study examined the effect of Gln treatment on liver injury resulting from TCDD. Our results indicate that although Gln prevented hepatic lesions, it could especially play a beneficial role by preventing MNs and changes in antioxidant status. The exposures of mice and rats to TCDD have resulted in increased production of reactive oxygen species (ROS) and lipid peroxidation (LPO) (Hassoun et al., 2000; Jin et al., 2008; Reyes-Hernánde et al., 2010; Shertzer, 2010). The greater expression of oxidative stress in liver exposed to TCDD might be due to inactivation of the antioxidant enzymes (Jin et al., 2008). It has been suggested that free radicals and ROS are quenched by amino acids, such as Gln (Roth et al., 2002). Overproduction of ROS and nitrogen species can tip the oxidant/antioxidant balance, resulting in the destruction of cell membranes, proteins, and DNA (Park et al., 2010). Therefore, under conditions of increased oxidative stress, dietary antioxidants become critical in maintaining a desirable oxidant–antioxidant balance.
Our results revealed that Gln alone in hepatic tissue exhibited significant antioxidant activity. Increased expression of GSH observed herein, was in agreement with the previous rat models for liver (Szijarto, 2008). The present study explored the ability of 15 mg/kg dose of Gln as a protective agent that induces improvement in the antioxidant capacity against TCDD. Gln supplementation resulted in particular protection of lipid/membrane structures in hepatocytes after TCDD treatments, since (i) the ratio of GSH in Gln/TCDD hepatocytes was reestablished to control/Gln levels; (ii) significant differences were observed in the activity of GSH-Px in the livers of Gln-fed animals and TCDD. Gln led increases of GSH-Px activity, alone and in TCDD models, with value of 28.37% and 22.83%, respectively. This means that Gln had significantly high antioxidant activity in liver. This is not surprising since Gln has been reported as an efficient scavenger and quencher of ROS in lipid bilayers (Kumar and Anandan, 2007). Therefore, Gln provides enhancement of the immune response (Müller et al., 2010) and protection against diseases such as cancer through scavenging of oxygen radicals (Thebault et al., 2010). However, the exact mechanisms of the protective effect of Gln against injury of organs and tissues are still incompletely understood (Jia et al., 2006). Our present findings suggested that the antioxidative protection from Gln was due to GSH and/or enhance enzymatic antioxidant defense against TCDD. GSH is one of the most abundant intracellular antioxidants in animal cells (Kidd, 1997) and detoxifies endogenous ROS and/or exogenous oxidative damage (Schauer et al., 2004). In female B6C3F1 mice, exposure to TCDD results in a significant depletion of GSH by oxidative stress in liver (Slezak et al., 2000). The observed decreases in GSH-related enzyme activities indicate that TCDD may induce oxidative stress in rat liver at relevant low doses of TCDD by altering GSH metabolic mechanisms at the cellular level. GSH-Px enzyme metabolizes peroxides such as hydrogen peroxide (H2O2) and protects cell membranes from LPO (Twaroski et al., 2001). It is proposed that amino acids may protect against different aspects of toxicity by serving as precursors for GSH, or by converting into tricarboxylic acid cycle intermediates is considered an important substrate for energy production (Ralpha et al., 2010). The energy potential of Gln is almost equivalent to that of glucose (Nelson and Cox, 2008). The liver is the major organ of Gln consumption. Gln provides carbons for energy and gluconeogenesis, nitrogen for ureagenesis, and substrate for nucleotide and GSH biosynthesis in order to support cell repair and detoxification reactions (Austgen et al., 1991). It is reported that the GSH content of cells becomes depleted if they are deprived of energy substrates. Thus, by providing an energy source, and by acting as a precursor to GSH, Gln may increase the GSH content of hepatocytes and protect them from toxic substances (Ralpha et al., 2010). An in vitro study by Babu et al. (2001) found that Gln prevented damage to the liver, and this was possibly mediated by GSH synthesis. Gln-supplemented nutrition significantly preserved hepatic GSH in an animal model of pre-infusion with Gln and hepatic injury (Hong et al., 1992). It was reported that there is a significant correlation between the Gln supply and intracellular GSH content (Roth et al., 2002). The administration of Gln reduced liver injury after bile duct ligation in the model of obstructive jaundice (Margaritis et al., 2005). In addition, preceding ingestion of a Gln suppressed liver injury in
The pathological effects in hepatic tissue of TCDD might also lead to disruption in the functional integrity of hepatocytes (Boverhof et al., 2006; Czepiel et al., 2010; Kopec et al., 2010; Sakamoto et al., 1995). In the present study, the TCDD elicited severe instances of liver damage (hepatocyte degeneration, sinusoidal dilatation, and lipid accumulation). However, Gln has pronounced effect on the relative potency of TCDD. Our findings not only supported that Gln was potent hepatoprotective agent but also explained the action of Gln on MNs. It was reported that the gene and chromosome data that could be phenotypically anchored to complementary histopathology could be used to further elucidate the mechanisms involved in the adaptive and toxic responses elicited by TCDD (e.g. lipid accumulation; N'Jai et al., 2008). TCDD-induced oxidative stress caused single-strand breaks in DNA from liver and brain tissues of rats (Hassoun et al., 2000, 2001). In our investigation, MN test was performed for the first time and is revealed that TCDD exposure increased the rate of MNs in hepatocytes. Micronucleus assay is a reliable test which indicated the chromosomal damage (Yilmaz et al., 2008), and MN production is one of the key pieces of evidence for the possible involvement of antioxidant activity in oxidative DNA damage (Tung-Kwang et al., 2010). ROS can alter vital cell components such as polyunsaturated fatty acids, proteins, and nucleic acids (Halliwell and Gutteridge, 1990). The increased production of ROS, LPO, and DNA and membrane damage are always associated with TCDD exposure (Shertzer, 2010). Gln acted as a potent oxygen radical scavenger shown to be beneficial to carcinoma prognosis (Todorova et al., 2010). Recently, Gln has been reported to possess anticancer properties (Halder et al., 2010; Kaufmann et al., 2008). Gln decreased DNA damage in breast cancer cells’ response to radiation in young healthy females since it is an important precursor of nucleic acids (Cano et al., 2010). And the amide nitrogen of Gln is the major component for the biosynthesis of nucleotides (e.g. purines and pyrimidines in DNA and RNA; Neu et al., 2002). Again, Gln appears to be a unique amino acid, an important precursor of amino sugars, and proteins (Gao et al., 2009).
Collectively, Gln treatment offered beneficial effects against TCDD-induced injury in hepatic tissue. The observed effect of Gln (at 15 mg/kg dose) on antioxidant status and MNs was reflected by a major protection against histopathological alterations. The data were essential to properly address the usage of Gln as a nutritional therapy for the prevention of TCDD toxicity.
Footnotes
Conflict of interest
The authors declared no conflicts of interest.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.