
Abstract
Cyclophosphamide (CP) is an alkylating agent that has been considered effective for cancer treatment. Lycopene, the pigment in tomato fruits, has beneficial effect in the treatment of some diseases. The goal of this study is to evaluate the protective effect of lycopene alone or combined with melatonin (Mel) in inhibiting the oxidative stress and toxic effect of CP in rats. Five groups of rats were included in this study; Group I served as the control. Rats in groups II-V were administrated with single dose of CP (150 mg/kg B.W) interperitoneally for 3 days. On the same day of CP administration, the rats in group III were fed a diet supplemented with lycopene (50 mg/kg of diet), rats in group IV were administered with a dose of 2.5 mg Mel/kg body weight (bw) injected subcutaneously and rats in group V were supplemented with lycopene and a dose of 2.5 mg Mel/kg bw injected subcutaneously. After 15 days the blood samples were collected. Results obtained showed that CP exerted its toxic effect by increasing the free radicals and reactive oxygen species that causes lipid peroxidation and cell damage, and this in turn is detected by elevation in nitric oxide (NO) and malondialdehyde (MDA), while the activities of antioxidants enzymes including (superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPx)) were significantly decreased as compared with the control rats. The combined treatment (Lyco + Mel) group showed potential reduction in these parameters more than those treated with lyco alone. The activities of SOD, CAT and GPx were found significantly high than lyco alone treated rats. A positive significant correlation between NO and MDA (r = .81). In conclusion, these results suggested that supplementation of diet with lycopene and Mel provided antioxidant defense with strong chemopreventive activity against Cp-induced cytotoxicity.
Introduction
Human body is equipped with various antioxidants such as superoxide dismutase (SOD), glutathione peroxidase (GPx), catalase (CAT), glutathione (GSH), ascorbic acid (vitamin C), α-tocopherol (Vitamin E), and so on, which can counteract the deleterious action of reactive oxygen species (ROS) and protect from cellular and molecular damage (Grinberg-Funes et al., 1990).
Cyclophosphamide (CP) is an alkylating drug that has been considered effective in the treatment of many cancer diseases. However, this drug may induce acute inflammation of the urinary bladder if precautions are not taken (Ahluwalia et al., 1994, Alfieri and Gardner, 1997). ROS, such as the nitric oxide radical (·NO), are included in the pathogenesis of CP-induced cystitis. It has also been reported that increase in ROS, such as NO, leads to bladder edema, inflammation and extravasation (Frasier et al., 1991). It was reported that CP-induced testicular oxidative stress manifested by a significant inhibition of peroxidase and CAT enzyme activities, as well as high levels of malondialdehyde (MDA) and conjugated dienes in the testis (Moncada et al., 1991).
Various molecules can inhibit the formation of free radicals associated with many diseases. Bioactive compounds from plant origin have the potential to subside the biochemical imbalances induced by various toxicity associated with free radicals. They provide protection without causing any side effects and therefore, development of drugs from plant products is desired. For that account natural antioxidants from plant sources have been viewed as promising therapeutic drugs (Pfeilschnifter et al., 1992).
Lycopene, the pigment principally found in tomato fruits and tomato products, has received much attention because of its beneficial effect in the treatment of many diseases (Pfeilschnifter et al., 1992). It was demonstrated that lycopene provided the best protection against singlet oxygen-induced cell damage (Regoli et al., 1994). Epidemiologic studies revealed that lycopene may show inverse relationships to cancer as with other common carotenoids (Maggi, 1997).
Melatonin (Mel), secreted by the pineal gland, is a powerful antioxidant that not only scavenges the hydroxyl radical (Souza-Filho et al., 1997) but also inhibits the production of NO synthase. Mel can enter the nucleus where it protects DNA from oxidation damage, thereby decreasing the incidence of cancer (Bray et al., 2004). Also, it modulates the immune response and inhibits the development of hormone-dependent cancer (Aruoma, 1994).
The goal of this study was to evaluate the protective effect of lycopene alone or combined with Mel in inhibiting the oxidative stress and toxic effect induced by CP in rats.
Materials and methods
Animals
A total of 60 male Sprague Dawley rats weighing (100–130 g) (colony of laboratory animals from the King Fahd Medical Research center, Jeddah, Kingdom of Saudi Arabia) were maintained and handled according to the recommendations of the Ethic Committee. Animals were housed in a room maintained at 22°C with a 12-h light–dark cycle with free access to foods. Rats were randomized into five groups (12 rats each); group I rats will served as the control. Rats in groups II-V were administrated with single dose of CP (150 mg/kg B.W) interperitoneally for 3 days (Sigma Chemicals, St Louis, MO, USA) (Cerutti, 1985).On the same day of CP administration, the rats in group III fed a diet supplemented with lycopene (50 mg/kg of diet) (Jagetia and Rao, 2006), rats in group IV were administered a dose of 2.5 mg Mel/kg body weight (bw) injected subcutaneously and the rats in group V were supplemented with lycopene and a dose of 2.5 mg Mel/kg bw injected subcutaneously. Mel was prepared by dissolving 50 mg in 2 ml ethanol and diluted to 100 ml with distilled water to make 0.2% solution (Vijayavel et al., 2006).
After 15 days the blood samples were collected from all animals. Serum was obtained after centrifugation at 12,000g, 10 min and stored at −80°C until analyses.
Biochemical analysis
NO measurement
Serum nitrite and nitrate were analyzed by the modified microassay as described by Vodovotz (1996). Briefly, nitrate was reduced to nitrite by cadmium and assayed by Griess reagent.
Malondialdhyde measurement
MDA was assayed by thiobarbituric acid method as described by Zima et al. (1995).
Assay of SOD activity
The SOD activity was assessed by the nitroblue tetrazolium (NBT) reduction method (Flohe and Otting, 1984); 0.1 ml of tissue supernatant was added to a reaction mixture containing 0.1 mM EDTA (200 µl), 0.12 mM riboflavin (50 µl) and 0.6 M phosphate buffer (pH 7.8) in a final volume of 3 ml. The optical density was measured at 560 nm.
Assay of CAT activity
The reaction mixture (2 ml) contained 1.95 ml of 10 mM H2O2 in 60 mM phosphate buffer (pH 7.0). The reaction was initiated by adding 0.5 ml supernatant to it, and the absorbance was taken for 3 min at 240 nm. Phosphate buffer (60 mM, pH 7.0) was used as a reference. The data are expressed as µmol H2O2 consumed/(min (mg protein) (Claiborne, 1985).
Assay of GPx activity
The activity of GPx was assayed by the method of Rotruck et al. (Ellman, 1959). The reaction mixture containing 0.2 ml of EDTA (0.8 mM, pH 7.0), 0.4 ml of phosphate buffer (10 mM), and 0.2 ml of tissue homogenate incubated with 0.1 M of H2O2 and 0.2 ml of GSH for 10 min. Oxidation of GSH by the enzyme is measured spectrophotometrically at 420 nm. The activity of GPx was expressed as µmol GSH oxidized/min/mg protein.
Statistical analysis
Data are presented as the mean ± standard deviation (SD). One-way analysis of variance (ANOVA) followed by Tukey multiple comparison method will be carried out to compare the mean value of different groups using SPSS 8 student version. Comparisons will made between group I with groups II, III, IV and group II with groups III and V. P < .05 was considered statistically significant.
Results
The protective effect of lycopene alone or combined with Mel against CP-intoxicated rats are shown in Table 1 Tables (1 and Table 2). In the CP-intoxicated group serum ALP and LDH were significantly increased as compared to the control group (p < 0.001). The elevated activities of serum ALP and LDH were significantly reduced in the animal groups treated with lycopene or Mele or combined (p < 0.001).
The level of total GSH in the control and CP-treated rats of different ages are shown in Table 2. In untreated rats, total GSH was decreased in CP-injected rats as compared with the control group. When CP was administered the total GSH decreased by 40%. The variations in the values for oxidized glutathione (GSSG) and thiobarbituric acid reactive substance (TBARS) levels in the kidney of rats injected with CP indicative of oxidative status and lipid peroxidation.
Serum malondialdhyde (MDA) and nitric oxide (NO) levels in all studied groups (mean ± SD).

Lyco: lycopene; Mel: melatonin; NS: nonsignificant.
* p < 0.05.
** p < 0.01.
*** p < 0.001.
aGroups II, III and IV compared with control group I.
bGroups III and IV compared with control group II.
The activities of erythrocyte superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GSPx) in all studied groups (mean ± SD).
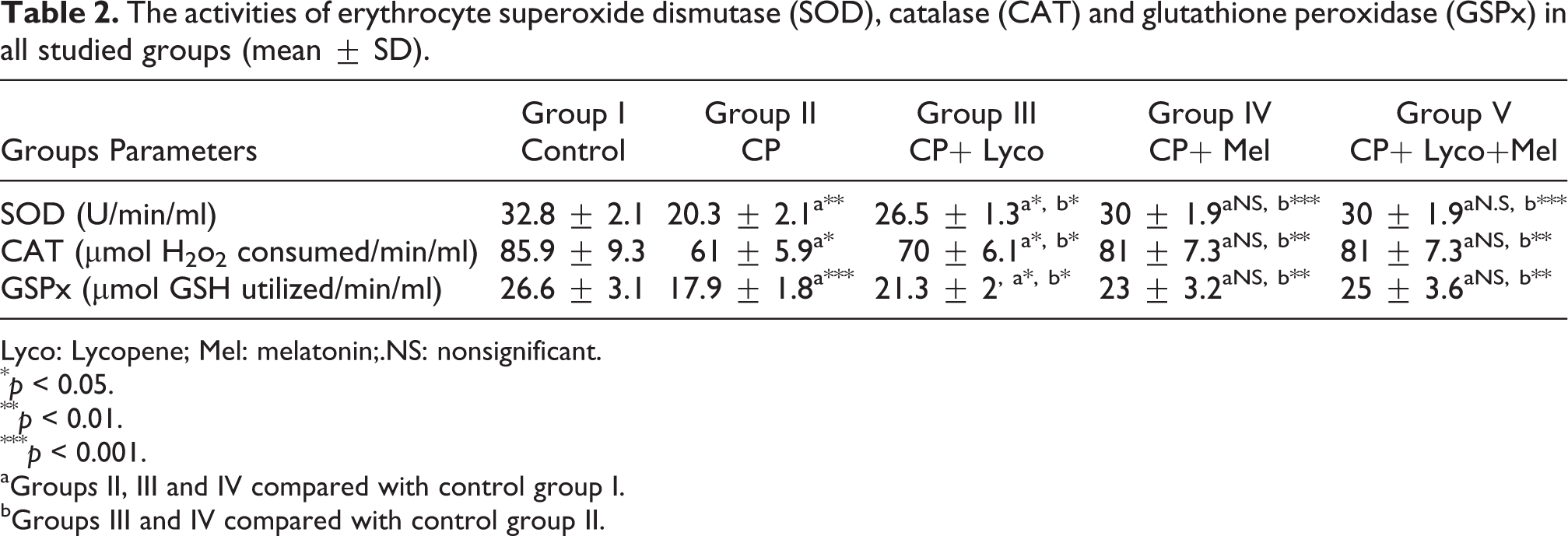
Lyco: Lycopene; Mel: melatonin;.NS: nonsignificant.
* p < 0.05.
** p < 0.01.
*** p < 0.001.
aGroups II, III and IV compared with control group I.
bGroups III and IV compared with control group II.
The effects of CP treatment on the activities of SOD, CAT and GPx were also assessed. In lyco-treated rats, SOD activity increased significantly (65%). When CP was given to the rats, SOD activity was significantly reduced (80%, as compared with its controls), while it was significantly elevated when lycopene was given for 10 days. Regarding CAT activity, following CP injection, the CAT activity significantly decreased (75%) in rats and significantly increased (65%) in lyco- and Mel-treated rats in comparison with the untreated animals.
Rats injected with CP reduced GPx activity as compared with control group, but the CP toxicity was ameliorated when lyco and Mel was given as compared with the untreated rats.
Discussion
CP is an alkylating agent with a wide spectrum of clinical uses that has been proved to be effective in the treatment of cancer and nonmalignant disease states. However, this drug may induce acute inflammation if precautions are not taken.
In the present study, CP increases superoxide anion and/or H2O2 formation in cultured rat hepatocytes (Fuhramn et al., 1997) and different kidney cells (Hussein et al., 2005) and it induces oxidative stress and lipoperoxidation in microsomal preparations from humans (Reiter et al., 1999).The deleterious toxic effects of CP is, at least in part, due to the increased production of free radicals and ROS.
The erythrocyte has a number of characteristics consistent with oxidative injury, and many studies have examined the effect of CP on the oxidative status in mammalian tissues. It has been associated with enhanced ROS generation and oxidative stress (Maestroni and Conti, 1989). The large increases in both GSSG and TBARS concentrations observed in CP injected rats, as compared with control rats, are in favor of a strong oxidative stress and enhanced ROS formation in rats. Similar increases in TBARS have been previously reported (Astrog et al., 1997). Nevertheless, it has also been reported that TBARS contents decrease in rats treated with epicatchin, suggesting that decreased lipoperoxidation in the kidney is not an inevitable consequence of CP.
Regarding antioxidant defenses, our data showed that the concentration of reduced GSH progressively increased in Lyco- and Mel-treated rats. Thus, the oxidative stress observed in rats could be related not only to the marked increase in the GSSG/GSH molar ratio but also to the decline in GSH levels. Increased generation of ROS and lipid peroxides has been reported following administration of GSH-depleting agents and after a reduction in liver GSH levels (Rao and Agarwal, 1999).
The Lyco enhanced the antioxidant enzymes, which may be an attempt by the organism to counterbalance the decreases in the reducing power mediated by GSH. However, in senescent animals, the total antioxidant capacity of kidney cells should not be sufficient to scavenge the ROS generated.
Lyco in human plasma has a half life of about 2–3days (Joseph et al., 2001). Only a few metabolites, such as 5,6-dihydroxy-5,6-dihydro lycopene, have been detected in human plasma (Bray et al., 2004). Owing to its lipophilic nature, lycopene was found to concentrate in LDL and VLDL fractions and not in HDL fraction of the serum (Aruoma, 1994).
The reduced activity of SOD and CAT in CP injected rats could be a consequence of inhibitory effects due to excess generation of ROS. SOD is inhibited by hydrogen peroxide (Gerster, 1997), and CAT by an excess of superoxide radical (Clinton, 1998) and CP enhances the formation of ROS, in turn (Cohen et al., 1978). A similar phenomenon has previously been reported to occur in isolated peroxisomes from rat liver and in the rabbit kidney after CP treatment, as well as under conditions of oxidative stress induced by chronic ethanol ingestion or liver parasitism (Tamarkin et al., 1981). Inhibition of the catalytic activities of proteins that express SOD and CAT activities could also be a consequence of the stronger CP-induced liver GSH depletion in old animals. It has been shown that CP reduces the content of protein sulfhydryl groups and causes protein thiol oxidation in rat liver cells (Ahluwalia et al., 1994), probably due to the CP-induced GSH depletion. Acausative mechanism proposed for the enzyme inhibition was observed (Alfieri et al., 1997). A causative mechanism proposed for the inhibition observed in other enzymes (Alfieri et al., 1997). CP might also reduce the protein levels of antioxidant enzymes, because a marked inhibitory effect of the drug on the synthesis of protein in the rat liver has been demonstrated (Frasier et al., 1991). The oxidative stress and enhanced TBARS formation were accompanied by decreased protein levels of SOD and CAT. Importantly, the CP-induced decreases in the enzymatic and nonenzymatic antioxidant defense systems in rats makes them more susceptible to oxidative damage and compromise the antioxidant capacity of the liver to adequately scavenge the ROS generated during CP therapy.
Our data indicated that either GSH depletion or disturbances in the GSH redox status appear in CP-treated rats. In addition, kidney is more susceptible to CP-induced oxidative stress, lipoperoxidative injury, enhanced GSH depletion and decreased antioxidant enzyme activities may contribute to this effect.
Oxidative stress induced due to the generation of free radicals and/or decreased antioxidant level in the target cells and tissues has been suggested to play an important role in carcinogenesis (Clinton, 1998). During cell membrane damage, various enzymes leak down to the circulatory fluid and their assessment in serum serves as markers in clinical studies. SOD is the first antioxidant enzyme to deal with oxyradicals by accelerating the dismutation of superoxide to hydrogen peroxide CAT is a peroxisomal hem protein that catalyses the removal of hydrogen peroxide formed during the reaction catalyzed by SOD. Thus, SOD and CAT acts mutually supportive antioxidative enzymes that provide protective defense against ROS (Cohen et al., 1978).
This study in the line of Cohen et al (Tamarkin et al., 1981), they reported that, Mel and retinoic acid have potential anticancer effect due to their ability to act as antioxidant or inhibit the production of NO by reducing NO synthase.
In conclusion, the toxicity following administration of CP to the rat appeared to ameliorate by Lyco and Mel by scavenging the free radicals and increasing the antioxidant activities.
Footnotes
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.