
Abstract
The protection afforded by melatonin (MLT) against diazinon (DZN)-induced micronucleus formation, an index of DNA damage, in human blood lymphocytes was investigated. Whole blood samples were collected from five volunteers and were incubated with MLT at different concentrations (100, 200, 300, and 400 µM final concentration) for 1 h. The samples were then incubated with 750 µM DZN for 1 h. Subsequently, the lymphocytes were cultured with a mitogenic stimulant to evaluate micronucleus formation in cytokinesis-blocked binucleated cells. The incubation of lymphocytes with DZN induces additional genotoxicity. Pretreatment with MLT at these doses significantly reduced the micronucleus frequency in cultured lymphocytes (p < 0.05–p < 0.0001). The maximum decrease in the frequency of micronuclei was observed at 400 µM of MLT, which caused a reduction of 87%. MLT also exhibited an excellent and dose-dependent radical-scavenging activity against 1,1-diphenyl-2-picrylhydrazyl free radicals. Our study revealed that MLT has a potent antigenotoxic effect against DZN-induced DNA damage, which may be due to the scavenging of free radicals and increased antioxidant status. Because MLT is a natural compound and is considered safe, it can be used as a supplement to protect people exposed to chemical or environmental hazards.
Introduction
There are many reports regarding the capability of hazardous environmental chemicals inducing genotoxic and carcinogenic effects on the mammary. The main mechanism is producing free radical species that induces damage to critical macromolecules such as DNA to promote chronic diseases such as cancers. (Tiwari, 2001). Diazinon (DZN; diethoxy-((2-isopropyl-6-methyl-4-pyrimidinyl)oxy)-thioxophosphorane) is a commonly used organophosphorus (OP) pesticide. It is synthetic chemical substance with broad spectrum insecticide activity (Sarabia et al., 2009). The toxic effects of DZN are due to the inhibition of the activity of acetylcholinesterase, an enzyme needed for proper nervous system function. DZN has been widely used throughout the world in agriculture and horticulture to control insects that feed on crops, ornamentals, lawns, fruits, vegetables, and other food products (El-Shenawy et al., 2010). Some reports have been published regarding DZN and its effects on the hematological and biochemical parameters in animal models (El-Shenawy et al., 2009, Kalender et al., 2005, 2006, Yehia et al., 2007). The toxicity of the OP insecticide DZN causes adverse effects on many organs (Kalender et al., 2005). Studies of DZN’s mutagenicity—its ability to cause genetic damage—showed that DZN can damage DNA in human blood cells, cells from laboratory animals, and bacteria (Grover et al., 2003). DZN exposure was found to increase the occurrence of a type of genetic damage called micronuclei (MN). MN may be induced by strand breaks in DNA due to oxidative stress (Fenech, 1993). The MN test using peripheral blood cells is used to detect chromosome breaks caused by DNA damage (Igarashi and Shimada, 1997).
Systems that protect the living body from oxidative stress-induced damage include the enzymes superoxide dismutase, catalase, and glutathione peroxidase, which remove reactive oxygen species (ROS); the proteins ferritin and transferrin, which mask heavy metal ions; antioxidants, including vitamin E and vitamin C; enzyme repair systems for damaged DNA; and the ubiquitin–proteasome system, which removes damaged proteins (Limón-Pacheco and Gonsebatt, 2009). Although the human body is equipped with self-defense mechanisms, such as detoxification processes involving various enzymes, exposure to high levels of dangerous chemicals can lead to mutagenic and carcinogenic events (Tiwari, 2001). The consumption of vegetables and fruits has been an effective strategy for reducing the genotoxicity and carcinogenicity induced by hazardous chemicals (Aune et al., 2009). The incidence of cancer is significantly reduced by the consumption of natural products (Gullett et al., 2010). The preventive effects of natural products are due to their antioxidant and free radical-scavenging activities and their effects on cellular signaling pathways (Ahmadi et al., 2008, Kim et al., 2011).
Melatonin (MLT; N-acetyl-5-methoxytryptamine), a pineal secretory product, influences circadian rhythmicity by acting on the suprachiasmatic nucleus (Reiter, 1991). MLT possesses strong antioxidant activity by which it protects cells, tissues, and organs from oxidative damage by ROS, especially the hydroxyl radical
Because MLT has excellent antioxidative properties, it is likely that MLT can protect against the toxicity of DZN; that is, an elevated level of MLT in the body may be prophylactic against damage. Therefore, this study was undertaken to assess the effects of MLT against genotoxicity induced by DZN in human peripheral blood lymphocyte using the micronucleus test.
Materials and methods
Chemicals
MLT was obtained from Sigma (St Louis, Missouri, USA). 1,1-Diphenyl-2-picrylhydrazyl radical (DPPH) was purchased from Sigma Chemicals Co. Butylated hydroxyanisole was purchased from Merck (Darmstadt, Germany). All chemicals were obtained from Merck and Sigma and were used without further purification.
Human blood samples
The protocol for this study was approved by the Research Committee of Mazandaran University of Medical Sciences, Sari, Iran. Informed consent was obtained from five healthy, nonsmoking human volunteers, who were all men aged between 25 and 35 years. Whole blood samples (7 mL) were collected in heparinized tubes and divided into seven 1-mL tubes. The blood samples were treated with 50 µL of MLT solution to give final concentrations of 100, 200, 300, and 400 µM and were then incubated for 1 h at 37°C. Subsequently, 750 µM of DZN was added to the blood samples, which were then incubated for 1 h. The positive control tube contained only DZN, and the control sample received no treatment. Another control group was treated with only high concentration of 400 µM MLT to observe the safety of the high concentration on human lymphocyte. After incubation, the blood samples were transferred for the micronucleus assay.
Micronucleus assay
Each sample of 0.5-mL duplicate aliquots were added to 4.5 mL of Roswell Park Memorial Institute (RPMI) 1640 culture medium (GIBCO, Grand Island, NY, USA) containing 20% fetal calf serum, 20 µL/mL phytohemagglutinin (Gibco), 50 U/mL penicillin, 50 µg/mL streptomycin, and 2 mM glutamine (Sigma). All cultures were incubated at 37 ± 1°C in a humidified atmosphere of 5% carbon dioxide and 95% air. Cytochalasin B (final concentration: 6 µL/mL) was added after 44 h of culture. After 72 h of incubation, the cells were collected by centrifugation for 8 min at 1000 r/min, resuspended in 0.075 M cold potassium chloride, and then immediately treated with a fixative solution (methanol:acetic acid, 6:1) three times. The fixed cells were dropped onto clean microscopic slides, air-dried, and stained with Giemsa solution. All slides were evaluated at 40× magnification to determine the frequency of MN in cytokinesis-blocked binucleated cells with well-preserved cytoplasm. Image of typical binucleated cells with and without a micronucleus are shown in Figure 1. The criteria for scoring MN were between 1/16th and 1/4th of the diameter of the main nuclei, a nonrefractile nature, no link to the primary nucleus, and no overlap with the primary nucleus (Hosseinimehr et al., 2009). Additionally, the two main nuclei of the cell had to be completely divided for the cell to be countable as a binucleated cell. In each blood sample group and for each volunteer, 1000 binucleated cells were scored from the treated and control cultures in duplicate to determine the frequency of micronuclei.
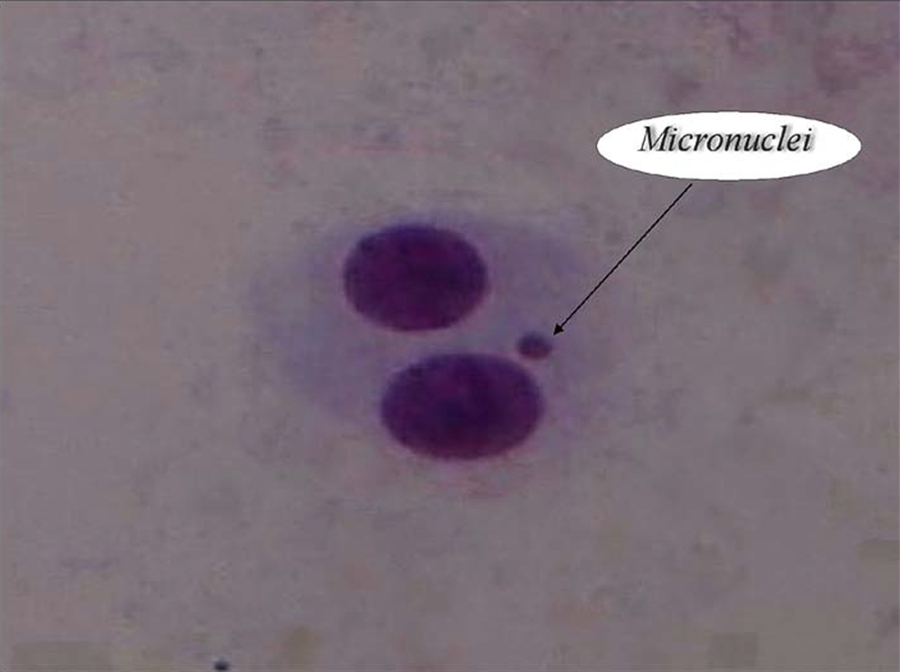
A typical binucleated lymphocyte with micronuclei.
Measurement of the free radical-scavenging activity of MLT
The free radical-scavenging capacity of MLT was determined based on the bleaching of the stable DPPH (Ahmadi et al., 2010). Different concentrations of MLT (0.0125–0.8 mg/mL) were added, in a volume of 1 mL, to 3 mL of a methanolic solution of DPPH (10 mg/250 mL). After 15 min of incubation at room temperature in the dark, the absorbance was recorded at 517 nm. The experiment was performed in triplicate. Butylated hydroxytoluene was used a standard antioxidant agent. The percentage of scavenged free radicals was calculated using the equation:
Statistical analysis
For each volunteer, the number of MN was recorded for each group. The data are presented as the mean ± SD. The data were analyzed using analysis of variance with Tukey’s honestly significant difference post hoc test. The value of p < 0.05 was considered significant.
Results
The data presented in Figure 2 show that there was a significant difference in the percentage of micronucleated binucleated cells between lymphocytes treated with DZN and the control cells (p < 0.0001). The percentage of MN in the lymphocytes of volunteers treated with 750 µM DZN was 9.32 ± 0.63%, whereas the percentage in nontreated control lymphocytes was 0.94 ± 0.11% (Table 1). MLT had a potent protective action against the DNA damage induced by DZN. The frequencies of MN in groups of cells that were pretreated with MLT at doses of 100, 200, 300, and 400 µM were 5.1 ± 0.7, 3 ± 0.33, 1.7 ± 0.44, and 1.18 ± 0.14%, respectively (Table 1). The results for MLT at all doses were significantly better than the result for the DZN sample (Figure 2). The total micronucleated binucleated cell values were 45, 67, 81, and 87% less for 100, 200, 300, and 400 µM concentrations of MLT, respectively, as compared to the controls. The maximum protective effect of MLT, which was observed at 400 µM, completely prevented the formation of MN induced by DZN and protected human lymphocytes against DZN-induced DNA damage (p < 0.0001). The protective effect of MLT and the reduction of the frequency of MN by MLT increased with increasing concentration.
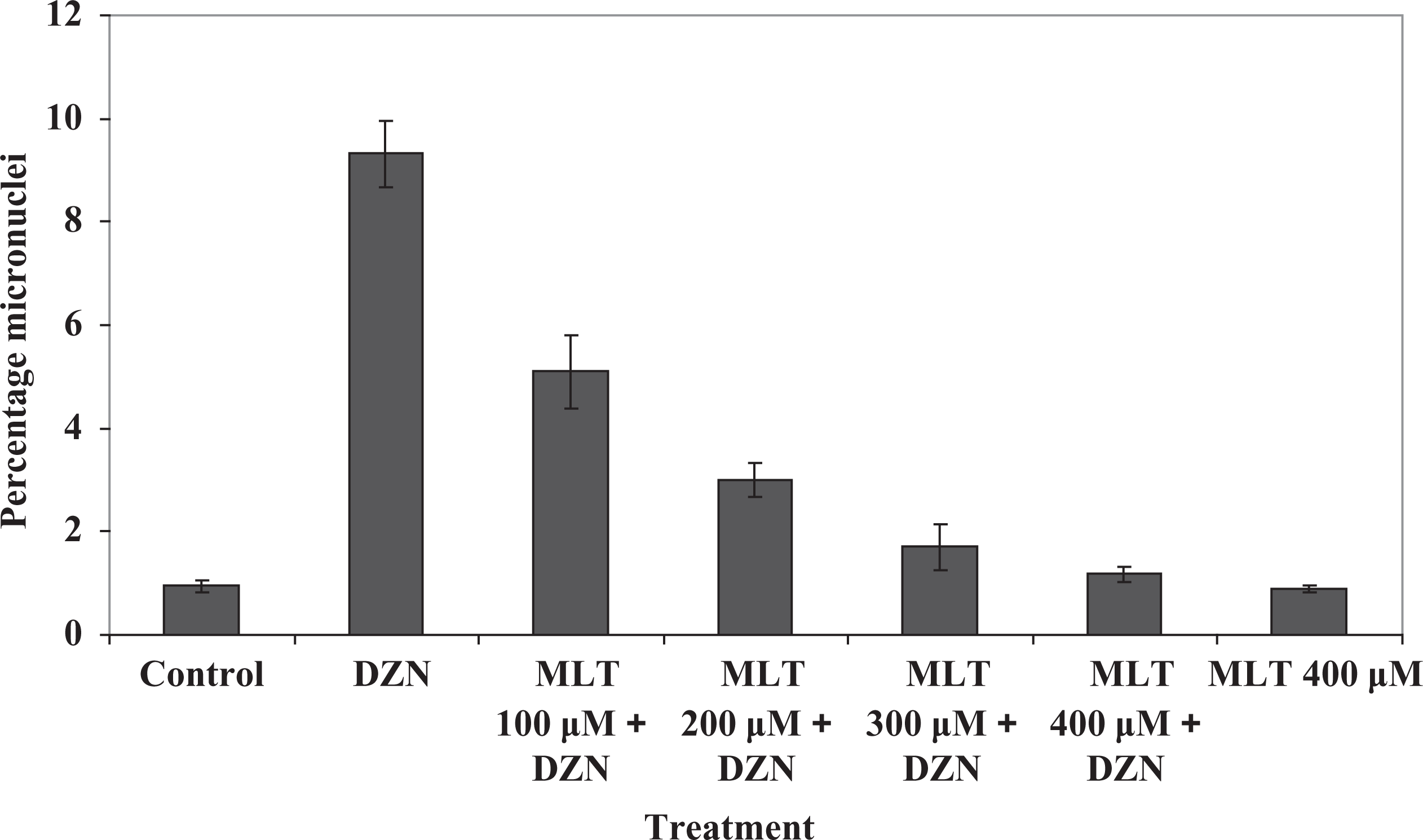
In vitro protection by MLT at different concentrations (100, 200, 300, and 400 µM) against genetic damage induced by 750 µM DZN in cultured whole blood lymphocyte. The data represent the mean ± SD of five human volunteers. p < 0.0001: Control sample compared with similarly administrated lymphocytes from the blood sample treated with DZN. p < 0.0001: DZN sample compared with DZN + MLT 300 and DZN + MLT 400 µM. p < 0.001: DZN sample compared with DZN + MLT 200 µM. p < 0.05: DZN sample compared to DZN + MLT 100 µM. p > 0.05: Control sample compared with similarly lymphocytes from the blood sample treated with MLT 400 µM. The data were analyzed using one-way ANOVA analysis and Tukey’s HSD test. MLT: melatonin; DZN: diazinon; ANOVA: analysis of variance; HSD: honestly significant difference.
The total number of MN induced in vitro by 750 µM DZN in cultured blood lymphocytes from human volunteers and the possible protective effects of different concentrations of MLT.
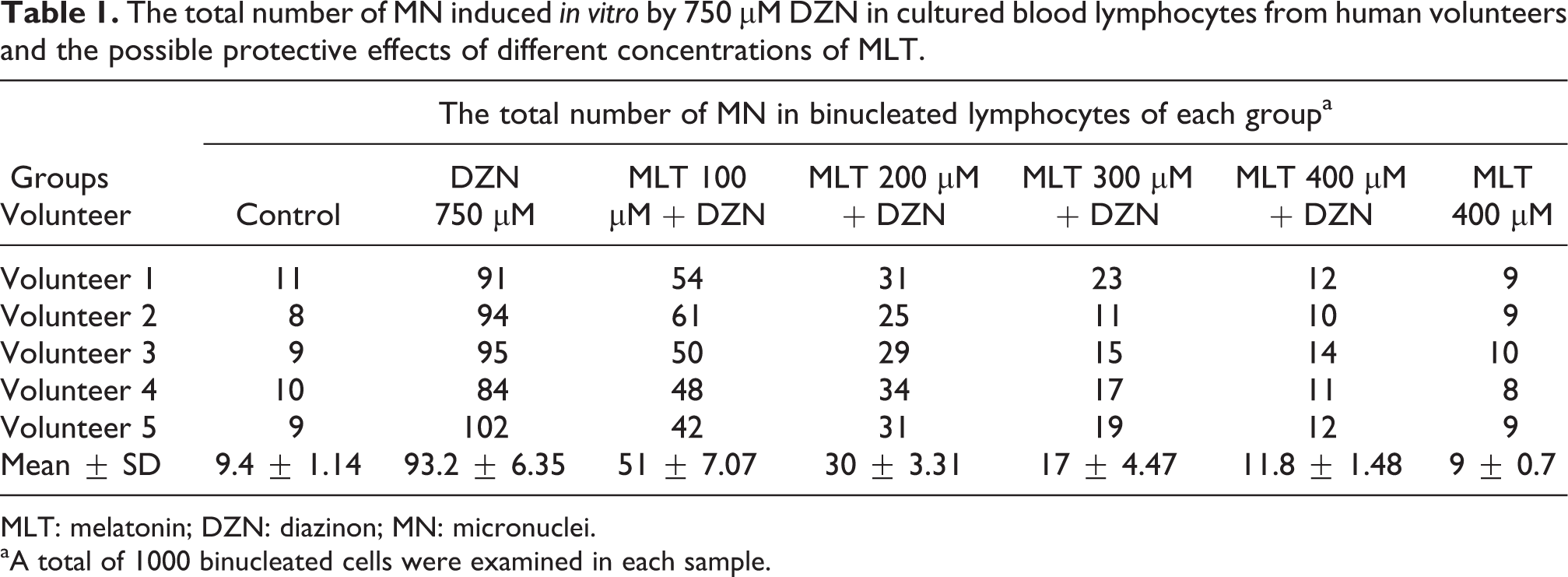
MLT: melatonin; DZN: diazinon; MN: micronuclei.
aA total of 1000 binucleated cells were examined in each sample.
The scavenging effects of MLT on DPPH radicals increased with increasing concentrations, reaching 98.6% at 0.8 mg/mL (Figure 3). Therefore, MLT has excellent free radical-scavenging and antioxidant activities.
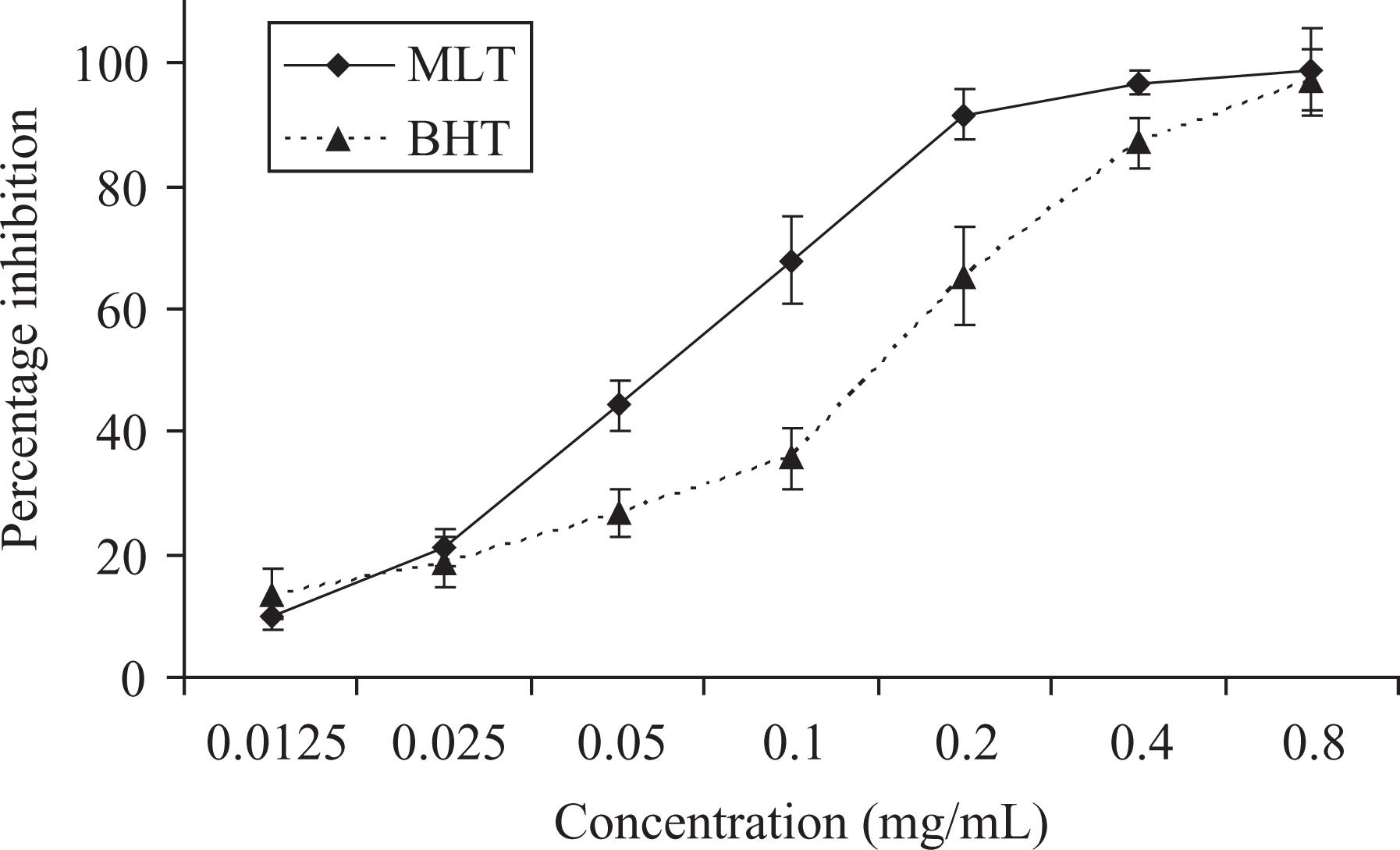
Scavenging effect of different concentrations of MLT and BHT on DPPH free radical at 517 nm. BHT: butylated hydroxytoluene; MLT: melatonin; DPPH: 1,1-diphenyl-2-picrylhydrazyl.
Discussion
Our results indicated that MLT pretreatment prevented the genotoxicity induced by DZN in human blood lymphocytes. DZN is one of the most commonly used OPs that are used to control a range of crop pests. This compound is also used as a veterinary ectoparasiticide in many countries. DZN is easily degraded, but under conditions of low temperature, low moisture, high alkalinity, and a lack of suitable microbiological degraders, it may remain biologically active in soils for 6 months or longer (Koprucu et al., 2006). The results of many studies have confirmed that DZN exposure induces DNA damage. Using the MN assay, the treatment of human blood cells (lymphocytes and erythrocytes) and skin fibroblasts with DZN has shown to result in a significant increase in the number of MN via a clastogenic mode of action, inducing single- and double-stranded DNA breaks (Čolović et al., 2010).
In the last several years, the MN assay has been extensively used to assess the cytogenetic damage induced by chemicals and radiation. With the improvements introduced by Fenech and Morley, this assay appears to be sufficiently sensitive, simple, and fast to detect agents that induce chromosome damage (Fenech and Morley, 1985). This method is now one of the best established in vivo and in vitro cytogenetic assays in the field of genotoxic biomonitoring (Decordier et al., 2009). In the present study, DNA damage was evaluated by the MN test. The administration of DZN resulted in DNA damage and induced MN formation in human blood lymphocytes. It is evident that the preincubation of lymphocytes with MLT for 1 h prevented DNA damage in the human lymphocytes. MLT at all doses used in this study reduced DZN-induced MN formation, which was used as a DNA damage index. These results suggest that MLT has a potent protective effect against DZN-induced genotoxicity.
The protective effect of the pineal secretory product MLT against oxidative damage has been examined in both in vivo (Melchiorri et al., 1994) and in vitro studies (Melchiorri et al., 1996). MLT also stimulates the endogenous antioxidative enzyme glutathione peroxidase, thereby contributing to enhanced cellular defenses against oxidative stress. Moreover, MLT has been shown to protect DNA from a physical agent, ionizing radiation (Pablos et al., 1995), and from a chemical toxin, lipopolysaccharide (Reiter et al., 1997), both of which are believed to damage the genetic material by free radical-based mechanisms. In a previous study, MLT was shown to protect the lungs and livers of paraquat-treated rats from oxidative stress, and MLT also reduced mortality following treatment with this herbicide (Melchiorri et al., 1994).
The primary reason for the induction of DNA damage is the increasing intracellular level of ROS; these toxic species potentially interact with macromolecules. The main characteristic of antioxidants is the ability to trap free radicals. Free radicals may oxidize nucleic acids, proteins, lipids, and DNA and can initiate degenerative diseases (Pan et al., 2011). It appears that the primary mechanism of chemoprotective agents is the direct and/or indirect interaction with these ROS. An excellent scavenging effect was observed for MLT. It appears that MLT scavenged and trapped free radicals induced by DZN and, through this mechanism, it could prevent the damage induced by free radicals. Therefore, MLT prevented the DNA damage and the formation of MN that could have been induced by free radicals produced by DZN in human lymphocytes. Protective effect of MLT is likely due to the antioxidative properties of the indolamine.
Due to the high lipophilicity of the MLT molecule and its low molecular weight (Reiter et al., 2007), this secretory product easily passes through cell membranes and affords on-site protection against locally generated free radicals directly at DNA sites. In fact, existing evidence suggests that there is preferential compartmentalization of MLT in the nucleus relative to the cytosol and cell membranes (Menendez-Pelaez et al., 2004).
In conclusion, MLT had a potent protective effect against the genotoxicity induced by DZN in human blood lymphocyte cells, and this protective effect may be the result of antioxidant activity and free radical-scavenging properties. Because MLT is safe and provides on-site protection against locally generated free radicals directly at DNA sites; this compound can be a worthwhile supplement to help defend the body against the negative effects, particularly DNA damage, induced by chemical and environmental hazards.
Footnotes
Conflict of interest
The authors declared no conflicts of interest.
Funding
This study was approved and supported by a grant from the Mazandaran University of Medical Sciences, Sari, Iran.