
Abstract
The study was aimed at evaluating the protective role of α-lipoic acid (ALA) on long-term exposure of rats to the combination of chlorpyrifos (CPF) and deltamethrin (DLT). Forty-two (42) male Wistar rats were divided into 6 exposure groups with 7 animals in each group: (I) soya oil (2 ml kg−1), (II) ALA (60 mg kg−1), (III) DLT (6.25 mg kg−1), (IV) CPF (4.75 mg kg−1), (V) (CPF + DLT) DLT (6.25 mg kg−1) and CPF (4.75 mg kg−1; 1/20th of the previously determined median lethal dose) and (VI) (ALA + CPF + DLT) pretreated with ALA (60 mg kg−1) and then co-exposed to CPF and DLT, 45 min later. The regimens were administered by gavage once daily for a period of 16 weeks. Sera obtained from blood collected at the end of the experimental period were used for the evaluation of serum glucose, total protein, albumin, urea, creatinine and the activities of alanine aminotransferase, aspartate aminotransferase, alkaline phosphatase, lactate dehydrogenase and acetylcholinesterase. The liver homogenate was used to assay for the activities of superoxide dismutase and glutathione peroxidase and the concentrations of malondialdehyde, cytokine and tumour necrotic factor α. The result showed that the combination of CPF and DLT resulted in marked alterations of these biochemical parameters in most cases compared to either of the pesticides singly, supplementation with ALA ameliorated these alterations.
Introduction
The need to improve agricultural yield and public health standards has resulted in widespread use of pesticides. The initial use of pesticides has been very effective in reducing pest infestation and increasing agricultural production and productivity. However, over time targeted pests have developed resistance to pesticides, necessitating increasing application or the use of several pesticides combination or mixtures to combat resistance by pest (Shonga et al., 2013). Mixing or combining pesticides with different modes of action may delay resistance development within pest population (Cloyd, 2010).
Pesticide combinations often applied in agricultural and public health programmes include organophosphates (OPs) and pyrethroids. The mixing of OP and pyrethroid classes of insecticides, which has become very common in the markets, has resulted in an increase in the prevalence of toxicity of pesticide combinations (El-Halwagy and Zaki, 2009). In the last few decades, various environmental problems have led to increased concern about potential toxicity from exposure to multiple chemicals, especially pesticides, including their residues detected in food or water (Yang et al., 1989). Recently, attention has been drawn not only to the effect of particularly toxic xenobiotics but also to their interaction with one another.
OPs and pyrethroids are the two most commonly used classes of pesticides worldwide accounting for 50% (Casida and Quistad, 2004) and 30% (Prasanthi et al., 2005), respectively, of all insecticide applications globally. Chlorpyrifos (CPF) and deltamethrin (DLT) are OP and pyrethroid insecticides, respectively, that have been widely investigated singly. However, their mechanisms of action, which involve the inhibition of acetylcholinesterase (AChE) (Goel et al., 2005) and impairing of ion transport on voltage-sensitive sodium channel (Soderlund et al., 2002), respectively, do not explain all the symptoms of their intoxication (Ojha and Srivastava, 2012). Oxidative stress, a putative molecular mechanism, which occurs when the production of reactive oxygen species (ROS) overrides the antioxidant capacity in the target cell resulting in oxidative damage has been recognized as one of the primary causes of subcellular toxicity of pesticides (Banerjee et al., 2001).
The presence of multiple chemical contaminants in the environment has made it necessary to investigate the effect of pesticides combination on humans and animals and to identify agents that may mitigate the adverse health consequence posed by long-term exposures.
Alpha-lipoic acid (ALA), a thiol compound naturally in plants and animals, possesses potent antioxidant properties (Goraca and Jozefowicz-Okonkwo, 2007) and has been reported to reach all areas of the central and peripheral nervous systems (Packer et al., 1997). ALA is a powerful metabolic antioxidant that is able to scavenge ROS and recycle other antioxidants in the body including vitamin C, vitamin E and glutathione (Wollin and Jones, 2003). The present study was aimed at evaluating the biochemical changes induced by chronic exposure to the combination of low level of CPF and DLT in rats and the ameliorating effect of ALA.
Materials and methods
Experimental animals
Young male Wistar rats weighing between 120 g and 150 g used for this experiment were obtained from the laboratory animal house of the Department of Veterinary Pharmacology and Toxicology, Ahmadu Bello University, Zaria, Nigeria. The animals were allowed to acclimatize for at least 2 weeks in the laboratory prior to the commencement of the experiment and were given access to standard rat chow and water ad libitum. The study was carried out according to the specification of the Ahmadu Bello University Animal Research Committee and Guide to the Care and Use of Laboratory Animals (1996). Ethical permission was also sort from the university’s ethical committee for experimentation.
Chemical preparation
CPF 20% emulsifiable concentration (EC), marketed as Termiphos® (Sabero Organics, Gujarat Limited, Sarigam, Gujarat, India) and DLT 1.25% EC marketed as Deltaforce® (Sabero Organics) were prepared by reconstituting in soya oil (S/oil; Grand Cereals and Oil Mills Ltd, Jos, Nigeria). ALA (600 mg/tablet; marketed and distributed by General Nutrition Corporation, Pittsburgh, Pennsylvania, USA) was obtained from a pharmaceutical store in Abuja, Nigeria and dissolved in S/oil, just prior to its daily administration.
Experimental protocol
The rats were weighed and randomly assigned into six groups of seven rats each. Group I (S/oil) served as the control and was given S/oil only at a dose of 2 ml kg−1, whilst group II (ALA) was given ALA only at a dose of 60 mg kg−1 (El-Halwagy and Hassanin, 2006). Group III (DLT) was administered with DLT only at a dose of 6.25 mg kg−1 (corresponding to 1/20th median lethal dose (LD50) value: 125 mg kg−1 determined previously by Uchendu et al. (2014)), whilst group IV (CPF) was administered with CPF only at a dose of 4.75 mg kg−1 (corresponding to 1/20th LD50 value: 95 mg kg−1 determined previously by Uchendu et al. (2014)). Group V (CPF + DLT) was co-exposed to CPF (4.75 mg kg−1) and DLT (6.25 mg kg−1), whilst rats in group VI (ALA + CPF + DLT) were pretreated with ALA (60 mg kg−1) and then co-exposed to CPF (4.75 mg kg−1) and DLT (6.25 mg kg−1) 45 min later. The different regimens were administered once daily by gavage for a period of 16 weeks. At the end of this study period, the rats were killed by severing the jugular vein after light ether anaesthesia and blood samples were collected into test tubes, incubated for 30 min and centrifuged at 800g for 10 min. The sera obtained were collected into clean test tubes and used for the evaluation of some serum biochemical parameters and the activities of AChE. The liver from each rat was harvested and homogenized in phosphate-buffered saline and the homogenates were used to assay for concentrations of malondialdehyde (MDA) and tumour necrotic factor (TNF) and the activities of antioxidant enzymes.
Serum biochemical evaluation
The sera samples were used for the evaluation of the concentrations of glucose, total proteins, albumin and activities of aspartate amino transferase (AST), alanine aminotransferase (ALT), alkaline phosphatase (ALP) and lactate dehydrogenase (LDH) using an autoanalyser Selectra XL (Netherlands).
Evaluation of serum AChE activity
The sera samples were also used to evaluate the activities of AChE according to the method of Ellman et al. (1961) using AChE kit obtained from Abcam® (Cambridge Science Park Cambridge CB4 OFL, UK).
Evaluation of hepatic TNF concentration
Cytokine assay was carried out to determine the concentrations of hepatic TNF-α from liver homogenate using a commercial rat TNF enzyme-linked immunosorbent assay kit obtained from Abcam®.
Evaluation of hepatic oxidative stress parameters
The superoxide dismutase (SOD) activity in the hepatic tissue homogenate was measured using the SOD kit from Northwest Life Science Specialties Vancouver LLC (Canada) based on the method described by Martin et al. (1987). The principle was based on the method of monitoring the autoxidation rate of haematoxylin. The SOD activity was determined by measuring ratios of autoxidation rates in the presence and absence of the sample (hepatic tissue homogenate) and the absorbance read at 560 nm after every 10 min and expressed as McCord-Fridovich ‘cytochrome c’ units.
The activity of glutathione peroxidase (GPx) was also determined in the hepatic tissue homogenate using the GPx kit from Northwest Life Science Specialties Vancouver LLC based on the method adapted from Paglia and Valentine (1967). The principle was as follows: GPx catalyses the oxidation of glutathione by cumen hydroperoxide. In the presence of glutathione reductase and nicotinamide adenine dinucleotide phosphate (NADPH), the oxidized glutathione is immediately converted to the reduced form with a concomitant oxidation of NADPH to NADP+. The decrease in absorbance of NADPH was measured at 340 nm.
The MDA concentration in the hepatic tissue homogenate as an index of lipid peroxidation was evaluated using the double heating method of Draper and Hadley (1990) as modified by Yavuz et al. (2004). The principle of the method was based on spectrophotometric measurement of the colour produced during the reaction of thiobarbituric acid (TBA) with MDA. The reaction was read at an absorbance using an ultraviolet spectrophotometer at 532 nm. The concentration of MDA was calculated by the absorbance coefficient of MDA-TBA complex, 1.56 × 105 cm−1 M−1, and expressed as nanomole per milligram protein. The tissue protein concentration in the hepatic tissue was determined using the method of Lowry et al. (1952).
Statistical analysis
Values obtained were expressed as mean ± SEM and subjected to one-way analysis of variance, followed by Tukey’s multiple comparison test using GraphPad Prism version 5.0. Values of p < 0.05 were considered significant. Percentages were used to indicate changes where necessary.
Results
Effect of treatments on glucose and serum proteins
There was a significant (p < 0.05) increase in serum glucose concentration in the CPF group compared to the S/oil, ALA and ALA + CPF + DLT groups (Table 1). Although the serum glucose concentration in the ALA + CPF + DLT group did not differ significantly (p > 0.05) compared to CPF + DLT group, it decreased by 14.0%.
Effect of treatments on biochemical parameters in Wistar rats.

CPF: chlorpyrifos; DLT: deltamethrin; ALA: alpha-lipoic acid; SOD: superoxide dismutase; GPx: glutathione peroxidase; MDA: malondialdehyde; AChE: acetylcholinesterase; TNF: tumour necrotic factor; ANOVA: analysis of variance.
a p: < 0.05: compared to S/oil group.
b p < 0.05: compared to ALA.
c p < 0.05: compared to ALA + CPF + DLT group (one-way ANOVA/Tukey post hoc).
d p < 0.05: compared to DLT.
e p < 0.05: compared to CPF.
f p < 0.05: compared to CPF + DLT.
No significant (p > 0.05) difference in serum total protein concentration was observed between the S/oil group and DLT, CPF or CPF + DLT groups (Figure 1). Serum total protein was decreased significantly (p < 0.05) in the CPF and CPF + DLT groups, compared to the ALA group. The serum total protein concentration in the ALA + CPF + DLT group, however, did not significantly (p > 0.05) differ from that in the CPF + DLT group, it decreased by 8.6%.
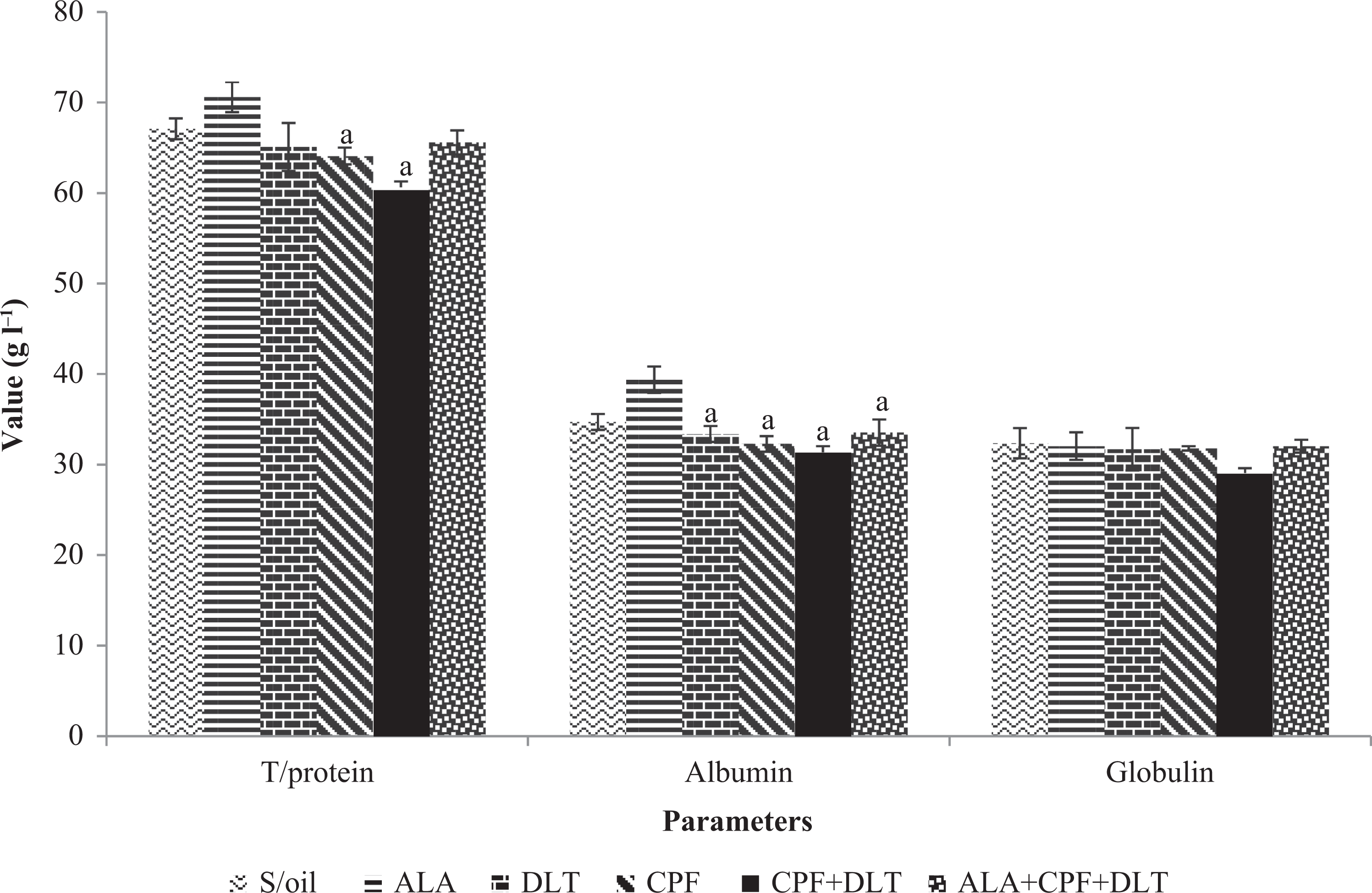
The effect of chronic exposure to S/oil, ALA, CPF and/or DLT and ALA, CPF and DLT (ALA + CPF + DLT) on serum proteins concentration in rats. *p < 0.05: compared to S/oil group; a p < 0.05: compared to ALA; b p < 0.05: compared to DLT; c p < 0.05: compared to CPF; d p < 0.05: compared to CPF + DLT; e p < 0.05: compared to ALA + CPF + DLT group (one-way ANOVA/Tukey post hoc). CPF: chlorpyrifos; DLT: deltamethrin; S/oil: soya oil; ALA: alpha-lipoic acid; ANOVA: analysis of variance.
Serum albumin concentration was significantly lower (p < 0.05) in the CPF, DLT, CPF + DLT or ALA + CPF + DLT groups, compared to the ALA group. Serum albumin concentration increased by 6.9% in the ALA + CPF + DLT groups compared to the CPF + DLT group.
There was no significant (p > 0.05) difference in serum globulin concentration between the groups (Figure 1). There were slight decreases in globulin concentration in the DLT (2.0%), CPF (1.8%) and CPF+DLT (10.3%) groups compared to the S/oil group.
Effect of treatments on liver-associated enzymes
There was a significant (p < 0.05) increase of ALT activity in the CPF, DLT and CPF + DLT groups compared with the S/oil group. The serum ALT activity significantly (p < 0.05) increased in the DLT, CPF, CPF + DLT and ALA + CPF + DLT groups, respectively, compared with the ALA group. However, the ALT activity showed an insignificant decrease in the ALA + CPF + DLT group (17.2%) compared to that of the CPF + DLT group (Figure 2).
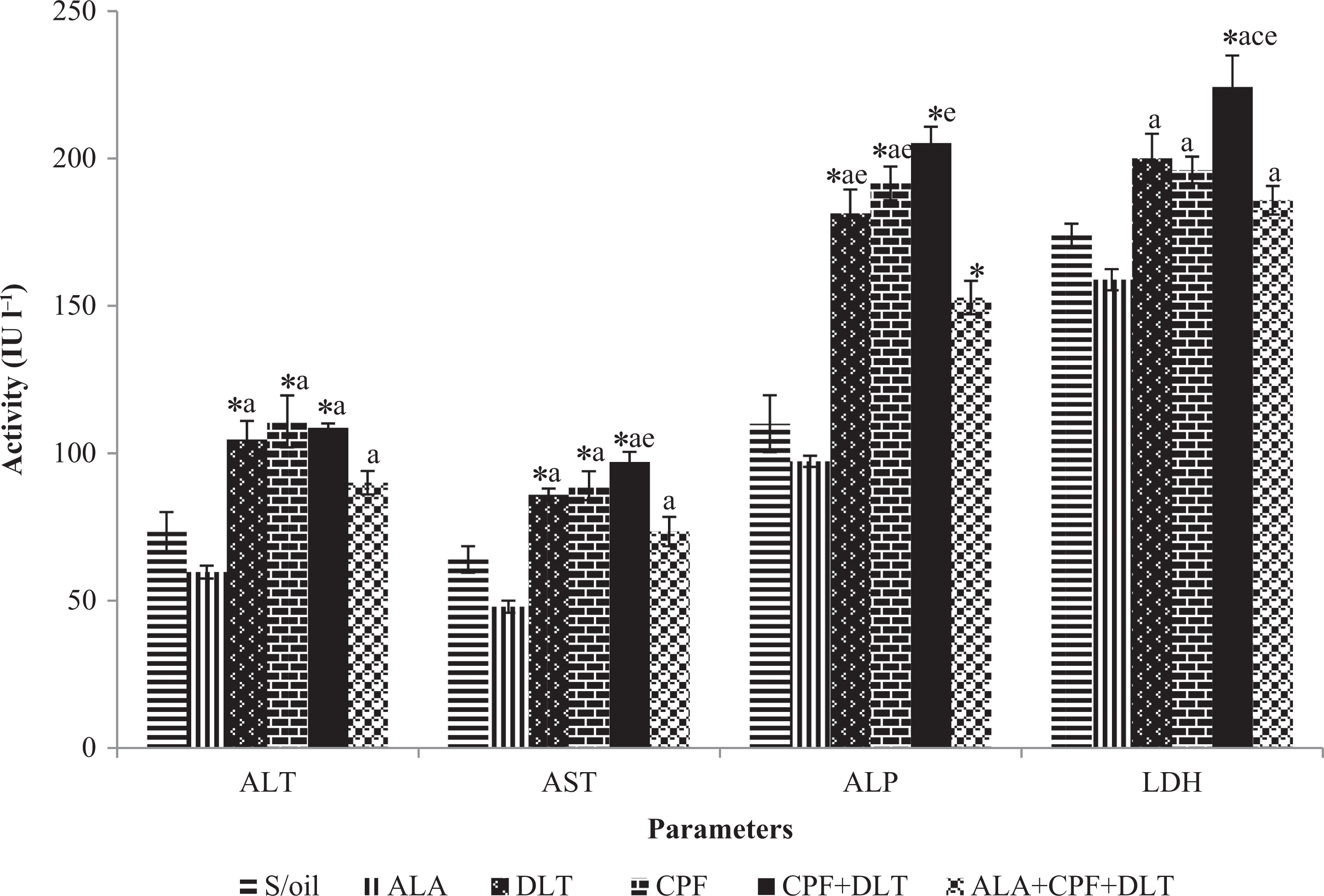
The effect of chronic exposure to S/oil, ALA, CPF and/or DLT and ALA, CPF and DLT (ALA + CPF + DLT) on serum liver enzymes activities in rats. *p < 0.05: compared to S/oil group; a p < 0.05: compared to ALA, b p < 0.05: compared to DLT; c p < 0.05: compared to CPF; d p < 0.05: compared to CPF + DLT; e p < 0.05: compared to ALA + CPF + DLT group (one-way ANOVA/Tukey post hoc). CPF: chlorpyrifos; DLT: deltamethrin; S/oil: soya oil; ALA: alpha-lipoic acid; ANOVA: analysis of variance.
There was a significant (p < 0.05) increase in serum AST activity in the DLT, CPF and CPF + DLT groups, respectively, compared with the S/oil group. The serum AST activity significantly (p < 0.05) increased in the DLT, CPF, CPF + DLT and ALA + CPF + DLT groups compared with the ALA group (Figure 2). There was a significant decrease (p < 0.05) in the ALA + CPF + DLT group compared with the CPF + DLT group.
The serum ALP activity in the DLT, CPF and CPF + DLT groups significantly (p < 0.05) increased compared with the S/oil group (Figure 2). There were also significant (p < 0.05) increases in the ALP activity in the DLT, CPF + DLT and CPF + DLT groups compared with the ALA group. The ALP activity significantly decreased (p < 0.05) in the ALA + CPF + DLT group compared with the CPF + DLT group.
The serum LDH activity in the CPF + DLT group significantly (p < 0.05) increased compared with the S/oil group. The serum LDH activity in the DLT, CPF, CPF + DLT and ALA + CPF + DLT groups was higher (p < 0.05) than that in the ALA group. There was a significant (p < 0.05) elevation in the serum LDH activity of the CPF + DLT group compared to the CPF group. There was a significant (p < 0.05) decrease of LDH activity in the ALA + CPF + DLT group compared to CPF + DLT group (Figure 2).
Effect of treatments on hepatic antioxidant enzymes
The hepatic SOD activity significantly (p < 0.05) decreased in CPF and CPF + DLT groups compared to the S/oil group. The SOD activity in the DLT, CPF, CPF + DLT and ALA + CPF + DLT groups was lower (p < 0.05) than that in the ALA group. There was, however, a significant increase (p < 0.05) in the CPF + DLT group compared to the ALA + CPF + DLT group (Table 1).
There was no significant (p > 0.05) difference in hepatic GPx activity between S/oil group and DLT, CPF and CPF + DLT groups. There was, however, a significant decrease (p < 0.05) of hepatic GPx activity in the DLT and CPF + DLT groups compared with the ALA group. The hepatic GPx activity in the ALA + CPF + DLT group increased by 20.6% compared with the CPF + DLT group (Table 1).
Hepatic MDA concentration rose (p < 0.05) in the CPF + DLT group compared to that of either the S/oil or ALA group. There were significant (p < 0.05) increases in hepatic MDA concentration in the DLT and CPF groups compared with the ALA group. A decrease of liver MDA concentration was observed in the ALA + CPF + DLT (39.6%) group compared to the CPF + DLT group (Table 1).
Effect of treatments on serum AChE and hepatic TNF concentration
There was a significant (p < 0.05) decrease of AChE activity in the DLT, CPF, CPF + DLT and ALA + CPF + DLT groups compared, respectively, to the S/oil and ALA groups. The AChE activity also decreased in the DLT (43%) and CPF + DLT (56.8%) groups, respectively, compared to the S/oil group. The AChE activity in the ALA + CPF + DLT group was higher by 38% than that of the CPF + DLT group (Table 1).
There were significant (p < 0.05) increases in hepatic TNF-α concentration in the DLT, CPF, CPF + DLT and ALA + CPF + DLT groups, respectively, compared to that of S/oil or ALA group. The hepatic TNF-α concentration significantly increased (p < 0.05) in the CPF + DLT group compared to either DLT or CPF group. There was a reduction in hepatic TNF-α concentration (p < 0.05) in the ALA + CPF + DLT group compared to that of the CPF and CPF + DLT groups, respectively (Table 1).
Discussion
Environmental pollutants such as pesticides are known to induce a broad spectrum of toxicological effects and biochemical dysfunctions culminating into serious health hazards. The present study revealed that chronic co-exposure to CPF and DLT resulted in hyperglycaemia. The hyperglycaemia observed was notably higher in the CPF group compared to either DLT or the co-exposed group. The increase in glucose level by pesticides treatment may indicate disrupted carbohydrate metabolism due to enhanced breakdown of liver glycogen (Raja et al., 1992) or probably due to the activation of the hypothalamus–pituitary–adrenal axis (sympathetic nervous system) during pesticide-evoked stress. This activation results in the secretion of glucocorticoid from the adrenal cortex which in turn increases blood glucose by induction of gluconeogenesis pathway (Rahimi and Abdollahi, 2007). Hyperglycaemia has been reported previously by several workers following DLT (Eraslan et al., 2007; Yousef et al., 2006) and CPF (Acker and Nogueira, 2012; Elsharkawy et al., 2013) intoxication. Hyperglycaemia has also been attributed to the inhibition of AChE by OPs, which can disturb the endocrine regulation of glucose (Matin and Siddiqui, 1982).
Pretreatment with ALA reduced the hyperglycaemia induced by CPF + DLT intoxication. This may be due to the ability of ALA to increase glucose utilization at both normal glucose and high glucose concentration and also to reduce protein glycosylation and lipid peroxidation (Jain and Lim, 2000) induced by CPF + DLT exposure.
The present study also revealed that chronic co-exposure to DLT and CPF caused a decrease in serum total protein concentration, apparently due to hypoalbuminaemia. The reduction in serum protein may also be attributed in part to the damaging effect of DLT and CPF on the liver cells, as confirmed by increased activities of liver-associated enzymes in the present study. Hypoproteinaemia has been reported in several studies following DLT (El-Magraby et al., 2010; Yousef et al., 2006) and CPF (Khan and Kour, 2007; Mansour and Mossa, 2010) poisoning in rats. The mild hypoalbuminaemia observed is consistent with the results obtained in previous studies following either DLT (Manna et al., 2004; Yousef et al., 2006) or CPF (Khan and Kour, 2007; Mansour and Mossa, 2010) intoxication. Hypoalbuminaemia may be due to a decrease in albumin synthesis by the liver as a result of hepatic impairment and/or increased urinary excretion due to impaired renal function. Yousef et al. (2006) suggested that the hypoproteinaemia and hypoalbuminaemia may reflect hepatocellular injury and disturbed protein and amino acid metabolism induced by pesticides.
Pretreatment with ALA ameliorated the deficit in serum total protein and albumin, induced by co-exposure to DLT and CPF, apparently due to the reduction of the induced oxidative stress, as ALA is known to enhance cellular antioxidant defence. Al-Attar (2010) also demonstrated in a study the ability of ALA to restore the hypoproteinaemia and hypoalbuminaemia induced by an OP, malathion. The apparent increase in total protein and albumin following pretreatment may be due to the radical scavenging and antioxidant recycling ability of ALA, which reduced LPO and protected the liver and kidneys from oxidative assaults induced by the pesticides.
The study revealed that exposure to CPF + DLT caused an increase in the activities of liver-associated enzymes such as AST, ALT, ALP, and LDH. The increase in AST activity observed was notably higher in the co-exposed group compared to that of the single pesticide exposure groups. Elevation in serum ALT activity was higher in the CPF group than either the co-exposed or DLT group. Increases in serum ALT activity have been reported by several workers in rats exposed to DLT (Aydin, 2011; Chargui et al., 2012) and CPF (Goel et al., 2005; Uzun and Kalender, 2013). ALT is highly liver specific and one of the most reliable indicators of hepatotoxic damage. The increase in activity of transferases may be due to pathological changes in the liver and other visceral and muscles involved in the production of these enzymes (Manna et al., 2004). Elevated activity of serum ALP is not limited to liver damage only; it is also associated with pathological changes in the bones, kidneys, intestines, and leucocytes. Therefore, the elevated level of ALP activity may be due to damage in any of the aforementioned organs as a result of direct effect of the pesticides or as a result of oxidative damage to them. The finding is consistent with that reported by several workers following either DLT (Manna et al., 2004; Mazmanci et al., 2011) or CPF (Khan and Kour, 2007; Uzun and Kalender, 2013) intoxications. Pesticides via oxidative stress are known to cause multiple organ injury that may lead to the elaboration of the enzyme in the blood stream. The high LDH activity may not be attributed to damage to only one organ, but it has been specifically linked to hepatocellular necrosis, leading to its leakage into the blood stream (Mansour and Mossa, 2010). Elevated serum LDH activity has been reported by several workers as a result of DLT (Manna et al., 2004; Saoudi et al., 2011) and CPF (Mansour and Mossa, 2010; El-Mazoudy et al., 2011) poisoning in rats. The elevation in serum LDH was notable in the co-exposed group compared to when the two pesticides were administered singly.
Pretreatment with ALA in this study was able to decrease the elevated activities of AST, ALT, ALP and LDH in the co-exposed group. This may partly be due to the antioxidant role of ALA, which resulted in the scavenging of ROS and neutralizing the effects of the oxidants before they could cause damage to the liver or other organs associated with the enzymes. This finding was further confirmed by the decrease of hepatic LPO in the pretreated group in the present study. Hepatoprotective effect of ALA has been demonstrated by Dwivedi et al. (2014) and Al-Attar (2010) following exposure of rats to dichlorvos and lead, and malathion, respectively.
The present study showed that co-exposure to DLT and CPF resulted in inhibition of hepatic SOD activity. This decrease may be due to the direct damaging effect of free radicals generated by the pesticides on the enzyme. Although liver is known to play a central role in detoxification, it is chronically exposed to xenobiotics and their derivatives and, thus, prone to oxidative damage (Khan and Kour, 2007). The decrease in hepatic SOD activity in the co-exposed group agrees with that reported by Tuzmen et al. (2008) in rats fed with a combination of CPF and DLT. Decreased hepatic SOD activity has been reported by several authors following exposure of rats to CPF (Khan and Kour, 2007; Zama et al., 2007) and DLT (Manna et al., 2004; Mazmanci et al., 2011). The present study revealed an alteration in hepatic GPx activities following chronic co-exposure of rats to CPF and DLT. The decrease in hepatic GPx observed may be due to oxidative inactivation of the enzyme protein because of accumulation of the insecticide in the tissue (El-Tawil and Abdel-Rahman, 2001). Inhibition of GPx activity is often accompanied by a depletion of GSH or increased hydrogen peroxide generation, which may result in oxidative stress (Demir et al., 2011). Several authors have demonstrated decreased hepatic GPx activity following exposure of rats to either DLT (Mazmanci et al., 2011; Rehman et al., 2006) or CPF (Uzun and Kalender, 2013; Verma and Srivastava, 2003). The reduction in the antioxidant enzymes of the hepatic tissues observed in the co-exposed group was more when compared to those of single pesticide exposure.
The amelioration of the alterations in hepatic antioxidant enzyme activities (SOD and GPx) induced by CPF + DLT exposure following pretreatment with ALA may be due the antioxidant properties of ALA. ALA is able to scavenge deleterious toxic radicals capable of altering antioxidant enzymes and causing tissue injury in the liver and/or accelerate their removal, thus improving the redox status of the liver. Being a potent antioxidant, ALA neutralizes oxidants and protects cell membranes by reducing oxidized ascorbic acid and glutathione, which in turn recycle vitamin E (Packer et al., 1995).
The pesticides, CPF and DLT, are lipophilic substances that may enhance LPO by their direct interaction with cellular membranes (Tuzmen et al., 2008). The present study revealed that co-exposure to the two pesticides notably increased MDA levels in the hepatic tissue of the intoxicated rats. Impairment of enzymatic antioxidant system apparently favours accumulation of ROS that are responsible for increased LPO on pesticide exposure. The increase of MDA concentrations observed in the hepatic tissues of rats co-exposed to CPF and DLT is consistent with that reported by Tuzmen et al. (2008) in rats fed with the combination of CPF and DLT. Several workers have demonstrated increased MDA concentration following DLT (Mazmanci et al., 2011; Rehman et al., 2006) and CPF (Alvarez et al., 2008; Ma et al., 2013) poisoning. The increase in LPO as evidenced by the high MDA concentrations in the liver following CPF and DLT co-exposure may have caused considerable ultrastructural damage of liver cells and eventual loss of their membrane integrity (Khan and Kour, 2007; Manna et al., 2004). The increased MDA concentration in the liver may also be due to the hepatic damage evidenced by increased AST, ALT, ALP, and LDH activities and the alterations in hepatic antioxidant enzyme system observed in this study. The increased MDA concentrations in the hepatic tissue observed were notably higher in the co-exposed group than those exposed to either CPF or DLT.
Pretreatment with ALA, however, reduced MDA in the hepatic tissue in the co-exposed group, through scavenging of free radicals and increasing the antioxidant activities in the liver of the rats to detoxify free radicals. This finding gives credence to previous reports, suggesting that ALA is a potent metabolic antioxidant source to quench free radicals (Packer et al., 1995, 1997).
Inhibition of AChE activity on OP exposure is an indicator of OP poisoning. OP compounds generally elicit their effects by inhibition of AChE. Recently, evidence has emerged that the reduction of AChE activity is not due exclusively to OP and carbamates, but that other environmental contaminants, such as pyrethroid insecticides, can induce AChE inhibition (Badiou and Belzunces, 2008). The present study revealed that CPF + DLT caused an inhibition of AChE in the serum of exposed rats. The degree of inhibition was notably higher in rats exposed to CPF than those exposed to either DLT or co-exposed to CPF and DLT. Similar inhibition of AChE activity in plasma was also reported by El-Halwagy and Zaki (2009) in rats exposed to the combination of an OP, diazinon and DLT, and they attributed it to the antagonistic effect of the OP and pyrethroid. The decrease in AChE activity observed in the serum of DLT-exposed rats agrees with the results of Kale et al. (2009), Yousef et al. (2006) and El-Demerdash (2007) who demonstrated that pyrethroids cause a decrease of AChE activity in the erythrocytes, plasma and brain of exposed rats. The decrease in AChE activity following CPF + DLT and DLT exposure may also be due to increased LPO, as LPO induced by pesticides other than OPs have been reported to indirectly affect membrane-bound enzymes such as AChE leading to a decrease in its activity (López et al., 2007).
The increased AChE activity, observed in the group pretreated with ALA, shows the AChE restoratory effect of ALA. ALA’s antioxidant ability to reduce LPO from oxidative stress may have protected the AChE from oxidative modifications since AChE is a membrane-bound enzyme.
The study further revealed that co-exposure of rats to CPF and DLT resulted in an inflammatory reaction with increased production of TNF-α in the hepatic tissue. The increased production of TNF-α (pro-inflammatory cytokine) may indicate that inflammation is involved in the pathogenesis of CPF and DLT-induced toxicity. TNF-α is a key cytokine involved in inflammation, immunity and cellular organization (Locksley et al., 2001). OP-induced tissue toxicity is due to the induction of inflammatory and production of inflammatory cytokines (Alluwaimi and Hussein, 2007). TNF-α–induced cytotoxicity also occurs through varying mechanisms, including the overproduction of ROS, as a result of pesticide exposure as observed by the increased MDA concentration and alteration in antioxidant enzymes in the present study, which in turn damages the cellular components such as protein, lipids and DNA (El-Gerbed, 2014). The elevation of TNF-α level was notably higher in the co-exposed group, indicating there was more inflammation and cell damage to the tissues of the CPF and DLT co-exposed rats than those occurring in the CPF- and DLT-exposed rats. El-Gerbed (2014) also demonstrated that DLT induced an elevation of TNF-α level in the kidneys of exposed rats.
Pretreatment with ALA caused a reduction in the hepatic TNF-α level of the ALA + CPF + DLT group. This reduction may be due to the ability of ALA to modulate the inflammatory and pro-inflammatory responses of pesticide-induced toxicity. Its ability to decrease LPO and restore altered antioxidant enzymes as observed in this study may have also contributed to its anti-inflammatory effect, as inflammation has been associated with increased LPO and altered antioxidant enzymes (Jesudason et al., 2008).
In conclusion, the study revealed that chronic exposure of rats to low level of the combination of CPF and DLT resulted in marked alterations in most of the biochemical parameters evaluated. These alterations may be partly due to oxidative stress and inflammation. The alterations were, however, mitigated by the supplementation with ALA. The study, therefore, showed that supplementation with ALA has beneficial effect on damages evoked by long-term co-exposure to CPF and deltamethrin, and perhaps other pesticides.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.