
Abstract
Homosalate (HMS) and 2-ethylhexyl 4-dimethylaminobenzoate (OD-PABA) are ultraviolet filters. We aimed to investigate the effects of dermal exposure to HMS and OD-PABA during the prenatal, lactation, and early infancy periods on pubertal development and thyroid function in male and female rats. The thyroid glands, uteri, testes, prostate glands, and seminal vesicles were excised and weighed, the reproductive organs were analyzed histologically, and the serum hormone levels were measured. In the prenatal period, the thyroxine (T4) levels increased in the female rats in the exposed groups (p < 0.05); the thyroid weights, reproductive organ weights, and gonadal hormone levels were not altered. In males, the testosterone levels decreased (p < 0.05), but the thyroid weights, T4 levels, prostate, and testis weights were not changed. In the lactation period, the weights of the thyroid glands increased in the exposed female groups (p < 0.05), but the T4, gonadal hormone levels, and reproductive organ weights were not changed. In the males, the thyroid gland weights, T4 levels, reproductive organ weights, and gonadal hormone levels were not changed. During infancy, the thyroid gland weights increased in the female rats in the exposed groups (p < 0.05), but the T4 levels, gonadal hormone levels, and reproductive organ weights were not affected. In the male rats in the exposed groups, the T4 levels were increased (p < 0.05), but the thyroid and reproductive organ weights, gonadal hormone levels were not affected. Organ histopathology was not affected in all groups. HMS and OD-PABA do not have endocrine disruptor effects on thyroid function and the pubertal development of female and male rats.
Introduction
Ultraviolet (UV) filters are chemical substances that are able to absorb UV radiation and thus are commonly used in sunscreen products to protect the skin against sunburn damage and skin cancer. These chemicals are generally found in cosmetics, creams, lipsticks, skin lotions, hair dyes and sprays, shampoos, and fragrances and are used to increase the light stability and durability of these products as well as to protect the skin. Human beings are exposed to UV filters through 2 primary routes, that is, directly through the use of cosmetics (dermal exposure) and indirectly through the food chain (Díaz-Cruz and Barceló, 2008; Schlumpf et al., 2001, 2008). UV filters have been detected in water sources, wastewaters, soil, sludge, and fish (Díaz-Cruz and Barceló, 2008; Schlumpf et al., 2001). The widespread use of UV filters has raised concerns regarding their adverse effects and toxicological profiles. Several experimental animal and in vitro studies have highlighted that some UV filters may exhibit adverse effects as endocrine disrupters (EDs) (Díaz-Cruz and Barceló, 2008). The UV filters used in cosmetics, such as benzophenone-3 (BP-3), 3-benzylidene-camphor (3-BC), 3-(4-methyl-benzylidene) camphor (4-MBC), 2-ethylhexyl 4-methoxy cinnamate (OMC), homosalate (HMS), 2-ethylhexyl 4-dimethylaminobenzoate (OD-PABA), and 4-aminobenzoic acid (PABA), have been suggested to act as EDs (Krause et al., 2012). Many of these substances, including those with both synthetic and natural origins, may exhibit estrogenic and/or antiandrogenic activities that may lead to serious alterations in the ecosystem and particularly serious alterations in the normal development of the human endocrine axis (Carou et al., 2009). EDs have been linked to effects on the functions of the reproductive and thyroid systems (Maerkel et al., 2007; Schmutzler et al., 2004). EDs act as agonists or antagonists of hormone receptors and may influence the expression levels of the steroid hormone receptor messenger RNAs and genes that are regulated by these hormones (Durrer et al., 2005; Klann et al., 2005; Maerkel et al., 2007; Schmutzler et al., 2004). Several effects of exposure to BP-3, 3-BC, 4-MBC and OMC on the reproductive system have been observed in extended, one-generation developmental studies. These effects include alterations in the weights and histology of the reproductive organs of both sexes (Axelstad et al., 2011; Durrer et al., 2007; Hofkamp et al., 2008; Schlumpf et al., 2004a). The age during exposure and the duration of exposure to endocrine disruptors are important factors. Previous in vitro and in vivo studies have shown that 4-MBC, BP-3, and OMC may interfere with the hypothalamic–pituitary–thyroid axis (Axelstad et al., 2011; Maerkel et al., 2007; Schmutzler et al., 2004, 2007). Several studies have noted the direct effects of oral treatment with UV filters on thyroid hormone synthesis in rat models (Klammer et al., 2005; Schlumpf et al., 2001; Seidlová-Wuttke et al., 2006). HMS and OD-PABA are the most common active ingredients used as UV filters (Nash, 2006; Sarveiya et al., 2004). Studies of the endocrine disrupting effects of HMS and OD-PABA on the pubertal development and thyroid functions of rat pups are limited (Schlumpf et al., 2001). Moreover, the effects of HMS and OD-PABA during the prenatal and lactation periods are not known. In the present study, the effects of dermal HMS and OD-PABA exposure on the thyroid gland and its functions, the reproductive system, and pubertal development were evaluated in a rat model at different time points during life that mimicked the prenatal, lactation, and early infancy periods using the Organization for Economic Cooperation and Development (OECD) enhanced test guideline 407 (Gelbke et al., 2007).
Methods
Chemicals
Both OD-PABA (CAS no 21245-02-3; 98% purity) and HMS (CAS no 118-56-9) were purchased from the Sigma-Aldrich Chemical Company (Munich, Germany). Liquid paraffin was used as a vehicle control.
Experimental animals
Wistar Hannover rats were purchased from the Bagcilar Training and Research Hospital of the Experimental Research and Skills Development Center, Istanbul. The animals were housed in polypropylene cages (43 cm × 30 cm × 15 cm) with laboratory-grade pine shavings as bedding. The rats were bred on a controlled light/dark cycle (lights on 02:00–16:00 h) at a controlled temperature (22°C ± 1°C) and were provided free access to food and water ad libitum. The Bagcilar Training and Research Hospital Animal Experiments Ethics Committee approved the study protocol (no. 2014-11). All animals were treated humanely and with particular care to alleviate suffering.
Experimental design
HMS and OD-PABA were applied as liquid solutions in paraffin. The OECD 427 test methods were used as the basis of these experiments. Under normal conditions, the exposure of humans to these chemicals is relatively limited; thus, doses ranging from 1 to 5 mg/cm2 were used in this study to mimic human exposure levels, as previously described. The recommended application doses of 0–10 mg/cm2 have been used in in vitro assessments of the protection afforded by sunscreen applied at 2 mg/cm2 in vivo (Janjua et al., 2004; Stokes and Diffey, 1999). In the present study, a 10% HMS paraffin solution and an 8% OD-PABA paraffin solution were topically applied to shaved areas on the rats’ back at doses of 2 mg/cm2. The back of each rat was shaved once per week to expose an area of 9 cm2 during the pregnancy and lactation periods or an area of 4 cm2 during infancy. The applications were performed once per day.
Prenatal period
To determine the effects of HMS and OD-PABA during the prenatal period, 15 adult female Wistar Hannover rats were randomly divided into 3 groups. Vaginal smears were taken to determine estrus. After estrus, the females were separated into 3 cages (5 per cage), and 1 male rat was added to each of the 3 cages. Mating vaginal smears were collected, and the identification of sperm on the smears was accepted as evidence of pregnancy. Gestational day 1 (GD 1) was defined as 1 day post-mating. Beginning on GD1, HMS was applied to 1 group, OD-PABA was applied to 1 group, and the third group served as the control and received applications of only paraffin. The daily applications continued until delivery. The animals were maintained in separate cages following the HMS, OD-PABA, and paraffin applications. Animals were maintained in separate cages after the first dose application until delivery. The daily application was made in separate cages. After delivery, no treatments were applied to the mothers or pups. The expected day of delivery, that is, GD 22, was designed as postnatal day (PND) 0 for the pups. The pups were weaned at PND 20 and identified by gender on PND 21. The animals were maintained in controlled conditions with free access to water and chow. On PND 21, the pups were separated by gender, and the pups of all mothers that had received applications were included in the study. Only 2 of the mothers that received OD-PABA, 3 of the mothers that received paraffin, and 4 of the mothers that received HMS applications gave birth, and all of the pups born to these mothers were included in the study. Beginning on PND 26, the pups were examined daily for signs of puberty.
Lactation period
To determine the effects of HMS and OD-PABA exposure during the lactation period, 15 adult female Wistar Hannover rats were randomly divided into 3 groups on GD1. All rats were monitored throughout the study and maintained in standard laboratory conditions with unlimited access to food and water. Each mother and her pups were maintained in a separate cage. From PND 2 to 21, HMS, OD-PABA, and paraffin were applied each day to the mothers in the appropriate groups. After application, each mother was separated for 1 h from their pups. Then the animals were continuously observed; they did not lick the skin of their mothers. On PND 21, the pups were identified by gender. Four male and four female offspring from each litter were included in the study. Beginning on PND 26, the pups were examined daily for signs of puberty.
Infancy period
On PND 21, naive pups whose mothers had not received HMS or OD-PABA during pregnancy or lactation were separated by gender. The male and female pups were separated and then randomly divided into 3 groups of 10. All of the selected pups were from different litters. HMS, OD-PABA, and paraffin were applied to the appropriate groups over 6 consecutive days from PND 21 to 26. The UV filters were applied once a day onto the back of pups with a soft brush once a day. After the application, pups were retained in separate cages for 30 min. After 30 min, the skin appeared dry, the pups were transferred to its home cage. Beginning on PND 26, the pups were examined daily for signs of puberty.
External physical signs of puberty onset
Vaginal patency is estrogen dependent and generally indicative of the occurrence of the first ovulation and the onset of estrous cyclicity in the rat. Throughout the dosing period, the animals were examined at the same time daily for vaginal opening. The age at complete vaginal opening was recorded. Beginning on the day of vaginal opening, daily vaginal smears were collected and observed under a low-power light microscope for the presence of leukocytes, nucleated epithelial cells, and cornified epithelial cells to determine the age at the first complete estrous cycle after vaginal opening. The vaginal smears were classified as diestrus (based on the presence of leukocytes), proestrus (based on the presence of nucleated epithelial cells), or estrus (based on the presence of cornified epithelial cells) (Stoker et al., 2004). Daily vaginal cytology was recorded for each female until the day of sacrifice.
The separation of the foreskin of the penis from the glans penis, that is, preputial separation (PPS), is an early reliable marker of the progression of puberty in male rats (Korenbrot et al., 1977). In the present study, PPS was monitored beginning on PND 26 and continued until all males exhibited separation. All males were examined once daily at approximately the same time each day.
Euthanasia, body weights, and organ weights
After the first estrus in the female pups and PPS in the male pups, the rats were killed by pentobarbital overdose. Subsequently, trunk blood was intracardially collected between 9:00 and 11:30 a.m. The thyroid gland, both testes, the prostate, and the seminal vesicles were excised from the male offspring and weighed. The thyroid gland, uterus, and bilateral oviducts and ovaries were excised from the female offspring and weighed.
Serum hormone levels
Serum was obtained by centrifugation (2,400 rpm, 20 min, 4°C) in a refrigerated device and then frozen at −70°C until the analyses of the serum thyroid-stimulating hormone (TSH), thyroxine (T4), follicle-stimulating hormone (FSH), luteinizing hormone (LH), testosterone, and estradiol levels. The serum TSH, T4, FSH, LH, testosterone, and estradiol levels were measured with rat-specific enzyme-linked immunosorbent assays (Eastbiopharm Co., Hangzhou, China) using a 1420 VICTOR3 absorbance reader (PerkinElmer, Inc., Waltham, MA, USA).
Histopathologic evaluation
Immediately following dissection at necropsy, the uterus, both ovaries, and the left testis were placed in 10% neutral-buffered formalin for 24 h. The tissues were then rinsed and stored in 70% alcohol until they were embedded in paraffin, sectioned, and stained with hematoxylin and eosin. Ten males and 10 females from each group were used for the pathological evaluation. The prenatal OD-PABA and the entire male control group were also used for the pathological evaluation.
Female rats
According to the guidelines, ovarian histology should include evaluations of the “presence/absence of tertiary/antral follicles, presence/absence of corpora lutea (CL), changes in corpus luteum development, and changes in the numbers of both primary and atretic follicles, in addition to any abnormalities/lesions, such as ovarian atrophy.” The endometrial morphology during estrous is characterized by the progressive degeneration and necrosis of both the glandular and luminal endometrial epithelia accompanied by leukocyte infiltration (Picut et al., 2015). The numbers and structures of corpus lutea in the ovaries, the numbers of Graafian follicles, the numbers of necrotic ova, and the appropriateness of the estrous cycles were compared between the control and experimental groups. In this article, of the histopathological results obtained from the female rats, only pictures of the histopathological results of the ovaries have been presented as a representative tissue in the figures.
In the uterus, the presence of mitotic figures and the glandular/ablated cell height at 1.5–3× the nuclear height ratios were evaluated and compared with the control group.
Male rats
Decreased numbers of elongated spermatids and decreased numbers of sperm in the testis are indicative of endocrine-mediated delays in puberty (Creasy, 2003). The left testis was evaluated. The seminiferous tubules were classified as follows: normal (concentric and normally organized germ cell layers present in the seminiferous epithelium) or abnormal (based on the presence of germ cells and cellular debris in the lumen, multinucleated formations, seminiferous epithelium with acidophilic cells, few germ cell layers, or vacuole formation or degeneration in the seminiferous epithelium). To evaluate the degree of maturation of the seminiferous epithelium, 100 random cross sections of the seminiferous tubules per animal were evaluated using an adaptation method of assigning values according to the most numerous mature germ cells in the tube. The maturation findings were compared between the control and experimental groups.
Statistical analysis
The statistical evaluations of the data were performed with SPSS version 22.0 (IBM Corp., Armonk, NY, USA). The descriptive statistics are reported as the means ± the standard deviations for the constant variables and as the frequencies or percentages for the categorical variables. The normality of the data distributions was assessed with the Kolmogorov-Smirnov test. The homogeneity of the variance between groups was assessed with Levene’s test. Analyses of variance (ANOVAs) were used to compare the differences between more than 2 groups when the variable was normally distributed and the variance was homogenous. Bonferroni tests were applied when the comparisons between groups were statistically significant. Welch’s tests were performed to compare the variables that were normally distributed but did not have equal variances. Dunnett’s T3 test was used to evaluate the statistically significant differences between these groups. The Kruskal-Wallis test was used to compare groups when the variables were not normally distributed, and the Mann–Whitney U test was used for multiple comparisons between groups that were shown to be significantly different. The results were assessed via the application of the Bonferroni correction (p < 0.05 was accepted as statistically significant). For the evaluation of the histopathological data, p values were obtained using the chi-square test.
Results
Female treatments
Prenatal period
Effect on the thyroid
The average thyroid gland weights in the HMS (p = 0.0001) and OD-PABA (p = 0.0001) groups were significantly reduced compared with the control group. The TSH levels in the control group were significantly increased compared with the OD-PABA group (p = 0.004). The control group exhibited significantly reduced T4 levels compared with both the HMS (p = 0.024) and OD-PABA groups (p = 0.0001; Table 1, Figure 1).
Effects of HMS and OD-PABA exposure during the prenatal, lactation, and infancy periods on rat thyroid functions and puberty in female offspring.
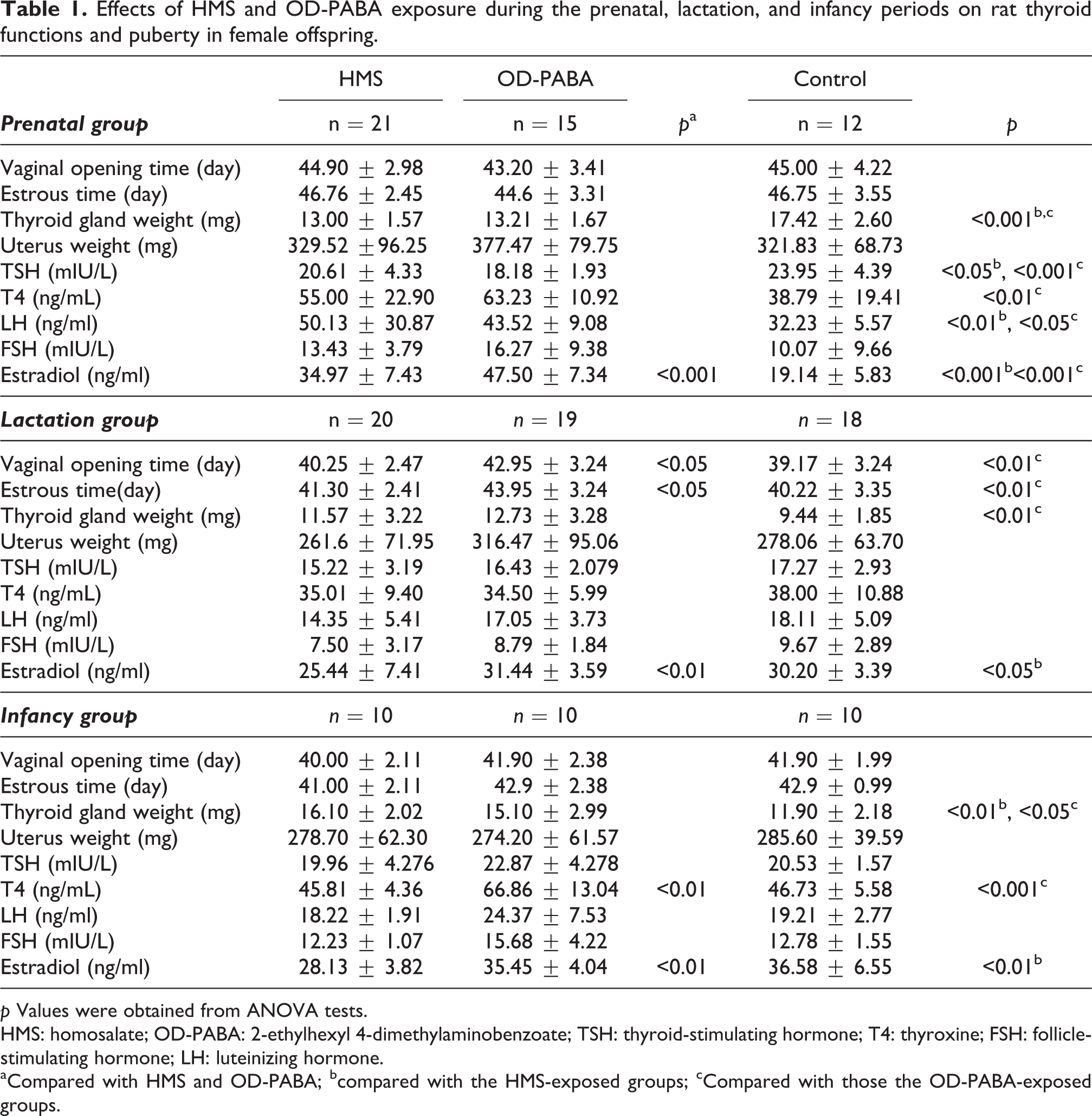
p Values were obtained from ANOVA tests.
HMS: homosalate; OD-PABA: 2-ethylhexyl 4-dimethylaminobenzoate; TSH: thyroid-stimulating hormone; T4: thyroxine; FSH: follicle-stimulating hormone; LH: luteinizing hormone.
aCompared with HMS and OD-PABA; bcompared with the HMS-exposed groups; cCompared with those the OD-PABA-exposed groups.
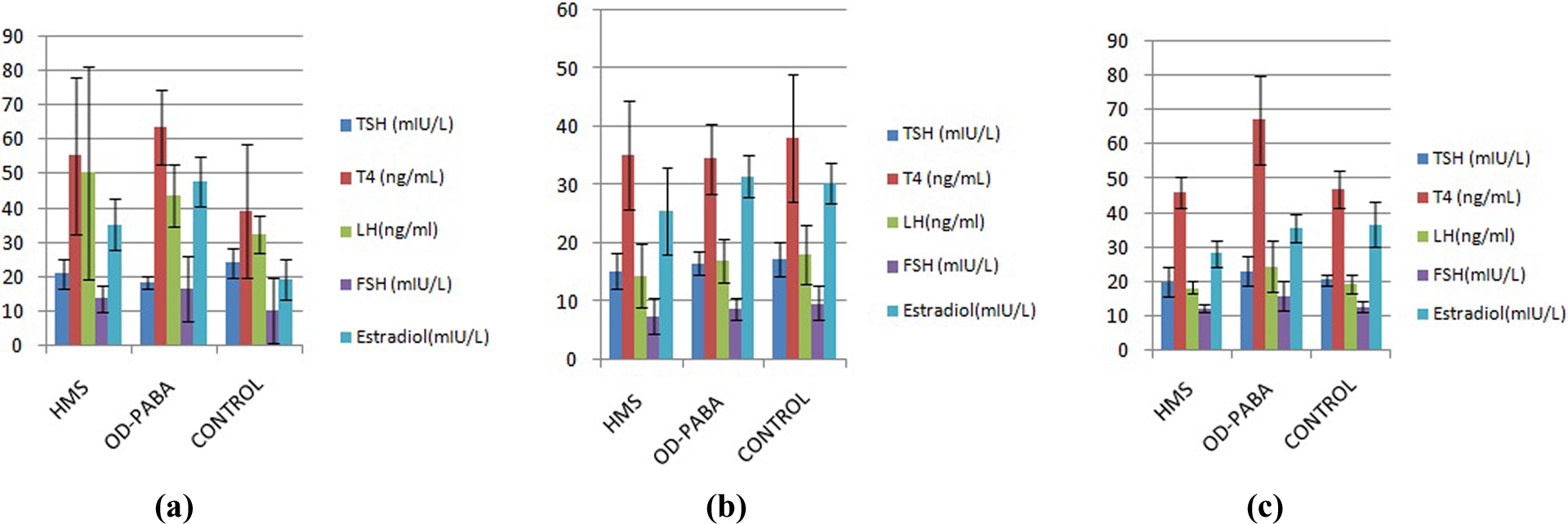
Comparisons of the hormone levels of the female offspring exposed (a) during the prenatal period, (b) during lactation, and (c) during infancy.
Effect on puberty
The LH levels of the control group were significantly reduced compared with the HMS (p = 0.049) and OD-PABA groups (p = 0.002). The estradiol levels of the OD-PABA group were significantly increased compared with the HMS group and control group (p = 0.0001; Table 1, Figure 1).
Histopathology
Although the Graafian follicle variable was significantly different across groups, no significant differences were observed in the comparisons between groups (Table 2). The structures of the Graafian follicles and corpora lutea were compatible with the stage of the estrus cycle (Figure 2). No significant differences between the control and experimental groups were noted in the histopathological examinations of the uteri (Table 3).
Effects of HMS and OD-PABA exposure on the reproductive organ histopathology of the female offspring during infancy.
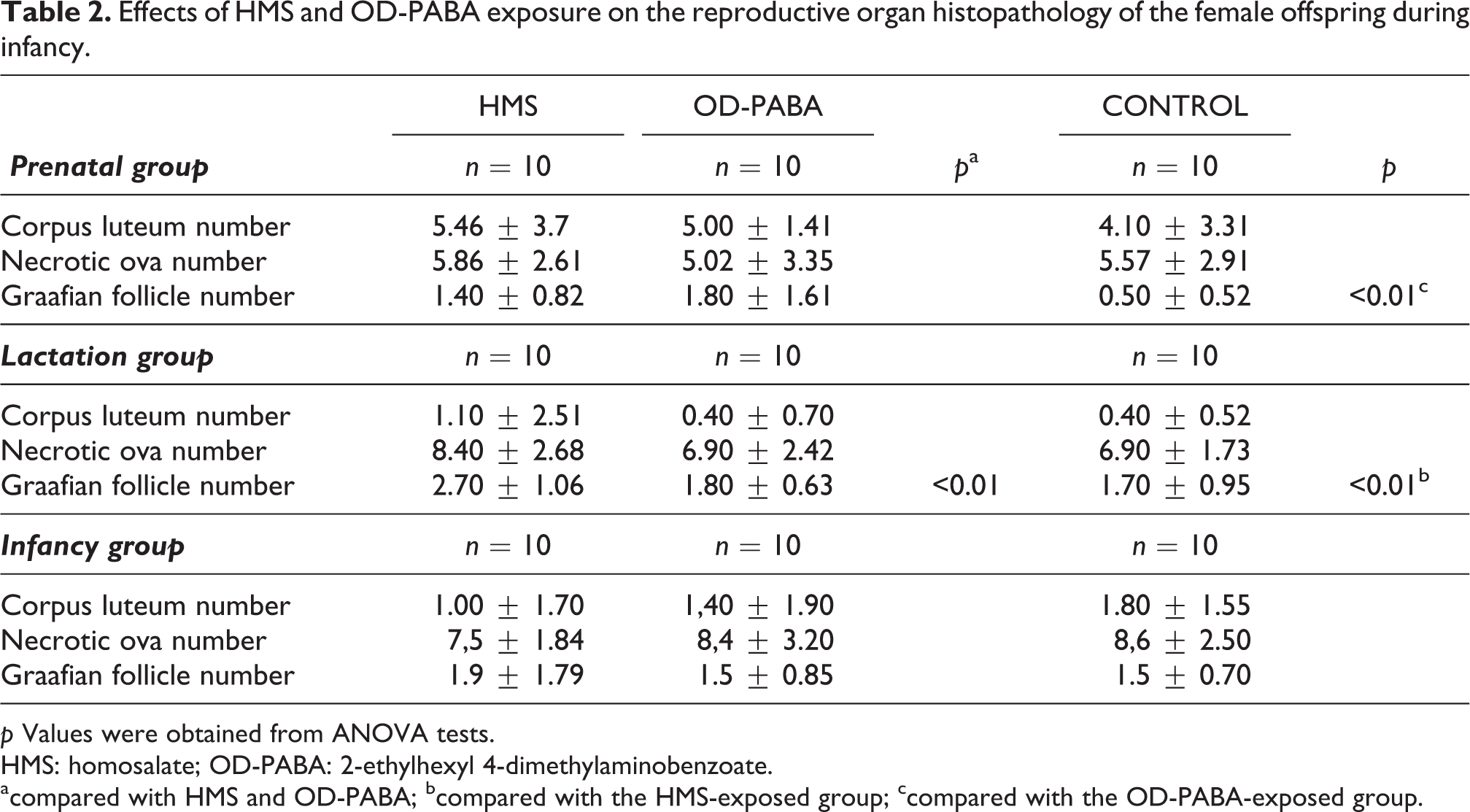
p Values were obtained from ANOVA tests.
HMS: homosalate; OD-PABA: 2-ethylhexyl 4-dimethylaminobenzoate.
acompared with HMS and OD-PABA; bcompared with the HMS-exposed group; ccompared with the OD-PABA-exposed group.
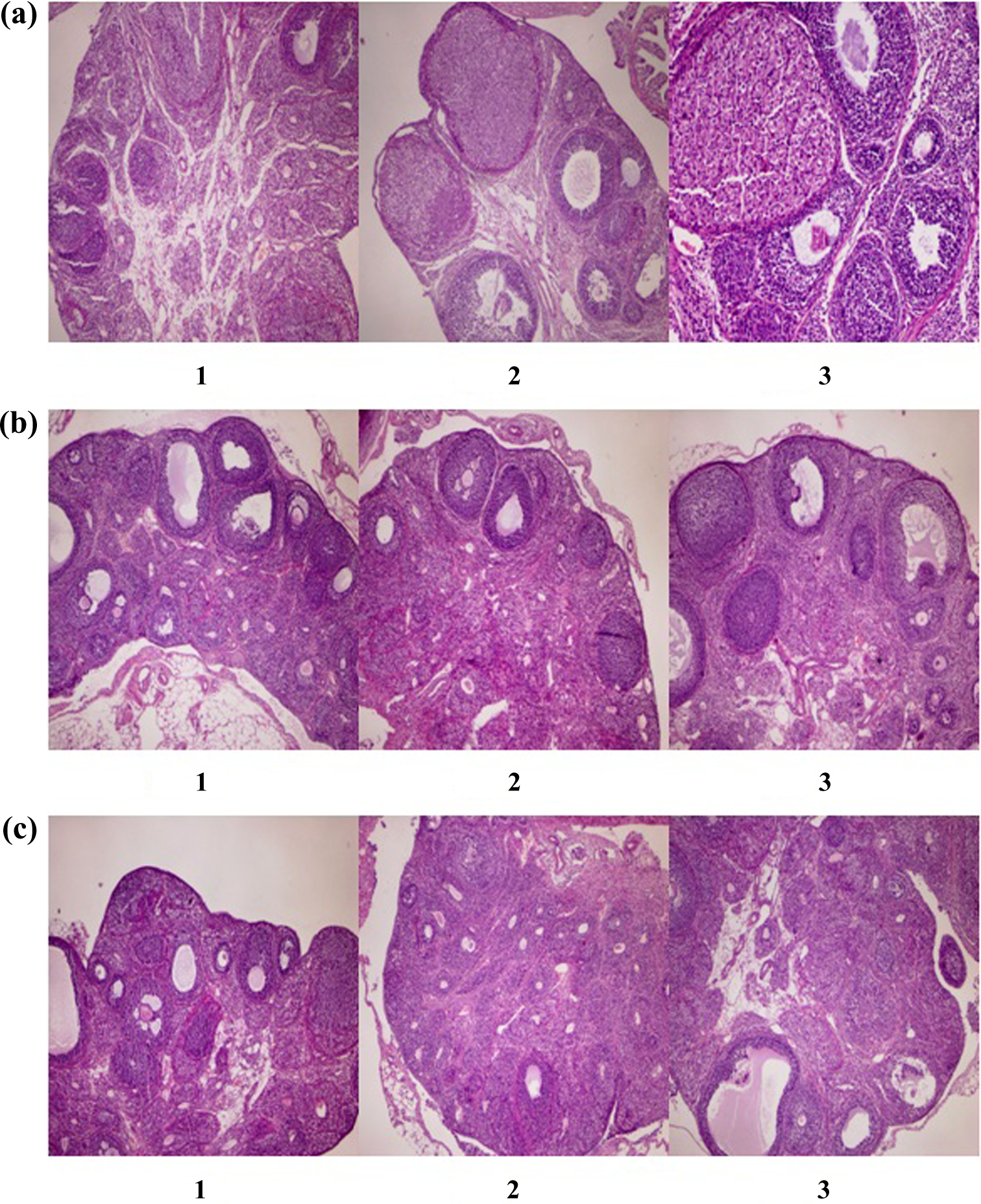
Comparisons of the ovary histopathological findings of female offspring exposed (a) during prenatal period, 1 = HMS exposure group, 2 = OD-PABA exposure group, 3 = control group (b) during lactation, 1 = HMS exposure group, 2 = OD-PABA exposure group, 3 = control group (c) during infancy, 1 = HMS exposure group, 2 = OD-PABA exposure group, 3 = control group. HMS = homosalate; OD-PABA = 2-ethylhexyl 4-dimethylaminobenzoate.
Histopathological findings of the uteri of 3 groups of rats in uterine specimen examination.
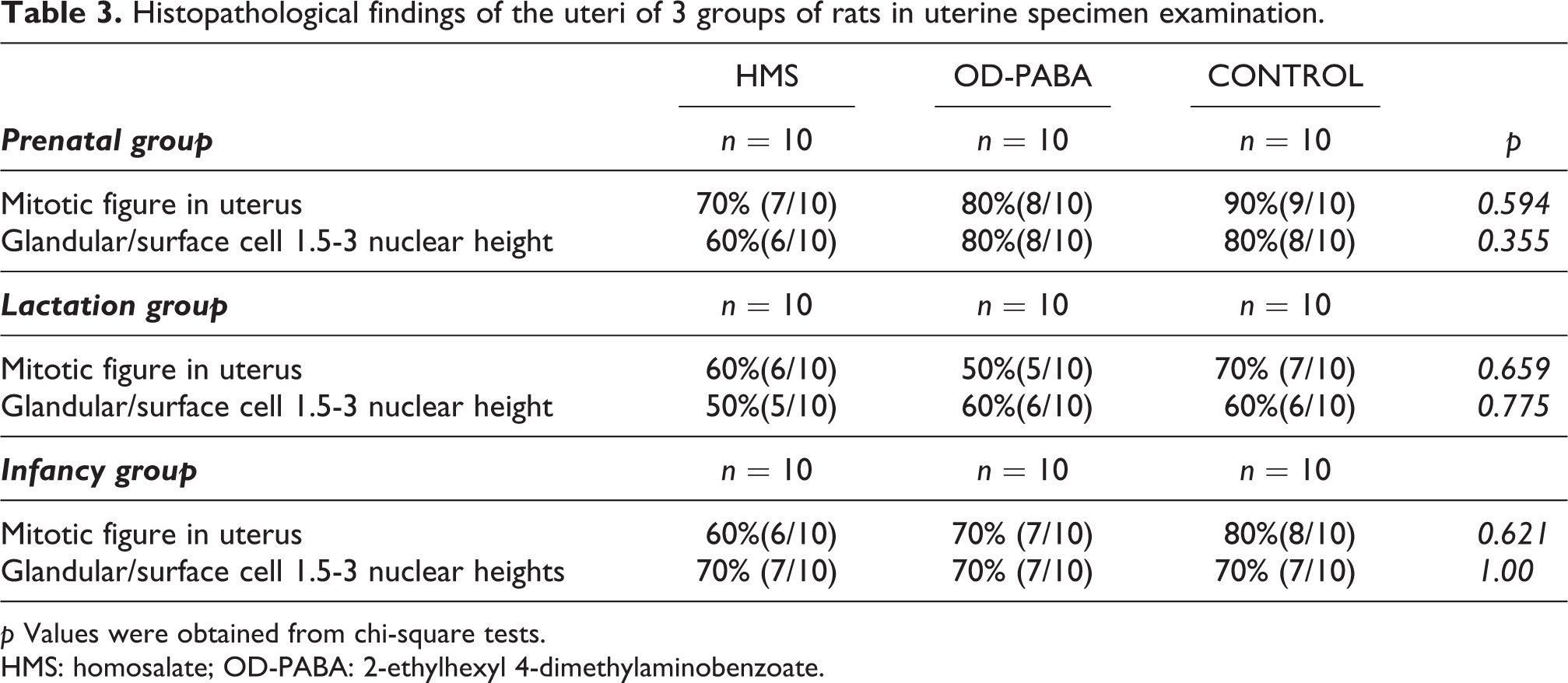
p Values were obtained from chi-square tests.
HMS: homosalate; OD-PABA: 2-ethylhexyl 4-dimethylaminobenzoate.
Lactation period
Effect on the thyroid
The thyroid gland weights in the OD-PABA group increased significantly compared with the control group (p = 0.006). No significant differences in the TSH and T4 levels were observed between the groups (Table 1, Figure 1).
Effect on puberty
The vaginal opening time of the OD-PABA group was significantly later than that of both the HMS (p = 0.020) and control groups (p = 0.001). The first estrous time occurred significantly earlier in the control group than in the OD-PABA group (p = 0.001) and significantly earlier in the HMS group than in the OD-PABA group (p = 0.025). The estradiol levels in the OD-PABA group increased significantly compared with the HMS group (p = 0.009), whereas the estradiol levels in the control group increased significantly compared with the HMS group (p = 0.045; Table 1, Figure 1).
Histopathology
The number of Graafian follicles in the HMS group increased significantly compared with the OD-PABA and control groups (p = 0.020; Table 2). The structures of the Graafian follicles and corpus lutea were compatible with the stage of the estrus cycle (Figure 2). No significant differences were noted between the control and experimental groups in the histopathological examinations of the uteri (Table 3).
Infancy period
Effect on the thyroid
The OD-PABA group exhibited greater thyroid weights than the control group (p = 0.021). The T4 levels in the OD-PABA group were significantly increased compared with both the HMS group (p = 0.0001) and control group (p = 0.0001; Table 1, Figure 1).
Effect on puberty
The estradiol levels of the HMS group were significantly decreased compared with the control (p = 0.005) and OD-PABA groups (p = 0.007; Table 1, Figure 1).
Histopathology
No significant differences were observed in the Graafian follicles or corpus lutea (Table 2). The structures of the Graafian follicles and corpus lutea were compatible with the stage in the estrus cycle (Figure 2). No significant differences were noted between the control and experimental groups in the histopathological examinations of the uteri (Table 3).
Male treatments
Prenatal period
Effect on the thyroid
The TSH levels in the control group were significantly increased compared with the HMS group (p = 0.033), and the T4 levels in the OD-PABA group were significantly increased compared with both the HMS group (p = 0.0001) and control group (p = 0.0001; Table 4, Figure 3).
Effects of HMS and OD-PABA exposure during the prenatal, lactation, and infancy periods on the thyroid hormone systems and puberty in the male offspring.
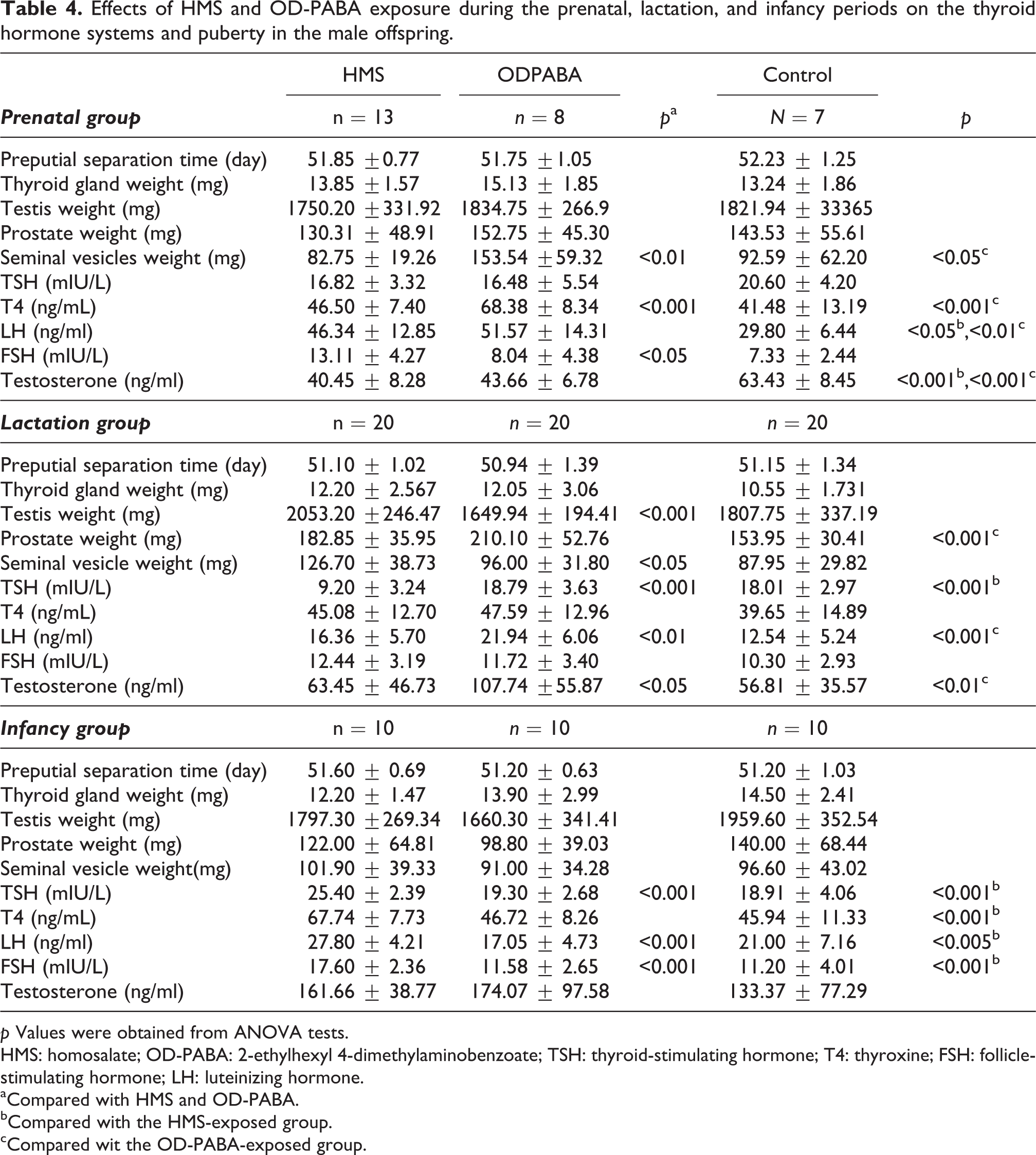
p Values were obtained from ANOVA tests.
HMS: homosalate; OD-PABA: 2-ethylhexyl 4-dimethylaminobenzoate; TSH: thyroid-stimulating hormone; T4: thyroxine; FSH: follicle-stimulating hormone; LH: luteinizing hormone.
aCompared with HMS and OD-PABA.
bCompared with the HMS-exposed group.
cCompared wit the OD-PABA-exposed group.
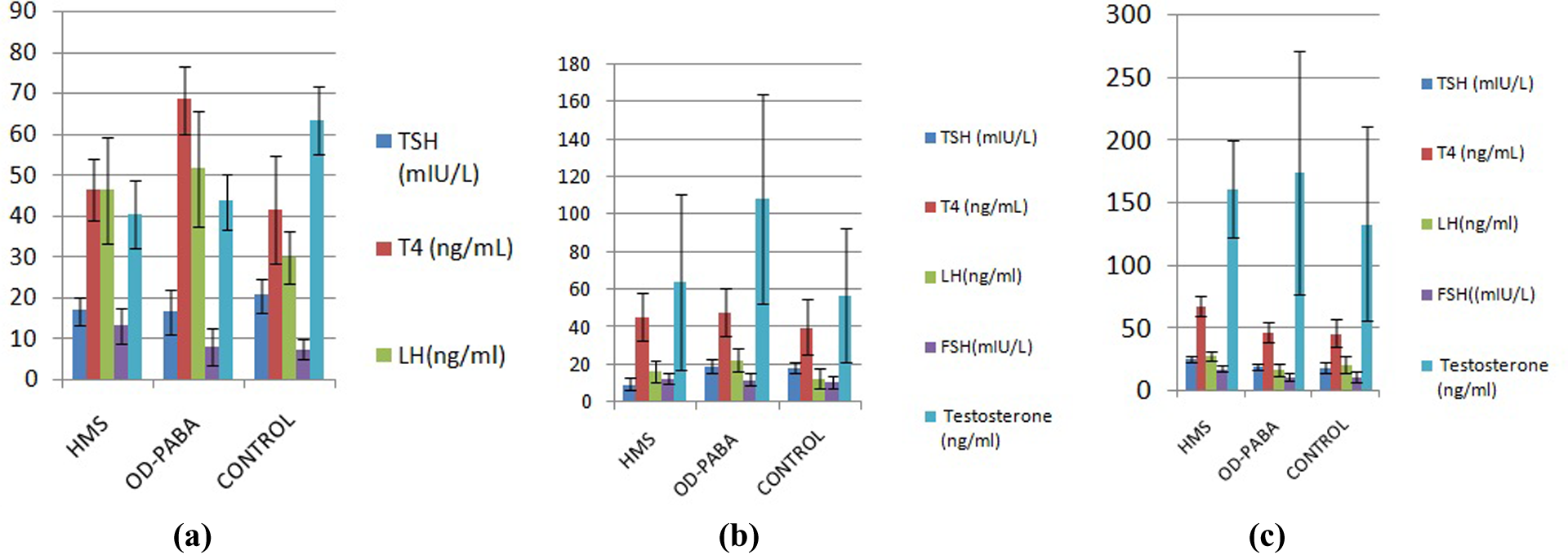
Comparisons of the hormone levels of the male offspring exposed during the (a) prenatal period, (b) during lactation, and (c) during infancy.
Effect on puberty
The weights of the seminal vesicles in the OD-PABA group were significantly increased compared with the HMS (p = 0.049) and control groups (p = 0.025). The LH values in the control group were significantly reduced compared with the OD-PABA (p = 0.010) and HMS groups (p = 0.02). The FSH levels in the HMS group were significantly increased compared with the control (p = 0.001) and OD-PABA groups (p = 0.010). The testosterone levels in the control group were significantly increased compared with the HMS (p = 0.0001) and OD-PABA groups (p = 0.002; Table 4, Figure 3).
Lactation period
Effect on the thyroid
The mean TSH levels in the HMS group were significantly reduced compared with the OD-PABA and control groups (p = 0.0001; Table 4, Figure 3).
Effect on puberty
The testis weights in the HMS group were significantly increased compared with the control (p = 0.016) and OD-PABA groups (p = 0.0001). The prostate weights in the control group were significantly reduced compared with the HMS (p = 0.027) and OD-PABA group (p = 0.001). The weights of the seminal vesicles in the HMS group were significantly increased compared with the OD-PABA (p = 0.019) and control groups (p = 0.002). The LH levels in the OD-PABA group were significantly increased compared with the HMS (p = 0.011) and control groups (p = 0.0001). The mean testosterone level in the OD-PABA group was significantly increased compared with the HMS group (p = 0.003) and control groups (p = 0.001; Table 4, Figure 3).
Infancy period
Effect on the thyroid
The HMS group exhibited significantly higher TSH levels than both the OD-PABA (p = 0.001) and control (p = 0.0001) groups. Additionally, the HMS group exhibited higher T4 levels than both the OD-PABA (p = 0.0001) and control (p = 0.0001) groups (Table 4, Figure 3).
Effect on puberty
The LH levels in the HMS group increased significantly compared with the OD-PABA (p = 0.001) and control (p = 0.031) groups. The FSH levels of the HMS group increased significantly compared with the OD-PABA (p = 0.001) and control (p = 0.0001) groups (Table 4, Figure 3).
Histopathology
The number of elongated spermatids did not decrease in the testes of any of the groups. There were no significant differences between the groups in the structures of the seminiferous tubules (Table 5, Figure 4).
Histopathological findings of the testes of the male offspring.
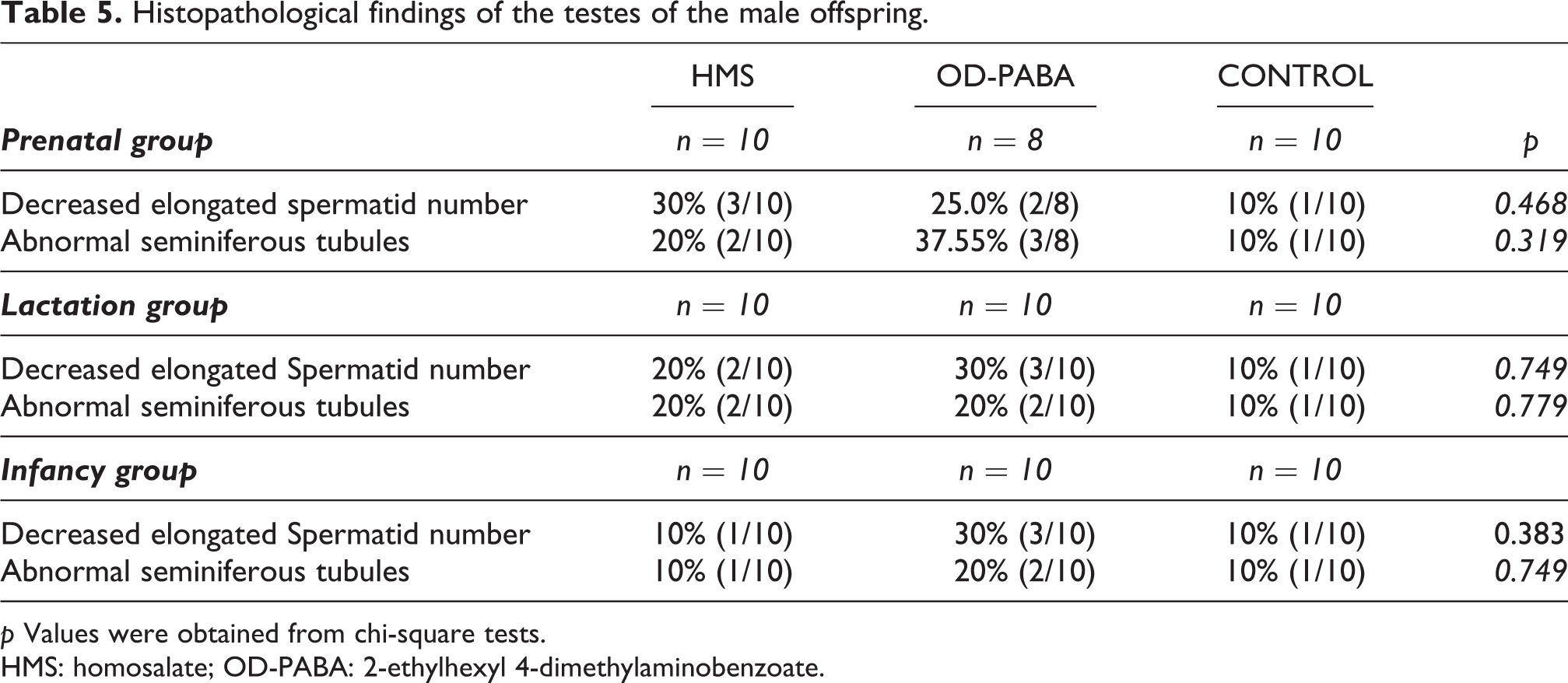
p Values were obtained from chi-square tests.
HMS: homosalate; OD-PABA: 2-ethylhexyl 4-dimethylaminobenzoate.
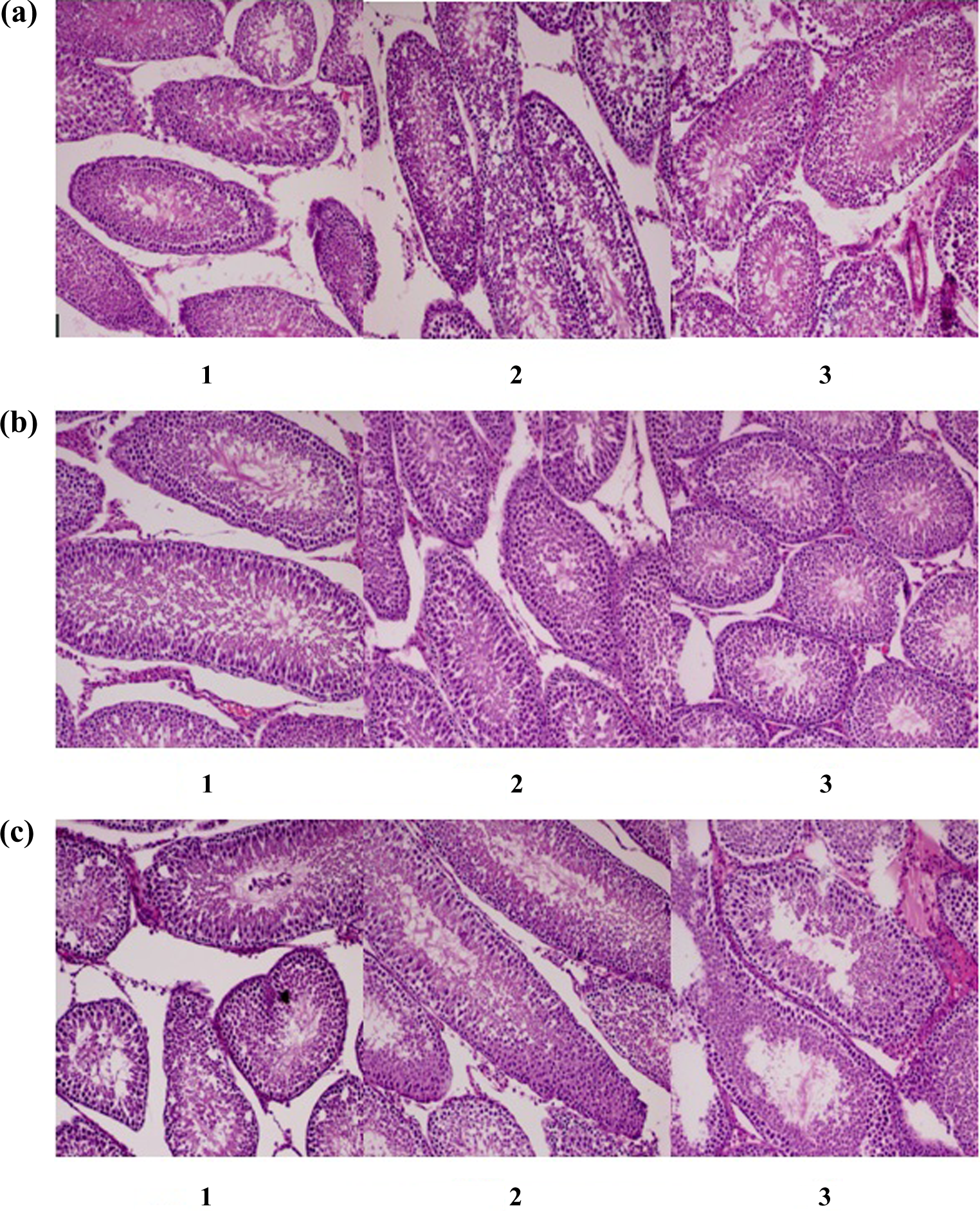
Comparisons of the testis histopathological findings of male offspring exposed (a) during prenatal period, 1 = HMS exposure group, 2 = OD-PABA exposure group, 3 = control group, (b) during lactation, 1 = HMS exposure group, 2 = OD-PABA exposure group, 3 = control group, (c) during infancy, 1 = HMS exposure group, 2 = OD-PABA exposure group, 3 = control group. HMS = homosalate; OD-PABA = 2-ethylhexyl 4-dimethylaminobenzoate.
Discussion
HMS and OD-PABA are important types of UV filters, but there is currently a lack of knowledge regarding the endocrine-disrupting effects of these compounds. For example, there is little information in the literature about the effects of exposure to these compounds on development during important critical windows, such as the prenatal, lactation, and postnatal periods. Similarly, the possible toxic effects of HMS and OD-PABA on the thyroid axis, which is the most important component of the endocrine system, have not been investigated. The effects of these compounds have been evaluated in previous studies that attempted to evaluate the effects of oral exposure on development (Schlumpf et al., 2001). However, sunscreen application constitutes the main source of human exposure to these compounds (Schauer et al., 2006). The present study was conducted to determinate the endocrine-disrupting effects of HMS and OD PABA in topical application conditions. Specifically, the prenatal, lactation and postnatal exposure periods, which are the most important critical windows of exposure to endocrine-disrupting compounds, were selected for evaluation in this study.
The UV filters were administered topically in the present study. Previous studies evaluating the effects of OD-PABA and HMS utilized the oral route of administration (Schlumpf et al., 2001). Similar applications have been discussed in evaluations of UV filters. Indeed, the dermal application of 3-BC has been determined to be much more effective (Søeborg et al., 2006). Additionally, a 3-fold greater estrogenic effect of 4-MBC has been observed in rats following topical application compared with oral exposure, which indicates a greater bioavailability of this compound due to biotransformation factors (Schlumpf et al., 2001; Völkel et al., 2006). The intradermal administration of 5% 4-MBC to immature rats increases the uterus weight (Schlumpf et al., 2001). The absorption and tissue penetration of HMS are known to be good following dermal application (Kim et al., 2014). The suggested dermal dose for humans is 2 mg/cm2, which is adequate for protection (Janjua et al., 2004; Stokes and Diffey, 1999), and this dose was topically applied to the rats in our study.
HMS and OD-PABA have been shown to have estrogenic and androgenic effects in in vitro studies (Schlumpf et al., 2001; Schreurs et al., 2002). It has been reported that immature rats exhibited uterotrophic effects in in vivo studies (Schlumpf et al., 2001; Schreurs et al., 2002); however, these findings have not been confirmed in other in vivo studies. The aim of the present study was to help determine the extent of these effects.
Endocrine disruption can also affect the timing of sexual maturation. Pre- and postnatal exposures to estrogenic chemicals typically delay sexual maturation in male offspring but cause precocious sexual maturation in female offspring. These effects have been noted after estradiol treatment (Biegel et al., 1998; Rodriguez et al., 1993) and in males exposed to UV filters, such as 4-MBC and 3-BC. However, UV filter exposure does not affect sexual maturation in female offspring, although it might cause significant alterations in the uterine and ovary weights (Schlumpf et al., 2004a). The estrogenic activities of BP-3, 3-BC, 4-MBC, and OMC have been confirmed in in vivo tests based on increased uterine weights in immature rats (Schlumpf et al., 2001, 2004b; Tinwell et al., 2002) and oophorectomized rats (Klammer et al., 2005). When these 3 UV filters are orally administered to immature rats, they exert estrogenic activity and increase the uterus weights in a dose-dependent manner.
In our study, we found that there was no difference in the vaginal opening times of the female rats that were exposed to UV filters in the prenatal and infancy periods. This result parallels those of studies that have stated that the time of the vaginal opening in female rats is not affected by UV filters. In the lactation group, vaginal opening was earlier compared to the control group. The uterus weights were not affected in any of the 3 study groups. We found that the estrogen, LH, and FSH levels were increased in the groups exposed to HMS and OD-PABA during the prenatal period. We observed that the estradiol levels of the female rats that were exposed to HMS in the lactation and early infancy periods were reduced, whereas the LH and FSH levels were not affected. Another study found that the uterus weight at PND 28 does not increase in female rats that have been exposed to OMC in the prenatal and postnatal periods, but the estradiol and progesterone levels decreased (Axelstad et al., 2011). That study also reported that the female rat reproductive system was affected. The vaginal opening time of female rats exposed to 4-MBC, which is known to have powerful estrogenic effects, during the prenatal period is 3 days earlier compared with controls, the gonadal weights are affected, and the serum LH and FSH levels increased. Exposure to 4-MBC during the prenatal period alters estrogen receptor expression in the adenohypophysis of the female rats, and this effect has been proposed to result from a positive feedback effect on gonadotropin secretion (Carou et al., 2009). For a compound to be considered to have an estrogenic effect, it is expected to elicit an increase in uterus weight and changes in estrogen-dependent parameters (Gelbke et al., 2004, 2007). Histological evaluations of the ovaries and uterus are essential for female pubertal assays (Gelbke et al., 2007). In our study, the numbers and structures of the corpus lutea in the ovaries, the number of necrotic ova, and the uterus histopathology were compatible with the phase of estrus and the findings in the control group.
Preputial separation is an early, reliable marker of the progression of puberty and an androgen-dependent external finding of sexual development in the male rat (Korenbrot et al., 1977). Thus, delays in PPS can indicate the antiandrogenic effect of endocrine disruptors (Monosson et al., 1999). In our study, there were no delays in PPS in any group. Reduced weights of the accessory sex organs, prostate, and seminal vesicles (including the secretory contents) are the most consistent indicators of the androgen environment (Chapin and Creasy, 2012). In the present study, we found that the weights of the seminal vesicles decreased in the male rats that were exposed to HMS during the prenatal period. The testis and prostate weights were not affected in the study groups.
Delays in male puberty and reduced prostate weight have been found to be the most sensitive indicators of reproductive toxicity following exposure to 3-BC and 4-MBC in extended, one-generation developmental studies in which rats were orally exposed for 10 weeks before mating, during pregnancy, and during lactation; thereafter, the offspring were subjected to continued oral exposure until adulthood (Durrer et al., 2007; Schlumpf et al., 2004a).
Spermatogenesis is initiated by hypothalamic stimulation due to LH and FSH secretion. Leydig cells produce testosterone, but sperm are generated in response to the FSH stimulus. LH plays a primary role in testosterone production. Increases in the circulating testosterone levels decrease the negative feedback and LH levels (Chapin and Creasy, 2012). The estrogenic effect of the central negative feedback mechanism in male rats acts on the hypothalamus to decrease the LH levels, which consequently results in reduced testosterone levels (Andrews et al., 2002; Roy et al., 1999; Rochira et al., 2006). BP-3, 3-BC, 4-MBC, HMS, OMC, and OD-PABA exhibit antiandrogenic activities in vitro, although the data regarding the individual compounds are conflicting. In contrast to other UV filters, which are primarily androgen antagonists, HMS exhibits both full agonistic and antagonistic androgen activities in vitro due to its full dose–response curve when binding to the human androgen receptor (hAR) and its inhibition of dihydrotestosterone action (Krause et al., 2012). In addition to its in vitro antiandrogenic activity, OMC has been shown to decrease the serum testosterone levels in immature rat offspring in a developmental study. Moreover, this study found that the testis and prostate weights were decreased and that these organs exhibited significant histopathological changes (Axelstad et al., 2011). The androgenic or antiandrogenic effects of UV filters have been tested in vitro, with the exception of OMC, for which the in vivo effects are not known (Krause et al., 2012). The testosterone levels were decreased in male rats that were exposed to OMC in the prenatal period. In vitro studies have revealed that OMC, which has an antiandrogenic effect, also has an in vivo effect. Moreover, this study found that OMC elicited decreases in the testis and prostate weights and significant histopathological changes in these organs (Axelstad et al., 2011). In the present study, we observed no significant differences in the histopathologic evaluations of the study groups. Moreover, the testosterone levels were reduced in the male rats that were exposed to OD-PABA and HMS during the prenatal period, whereas the LH levels increased, which indicated that exposure to this compound did not cause a predominantly estrogenic effect. Moreover, the decrease in the testosterone level may be attributed to an antiandrogenic effect. A decrease in testosterone is compatible with the delay in the PPS time and the decrease in seminal vesicle weight in the male pups that were exposed to HMS during the prenatal period because both parameters are dependent on androgen. These results are consistent with those of other studies that have reported the relationships between the serum testosterone levels with androgen-dependent parameters (Laws et al., 2007; Stoker et al., 2002).
Infants are exposed to UV filters through breast milk. In one study, the highest concentration of 4-MBC found in human milk was 48.37 ng/g of lipids (Schlumpf et al., 2010), which is nearly four times lower than the concentration of 4-MBC in rat milk (208.6 ng/g lipid) (Schlumpf et al., 2008). A pubertal delay and prostate weight reductions were observed in male rats exposed to 4-MBC and 3-BC during the lactation period (Durrer et al., 2007). In a study that also included OD-PABA and HMS, the UV filters were present in the breast milk (Schlumpf et al., 2008). In a study by Axelstad et al. (2011), the testosterone levels decreased in male rats exposed to OMC during the postnatal period. In our study, we did not observe any influence after the lactation period in the male or female rats.
The effects of EDs on the thyroid were evaluated based on developmental parameters. According to the Endocrine Disruptor Screening and Testing Advisory Committee of the EPA (Gray, 1998) and the enhanced OECD Test Guideline 407 (Andrews et al., 2002; Gelbke et al., 2007), thyroid toxicity was evaluated during male and female pubertal development. In these guidelines, the weight and histology of the thyroid gland and the serum T4 and TSH levels are the major parameters that are affected by EDs (Gelbke et al., 2007; Schmutzler et al., 2007). Endocrine disruptors affect thyroid function by decreasing iodide uptake, decreasing thyroid peroxidase activity, serving as transuterine agonists or antagonists, or decreasing thyroid hormone distribution (Gelbke et al., 2007). OMC, 4-MBC, and BP-3 are the best known UV filters that influence the hypothalamic–pituitary–thyroid axis (Axelstad et al., 2011; Durrer et al., 2007; Maerkel et al., 2007; Klammer et al., 2005; Krause et al., 2012). Exposure to OMC and 4-MBC during the prenatal period may influence both the developing reproductive and thyroid hormone systems of offspring rats (Axelstad et al., 2011; Carou et al., 2009). It has been reported that male offspring rats that are exposed to OMC during the pre- and postnatal periods exhibit reduced body weights and T4 and testosterone levels. The weight of thyroid gland increased in both sexes, whereas the weights of testis and prostate decreased, and the testis and prostate histology were also affected (Axelstad et al., 2011). Moreover, in high-dose OMC exposure experiments, the T4 levels are decreased in a dose-dependent manner, but the TSH levels do not change (Klammer et al., 2005). Additionally, 4-MBC and OMC have been found to have adverse effects on the thyroid axis in 90-day exposure studies (Schmutzler et al., 2004, 2007). An extended, one-generation developmental study in rats confirmed that alterations in the TSH and T3 levels following 4-MBC exposure result in increased thyroid weights in the offspring (Maerkel et al., 2007). The administration of low-dose 4-MBC causes goiters in the treated rats and their offspring (Schlumpf et al., 2004a).
The evaluation of the histology of the thyroid gland is important in ED investigations. Follicular colloid and follicular cell responses against thyrotoxicants are important. Although no histological evaluation was performed in our study, no increases were observed in the thyroid gland weights or decreases in the levels of T4 in the female or male offspring.
The thyroid gland plays significant roles in male and female pubertal development. Hypothyroidism causes delays in sexual maturation and the onset of puberty in male rats. Additionally, hypothyroidism can lead to atrophy of the testes and prostate and reduced gonadotropin and testosterone levels (Tamura et al., 1998). Moreover, pubertal development and gonadal hormones affect the thyroid gland. Androgen-sensitized thyroid follicular cells are affected by TSH (Banu et al., 2002). Thyroid endocrine functions begin at PND 16 in rats, and TSH exhibits a bimodal pattern during the postnatal period (Choksi et al., 2003). The initial peaks in TSH are observed during the first 2 phases of life, the TSH level peaks again from PND 30 to 50, and normal levels are reached by PND 60 (Goldman et al., 2000). Thyroperoxidase expression is higher in young male rats than female rats (da Costa et al., 2001), which indicates that as the gonadal hormone levels decrease in aging rats, a concomitant decrease in thyroid peroxidase occurs. However, there is little evidence indicating that the serum TSH and T4 concentrations fluctuate over the estrous cycle (Fukuda et al., 1975; Kieffer et al., 1976).
In our study, the thyroid axes and pubertal development periods of the male and female offspring rats were not affected.
Conclusions
The maximum permitted applications of dermal OD-PABA and HMS to the mothers during the prenatal and lactation periods and to both the female and male pups during infancy did not disrupt the thyroid gland, its function, the pubertal developmental stage, or the gonadal steroid levels. Today’s modern life makes exposure to endocrine disruptors inevitable. Having knowledge about the toxicity profiles of these compounds, we are exposed to intentionally or unintentionally, will protect us from the health risks that may arise. The results obtained in this study will contribute to the formation of toxicological information about OD-PABA and HMS. The previous studies have shown that other UV filters such as OMC, 4-MBC, and 3-BC affected pubertal development and thyroid functions. Future exposure studies completed on the dermal route, the main route of UV filters exposure, will shed some light on the better understanding of toxicological profiles of these compounds.
Footnotes
Acknowledgments
The authors wish to thank the Bagcilar Training and Research Hospital for providing support, materials, and equipment for the work.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.