
Abstract
Phenolic compounds of essential oils from the family Lamiaceae are commonly used substances in the food industry because of their flavouring, antimicrobial and antioxidant properties. In this context, it has become important to have healthy and safe products for consumers who are exposed to these phenolic compounds. The present study was aimed to investigate the toxic effects of carvacrol, thymol and their mixture on human gastric carcinoma (AGS) cells. Cells were analyzed after 24 h of exposure to different concentrations of carvacrol, thymol and their mixture by the ATP cell viability, 2′,7′ dichlorodihydrofluorescein diacetate (H2DCF-DA), reducte glutatione/oxide glutathione ((GSH)/GSSG-Glo) and comet assays. Apoptosis induction was studied by acridine orange/ethidium bromide staining and western blotting. Carvacrol, thymol and their mixture induced cytotoxicity, genotoxicity, apoptosis, increased reactive oxygen species (ROS) and decreased GSH levels after 24 h of their exposure in a dose-dependent manner. A close negative relationship was found between cell viability and ROS generation. We examined dose-dependent cytotoxic effects of carvacrol, thymol and their mixture in human AGS cells. Increased intracellular ROS causes oxidative stress in cells. The results indicated that these compounds should be used carefully in the food industry.
Introduction
A recent food packaging technology namely ‘active packaging’ allows the use of active substances in the food packaging films. This convenient method consists of incorporation of some antimicrobial substances into the food packaging films. Microbial protection of food is facilitated through the stability of the antimicrobial substances including essential oils, ensuring the control of their release into the food over time (Li et al., 2006). However, the use of such packaging materials is not a substitute for good sanitation practices. They should enhance food safety as an additional hurdle for the growth of pathogenic microorganisms (Pérez-Pérez et al., 2006). On the other hand, a too slow release of antimicrobial substances might cause insufficient microbial inhibition in the food, while their too fast release might be the reason why the inhibition is not sustained. The release rate of antimicrobial substances used in food packaging depends on the environmental conditions, the polymer type, the film preparation method and the interactions between polymeric and antimicrobial materials (Cagri et al., 2004). Therefore, the antimicrobial substances used in the food packaging are also important in terms of how safe they are for human consumption.
Carvacrol (5-isopropyl-2-methylphenol) and thymol (2-isopropyl-5-methylphenol) are two major phenolic compounds of most essential oils from the family Lamiaceae plants (Llana-Ruiz-Cabello et al., 2014a). Like other phenolic monoterpenes, both compounds also exhibit a range of biological activities including anticancer, antimicrobial, antifungal and antioxidant effects. They have been widely used alone or in combination in medical practice as antiseptics; in agriculture against nematodes; in cosmetics for bath, skin, personal care, hair products, hair dyes and mouthwash products; and in the food industry especially for food packaging (Llana-Ruiz-Cabello et al., 2015a). Microencapsulation of both compounds into polymer films used for food packaging was recommended because of their antimicrobial activity (Guarda et al., 2011).
According to good manufacturing practices, carvacrol and thymol can be utilized as flavouring substances in the food industry without any restriction (Regulation (EC) 1334/2008). Both compounds are accepted safe for human consumption by the European Commission (Commission Decision, 1999) and additionally as approved food additives by the US Food and Drug Administration (FDA, 2013). However, their use in lower concentrations is for food flavour agents, while their higher concentrations were applied to food packaging systems (Stammati et al., 1999). In the course of time, carvacrol and thymol may release from the packaging material into the food, and humans consume foods exposed to these compounds at higher concentrations. The use of these compounds in combination has showed high antibacterial effect in vitro (Pei et al., 2009). Additionally, the mixture of carvacrol/thymol should also be evaluated in order to determine its safety for consumers. Some in vivo studies reported acute and subacute toxic effects of these compounds in different animal species (Hagan et al., 1967; Jenner et al., 1964). However, the toxic mechanism of these compounds has not been completely elucidated so far. Therefore, considering the increasing use of these compounds in food packaging technology, new studies are required to understand their safety for human health (Llana-Ruiz-Cabello et al., 2015b; Stammati et al., 1999).
On the other hand, some studies were focused on their beneficial effects, but besides these effects they may also have toxic ones, such as cytotoxic, genotoxic/mutagenic, apoptotic and pro-oxidant activity. Different in vitro models have been used to assess the toxicity of these compounds (Llana-Ruiz-Cabello et al., 2015a). Horváthová et al. (2006) reported cytotoxic effects of carvacrol and thymol on hepatoma (HepG2) and colon carcinoma (Caco-2) cells, which were not associated with a genotoxic effect. An electron microscope observation of Caco-2 cells exposed to thymol resulted in ultrastructural changes of cells such as lipid degeneration, mitochondrial damage, nucleolar segregation and apoptosis (Llana-Ruiz-Cabello et al., 2014a). However, other investigations have described apoptosis induction by carvacrol and thymol on different cancer cell lines (Arunasree, 2010; Deb et al., 2011; Fan et al., 2015; Hsu et al., 2011; Kang et al., 2016). Apoptosis is a programmed gene-directed form of cell death and is characterized by specific morphological and biochemical properties (Fan et al., 2015). Several protein families, including the upstream Bcl-2 family (e.g. the antiapoptotic Bcl-2 and proapoptotic Bax) and the downstream caspase family (e.g. caspase 9 and caspase 3), play roles in apoptosis cases. Carvacrol-induced apoptosis was reported by direct activation of the mitochondrial pathway on HepG2, breast cancer (HeLa and SiHa), Caco-2 (HCT116 and LoVo) and gastric adenocarcinoma (AGS) cells (Arunasree, 2010; Fan et al, 2015; Gunes-Bayır et al., 2017; Yin et al., 2012). The same mechanism was also reported for thymol on human glioblastoma, promyelocytic cancer and AGS cells (Deb et al., 2011; Hsu et al., 2011; Kang et al., 2016).
Free radicals, namely reactive oxygen species (ROS), were produced by the metabolism of oxygen (Mahalingam and Fedoroff, 2003). They have roles in the destruction of macromolecules and the modifying of cell signaling and other functions. Overproduction of ROS was seen at the onset and progression of cancers and chronic diseases. Generation of ROS can lead to the permeabilization of the mitochondrial membrane and thus induce apoptosis by releasing apoptotic proteins. At the same time, some plants including their extracts or essential oils affect the generation of ROS, and so may activate apoptosis. Carvacrol- and thymol-induced oxidative stress is also a toxic mechanism for some cell lines (Ozkan and Erdogan, 2012; Undeger et al., 2009). In our previous study, it was demonstrated that carvacrol has pro-oxidant activity through increased ROS generation and decreased glutathione (GSH) levels in human AGS cells (Gunes-Bayır et al., 2017). GSH is an antioxidant that plays a role in the scavenging ROS in cells. Thymol-induced ROS generation in AGS cells was also reported without any performed significance analyses (Kang et al., 2016). However, little is known with respect to the mixture of carvacrol and thymol. Only one study reported the ROS generating effect of this mixture on human Caco-2 cells (Llana-Ruiz-Cabello et al., 2015b). Additionally, the genotoxic effect of carvacrol, thymol and their mixture on AGS cells has not been examined so far. A few genotoxicity investigations were reported for carvacrol and thymol on human Caco-2, HepG2 and lymphocytes (Aydin et al., 2014; Horváthová et al., 2006; Llana-Ruiz-Cabello et al., 2014b), but not on human AGS cells.
Earlier, it was reported that both substances are rapidly metabolized and excreted in rats (Austgulen et al., 1987). Their excretion after 24 h was very limited and the molecules were unchanged. Ring hydroxylation of these compounds was identified, and that is the reason why thymol and carvacrol metabolism is fast. The biological activities of polyhydroxylated compounds generally seem to be dependent on their chemical properties such as structure and lipophilicity, which can affect their uptake into cells or influence their interaction with proteins and enzymes. A previous study demonstrated that thymol and carvacrol were almost completely absorbed in the stomach and proximal small intestine of pigs (Michiels et al., 2008). Therefore, it is important and useful to investigate both compounds and their mixture for their toxic mechanisms in the stomach cell line AGS, which is a commonly used gastric model for human stomach research (Moese et al., 2004).
Considering the limited number of studies on toxic effects and increased implementation of carvacrol, thymol and their mixture in the food industry, the evaluation of their safety is very important for human gastric cells. Therefore, the aim of this study was to investigate, for the first time, the cytotoxic, genotoxic, apoptotic and ROS generating effects caused by wide range of concentrations of major polyphenols from the family Lamiaceae plants on human gastric cells together.
Materials and methods
Cell culture
Human AGS cells derived from human Caucasian AGS cells were purchased from the American Type Culture Collection (Manassas, Virginia, USA). Cells were cultured in Ham’s F-12 K (Kaighn’s) Medium supplemented with 10% fetal bovine serum (FBS) and antibiotics (100 U/ml penicillin, 100 µg/ml streptomycin) at 37°C in a humidified atmosphere of 5% carbon dioxide (CO2). Cells were grown to near confluence in 75-cm2 plastic flasks and harvested weekly.
Chemicals and reagents
FBS, penicillin-streptomycin (10,000 U/ml), 0.25% Trypsin-EDTA and phenol red were supplied by Life Technologies (Life Tech, California, USA). Ham’s F-12 K (Kaighn’s) Medium was obtained from Gibco Invitrogen Corporation (Carlsbad, California, USA). GSH/GSSG-Glo assay was obtained from Promega (Madison, Wisconsin, USA). Bax (N-20), Bcl-2 (C 21), caspase 3 (H-277) and caspase 9 (H-170) antibodies were provided by Santa Cruz Biotechnologies (Santa Cruz, California, USA). β-actin (clone AC-15), 2′,7′ dichlorodihydrofluorescein diacetate (DCFH-DA), ethidium bromide (EB), acridine orange (AO) solution, carvacrol and thymol used in this study were provided by Sigma Chemical Co. (St Louis, Missouri, USA).
Test solutions
Test solutions were prepared according to Llana-Ruiz-Cabello et al. (2014a, 2014b, 2015b) and Fan et al. (2015). Carvacrol and thymol were separately mixed in 100% DMSO, which made the stock solutions of carvacrol (600 µM) and thymol (600 µM). The different concentrations (10, 20, 30, 50, 100, 200 and 400 µM) of both compounds were prepared from the stock solutions in DMSO. For the preparation of the mixture of both compounds, carvacrol and thymol were mixed in a proportion of 10:1 (0-200:0-20 µM). Test solutions were added to the cell culture medium, and so the final concentration of DMSO in cell culture medium was 0.1%.
Determination of cytotoxicity
The CellTiter-Glo Luminescent Cell Viability Assay (Promega; Madison, Wisconsin, USA) was utilized, which is based on quantitation of the ATP presence in cells; 100 µl of complete medium and 15 × 103 cells were placed in each well of a 96-well microtiter plate. Cells were treated without (the negative control) or with test solutions of carvacrol, thymol and their mixture. After 24 h of incubation at 37°C, CellTiter-Glo Reagent was added into each well containing AGS cells. The amount of ATP was determined by a luminometer (Varioskan Flash Multimode Reader, Thermo; Waltham, Massachusetts, USA). The light emitted in the presence of ATP was quantitated in relative light units. The intensity of emitted light quants was directly related to the ATP content in the tested sample. The cell viability was expressed as a percentage compared with the negative control group designated as 100% (Scudiero et al., 1988). The half maximal growth inhibitory concentration (IC50) value of carvacrol, thymol and their mixture was estimated from the concentration–response curves by non-linear regression analysis. All experiments were repeated three times and standard deviation (SD) was within 5%.
Determination of ROS activity
The intracellular ROS generation was determined by the oxidant-sensitive fluorescent probe DCFH-DA assay; 200 µl of complete medium and 15 × 103/ml AGS cells were placed into a 96-well plate. After 24 h of incubation, growth media was replaced with growth medium containing reduced FBS (5%, v/v). Culture plates were incubated in the absence (negative control) or presence of different test solutions from carvacrol, thymol and their mixture (10:1) for further 24 h. The cells were washed with cold phosphate buffered saline (PBS) and incubated with 100 mM DCFH-DA for 30 min at 37°C. Fluorescence intensity was measured using the fluorescence plate reader (Varioskan Flash Multimode Reader; Thermo) at E x/E m = 488/525 nm. The measurements were performed triplicate for ensuring reproducibility. The result of the assay was expressed as percentage of the relative fluorescence with respect to the control group.
Determination of GSH activity
The GSH/GSSG-Glo™ Assay (Promega; Madison, Wisconsin, USA) was utilized to determine and quantify GSH levels in AGS cells in this study. The assay is based on the luminescence measurements in multiwell plates; 100 µl of complete medium and cells at a density of 15 × 103/ml were placed in each well of a 96-well microtiter plate. Cells were treated without (the negative control) or with test solutions of thymol. After 24 h incubation, of AGS cells at 37°C, treatments were removed and the total GSH Lysis Reagent was added into each well. Luciferin Generation Reagent was added to all wells, and the well contents were mixed. After 30 min of incubation, Luciferin Detection Reagent was added to all wells; assays are mixed and incubated for 15 min. The amount of the GSH was measured using a luminometer (Varioskan Flash Multimode Reader; Thermo). The light emitted in the presence of GSH was quantitated in relative light units. The intensity of emitted light quants was directly related to GSH content in the tested sample. All experiments were repeated three times, and the SD was within 5%.
Determination of apoptosis
AO/EB fluorescent staining was used to study the apoptotic activity of carvacrol, thymol and their mixture (10:1) at the cellular level. Carvacrol (0-100 µM), thymol (0-100 µM) and their mixture (0-100:0-10 µM) were added and incubated three times for 24 h following the placing 18 × 104/ml of AGS cells in a 6-well plate. The cells were harvested and washed twice with PBS. A solution of AO/EB dye was added to the cell suspension, which was evaluated for nuclear morphology using a fluorescence microscope (Leica DM 1000; Solms, Germany). Finally, cells having normal green nuclei as viable, cells having green nuclei with fragmented chromatin as apoptotic and cells having orange/red nuclei as necrotic were determined. According to this method, multiple photos were taken at randomly selected areas, and a minimum of 100 cells was counted.
Apoptotic activity of both compounds and their mixture at the protein expression level was examined by western blotting as described earlier (Gunes-Bayır et al., 2017). All samples were also blotted for normalizing protein amounts of β-actin. The expression level of proteins was detected using Amersham ECL Plus Western blotting detection reagents (GE Healthcare, Piscataway, New Jersey, USA) and captured with imaging system (Vilber Lourmat, France).
Determination of genotoxicity
Genotoxic effects of carvacrol, thymol and their mixture (10:1) was investigated using a comet assay according to the method of Singh et al. (1988) with a slight modification. Before carrying out the comet assay, a trypan blue exclusion test was performed to determine the number of viable cells present in a cell suspension (Strober, 2001). Approximately 2 × 105 cells per well were seeded into 6-well plates with cell culture medium and incubated at 37°C in 5% CO2. After 24 h of incubation, below IC50 concentrations of carvacrol, thymol and their mixture (10:1) were added onto the cells and incubated for a further 24 h at 37°C. DMSO (1%) was used as a negative control, and 50 µM hydrogen peroxide was applied as a positive control. Cells were washed with PBS and harvested using trypsin/EDTA. Cells were collected for centrifugation at 400 × g for 5 min at 4°C. The supernatant was discharged, and the cell density was adjusted to 2 × 105 cells/ml using cold PBS; 10 ml of cell suspension were mixed with 90 µl of 0.6% low-melting agarose and added to the slides pre-coated with 1% normal melting agarose. After solidification of the agarose, the slides were immersed in lysis solution (2.5 M sodium chloride, 100 mM Na2EDTA, 10 mM Tris-HCl, pH 10, 1% Triton X-100 and 10% DMSO at 4°C) for 1 h for removing cellular proteins. After that, slides were washed with cold PBS and placed in a horizontal electrophoresis box. DNA was allowed to unwind for 40 min in freshly prepared alkaline electrophoresis buffer containing 300 mM sodium hydroxide and 10 mM Na2EDTA (pH > 13). After the unwinding time, the electrophoresis was run at 300 mA for 25 min at 4°C under minimal illumination to prevent further DNA damage. The slides were washed three times with a neutralization buffer (0.4 M Tris, pH 7.5) for 5 min at 4°C and treated with ethanol for 5 min before staining. Dried microscope slides were stained with EB (2 µg/ml in distilled water; 70 µl/slide), covered with a coverslip and analyzed using a fluorescence microscope (Leica DM 1000; Solms) at a 200× magnification with epifluorescence. The microscope was equipped with a rhodamine filter (with an excitation wavelength of 546 nm; and a barrier of 580 nm). A computerized image analysis system (Comet Assay IV; Perceptive Instruments) was performed. According to Hartmann et al. (2003), the percentage of tail DNA was used as a parameter for measurement of DNA damage (DNA strand breaks). One hundred comets were scored per each sample in one electrophoresis. Additionally, all of these procedures were completed with the same biochemistry staff. DNA damage was detected using a single observer who was not aware of the subject’s status.
Statistical analyses
The Statistical Package for the Social Sciences version 23.0 (SPSS Inc, Chicago, Illinois, USA) computer program was used for statistical analysis. All experiments were performed in triplicate. The comparison of the results from the experiments was done by one-way analysis of variance, and post hoc analysis of dose differences was carried out by Tukey’s test. The results were presented as mean with SDs (mean ± SD) and calculated in percentages with respect to the control cells. A p value of ≤ 0.05 was considered to be statistically significant. Additionally, a non-linear regression analysis was performed by concentration–response curve in order to estimate IC50 values for carvacrol, thymol and their mixture (10:1).
Results
Cytotoxic effect
Cell viability in AGS cell culture of three different experiments was greater than 95% before performing all experiments (Figure 1(a)). Human AGS cells exposed to carvacrol (10-600 µM) were investigated in cytotoxicity assays after 24 h of incubation. The assays showed significant changes in relation to the control group from 50 µM to 600 µM (p ≤ 0.001; Figure 1(b)). The IC50 value for carvacrol was 82.57 µM ± 5.58 µM.
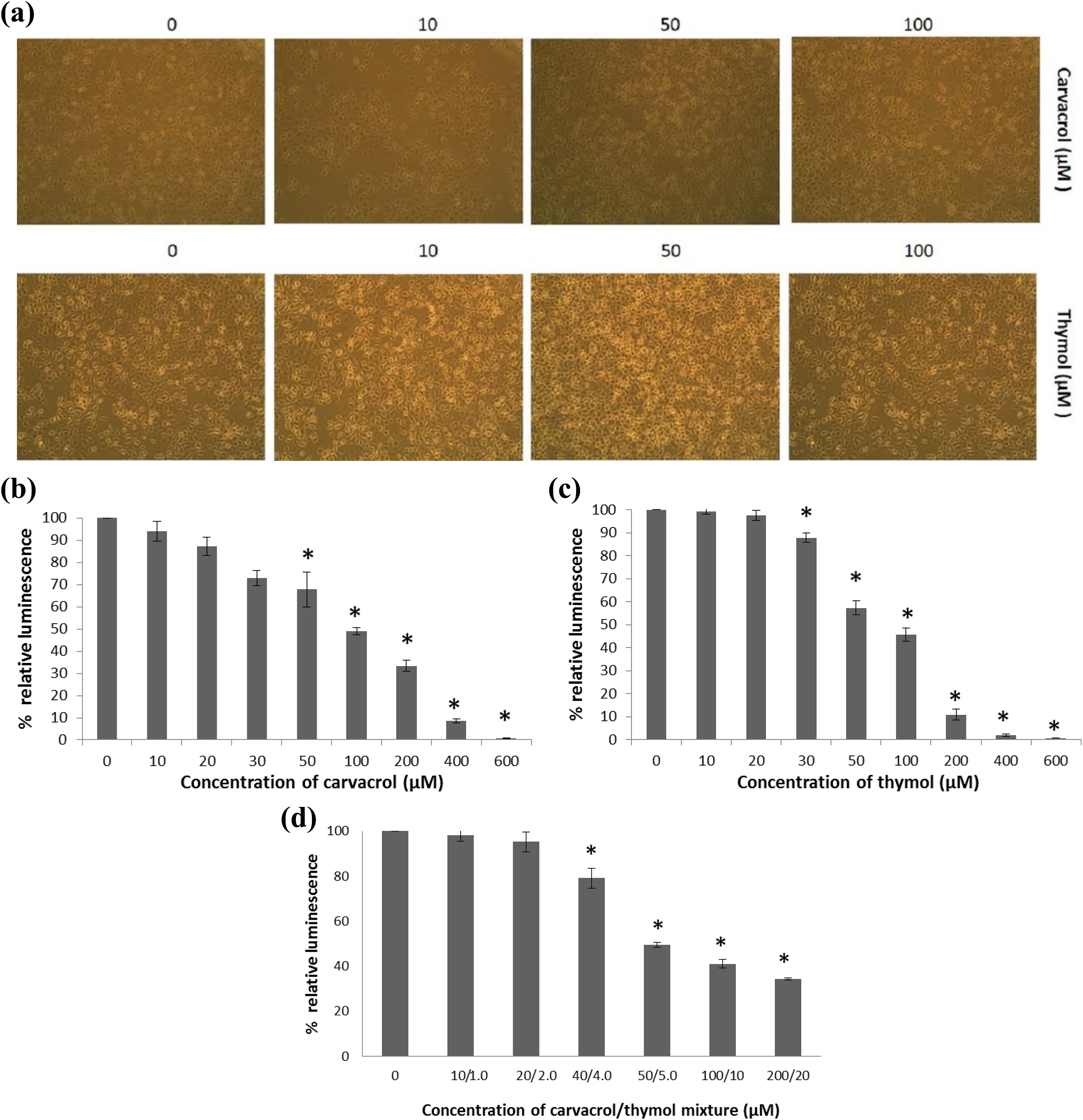
Cytotoxicity assay. (a) Human AGS cells before performing all experiments. The cytotoxic effect of carvacrol (0–600 µM) (b), thymol (0–600 µM) (c) and their mixture (0:0–200:20 µM) (d) on the human AGS cell line after 24 h of exposure. Both compounds and their mixture induced cytotoxicity in a dose-dependent manner. All values are expressed as mean ± SD. *p ≤ 0.001: significantly different from control cells. AGS: gastric adenocarcinoma; SD: standard deviation.
Cells exposed to thymol (10–600 µM) showed significantly reduced cell viability with respect to the control group (p ≤ 0.001; Figure 1(c)). A significant change was obtained between the control group and cells exposed to thymol (30–600 µM; p ≤ 0.001). The IC50 value for thymol was found 75.63 µM ± 4.01 µM.
Cells exposed to the mixture carvacrol/thymol, in a proportion 10:1, showed also significantly reducing cell viability in a dose-dependent manner (p ≤ 0.001; Figure 1(d)). Differences were found to be significant between the control cells and cells exposed to the mixture carvacrol/thymol (40:4–200:20 µM; p ≤ 0.001). The IC50 value for 10:1 mixture was found 84.71 µM ± 6.88 µM.
Further studies assessing apoptosis and genotoxicity were performed using 10-100 µM of carvacrol, thymol and their mixture in the selected proportions (10:1, 20:2, 40:4, 50:5, 100:10 and 200:20 µM).
Effect of carvacrol on ROS level
The measurement of ROS generation was carried out using the oxidation-sensitive probe DCFH-DA. Human AGS cells experienced a significant increase (p ≤ 0.001) in ROS levels when they were exposed to 50–600 µM of carvacrol (Figure 2(a)) and 30–600 µM of thymol (Figure 2(b)). An increase in ROS generation in the mixture carvacrol/thymol (40:4–200:20 µM) was also significantly seen in comparison to control cells after 24 h of their incubation (Figure 2(c)). A positive correlation was found between ROS generation and increased concentration of carvacrol, thymol and their mixture in human AGS cells.
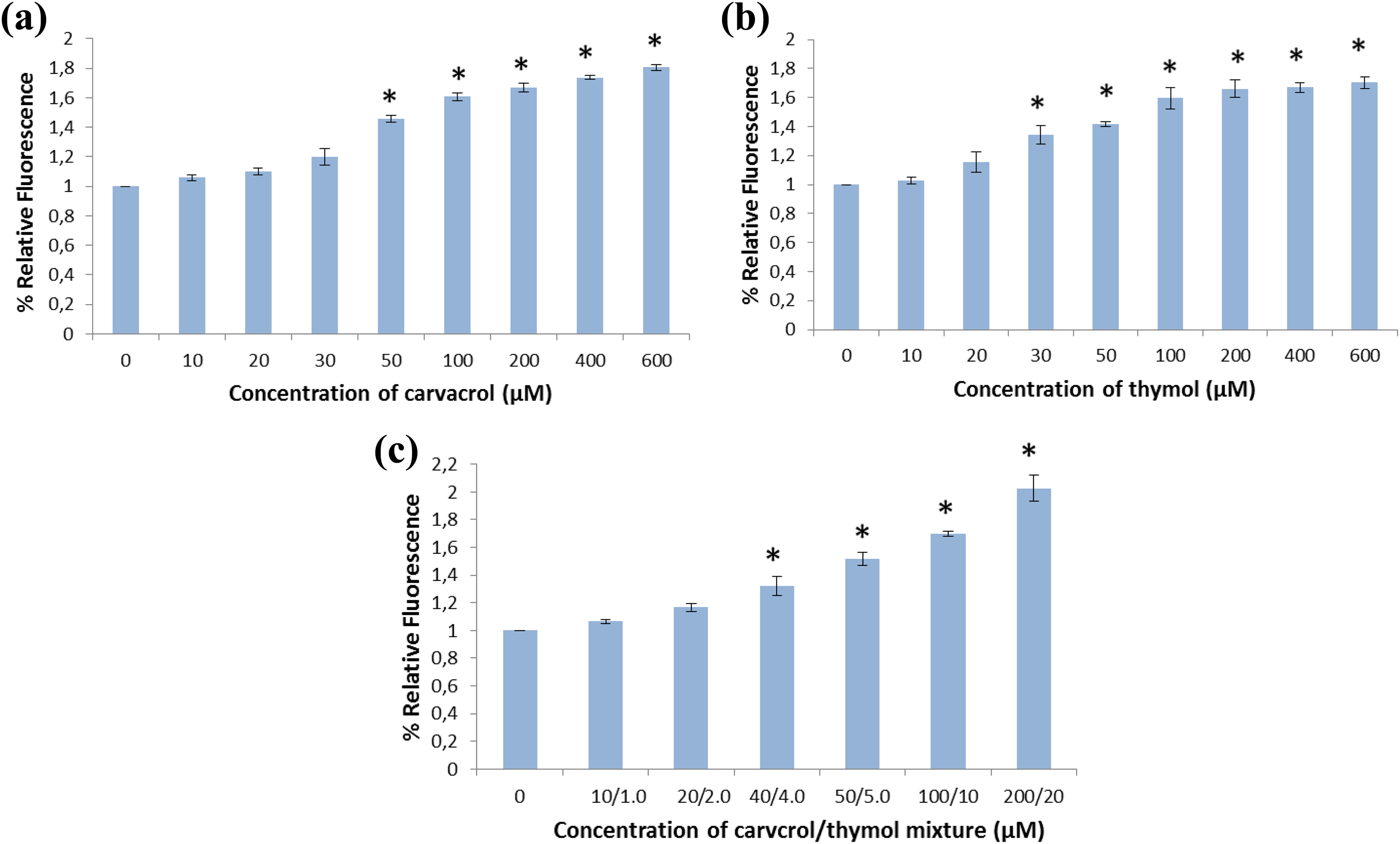
The DCFH-DA assay. (a) ROS level in human AGS cells exposed to carvacrol (0–600 µM) after 24 h of incubation. Carvacrol induced intracellular ROS generation in a dose-dependent manner. (b) ROS level in human AGS cells exposed to thymol (0–600 µM). Thymol induced intracellular ROS generation in a dose-dependent manner. (c) ROS level in human AGS cells exposed to mixture of carvacrol/thymol (0:0–200:20 µM). The mixture induced intracellular ROS generation in a dose-dependent manner. All values are expressed as mean ± SD. *Differences were considered significant compared to the control cells from p ≤ 0.001. ROS: reactive oxygen species; AGS: gastric adenocarcinoma; SD: standard deviation; DCFH-DA: 2′,7′ dichlorodihydrofluorescein diacetate.
Effect of carvacrol on GSH level
GSH level was investigated by the GSH/GSSG-Glo Assay. Carvacrol, thymol and their mixture were found to reduce GSH levels in the human AGS cells after 24 h of incubation. GSH level was significantly decreased after carvacrol exposure (Figure 3(a)). Significant changes in GSH levels were observed between the control and carvacrol (20–100 µM) exposed cells (p ≤ 0.001). The measured relative luminescence was shown in percentages (Figure 3(b)). Significant changes in GSH levels were also observed between the control and thymol (20–100 µM) exposed cells (p ≤ 0.001). Also, significant differences were recorded at 10:1.0–100:10 µM of the mixture (p ≤ 0.001; Figure 3(c)). A negative correlation was found between relative GSH level and increasing dose of both compounds and their mixture.
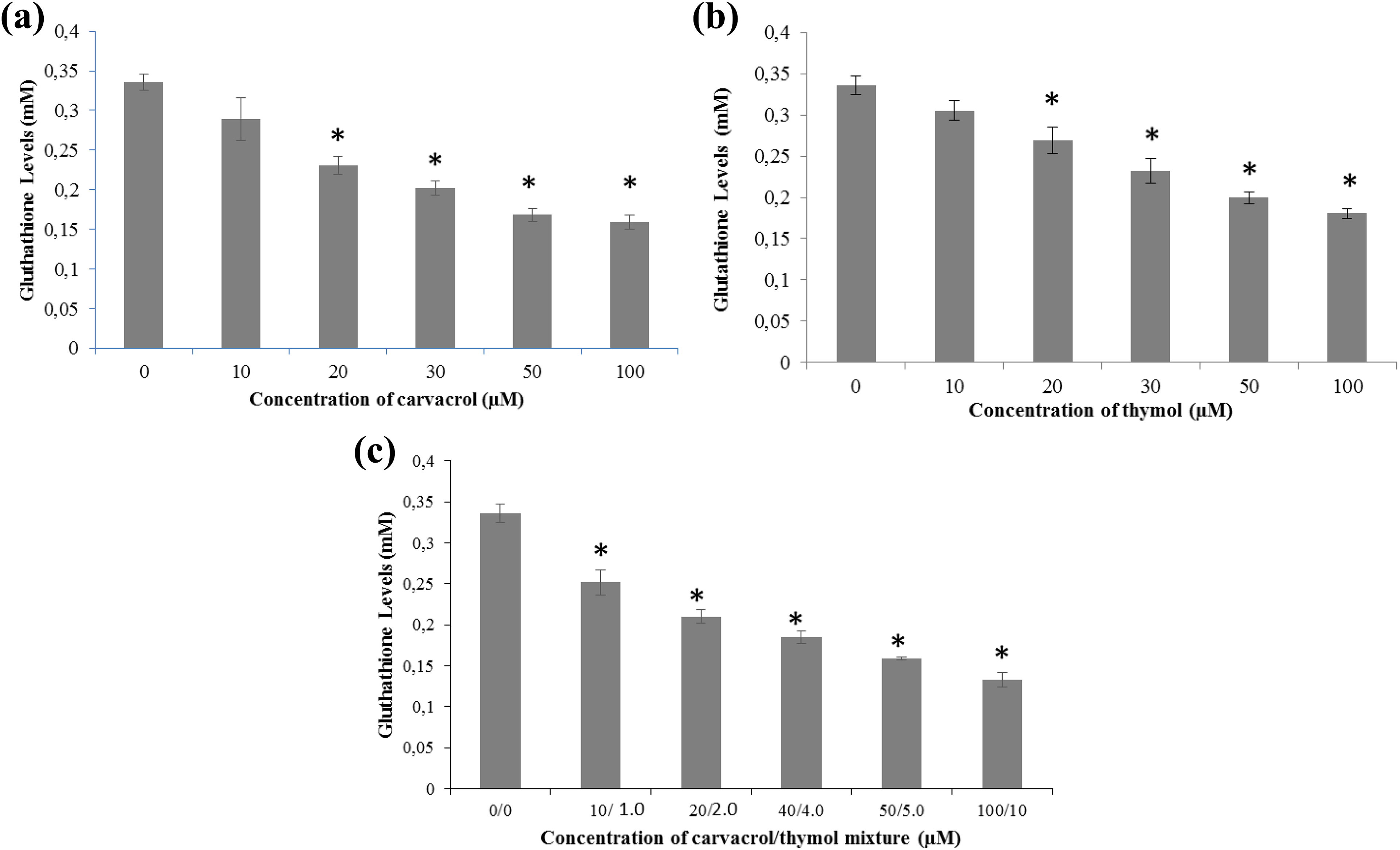
GSH levels in AGS cells after 24 h of exposure to carvacrol (0–100 µM) (a), thymol (0–100 µM) (b) and their mixture (0–100:10 µM) (c). All values are expressed as mean ± SD. *p ≤ 0.001: differences were considered significant compared to the control group. GSH: glutathione; AGS: gastric adenocarcinoma; SD: standard deviation.
Apoptotic effect
An AO/EB staining was performed to evaluate viable, apoptotic and necrotic cells after 24 h of exposure to different concentrations of carvacrol, thymol and their mixture. It was found that cell morphology became abnormal with cell nuclear shrinkage and chromatin condensation after treatments (Figure 4(a)). Significant changes with respect to the control group were observed for viable, apoptotic and necrotic cells after AGS cells exposed to carvacrol, thymol and their mixture (p ≤ 0.001; Figure 4(b) to (d)). At the concentration of 100:10 µM of the mixture, the percentage of apoptotic cells was higher than their viable and necrotic cells. The results showed that apoptotic and necrotic effects of both compounds and their mixture were in a dose-dependent manner.
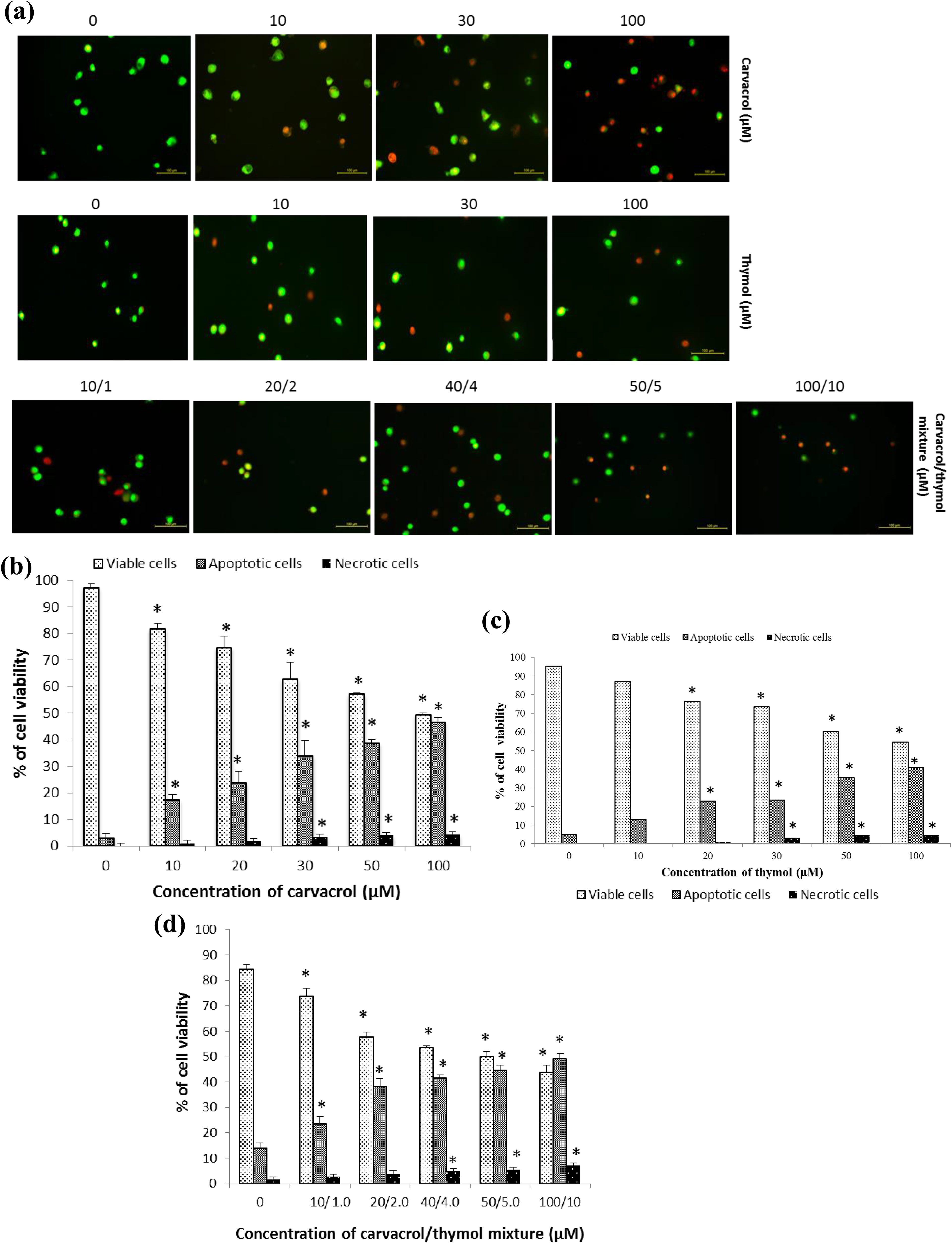
Detection of apoptosis. (a) V, A and N cells were detected using a fluorescence microscope observation following the AO/EB staining. Significant changes in human AGS cells exposed to carvacrol (0–100 µM) (b), thymol (0–100 µM) (c) and their mixture (0:0–100:10 µM) (d) after 24 h of their exposure. The effect of both compounds and their mixture on the cell viability compared with control cells are given in percentages. All values are expressed as mean ± SD. Differences were considered significant with respect to the control cells from p ≤ 0.001. V: viable cells; A: apoptotic cells; N: necrotic cells; AGS: gastric adenocarcinoma; SD: standard deviation; EB: ethidium bromide; AO: acridine orange.
Apoptotic effects of both compounds and their mixture were studied at the protein expression level by western blotting. Apoptotic (caspase 9 and caspase 3), pro-apoptotic (Bax) and anti-apoptotic (Bcl-2) proteins play a role in the regulation of apoptosis. Therefore, their changes in cells with carvacrol (0–50 µM), thymol (0-50 µM) and their mixture (0:0–50:5 µM) were studied after 24 h of their incubation in AGS cells. The density of caspase 3, caspase 9 and Bax proteins was significantly increased, whereas the density of Bcl-2 protein was decreased in a dose-dependent manner (p ≤ 0.001; Figure 5 (a) to (c)). It was suggested that both compounds and their mixture cause apoptosis by decreasing Bcl-2 protein level and increasing Bax, caspase 3 and caspase 9 proteins’ level. β-actin was positive for all.
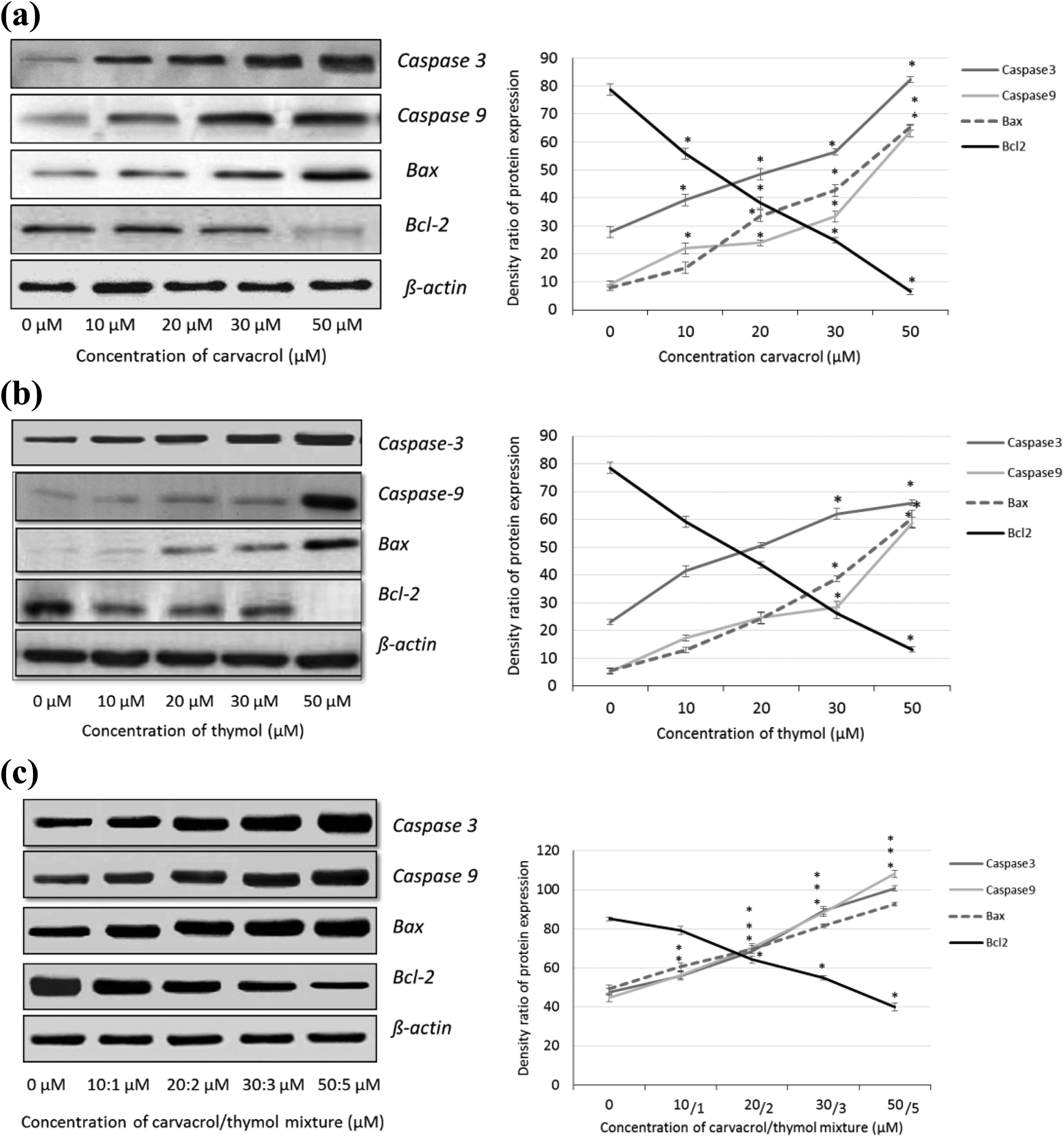
Detection of apoptosis by western blotting. Significant changes were found in AGS cells exposed to carvacrol (0–50 µM) (a), thymol (0–50 µM) (b) and their mixture (0:0–50:5 µM) (c) after 24 h of their exposure. For each experiment, the expression of Bax, Bcl-2, caspase 9 and caspase 3 in comparison with β-actin is demonstrated. Density ratio of expressed proteins is presented in percentages. All values are expressed as mean ± SD. *p ≤ 0.001: significantly different from the control cells. AGS: gastric adenocarcinoma; SD: standard deviation.
Genotoxic effect
After 24 h of exposure to different concentrations of carvacrol, thymol and their mixture in AGS cells, DNA damage was detected by a specific assay, the comet assay Hartmann et al. (2003). DNA damage was determined by tail intensity and was seen as a comet formation (Figure 6(a)). A dose-dependent genotoxic effect of carvacrol, thymol and their mixture was found. Significant changes in the tail of DNA were seen between the control group and cells exposed to carvacrol (30–100 µM), thymol (20-100 µM) and their mixture (20:2–100:10 µM; p ≤ 0.001; Figure 6(b) to (d)).
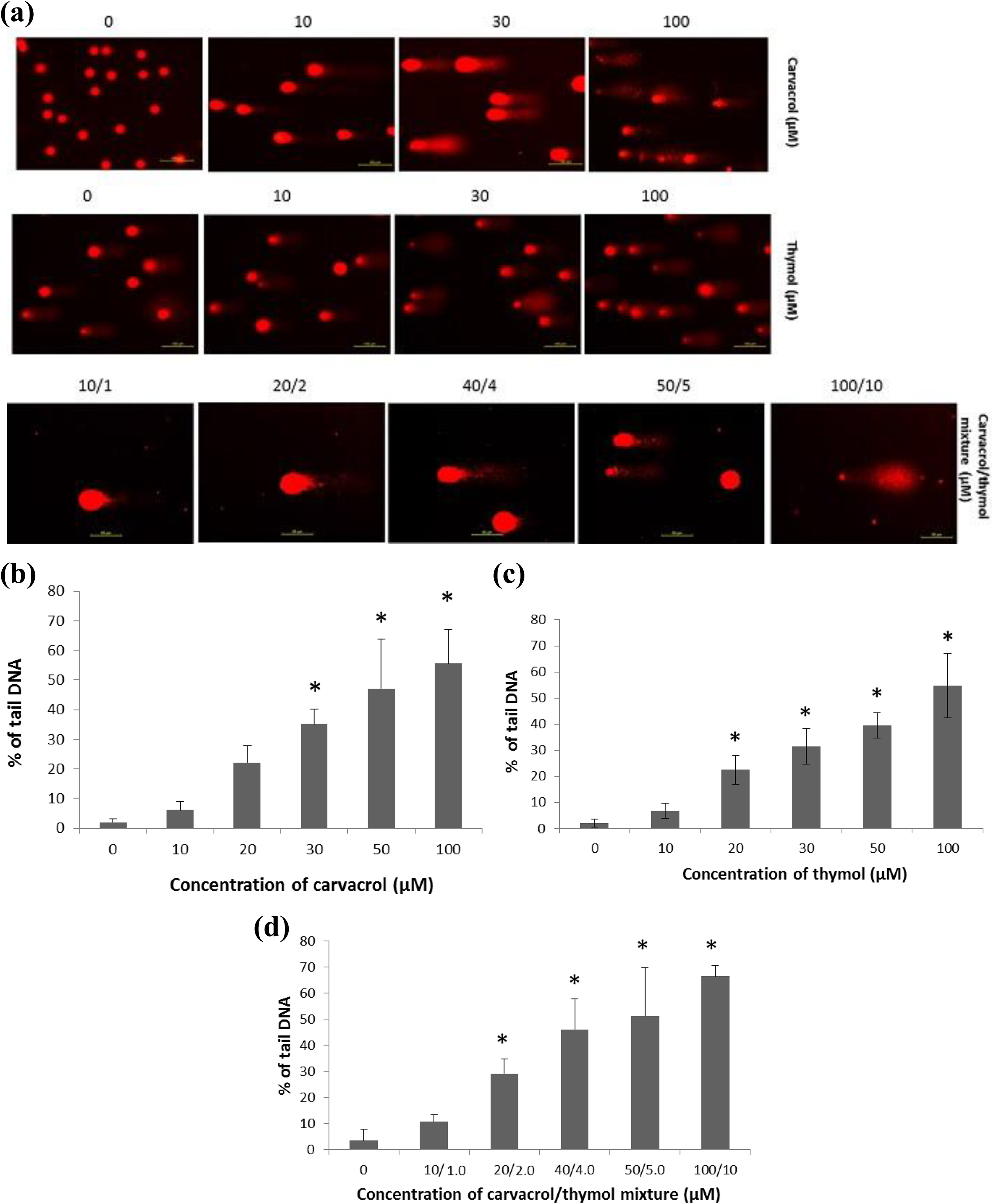
Comet assay. (a) Increased comet formation shows increased DNA damage. The genotoxic effect on human AGS cells exposed to carvacrol (0–100 µM) (b), thymol (0–100 µM) (c) and their mixture (0:0–50:5 µM) (d) after 24 h of incubation. Both compounds and their mixture induced DNA damage in a dose-dependent manner. The level of DNA damage is expressed as percentage of tail DNA. All values are expressed as mean ± SD. *p ≤ 0.001: significantly different from the control cells. AGS: gastric adenocarcinoma; SD: standard deviation.
Discussion
In the food industry, synthetic compounds used for food packaging such as butylated hydroxytoluene or butylated hydroxyanisole have caused pathological alterations, with teratogenic and carcinogenic potential in vivo (Branen, 1975). Therefore, alternative substances should be investigated for substitution of these synthetic compounds. Carvacrol and thymol at lower concentrations are usually used as flavouring substances in the food industry and are categorized as generally regarded as safe (GRAS) by US Food and Drug Administration (FDA, 2013). However, their use in food packaging may require higher concentrations, which increases the exposure of humans to these compounds (Stammati et al., 1999). Earlier, the usefulness of carvacrol and thymol as antimicrobial substances in food packaging was examined in order to determine their possible controlled release of active substances into the food over time (Kurek et al., 2014; Ramos et al., 2012, 2013). Because of the increased application of these compounds in the food industry, an increasing oral intake of carvacrol and thymol is feasible for humans. Additionally, according to European Commission Regulation No 450/2009, only authorized substances, which they included in the community list, may be used in the food industry (Regulation (EC), 2009). Interestingly, essential oils from the family Lamiaceae plants or their extracts used in food packaging were not included on this list. On the other hand, a safety assessment of substances intended to come into contact with food requires genotoxicity and sub-chronic toxicology investigations (EFSA, 2016). In this context, a subchronic 90-day toxicity study was reported in rats exposed to the Origanum vulgare essential oil (OEO) orally (50, 100 and 200 mg/kg) (Llana-Ruiz-Cabello et al., 2017). OEO was found safe for oral consumption by rats and can be used in active food packaging. This study determined a no observable adverse effect level forOEO. However, the chemical composition of plants is related to soil, climate, cultivation, growing/harvested season and geographical region (Undeger et al., 2009). Additionally, carvacrol (100 mg/kg) prevented diet-induced obesity in mice fed with high-fat diet (Cho et al., 2012). Therefore, an accurate toxicological evaluation of these compounds is necessary. The cytotoxic, cell morphological effects, and the pro-oxidant/antioxidant role of carvacrol, thymol and their mixture on the intestinal Caco-2 cell line were reported (Llana-Ruiz-Cabello et al., 2014a, 2015b). The gastrointestinal tract, especially the stomach, would be the first target of these substances, if they were consumed by humans. It should be noted that these substances were almost completely absorbed in the pig stomach (Michiels et al., 2008). The present study aimed to investigate the toxic effects of carvacrol, thymol and their mixture on human gastric AGS cells. Furthermore, it is the first study investigating the mixture of carvacrol and thymol on human AGS cells.
In this study, the cytotoxic effect was investigated in human AGS cells exposed to carvacrol, thymol and their mixture after 24 h of incubation. A dose-dependent cytotoxic effect for carvacrol and the mixture carvacrol/thymol was found in agreement with the earlier reports, which were investigated in different cell lines (Llana-Ruiz-Cabello et al., 2015a). Recently, it was reported that 400 µM of thymol reduced AGS cell viability by 50% (Kang et al., 2016) using the MTT assay, but no significance analysis was done. In our study, thymol was investigated up to 600 µM, and this compound (30–600 µM) showed statistically significant cytotoxic effects on human AGS cells in a dose-dependent manner. In addition, 400 and 600 µM of thymol reduced AGS cell viability significantly by 2% and 0.6%, using the sensitive ATP cell viability assay. This difference may depend on the sensitivity of the method used. It was also reported that cell viability decreased in a dose-dependent manner when the human glioblastoma and human lung cancer cell lines were exposed to thymol (Hsu et al., 2011; Ozkan and Erdogan, 2011). Nevertheless, no cytotoxic effect was recorded for thymol (0–1 mM) in mouse cortical neurons (García et al., 2006). Thymol (0–250 µM) did not affect the cell viability in human intestinal cells, but morphological changes of cells were observed under an electron microscope (Llana-Ruiz-Cabello et al., 2014a).
Generally, phenolic substances may have either antioxidant or pro-oxidant activity depending on cell resistance, concentration and time (Ozkan and Erdogan, 2012). Therefore, we investigated the ROS generating effect of carvacrol, thymol and their mixture on AGS cells by the DCFH-DA assay, which is a reliable and efficient assay for quantifying cellular oxidative stress (Wang and Joseph, 1999). In this study, the dose-dependent ROS generating effect was found in AGS cells exposed to carvacrol, thymol and their mixture after 24 h. The significant increases were determined at higher concentrations of these substances with respect to the control cells. Similarly, Ozkan and Erdogan (2012) reported that carvacrol and thymol at higher concentrations exhibited a pro-oxidant effect on human lung cancer cells. In our previous study, carvacrol had a GSH reducing effect on human AGS cells, confirming the pro-oxidant activity of carvacrol (Gunes-Bayır et al., 2017). On the other hand, carvacrol (100 µM) caused intracellular ROS generation, while thymol (1–100 µM) did not, in V79 Chinese Hamster lung fibroblast cells after 24 h of exposure (Aydin et al., 2014). Lately, it was described that thymol (0–400 µM) increased the ROS level in AGS cells in a dose-dependent manner (Kang et al., 2016). In another study, carvacrol and its mixture with thymol showed increased ROS generation in Caco-2 cells, but thymol (250 µM) did not. These differences may occur because of different cell types used in the researche and the reported high absorption rate of thymol in the stomach and proximal small intestine of pigs (Michiels et al., 2008).
Apoptosis at the cellular level was investigated using AO/EB staining, which allowed the observation of nuclear changes and apoptotic body formation in cells. Cells were counted to quantify apoptotic, viable and necrotic cells under a fluorescence microscope (Kasibhatla et al., 2006). After 24 h of exposure to carvacrol (0–100 µM), thymol (0–100 µM) and their mixture (0:0–100:10 µM), the viability of human AGS cells decreased significantly, while the number of apoptotic and necrotic cells increased. Similarly, it pro-apoptotic activity of carvacrol was also reported in HepG2, breast and Caco-2 cells (Arunasree, 2010; Fan et al., 2015; Llana-Ruiz-Cabello et al., 2015a; Yin et al., 2012) and of thymol in promyelotic cancer (25–100 µM), human glioblastoma (200–800 µM) and AGS cells (100–400 µM) (Deb et al., 2011; Hsu et al., 2011; Kang et al., 2016). In contrast, human Caco-2 cells exposed to thymol (0–250 µM) did not show any apoptotic effects (Llana-Ruiz-Cabello et al., 2014a), but in agreement with our study, the mixture of carvacrol/thymol had apoptotic effect on Caco-2 cells at 150:15 µM more than at 300:30 µM. In our study, apoptosis was observed in human AGS cells exposed to thymol in a dose-dependent manner, in agreement with other studies, which were investigated in human promyelotic leukemia (HL-60), human glioblastoma, HepG2, non-small cell lung cancer (H1299) and human AGS cells (Deb et al., 2011; Hsu et al., 2011; Kang et al., 2016; Ozkan and Erdogan, 2011, 2012). However, no apoptotic effect for thymol was reported for human Caco-2, human HepG2 and mouse cortical neuron cells (García et al., 2006; Llana-Ruiz-Cabello et al., 2014a; Stammati et al., 1999). On the other hand, electron microscopic observations of Caco-2 cells exposed to thymol showed apoptotic appearances (Llana-Ruiz-Cabello et al., 2014a). Although several authors have been reported the apoptotic effect of carvacrol and thymol, in our study for the first time, the apoptotic effect of their mixture on human AGS cells was described. The apoptotic effect of the mixture on human AGS cells was found up to 20:2 µM in a dose-dependent manner, whereas this effect on human Caco-2 cells was reported up to 150:15 µM (Llana-Ruiz-Cabello et al., 2014a). Furthermore, the apoptosis-induced effect of carvacrol, thymol and their mixture was also confirmed by western blotting as in previous reports (Arunasree, 2010; Deb et al., 2011; Fan et al., 2015; Yin et al., 2012).
Phenolic compounds can affect the cellular redox status because of their pro-oxidant properties. This may result in cytotoxicity with non-genotoxic effects or with genotoxic effects depending on time and concentration (Bakkali et al., 2008). Besides, some polyphenols at lower concentrations can have an antioxidant effect, while their higher concentrations cause DNA damage due to their pro-oxidant effect. In our study, the genotoxic effect of carvacrol, thymol and their mixture was investigated using the comet assay, which is widely used as a method for detecting DNA damage (Hartmann et al., 2003). Carvacrol showed a genotoxic effect on human AGS cells in a dose-dependent manner. This finding is in accordance with the previous results obtained from mouse myoblast, human lymphocytes, human breast cancer and human Caco-2 cells exposed to carvacrol (Llana-Ruiz-Cabello et al., 2015a). Our study also demonstrated that thymol had a genotoxic effect on human AGS cells in a dose-dependent manner. Similar results were obtained from human HepG2 and non-small lung cancer cells following exposure to thymol (Ozkan and Erdogan, 2011, 2012). Carvacrol and thymol at concentrations < IC50 showed cytotoxic effects on HepG2 and Caco-2 cell lines that were not associated with the genotoxic effects (Horváthová et al., 2006). On the other hand, thymol (0–250 µM) had no genotoxic effect on Caco-2 cells (Llana-Ruiz-Cabello et al., 2014b). Similarly, no genotoxic effect was reported in V79 Chinese hamster lung fibroblast cells exposed to carvacrol and thymol at lower concentrations (1–25 µM) (Undeger et al., 2009). However, genotoxicity was found up to 25 µM of carvacrol and thymol in V79 cells. The conversion from antioxidants to pro-oxidants occurs at higher concentrations, suggesting a concentration-dependent effect of polyphenols. Considering many studies, the genotoxic effect of carvacrol and thymol was reported on different cell lines, but our study shows for the first time that carvacrol, thymol and their mixture have the genotoxic effect on human AGS cells.
Conclusions
To the best of our knowledge, this study is the first that presents cytotoxic, genotoxic, apoptotic and ROS generating and GSH reducing effects of carvacrol, thymol and their mixture on AGS cells. Both compounds and their mixture (10:1) showed a significant reduction in cell viability due to their cytotoxic effects. The apoptotic, genotoxic and ROS generating effects were also found in a dose-dependent manner. Because of their increased use at higher concentrations in the food industry, further studies are required to explain their toxicity in terms of normal/healthy cells in vitro and suitable in vivo models.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Bezmialem Vakif University, the Unit of Scientific Research Projects (grant number 12.2014/3).