
Abstract
It is well-documented that lead (Pb) toxicity can affect almost all systems in living organisms. It can induce selective autophagy of mitochondria (mitophagy) by triggering reactive oxygen species production. Emerging evidence has suggested that Pb-induced autophagy can also be activated by the endoplasmic reticulum (ER) stress pathway. However, the interplay between ER stress and mitophagy remains to be elucidated. In this study, human embryonic kidney HEK293 cells were employed to investigate the role of ER stress in Pb-induced mitophagy. The results showed that the cell viability was decreased and cell damage was induced after exposure to Pb (0, 0.5, 1, 2, and 4 mM) for 24 h in a dose-dependent manner. Moreover, the expression of LC3-Ⅱ was significantly increased, and the expression of HSP60 was dramatically decreased after exposure to 1 mM and 2 mM Pb, indicating the induction of mitophagy following Pb exposure. Meanwhile, the expressions of activating transcription factor 6, inositol-requiring protein-1α, CCAAT/enhancer binding protein homologous protein, and glucose-regulated protein 78 were dramatically increased after Pb treatment, signifying the initiation of ER stress. Notably, the mitophagic effect was significantly compromised when ER stress was inhibited by 0.5 mM 4-phenylbutyrate, which was evidenced by lesser decreases in HSP60 expression and level of LC3-Ⅱ, suggesting Pb-induced mitophagy may be activated by the ER stress. Taken together, these findings provide a better understanding of Pb toxicity and suggest that Pb-induced ER stress may play a regulatory role in the upstream of mitophagy.
Introduction
Lead (Pb), a toxic heavy metal and ubiquitous environmental pollutant, has been linked with many anomalies in living organisms (Gidlow, 2015). Chronic exposures of Pb affect almost all systems in human such as renal, nervous, and reproductive systems. Among these systems, the renal system appears to be the most affected by Pb toxicity (Chu et al., 2018; Matovic et al., 2015). For instance, previous studies have reported that Pb exposures are responsible for kidney damage and renal dysfunction in a number of animals such as rats (Liu et al., 2012) and chickens (Jin et al., 2017). At the cellular level, Pb2+ disrupts calcium homeostasis by competing with Ca2+, ultimately knocking Ca2+ out of mitochondria and causing opening of the mitochondrial permeability transition pore. This event causes mitochondrial damage, increases reactive oxygen species (ROS) production, and oxidative stress including altered lipid metabolism (Kapoor and Rossum, 1984). More specifically, opening of mitochondrial permeability transition pores results in the collapse of mitochondrial membrane potential and decreased ATP production in rat proximal tubular cells (Chu et al., 2018).
Previously, we have reported that Pb toxicity could induce mitochondrial damage due to the burst of ROS that triggers mitophagy, that is, selective removal of damaged mitochondria (Gu et al., 2018). It is a well-characterized natural process activated in cells to respond to various stresses and to protect cells from harmful effects of damaged mitochondria. Removal of damaged mitochondria through mitophagy requires the induction of general autophagy as demonstrated by increased LC3-II/LC3-I ratios. During mitophagy, the mitochondrial mass, which can be reflected by the expression of mitochondrial marker proteins such as HSP60, decreases as a consequence (Williams and Ding, 2018).
More recent studies have unveiled that some of the toxic effects of Pb are also localized to the endoplasmic reticulum (ER), which controls a wide range of cellular processes (Corsetti et al., 2017; Jia et al., 2018). A variety of stimuli such as calcium deprivation, oxidative stress, and exogenous toxins would initiate ER stress via a coordinated stress response called the unfolded protein response (UPR) (Gökçe et al., 2020). It has been found that UPR is restricted by three transmembrane ER stress sensors in mammalian cells, namely protein kinase R-like ER kinase (PERK), inositol-requiring protein-1α (IRE1α), and activating transcription factor 6 (ATF6) (Walter and Ron, 2011). The induction of ER-resident molecular chaperone glucose-regulated protein 78 (GRP78), also referred to as binding immunoglobulin protein (Bip), and the upregulation of the CCAAT/enhancer binding protein homologous protein (CHOP) are probably the most sensitive markers of ER stress (Zheng et al., 2014). Previous studies have shown that Pb exposure could induce ER stress in the nervous system and kidneys as well (Wang et al., 2018).
Interestingly, there are some reports concerning the interplay between ER stress and autophagy, which are connected in various ways, including ER stress-mediated autophagy activation and the formation of autophagosomes at the ER (Khaminets et al., 2015; Lee et al., 2015). However, there are only a few studies that focused on the interplay between mitophagy and ER stress, especially induced by Pb. Therefore, the present study aimed to explore how ER stress and mitophagy are interconnected in response to Pb. Our study demonstrated a regulatory role for ER stress in Pb-induced mitophagy in HEK293 cells. The findings of this study offer new insights into Pb cytotoxicity and the molecular mechanism of mitophagy induced by Pb.
Materials and methods
Chemicals and antibodies
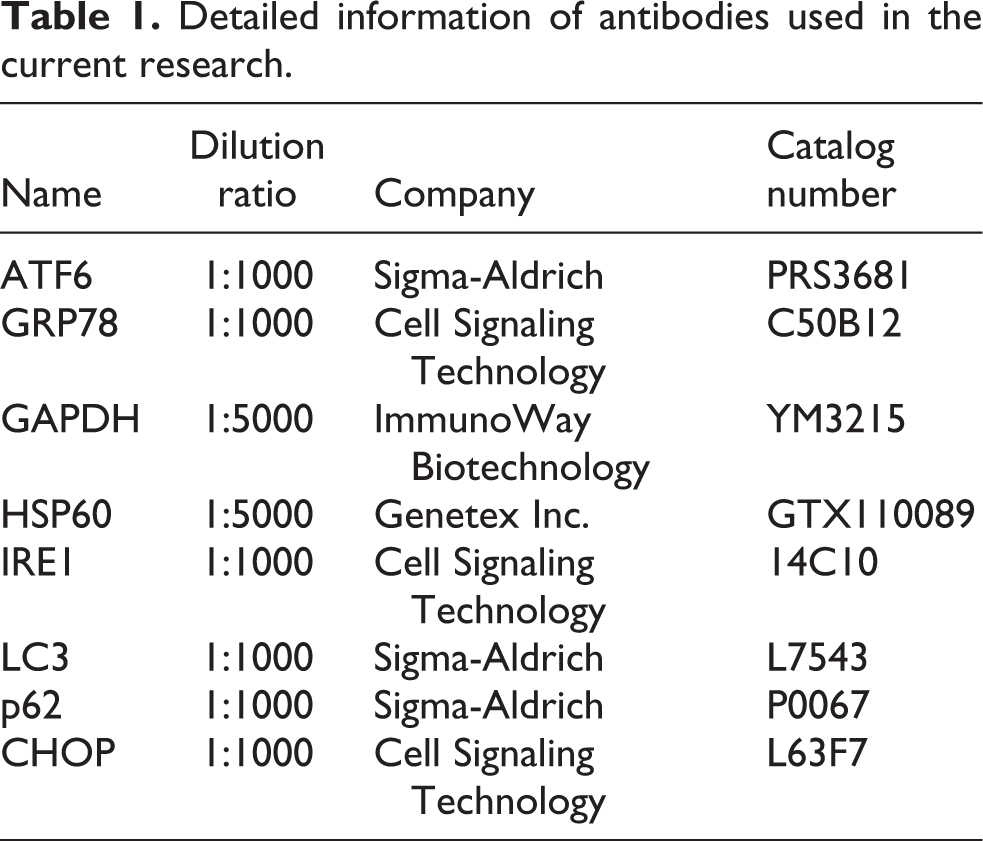
Lead acetate (Pb(Ac)2), carbonyl cyanide m-chlorophenylhydrazone (CCCP), tunicamycin (Tu), and 4-phenylbutyrate (4-PBA) were obtained from Sigma-Aldrich (St. Louis, Missouri, USA). The antibodies applied for Western blotting are listed in Table 1.
Detailed information of antibodies used in the current research.
Cell culture
HEK293 cells were purchased from the Cell Bank of Type Culture Collection of Chinese Academy of Sciences (Shanghai, China) and grown in Eagle’s Minimum Essential Medium (MEM) with nonessential amino acids (ThermoFisher Scientific, Waltham, MA, 41500034), supplemented with 10% fetal bovine serum (Royabio, Lanzhou, China, RY10021-G920), and 100 IU/mL of penicillin-streptomycin (Hyclone, Logan, Utah, SV30010). Cells were cultured in an incubator with 5% CO2 at 37°C.
Cell viability and lactate dehydrogenase assay
Cell viability was measured using a Cell Counting Kit-8 (CCK8). Briefly, cells were seeded in a 96-well plate (1×104 cells/well) and treated with Pb (0, 0.5, 1, 2, and 4 mM) for 24 h. Ten microliters of CCK8 solution (5 mg/mL) was added to each well and incubated at 37°C for 2–4 h and the absorbance was measured at 450 nm using a SpectraMax M2 spectrometer (Molecular Devices, Sunnyvale, California, USA).
Lactate dehydrogenase (LDH) release was measured using LDH assay kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer’s instructions.
Western blotting
HEK293 cells were treated with Pb (1 and 2 mM) for 24 h or CCCP (50 μM) for 6 h. After Pb treatment, the cells were lysed using lysis buffer for Western blotting (Beyotime biotechnology, Shanghai, China, P0013) supplemented with 1×PhosSTOP™ (Roche, Basel, Switzerland, PHOSS-RO) and 1×cOmplete™ ULTRA (Roche, Basel, Switzerland, 05892970001). The protein samples were separated by SDS-PAGE and transferred to a polyvinylidene fluoride (PVDF) membrane. After blocking with 5% nonfat milk for 1 h at room temperature, membranes were incubated with the primary and secondary antibodies sequentially. Finally, the protein levels were visualized using Immobilon Western chemiluminescent HRP substrate (Millipore, Billerica, MA, WBKLS0100) and quantified with ImageJ software (NIH, Bethesda, USA).
Statistical analysis
All statistical analyses were performed using SPSS 19.0 (IBM, NY, USA) and graphed with Origin 8.0 (OriginLab, USA). Statistical comparisons were made using one-way analysis of variance followed by the least significant difference post hoc test. Differences were considered significant at p < 0.05 and highly significant at p < 0.01.
Results
Pb inhibits cell growth and induces cell damage in HEK293 cells
To investigate the cytotoxic effect of Pb, HEK293 cells were treated with Pb (0, 0.5, 1, 2, and 4 mM) for 24 h. The results demonstrated that the cell viability was decreased in a dose-dependent manner. In the -2mM and 4-mM treatment groups, the cell viability was significantly lower than that of the control group (p < 0.01; Figure 1(a)). As expected, the LDH release, an indicator of cell injury, was increased gradually with the 2-mM and 4-mM treatment groups, showing a significant difference compared with the control group (p < 0.05; Figure 1(b)).
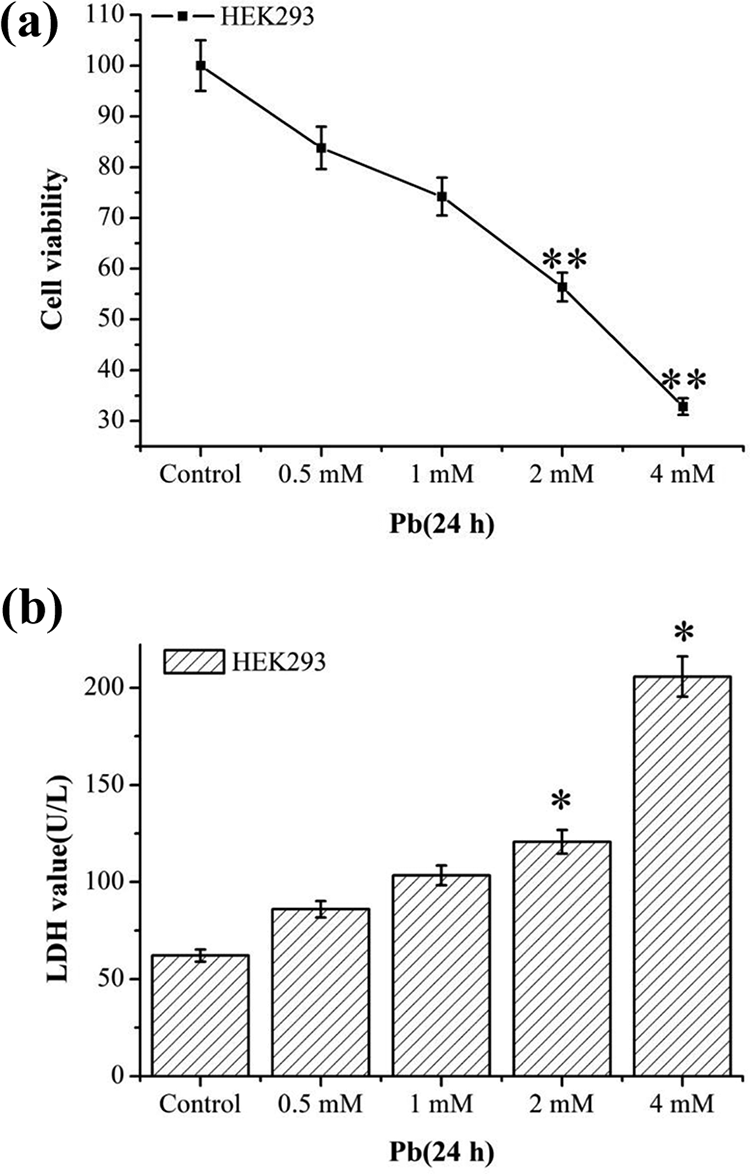
Effects of Pb on the cell viability and cell injury of HEK293 cells. HEK293 cells were treated with 0, 0.5, 1, 2 and 4 mM Pb for 24 h. Cell viability was measured by CCK8 kit (a). The LDH was measured by LDH assay kit (b). Statistical results are mean ± SEM of three independent experiments (n = 3). *p < 0.05 and **p < 0.01 indicate the significant difference compared with the control group. Pb: lead; CCK8: Cell Counting Kit-8; LDH: lactate dehydrogenase; SEM: standard error mean.
Pb induces mitophagy in HEK293 cells
To verify the effect of Pb on mitophagy, we first detected the expression of LC3 in HEK293 cells. The results showed that Pb exposure significantly increased the LC3-II/LC3-I ratio in a concentration-dependent manner, suggesting the formation of autophagosome (Figure 2(a)). Moreover, the expression of mitochondrial matrix protein HSP60 was decreased gradually in the 2-mM Pb and CCCP treatment groups (p < 0.01; Figure 2(b)), suggesting that Pb exposure decreases mitochondrial mass, demonstrated by decreased mitochondrial proteins, that is, HSP60.
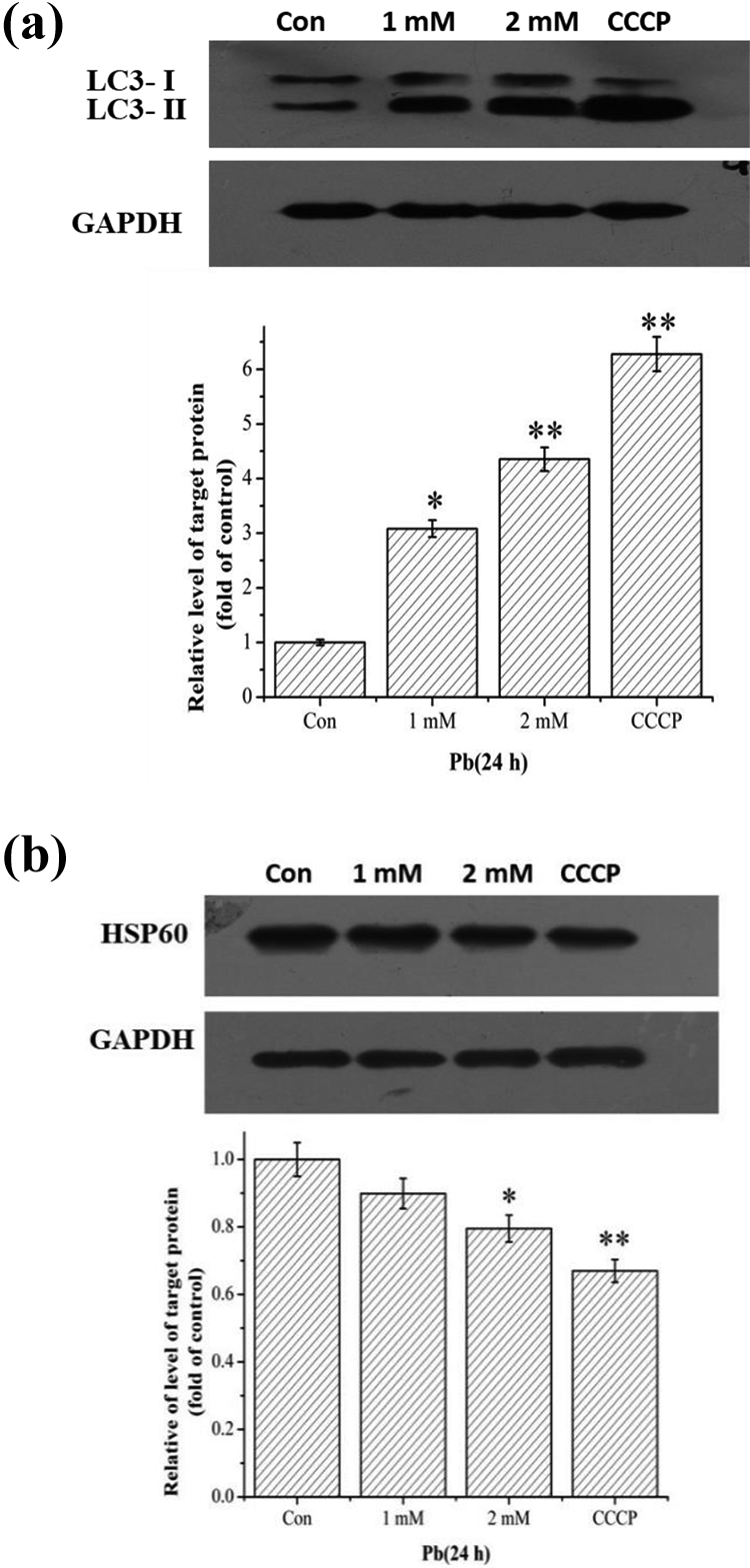
Effects of Pb on mitophagy. HEK293 cells were treated with 1 mM and 2 mM Pb for 24 h, or 50 μM CCCP for 6 h. The expression levels of LC3 (a) and HSP60 (b) were detected by Western blotting. Statistical results are mean ± SEM of three independent experiments (n = 3). *p < 0.05 and **p < 0.01 indicate the significant difference compared with the control group. Pb: lead; CCCP: carbonyl cyanide m-chlorophenylhydrazone; SEM: standard error mean.
Pb causes ER stress in HEK293 cells
To verify whether Pb could induce ER stress in HEK293 cells, we detected the expression level of ER stress–related proteins. Our results showed that the expression of ATF6 increased gradually in a concentration-dependent manner (Figure 3(a)). Similarly, the expression level of IRE1α was significantly increased in the 2 mM Pb or CCCP treatment group (Figure 3(b)).
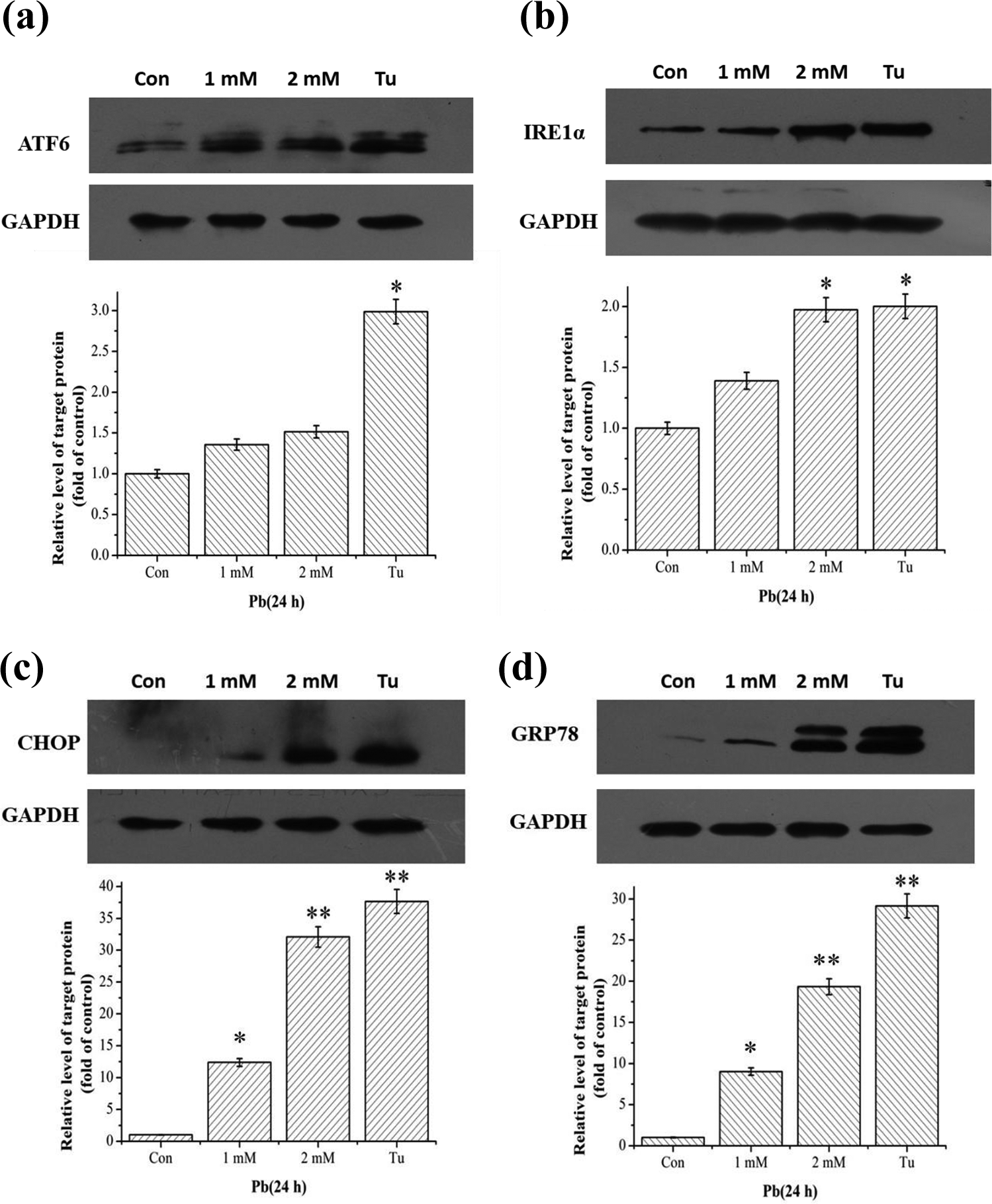
Effects of Pb on ER stress signaling pathway. HEK293 cells were treated with 1 mM and 2 mM Pb or 2 μg/mL Tu for 24 h. The expression levels of ATF6 (a), IRE1α (b), CHOP (c), and GRP78 (d
We further detected the expression level of CHOP, a downstream molecule of the ATF6 and PERK pathways, and the expression level of GRP78. The results showed that the expression levels of CHOP and GRP78 were increased in a concentration-dependent manner (p < 0.05) in the -2mM Pb or 2-μg/mL Tu (positive control) treatment groups (Figure 3(c) and (d)). In conclusion, these data demonstrated that Pb could induce ER stress by activating the ATF6 and IRE1 signaling pathways in HEK293 cells.
Blockage of ER stress attenuates Pb-induced mitophagy in HEK293 cells
4-PBA, an ER stress inhibitor, was used to elucidate the role of ER stress in the process of mitophagy induction. We first detected the effective dose of 4-PBA on ER stress response in HEK293 cells. The results suggested that 0.5 mM 4-PBA can effectively inhibit ER stress in HEK293 cells (Figure 4(a)). Therefore, we selected 0.5 mM 4-PBA to pretreat the cells in the following study.
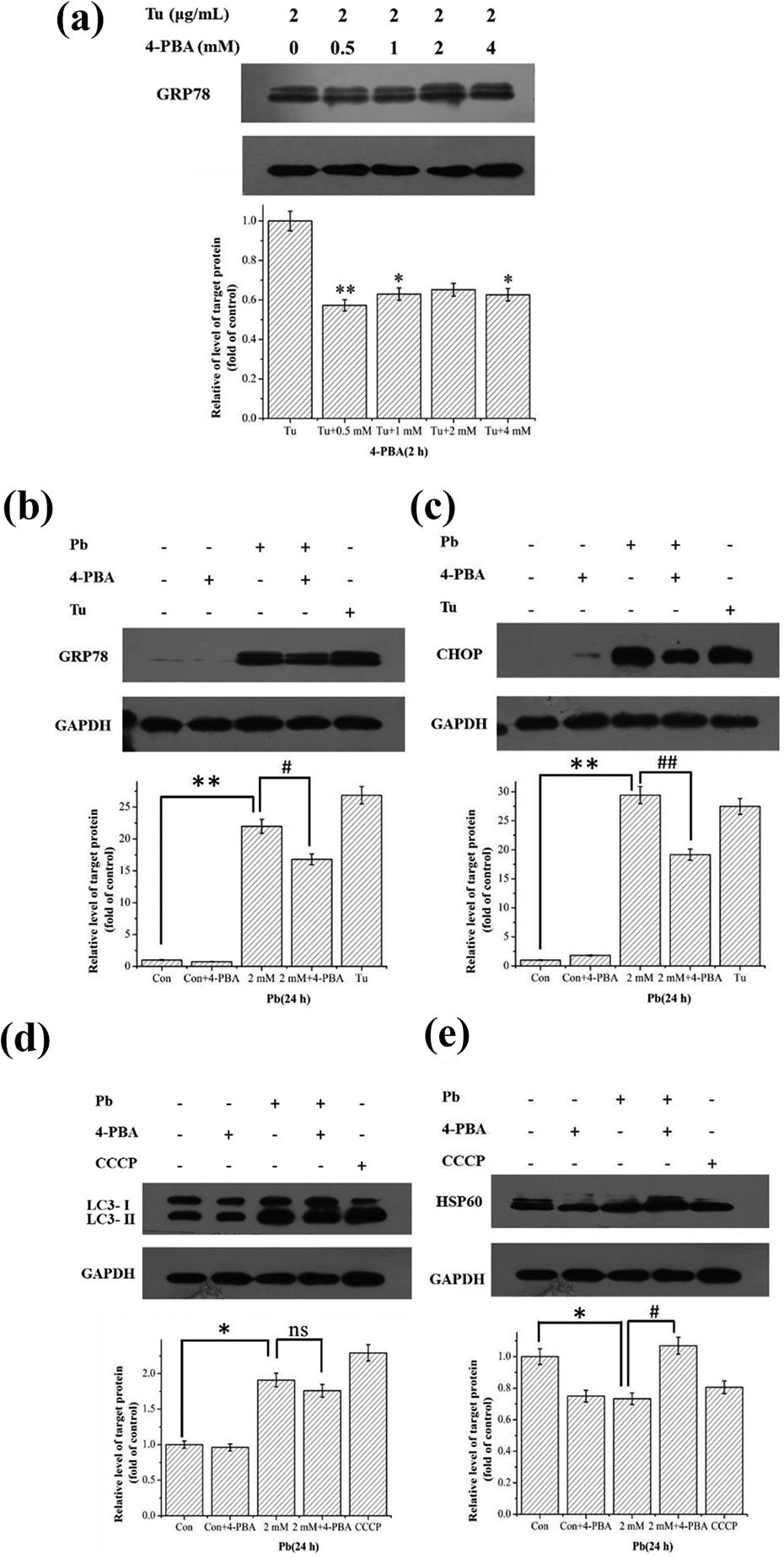
Inhibiting ER stress compromised mitophagy in HEK293 cells. HEK293 cells were pretreated with 0, 0.5, 1, 2, and 4 mM 4-PBA for 2 h, then treated with 2 μg/mL Tu for 24 h. The expression level of GRP78 was detected by Western blotting (a). HEK293 cells were pretreated with 0.5 mM 4-PBA for 2 h, and then treated with 2 mM Pb for 24 h. The expression levels of GRP78 (b), CHOP (c), LC3 (d), and HSP60 (e) were detected by Western blotting. Statistical results are mean ± SEM of three independent experiments (n = 3). *p < 0.05 and **p < 0.01 indicate the significant difference compared with the control group. #p < 0.05 indicates the significant difference between the 4-PBA + Pb and Pb alone. ER: endoplasmic reticulum; 4-PBA: 4-phenylbutyrate; Tu: tunicamycin; GRP78: glucose-regulated protein 78; CHOP: CCAAT/enhancer binding protein homologous protein; SEM: standard error mean; Pb: lead.
To determine whether 4-PBA could inhibit Pb-induced ER stress, HEK293 cells were pretreated with 0.5 mM 4-PBA for 2 h, followed by treatment with 2 mM Pb for 24 h. Our results showed that 4-PBA could effectively inhibit the increased expression of GRP78 and CHOP induced by Pb treatment (Figure 4(b) and (c)).
Furthermore, the mitophagic effect was evaluated under the condition of inhibited ER stress by 4-PBA. The cells were pretreated with 0.5 mm 4-PBA for 2 h, then treated with 2 mM Pb or 2 μg/mL Tu for 24 h. The results showed that inhibition of ER stress slightly decreased the conversion of LC3-I to LC3-II (Figure 4(d)) and significantly inhibited the degradation of mitochondrial maker proteins HSP60 (Figure 4(e)), suggesting that ER stress may play an important role in the activation of mitophagy.
Discussion
Pb toxicity can cause multiple toxic insults to various tissues and organs, whereas kidneys are notably the main target organ in this regard. Many studies have found that Pb exposures at low concentrations can increase the risk of chronic renal diseases, which may even lead to renal dysfunction (Chu et al., 2018; Liu et al., 2018). In the present study, we showed that Pb(Ac)2 exposure can decrease the viability of human embryonic kidney cells (HEK293) in a dose-dependent manner. Moreover, a previous study has found that Pb(Ac)2 exposure promoted the release of LDH, an important marker of cell damage, in primary rat proximal tubular cells (Song et al., 2017). Similarly, we found that the release of LDH increased gradually in a concentration-dependent manner after exposing HEK293 cells to different concentrations of Pb(Ac)2 for 24 h. Moreover, we found that ER stress–related proteins such as ATF6 and IRE1α were upregulated in the Pb treated group, indicating that ER stress might involve in Pb-induced toxicity. The results were further verified by blocking ER stress using ER stress inhibitors, that is, 4-PBA. ER stress inhibition resulted in decreased clearance of damaged mitochondria employing the role of ER stress in mitochondrial clearance.
Environmental pollutants can lead to mitochondrial damages, and mitophagy can be initiated to protect cells when the damage is reached to a certain extent (Gu et al., 2018). LC3-II and HSP60 are usually considered as important markers for the detection of mitophagy (Kliosnky, 2016). The present study showed that Pb(Ac)2 significantly elevated the protein levels of LC3-II and decreased that of HSP60, indicating that Pb(Ac)2 could induce mitophagy in HEK293 cells. It is noteworthy that more precise and quantitative assays of mitophagy have been well evaluated in our previous work (Gu et al., 2018), which further supported that Pb indeed induced mitophagy in HEK293 cells.
Various toxicants have been found to be able to induce ER stress (Hosohata et al., 2019). Arsenate has been shown to induce ER stress by elevating the expression levels of GRP78 and CHOP (Chen et al., 2017). Consistent with these studies, we found that the expressions of ATF6, IRE1, CHOP, and GRP78 were gradually increased in a concentration-dependent manner after Pb(Ac)2 exposure, indicating that Pb(Ac)2 can induce ER stress by the activation of the ATF6 and IRE1 signaling pathways. Nevertheless, other ER stress effectors such as the PKR-like eukaryotic initiation factor 2α kinase (PERK), eIF2α phosphorylation, or activated transcription factor 4 (ATF4) should be evaluated in the future study.
Previous studies have shown that ER stress could induce autophagy (Fernandez et al., 2015; Yang et al., 2016). For example, ER stress was involved in autophagy activation in mouse models of acute lung injury and human lung cancer cells (Xie et al., 2015; Zeng et al., 2017). Under ER stress, autophagy is generally considered as a cytoprotective response to the overload of unfolded or misfolded proteins (Lv et al., 2015). To explore the role of ER stress in Pb-induced mitophagy of HEK 293 cells, we examined the mitophagy induction under the conditions of inhibited ER stress. Our findings verified that inhibition of ER stress in Pb(Ac)2-treated cells decreased the conversion of LC3-I to LC3-II, increased the expression level of HSP60, suggesting that Pb(Ac)2-induced mitophagy is compromised by blocking ER stress activation.
In summary, our results showed that Pb could not only induce mitophagy but also induce ER stress. Pb-induced ER stress may play a regulatory role in the upstream of mitophagy. These findings enhance the understanding of Pb cytotoxicity and offer insights into the molecular mechanism of mitophagy induced by Pb.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by National Natural Science Foundation of China (31300941), Natural Science Foundation of Gansu Province (17JR5RA167), and Fundamental Research Funds for the Central Universities (lzujbky-2018-kb18).