
Abstract
This study investigated hepatic oxidative damage in rats following long-term manganese (Mn) exposure and clarified the underlying mechanisms. Forty-eight rats (SPF, male) were randomly assigned to receive low (10 mg/kg, n = 16) or high doses of Mn (50 mg/kg, n = 16) or sterilized distilled water (control group, n = 16). Rats were euthanized after 12 months, and liver Mn levels and histopathological changes were determined. Serum aspartate aminotransferase (AST) and alanine aminotransferase (ALT) levels and liver malondialdehyde (MDA), glutathione peroxidase (GSH-PX), nuclear factor E2-related factor-2 (Nrf2), heme oxygenase-1 (HO-1), and NAD(P)H quinine oxidoreductase-1 (NQO1) levels were also determined. The Mn concentration and relative liver weights were significantly higher in the high-dose Mn group than in the control and low-dose Mn exposure groups. Low-dose Mn exposure resulted in mild expansion of hepatic sinuses and intact nuclei, whereas high-dose exposure led to pathological alterations in hepatocytes. High-dose Mn treatment significantly increased AST, ALT, and MDA activities and decreased GSH-PX activity. Additionally, liver Nrf2, HO-1, and NQO1 protein expression were markedly reduced by Mn exposure. Under the study conditions, long-term low-dose Mn exposure resulted in slight pathological changes in liver structure, but high-dose Mn exposure affected both liver structure and function, which might be related to the inhibition of Nrf2 expression, suppression of the transcription of its underlying antioxidant genes, and down regulation of the corresponding proteins. Consequently, the antioxidant capacity in the rat liver was weakened.
Introduction
Previous studies on acute and subacute manganese (Mn) exposure indicated that high Mn levels damage the structure of liver tissue and affect its physiological function (Foster et al., 2015), and these changes were associated with oxidative stress. Researchers found that 2 days of Mn exposure significantly increased aspartate aminotransferase (AST) and alanine aminotransferase (ALT) levels in rat blood serum, as well as lipid peroxidation (LPO) and rat liver Mn concentrations (Montes et al., 2016). Furthermore, liver malondialdehyde (MDA) levels in grass carp and silver carp were increased following 30 days of Mn exposure. The total antioxidant capacity of the liver and catalase activity decreased after an initial increase during Mn exposure (Costa-Silva et al., 2018). Additionally, after 20 mg/mL manganese chloride (MnCl2) was orally administered to Wistar rats, the activities of superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GSH-PX), and nonenzymatic antioxidants were significantly reduced, indicating that Mn weakened the resistance to oxidative liver damage (Chtourou et al., 2013). After 60 days of Mn exposure in rats, liver LPO levels were significantly increased, whereas SOD and GSH-PX levels were decreased. Meanwhile, GSH-PX activity was enhanced in rats exposed to 45 mg/kg Mn (Chen et al., 2006). Another study reported that 90 days of MnCl2 exposure resulted in elevated rat liver MDA levels and markedly decreased GSH and GSH-PX contents, and the liver appeared to undergo pathological changes including cell necrosis and hepatic lobular arrangement disorder (Vieira et al., 2012).
A growing number of studies have shown that the nuclear factor E2-related factor-2 (Nrf2) signaling pathway plays a key role in oxidative damage, as Nrf2 and its underlying signaling pathway can protect cells via an antioxidant response mechanism (Keum and Choi, 2014). Various oxygen free radicals can activate Nrf2 signaling and the expression of subsequent antioxidant genes, promoting the synthesis of antioxidant enzymes such as GSH-PX, heme oxygenase 1 (HO-1) and NQO1 (Mondal et al., 2019; Toughan et al., 2018; Yang et al., 2019; Zhao et al., 2018). Antioxidative activity is enhanced via the increased expression of Nrf2 and upregulation of phase Ⅱ detoxification and antioxidant enzymes. Therefore, the Nrf2 signaling pathway may also play an essential role in the development of liver antioxidant damage. Previous research illustrated that Nrf2 and GPX-1 expression in rat testes was significantly decreased after 4 weeks of Mn exposure (Yang et al., 2019). Therefore, the Nrf2 signaling pathway could play an essential role in Mn-induced liver damage.
In daily life, long-term excessive Mn exposure is common among occupational workers, but current studies have focused on short-term high-level Mn exposure. Consequently, whether long-term exposure to excessive Mn levels has a negative impact on overall health has become a popular research topic. In addition, current studies including epidemiological surveys and animal experiments have mainly focused on neurotoxicity and motor dysfunction induced by long-term excessive Mn exposure (Beaudin et al., 2015; Bjørklund et al., 2017). Because the liver has higher adaptability to toxicological insults than the neural system, little research has explored the liver changes following long-term high-level Mn exposure. In particular, whether this exposure results in hepatic oxidative damage is unclear, and the mechanism by which Nrf2 signaling changes in the liver under long-term excessive Mn exposure remains to be studied. Therefore, our animal model simulated approximately one-third to one-half of the human life cycle under long-term excessive Mn exposure to mimic such occupational exposure in humans. The effects of different Mn doses, including hepatic oxidative damage and changes in antioxidant proteins such as HO-1, NQO1, and Nrf2, were examined.
Methods
The present study followed the guidelines for laboratory animal management and animal ethical protection during feeding, and the experimental protocol was approved by the Zunyi Medical University Institutional Animal Care and Use Committee (2014-2-001). The present study followed the Guidelines for the Management and Use of Laboratory Animals (Eighth Edition). 48 SPF male SD rats, weighing 260-300g, were provided by the Experimental Animal Center of Third Military Medical University (SCXK(YU)2012-0005). All rats were maintained at a temperature of 22°C ± 4°C and relative humidity of 50–60% under a 12-h/12-h light/dark cycle with unrestricted access to food and water. After 1 week of acclimation, the rats were randomly assigned to receive high-dose Mn (50 mg/kg, n = 16), low-dose Mn (10 mg/kg, n = 16), or sterilized distilled water (control group, n = 16). Mn or water was orally administered once daily five times per week (M-F) for 12 months.
Hematoxylin and eosin staining
Liver tissue rinsed with phosphate buffer and then stepwise dehydrated with 85%, 90%, 95% alcohol, anhydrous ethanol, alcoholic benzene, xylene, and wax. The dehydrated tissues were embedded by paraffin, sectioned with a thickness of 5 μm, dewaxed, and washed with alcohol (95%, 90%, 85%) and distilled water. The sections were stained with hematoxylin and eosin (HE) to observe their morphology.
AST and ALT determination
This protocol was then performed in strict accordance with the description instructions of commercially available AST and ALT activity test kits (Beijing Luzesen BC1565-100 and Beijing Luzesen BC1555-100, respectively).
MDA content and GSH-PX activity
Approximately 0.1 g of liver tissue was added to 1 mL of the extract for homogenization in an ice bath and centrifuged at 8000 × g for 4 min at 4°C (L500 desktop centrifuge, Hunan Xiangyi Laboratory Instrument Development Co., Ltd., Xiangyi, China). The supernatant was removed and placed on ice for testing. MDA content and GSH-PX activity were determined in strict accordance with the instructions of commercially available MDA (Beijing Luzesen BC0025-100, Beijing Soleibao Technology Co., Ltd.) and GSH-PX content test kits (Beijing Luzesen BC1175-100).
Graphite furnace atomic absorption
We added 2 mL of concentrated nitric acid to accurately weigh liver tissue samples that were then placed under a fume hood at room temperature until digestion, as indicated by a clear bright yellow color. Digested samples were transferred to clean dry beakers and heated on a hot plate at 100°C to near dryness. The samples were cooled to room temperature and then diluted with 1% dilute nitric acid to a volume of 5 mL. The sample was subjected to graphite furnace atomic absorption spectrometry, soaked in 30% nitric acid solution overnight, repeatedly rinsed with pure water, and baked for 10 min.
Western blot
Lysis buffer was added to liver tissues, which were shaken on ice for 2 h. The total protein was separated via centrifugation at 13685x g for 2 h at 4°C. The protein concentration was calculated using a bicinchoninic acid protein assay kit (Pierce, Rockford, Illinois, USA). Protein samples were diluted with sodium dodecyl sulfate polyacrylamide gel electrophoresis sample loading buffer (Beyotime Institute of Biotechnology, China), and the electrophoresis gel was heated at 100°C for 5 min before loading the samples. The antibodies targeted Nrf2, HO-1, NQO1, and β-actin. Concentrated gel was prepared with 5% antibodies, and a separation gel was prepared with 12% β-actin, HO-1, NQO1, and 10% Nrf2 antibodies. The PVDF membrane with sample protein was washed in Tris buffer solution Tween-20 (TBST) buffer (Solarbio Life Sciences, China) and incubated with alkaline phosphatase-conjugated secondary antibody (1:5000; Zhongshan Golden Bridge Biotechnology, China) for 1 h at room temperature (37°C). A suitable amount of luminescent liquid was pipetted to cover the PVDF membrane (7 Seas Biotech, China), and the image was exposed and collected on an ECL illuminator.
Data analysis
Statistical analysis was performed using SPSS 18.0. The visceral coefficient was calculated as follows: visceral coefficient = organ weight (g)/body weight (100 g) × 100%. One-way analysis of variance was used for between-group comparisons. If there were significant differences, the least significant difference method was used with pairwise comparisons. Differences were considered statistically significant at p < 0.05.
Results
Survival of Sprague Dawley rats exposed to Mn
Figure 1 presents the survival of Sprague Dawley rats exposed to Mn. By the end of the experiment, the mortality rate was 18.75% in the high-dose Mn group versus 6.25% in both the low-dose Mn and control groups.
Euthanasia of SD rats at different time points. SD: Sprague Dawley.
Mn content in rat livers after 12 months of Mn exposure
Figure 2 reveals that the Mn content in the liver was greater in the high-dose Mn group than in the low-dose Mn and control groups (p < 0.05). There was no difference in Mn content between the low-dose Mn and control groups (p > 0.05).
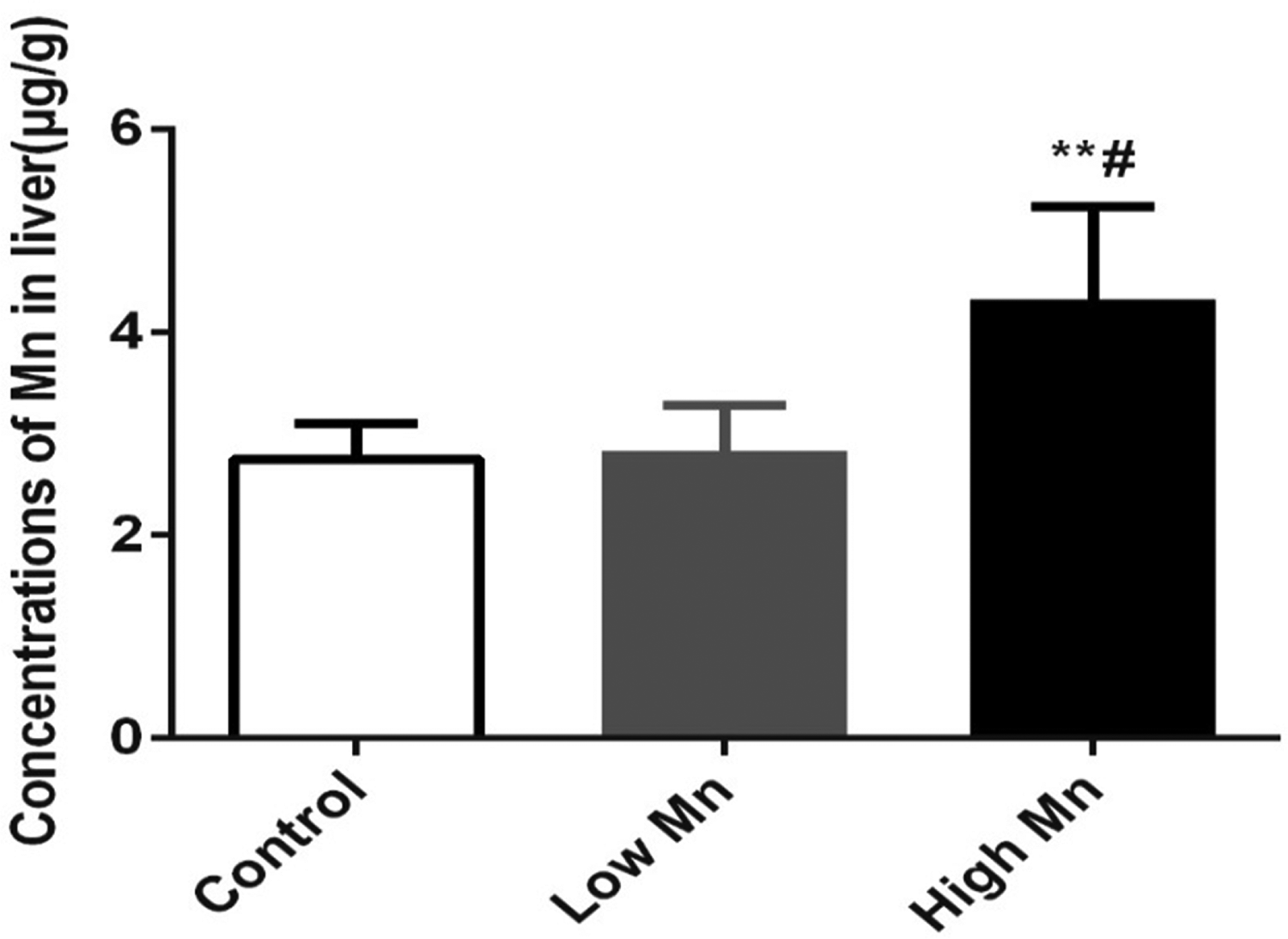
Mn accumulation in rat liver after 12 months Mn exposure (n = 6, mean ± SD), chronic Mn exposure increased accumulation in low Mn and high Mn exposure groups. Values were presented as mean ± SD. µg/g is wet weight. **p < 0.01 vs. control group; # p < 0.05 vs. low Mn exposure group. SD: standard deviation.
Liver organ coefficient increased after 12 months of Mn exposure
Figure 3 reveals that the liver organ coefficient was significantly higher in the high-dose Mn group than in the control and low-dose Mn groups (p < 0.05). The coefficient did not significantly differ between the low-dose Mn and control groups (p > 0.05).
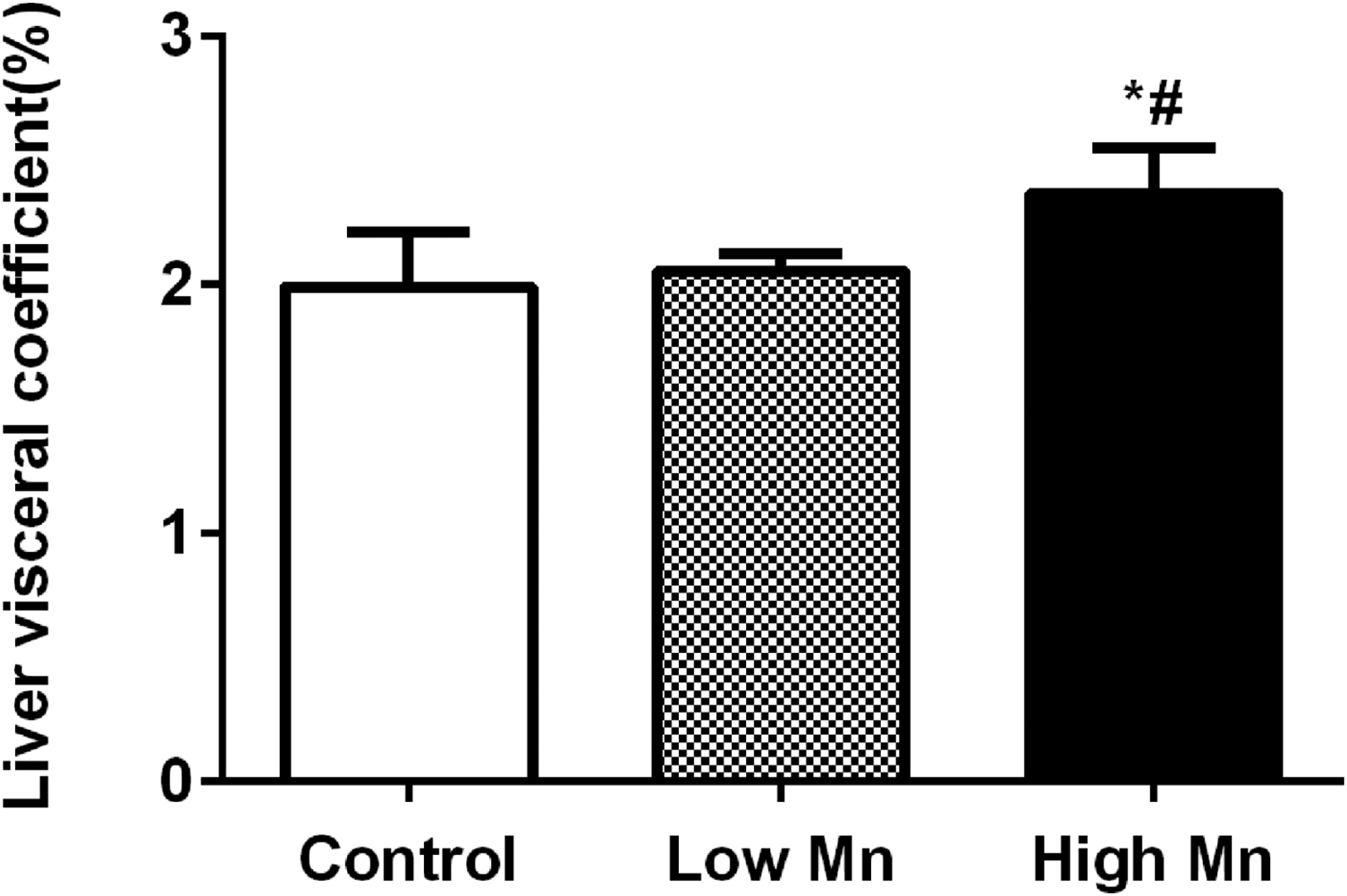
Changes of rat liver organ coefficient after 12 months Mn exposure, liver organ coefficient increased both in low Mn and high Mn exposure groups (n = 6 per group), values were presented as mean ± SD. *p < 0.05 vs. control group; # p < 0.05 vs. low Mn exposure group. SD: standard deviation.
Pathological changes were observed via HE staining of rat liver tissue after 12 months of Mn exposure
Hematoxylin and eosin (HE) staining showed that liver tissue in the control group was normal, featuring a regular liver lobule, normal hepatocytes, an intact hepatic cord, intact cell membranes, and clear nuclei (Figure 4(a)). In the low-dose Mn group, the structure of the hepatic lobule and hepatic cord was relatively complete, and there were no obvious abnormalities in the slices of liver tissue (Figure 4(b)). However, in the high-dose Mn group, the hepatic lobular structure was destroyed, including a fractured hepatic cord, degenerated hepatocytes, necrosis, ruptured cell membranes, smaller nuclei, and damaged central venous intimae. The cell gap widened, the number of inflammatory cells was increased, and hepatic sinusoids were expanded and congested (Figure 4(c)).
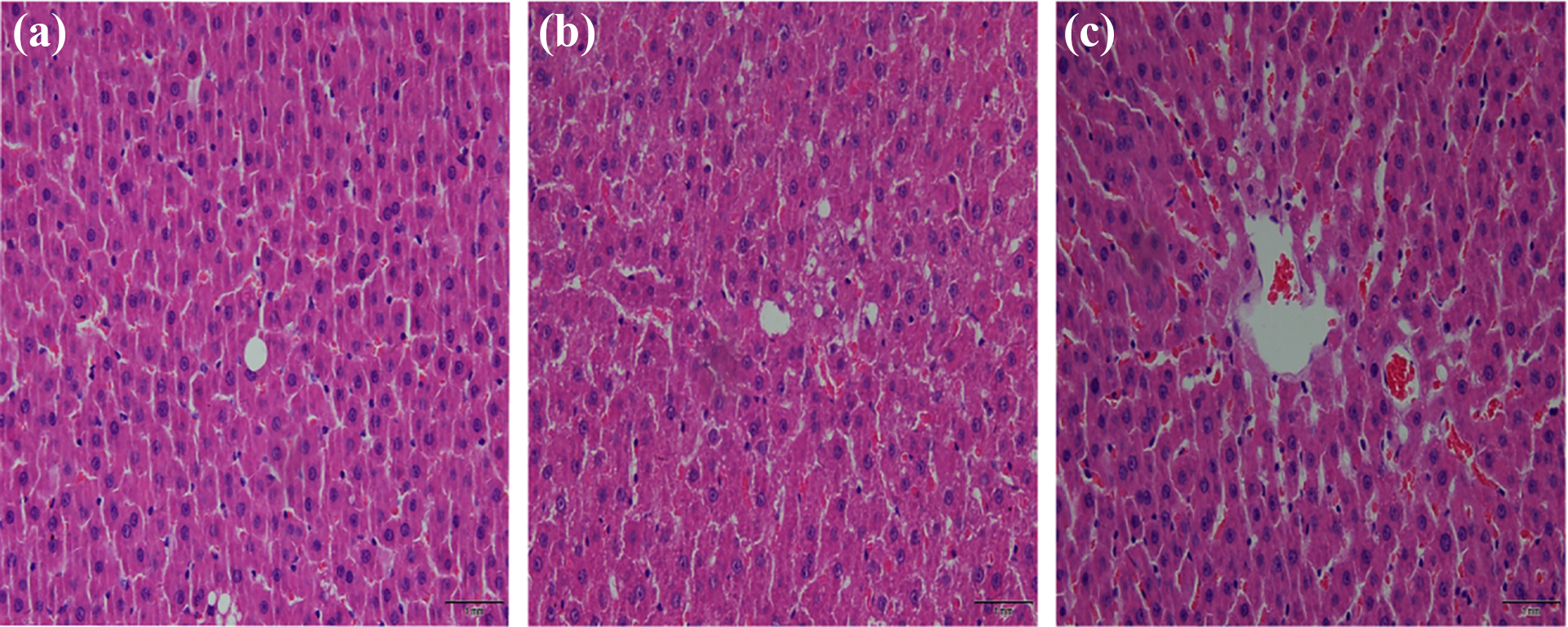
HE staining photographs show histopathological changes in striatum in different groups after 12 months of Mn exposure. (a) Control group. (b) Low-Mn exposure group. (c) High-Mn exposure group. HE staining, ×40. HE: hematoxylin and eosin; Mn: manganese.
AST and ALT activities in rat serum were increased by Mn exposure
Table 1 illustrates that the serum activities of AST and ALT were greater in the high-dose Mn exposure group than in the control and low-dose Mn groups (all p < 0.05). More critically, both AST and ALT activities were significantly greater in the low-dose Mn and control groups (both p < 0.05).
Mn-induced increases of GOT and GPT activities in rat blood serum (n = 6).a
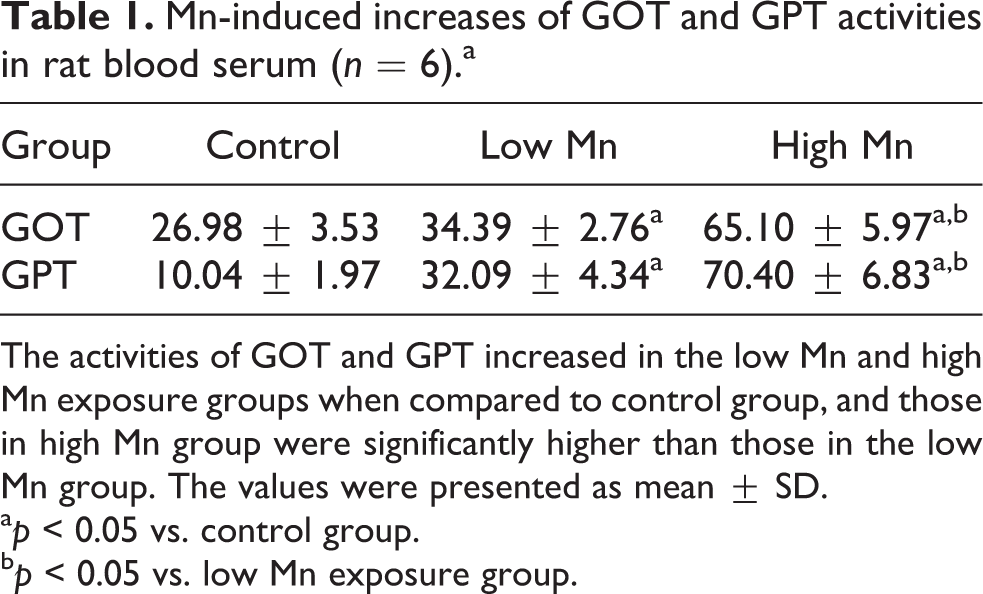
The activities of GOT and GPT increased in the low Mn and high Mn exposure groups when compared to control group, and those in high Mn group were significantly higher than those in the low Mn group. The values were presented as mean ± SD.
a p < 0.05 vs. control group.
b p < 0.05 vs. low Mn exposure group.
MDA and GSH-PX activities in the rat liver changed differently in the high- and low-dose Mn groups
Table 2 shows that MDA activity was significantly greater in the high-dose Mn group than in the control and low-dose Mn groups (both p < 0.05). MDA activity did not differ between the low-dose Mn and control groups (p > 0.05). Furthermore, GSH-PX activity was significantly lower in the high-dose Mn group than in the control and low-dose Mn groups (both p < 0.05), and GSH-PX activity was lower in the low-dose Mn group than in the control group (p < 0.05).
Mn-induced changes in MDA and GSH-PX activities in rat liver (n = 6).a
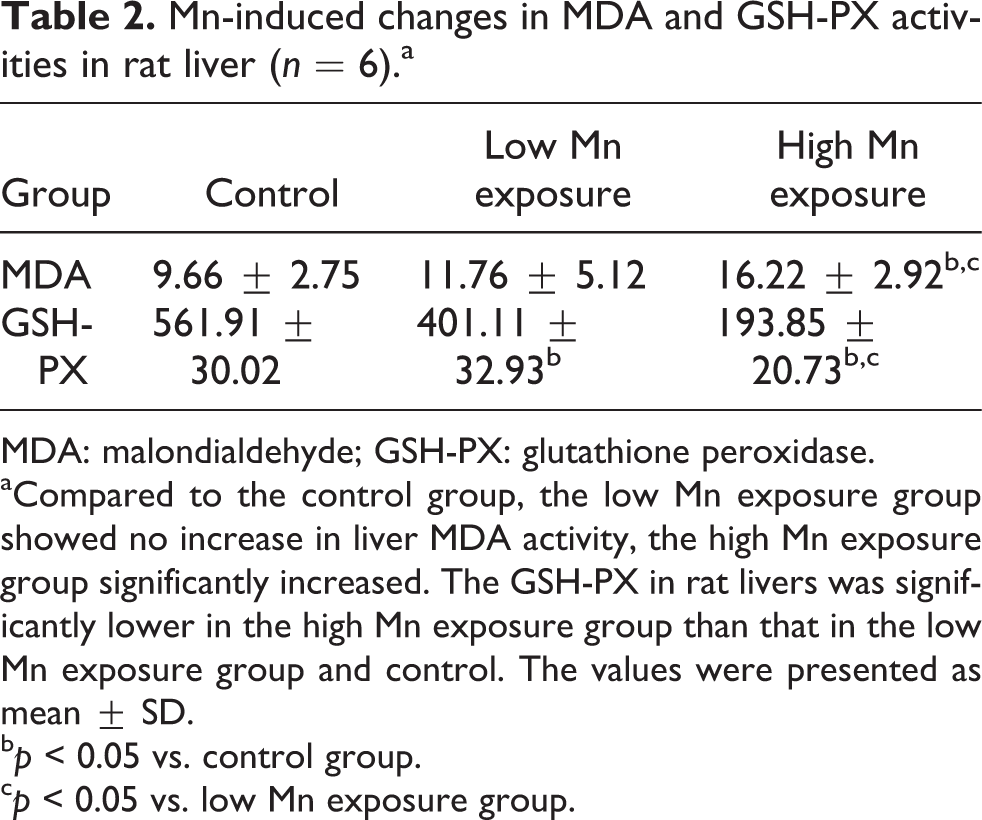
MDA: malondialdehyde; GSH-PX: glutathione peroxidase.
aCompared to the control group, the low Mn exposure group showed no increase in liver MDA activity, the high Mn exposure group significantly increased. The GSH-PX in rat livers was significantly lower in the high Mn exposure group than that in the low Mn exposure group and control. The values were presented as mean ± SD.
b p < 0.05 vs. control group.
c p < 0.05 vs. low Mn exposure group.
NQO1, HO-1, and Nrf2 expressions in the rat liver were decreased by Mn exposure
As presented in Figure 5, HO-1, NQO1, and Nrf2 protein expression in the liver was significantly less in the low- and high-dose Mn groups than in the control group (all p < 0.05). Furthermore, the levels of the aforementioned proteins were significantly less in the high-dose Mn group than in the low-dose Mn group (all p < 0.05).
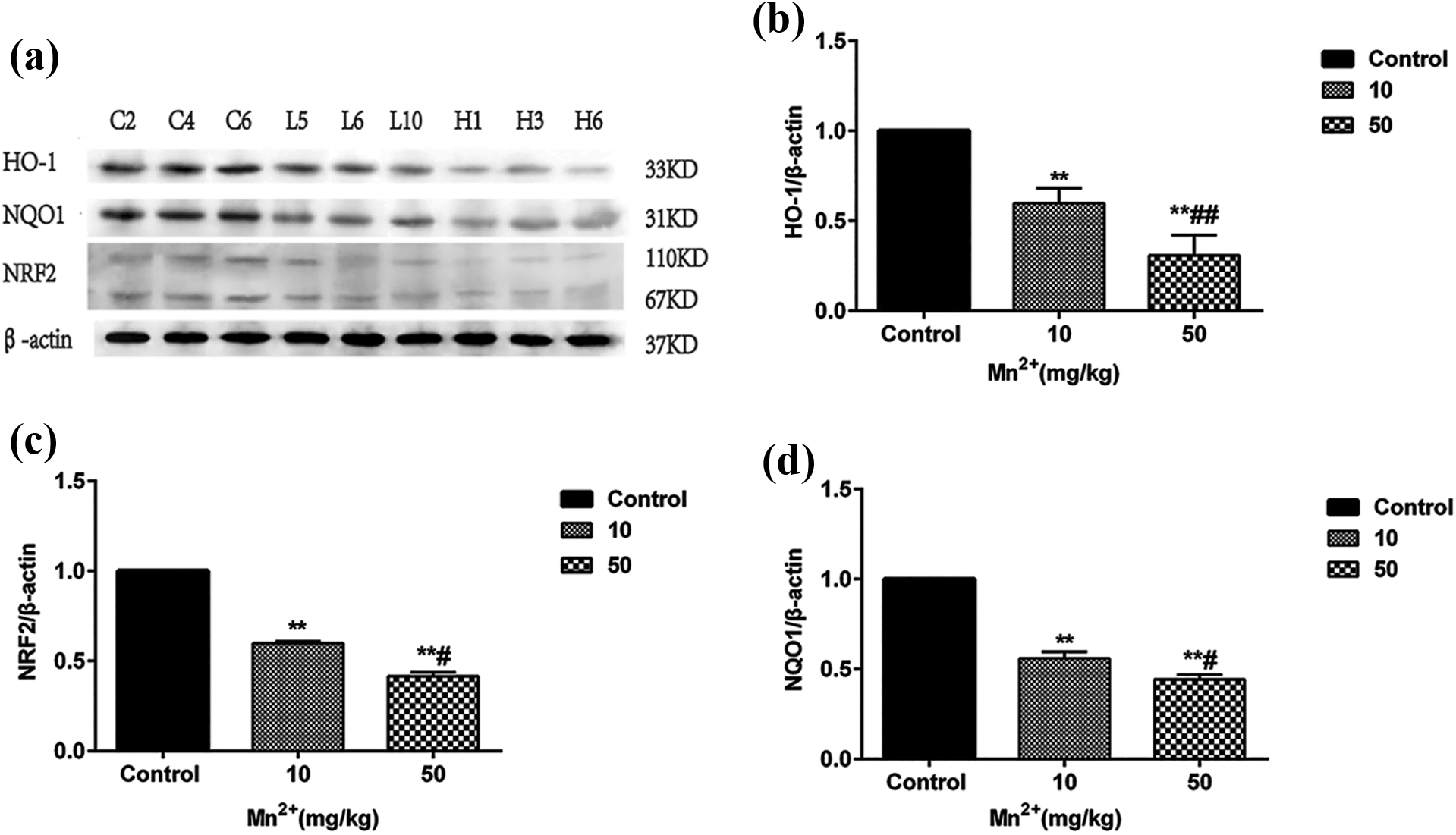
The protein expressions of HO-1, NQO1, and Nrf2 in the low Mn exposure group and the high Mn exposure group decreased after 12 months Mn exposure, (a) protein strip ((b) to (d)) quantitative analysis of HO-1, NQO1, and Nrf2 protein expression by immunoblotting respectively and comparison of protein expression between groups. Values were presented as mean ± SD. **p < 0.01 vs. control group; # p < 0.05 vs. low Mn exposure group; ## p < 0.01 vs. low Mn exposure group. HO-1: heme oxygenase-1; NQO1: NADPH quinine oxidoreductase-1; Nrf2: nuclear factor E2-related factor-2; Mn: manganese; SD: standard deviation.
Discussion
Our study found that Mn markedly accumulated in the livers of rats in the high-dose Mn group but not in the low-dose Mn group, consistent with the results of Serrazina et al. (2018). Thus, it can be speculated that long-term low-dose Mn exposure may accelerate Mn metabolism and maintain the relative stability of Mn content in the liver, resulting in only slight liver damage.
Our experimental results supported the aforementioned report and demonstrated a notable increase of the liver organ coefficient only in the high-dose group, consistent with the findings of Schmitz et al. (2019). In addition, some abnormal changes in the liver, such as broken hepatic lobule structures and hepatocyte cords, dilated and congested hepatic sinusoids, broken hepatocyte membranes, and reduced nuclei size were only observed in the high-dose Mn group, in line with previous findings (Schmitz et al., 2019; Vieira et al., 2012). Increases in the organ coefficient, the ratio of an organ’s weight to body weight, are indicative of hyperemia, edema, and hypertrophy of tissues and organs (Karimi et al., 2019). Our results revealed that chronic high-dose Mn exposure severely damaged liver tissues, whereas low-dose Mn exposure caused few pathological changes in the liver.
Similarly, AST and ALT levels were dramatically higher in the high-dose Mn group than in the other groups, whereas their levels were also increased by low-dose exposure relative to the control values. As classical amino acid metabolizing enzymes, AST and ALT liver levels increase when liver damage occurs (Li et al., 2019). Previous studies also found that AST and ALT levels in rat serum were increased significantly by 60 days of Mn exposure (Chen et al., 2006). Hence, we concluded that long-term excessive Mn exposure changes liver structure and damages liver hepatocytes.
We further tested several indicators of oxidative damage, including MDA and GSH-PX. LPO levels can be determined by detecting MDA activity (Samarghandian et al., 2017). GSH-PX, one of the main enzymes of the glutathione redox reaction, can specifically catalyze the reaction of reduced glutathione with reactive oxygen species (ROS) to produce glutathione, which protects cells against damage, helping to maintain normal cell function. More critically, GSH-PX is also related to liver protection, and the antagonism of harmful metal ions to the body. Shen and Jia found that MDA activity was increased by approximately 26% and 78% after 30 and 90 days of Mn exposure, respectively, resulting in rat liver damage (Shen and Jia, 2011). They also found that GSH-PX activity in liver tissue was significantly decreased, and a dose–response relationship with antioxidant damage was noted. Our experimental results, similar to previous acute and subacute experiments, illustrated that only high-dose Mn exposure significantly increased MDA activity. Additionally, both high- and low-dose Mn exposure significantly decreased GSH-PX activity, although the decrease was slightly less in the low-dose group. Given these findings, we believe that long-term high-level Mn exposure leads to liver decompensation, and Mn accumulation in liver tissue results in liver structural and functional changes through oxidative damage. However, long-term low-level Mn exposure may be mitigated by accelerated liver metabolism, resulting in less obvious damage.
Furthermore, we detected changes in the expression of proteins involved in Nrf2 signaling to explore the mechanism of Mn-induced oxidative injury. The transcription factor Nrf2, one of the core regulators of the antioxidative response to exogenous stimuli, contains six distinct functional regions, named Neh1 to Neh6 (Nrf2-ECH homology). The Neh2 region, which combines with the cytoplasmic protein Kelch-like ECH-associated protein 1 (Keap1), is relatively stable and anchored in the cytoplasm. Beyond that, Nrf2 uncouples from Keap1 and translocates to the nucleus following the phosphorylation of various protein kinases. The Neh1 region binds with Maf protein in the nucleus to form a heterodimer and then recognizes and combines with ARE protein, thus activating the expression of downstream antioxidant genes and phase Ⅱ detoxifying enzymes to produce antioxidants following stimulation by oxidants or electrophiles (Mondal et al., 2019; Woolbright and Jaeschke, 2017). Similarly, it has been demonstrated that MnCl2 concentrations of 12.5, 25, and 50 mg/kg can induce differences in the expression of Nrf2-related proteins, resulting in increased ROS levels in the glial cells of mice. Consequently, low Mn concentrations activated the Nrf2 pathway and increased HO-1 and NQO1 expression, whereas high Mn concentrations inhibited these changes (Qi et al., 2019). Although Mn is an essential trace nutrient for organisms because of its role in the function as an enzymatic cofactor and ROS scavenger, these effects are confined to low concentrations and appropriate exposure times (Zeinert et al., 2018). Our experiment demonstrated that HO-1, NQO1, and Nrf2 expression was significantly reduced by Mn exposure. Meanwhile, Nrf2 pathway activity and the expression of its underlying proteins were significantly less in the high-dose group than in the low-dose group, demonstrating that excessive Mn levels could inhibit Nrf2 signaling in the liver (Figure 6), suppress the transcription of underlying antioxidant genes, and decrease corresponding protein levels and antioxidant capacity of rat livers. The present study indicated this might be one of the mechanisms of Mn-induced liver oxidative injury.
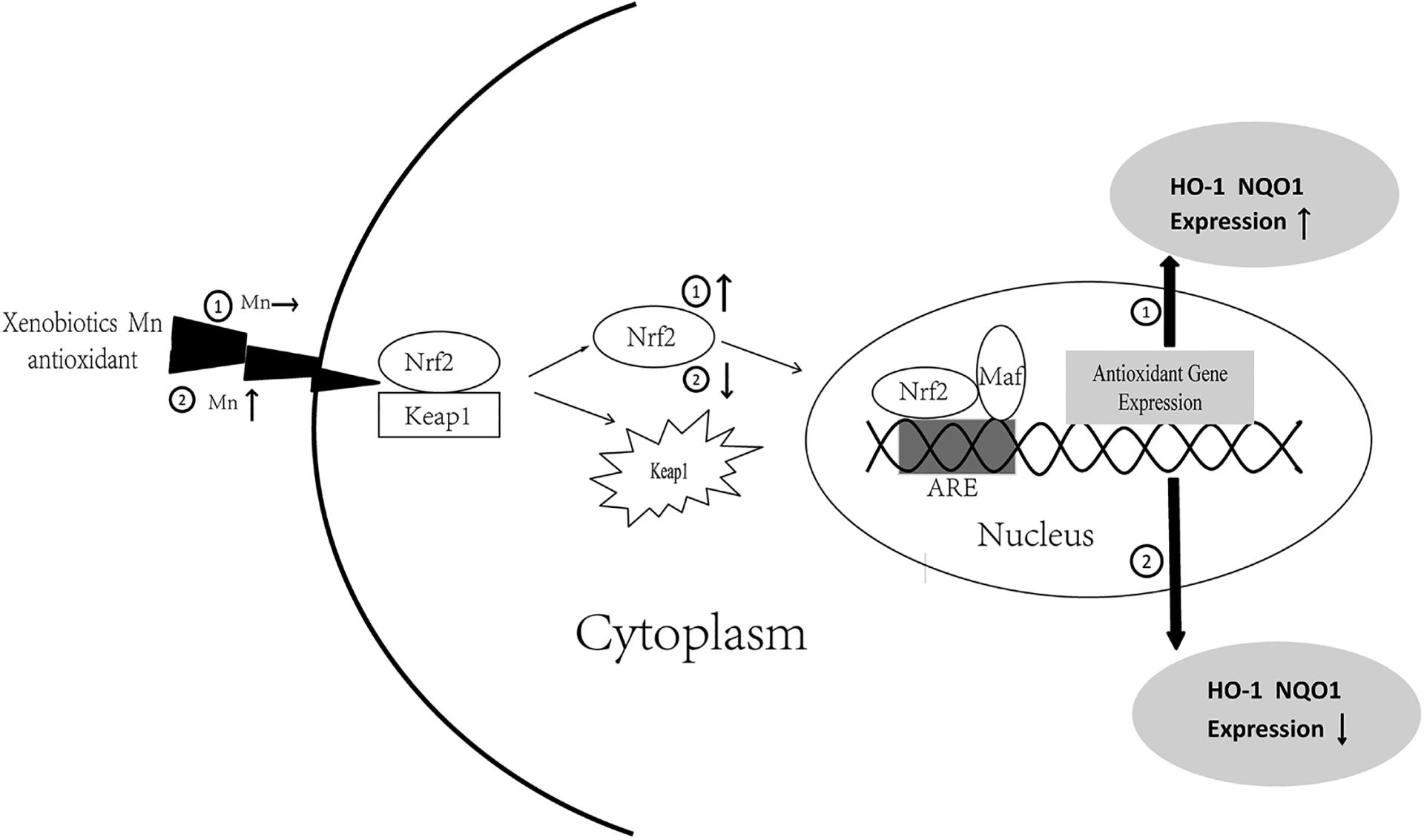
Nrf2 pathway and its underlying proteins change after 12 months Mn exposure. ① Trace Mn exposure can activate Nrf2 in cytoplasm and promote its combination with Maf protein and ARE to accelerate the expression of HO-1 and NQO1, subsequently producing antioxidant gene to play a protective role in the cell. ② Excessive Mn exposure inhibited Nrf2 in cytoplasm, subsequently decreased its chance to bind with Maf protein and ARE to translate antioxidant genes, resulting in the decline of the expression of HO-1 and NQO1.
Conclusion
Serum AST and ALT were changed by long-term low-dose Mn exposure, but liver damage was not clearly observed. This may be related to the acceleration of Mn metabolism in the liver. Furthermore, high-dose Mn exposure caused Mn accumulation in the liver and dramatically damaged liver structure and possibly function, which could be correlated with the inhibited expression of Nrf2 and its underlying proteins and resulting in decreased ability of rat livers to resist oxidative damage.
Footnotes
Acknowledgments
The authors sincerely thank the Committee of National Natural Science Foundation.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by The National Natural Science Foundation of China (81660552).